

Novel Saccharomyces cerevisiae × Saccharomyces mikatae Hybrids for Non-alcoholic Beer Production
 , ,
, ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Media and Solutions
2.2. Microorganisms
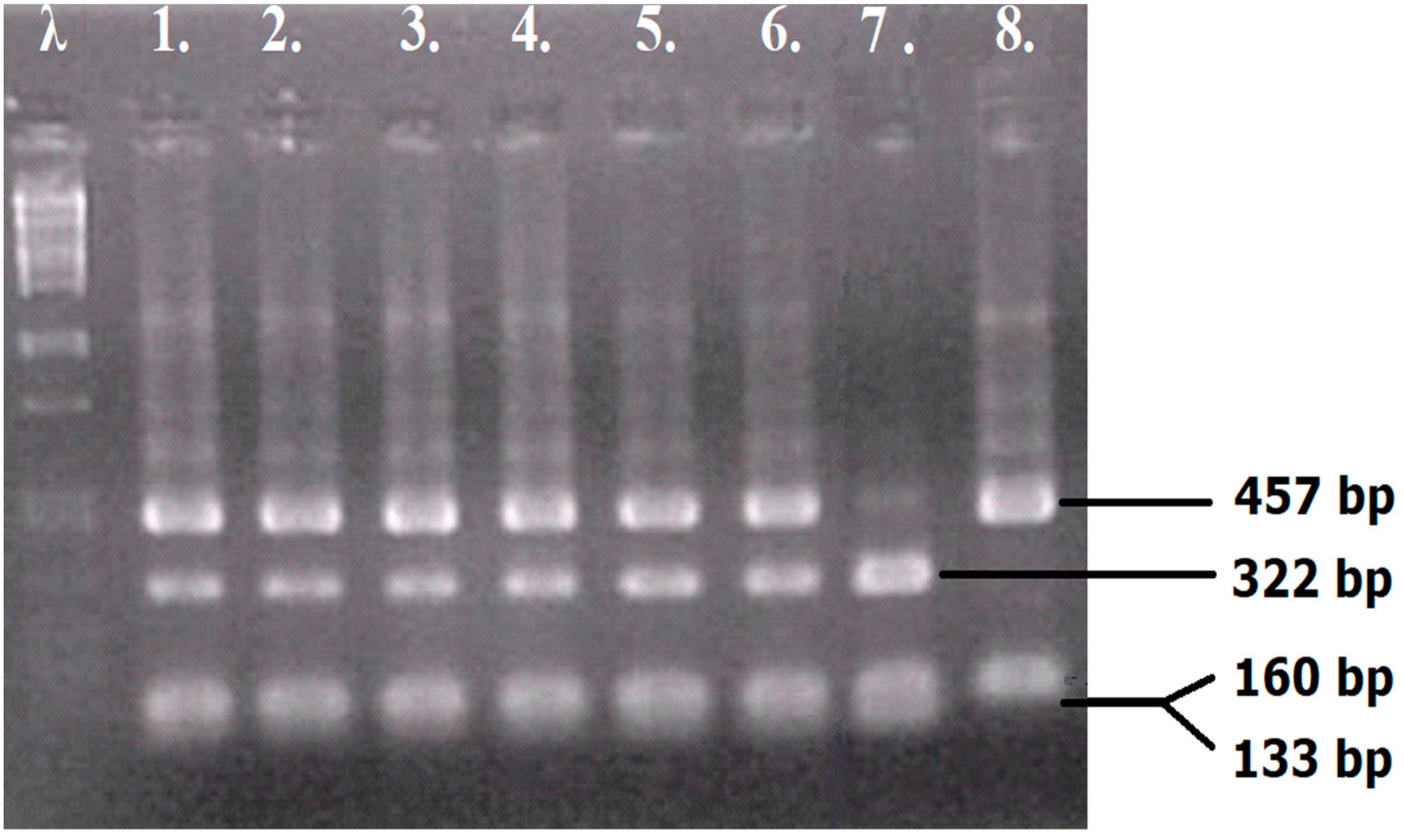
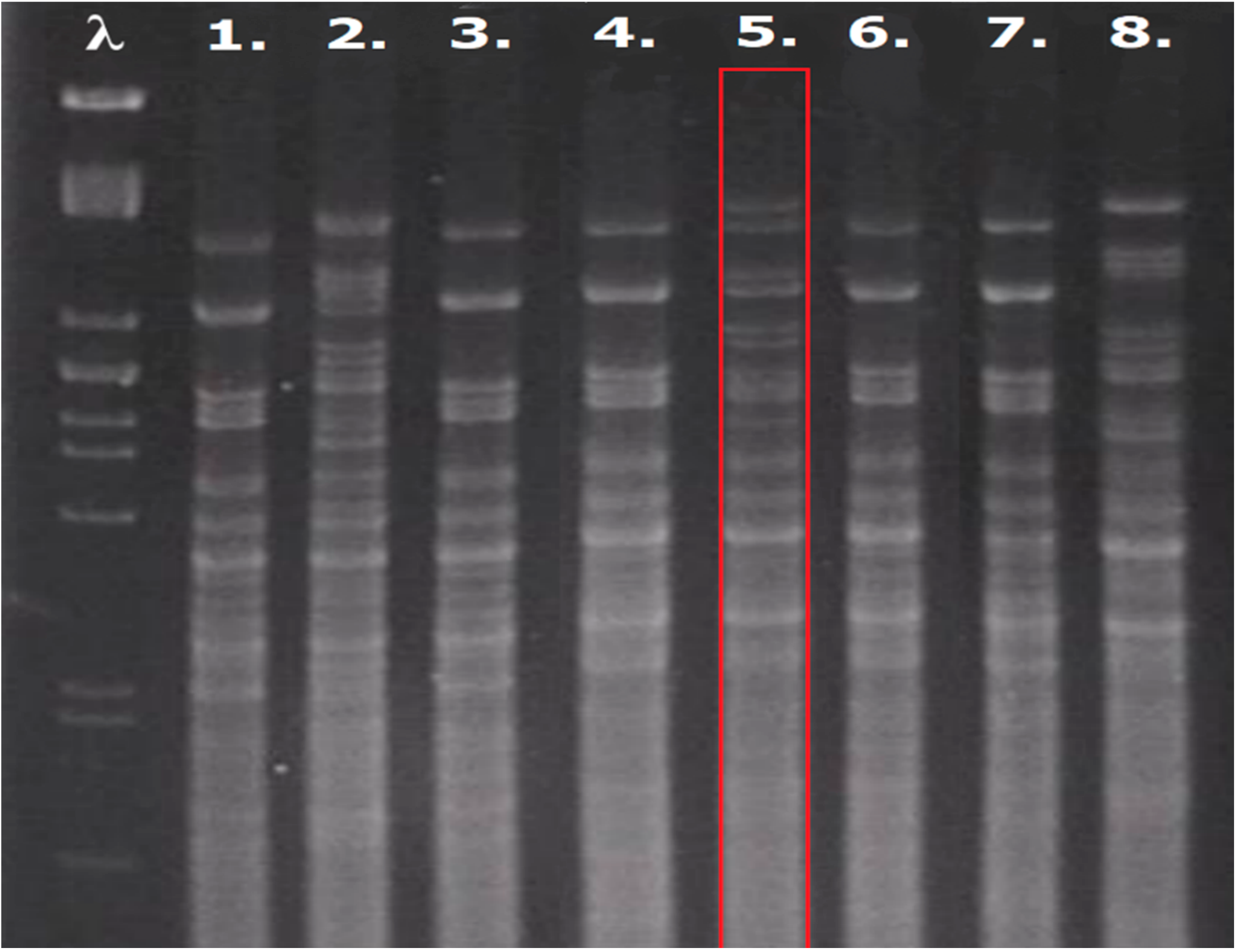
2.3. The Construction and Verification of Hybrids
2.3.1. Yeast Starters
2.3.2. Sugar Fermentation
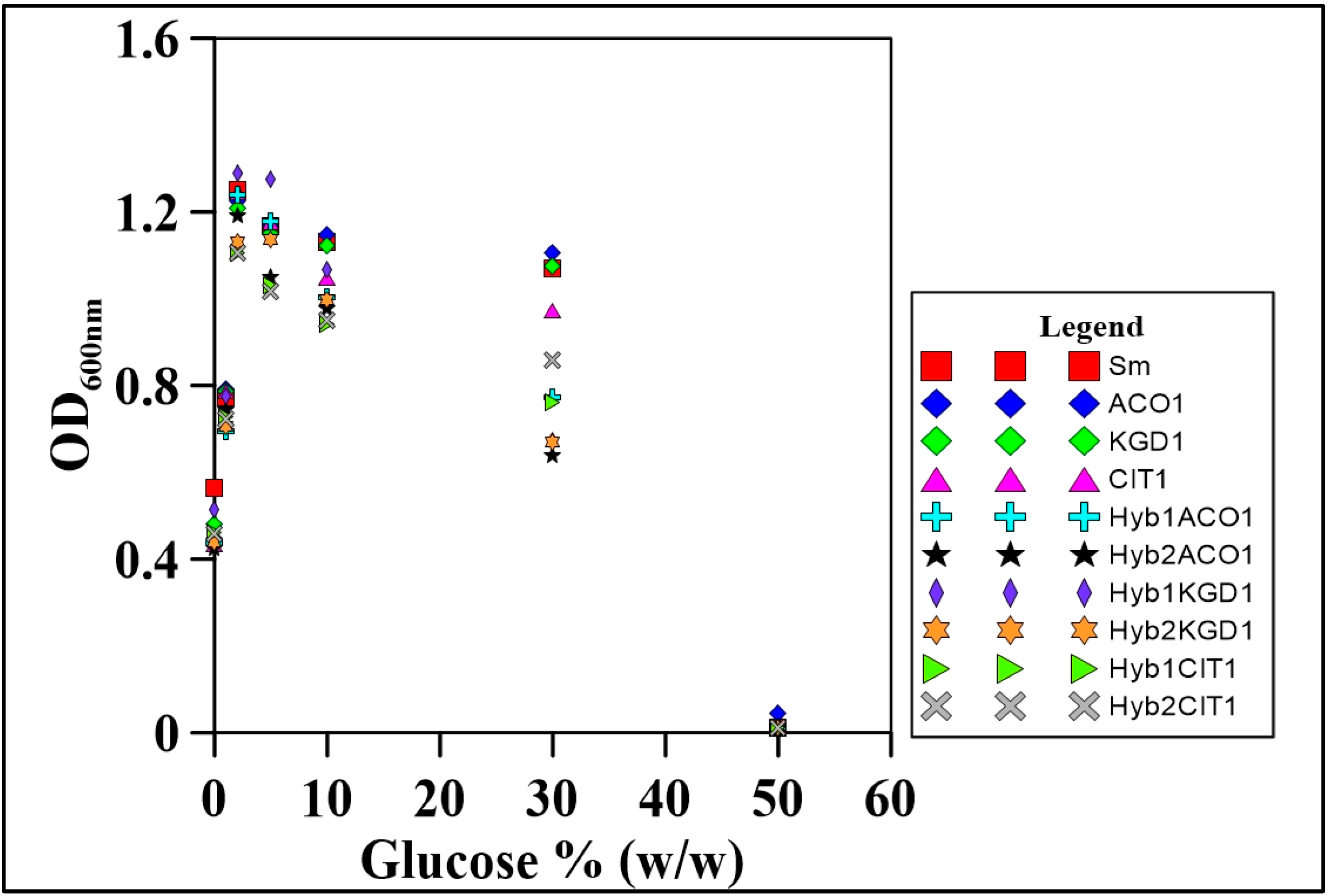
2.3.3. Osmotolerance
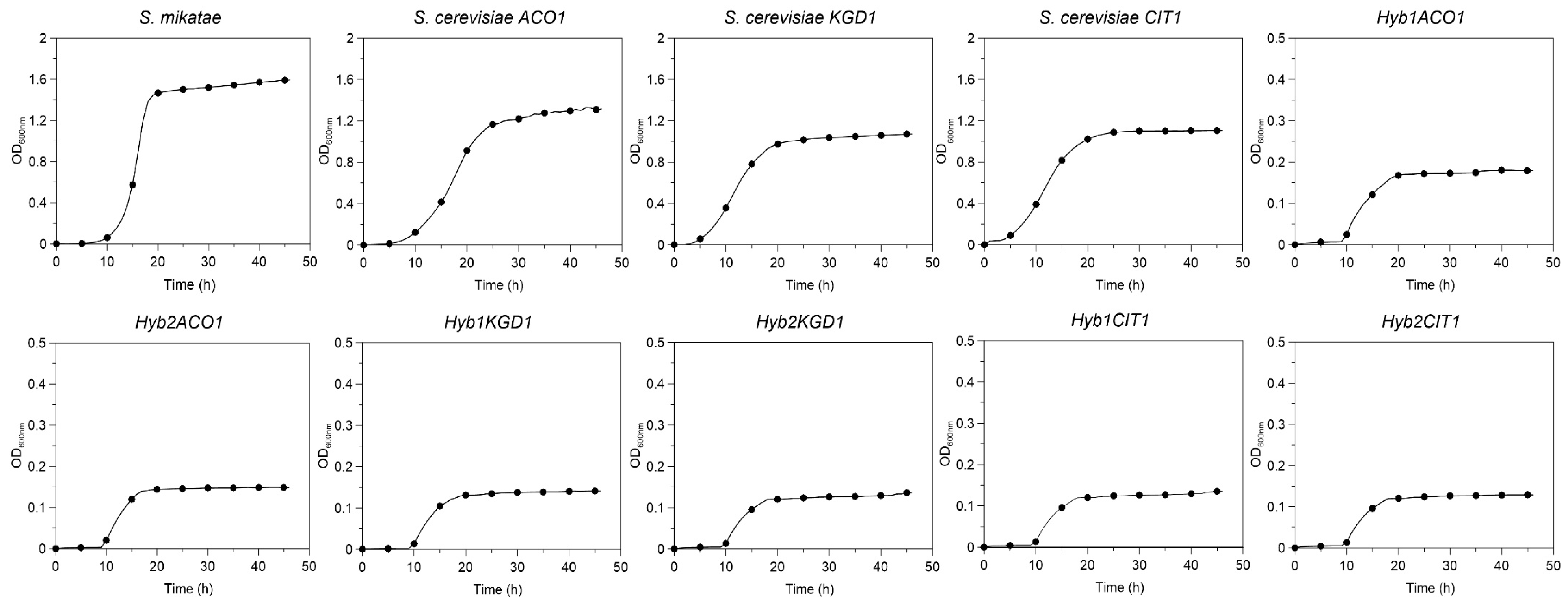
2.3.4. Aerobic Growth
2.3.5. Phenotype verification of Saccharomyces cerevisiae Mutants
2.4. Beer Production
2.5. Analytical Methods
2.5.1. Analysis of Beer by HPLC-RID
2.5.2. Analysis of Beer by HS-SPME-GC-MS
2.6. Statistical Methods
3. Results and Discussion
3.1. Phenotype Verification of Saccharomyces cerevisiae Mutants
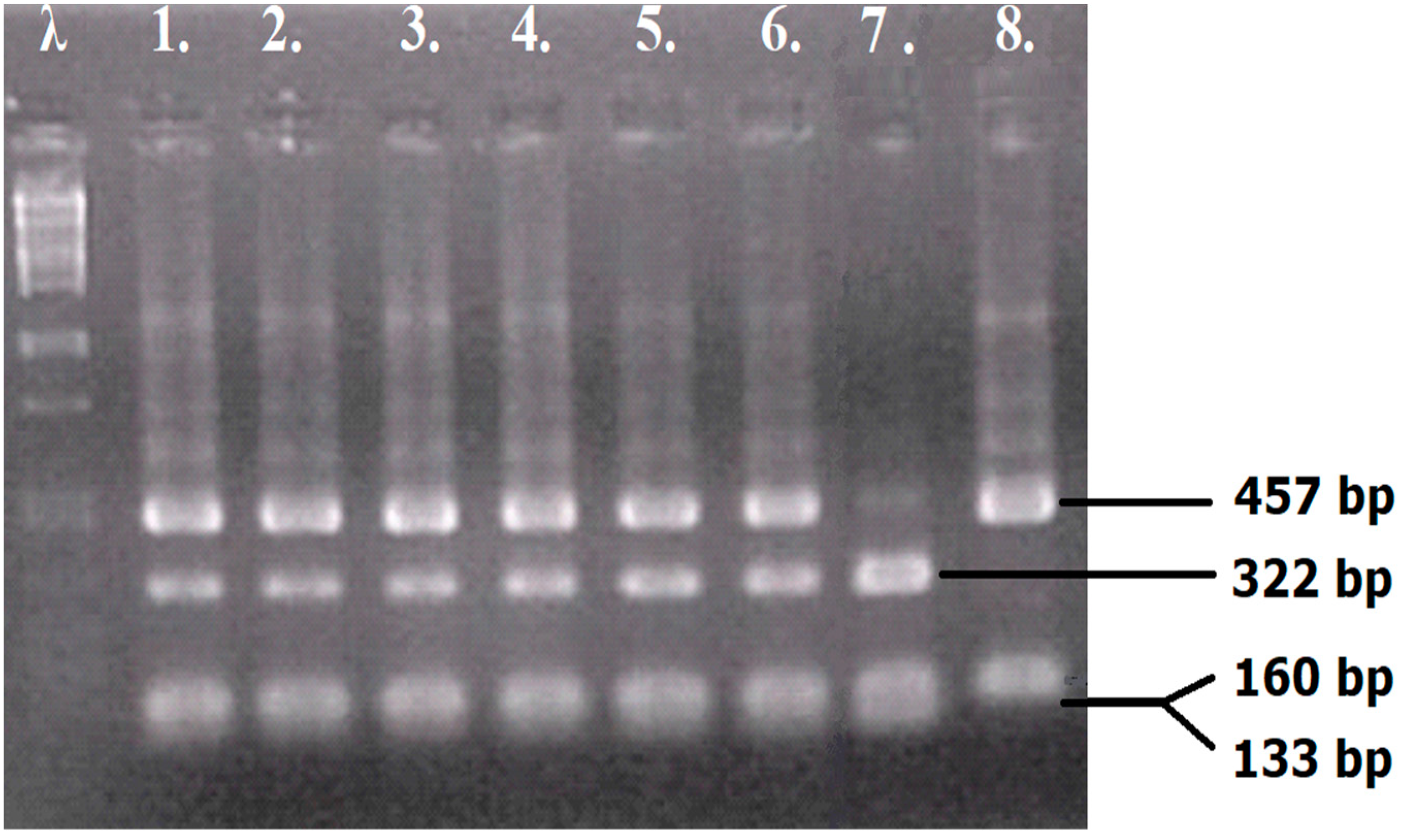
3.2. The Preparation of Yeast Hybrids
3.3. Saccharide Fermentation Tests
3.4. Osmotolerance
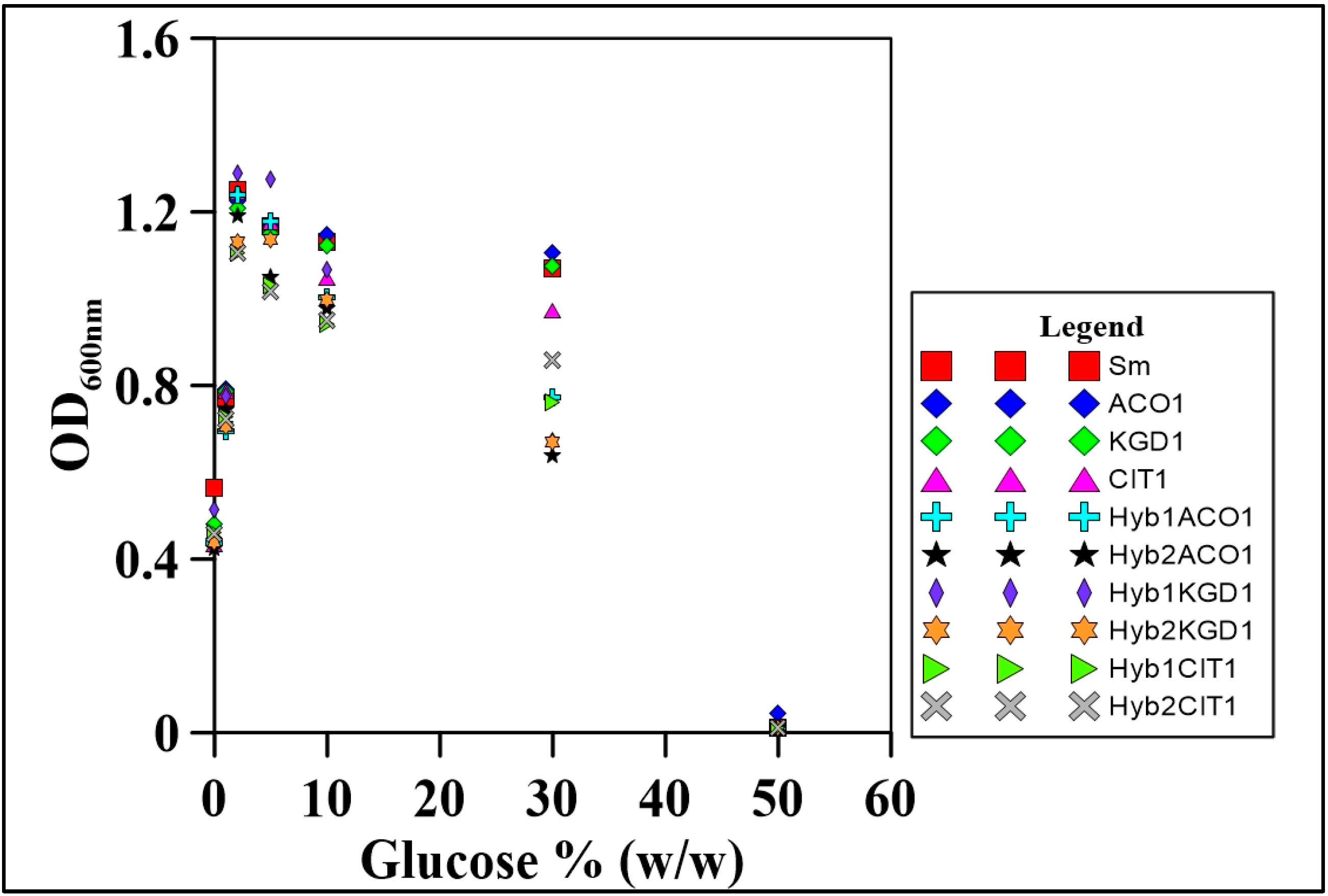
3.5. Aerobic Growth
3.6. Beer Production
3.7. Production of Volatile Organic Compounds (VOCs)
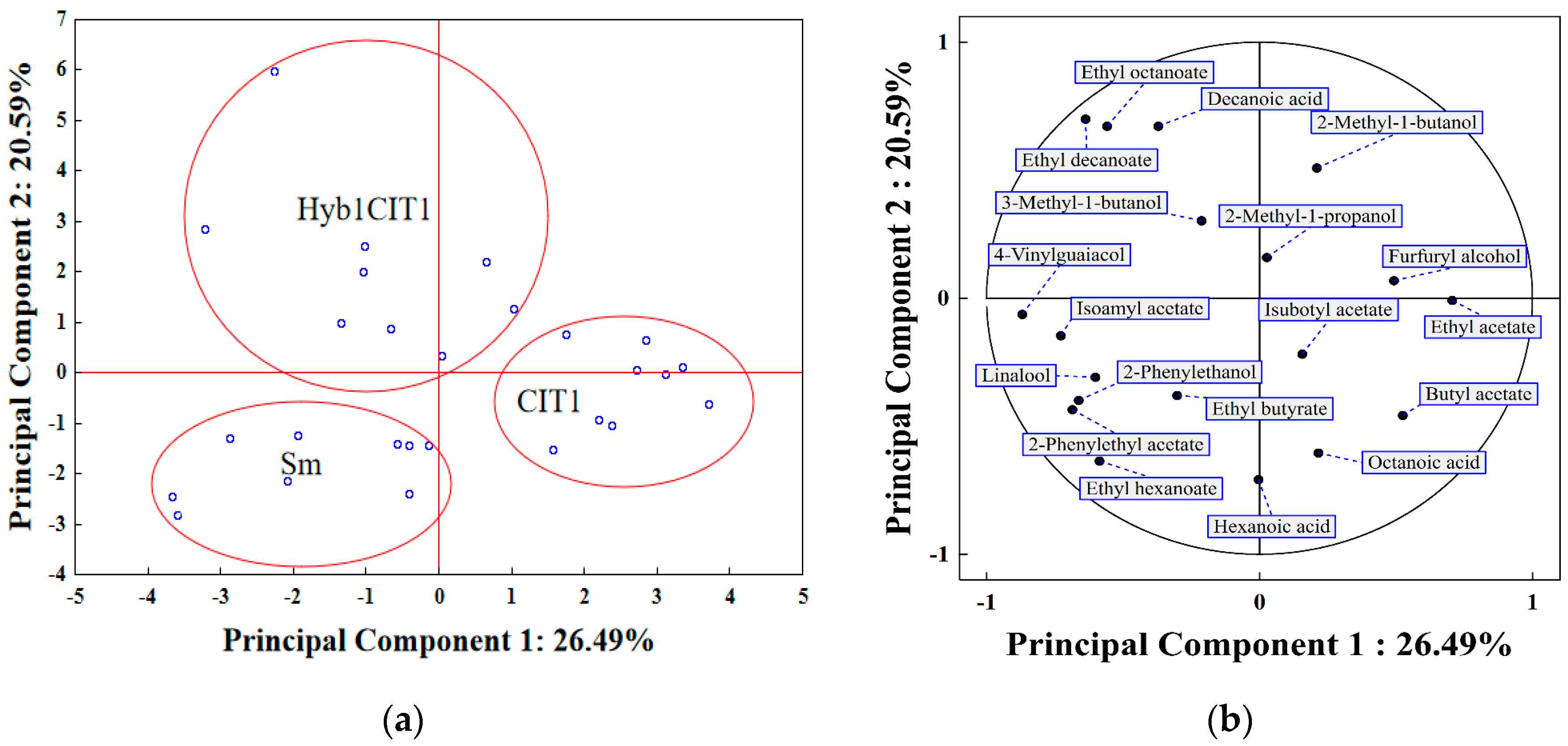
3.8. Principal Component Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Staub, C.; Contiero, R.; Bosshart, N.; Siegrist, M. You Are What You Drink: Stereotypes about Consumers of Alcoholic and Non-Alcoholic Beer. Food Qual. Prefer. 2022, 101, 104633. [Google Scholar] [CrossRef]
- Karaoglan, S.Y.; Jung, R.; Gauthier, M.; Kinčl, T.; Dostálek, P. Maltose-Negative Yeast in Non-Alcoholic and Low-Alcoholic Beer Production. Fermentation 2022, 8, 273. [Google Scholar] [CrossRef]
- Varela, J.; Varela, C. Microbiological Strategies to Produce Beer and Wine with Reduced Ethanol Concentration. Curr. Opin. Biotechnol. 2019, 56, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Bellut, K.; Michel, M.; Zarnkow, M.; Hutzler, M.; Jacob, F.; Atzler, J.J.; Hoehnel, A.; Lynch, K.M.; Arendt, E.K. Screening and Application of Cyberlindnera Yeasts to Produce a Fruity, Non-Alcoholic Beer. Fermentation 2019, 5, 103. [Google Scholar] [CrossRef] [Green Version]
- De Francesco, G.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Screening of New Strains of Saccharomycodes Ludwigii and Zygosaccharomyces Rouxii to Produce Low-Alcohol Beer: Screening of New Strains of S. Ludwigii and Z. Rouxii. J. Inst. Brew. 2015, 121, 113–121. [Google Scholar] [CrossRef]
- Brányik, T.; Silva, D.P. A Review of Methods of Low Alcohol and Alcohol-Free Beer Production. J. Food Eng. 2012, 108, 493–506. [Google Scholar] [CrossRef]
- Methner, Y.; Hutzler, M.; Zarnkow, M.; Prowald, A.; Endres, F.; Jacob, F. Investigation of Non- Saccharomyces Yeast Strains for Their Suitability for the Production of Non-Alcoholic Beers with Novel Flavor Profiles. J. Am. Soc. Brew. Chem. 2022, 80, 341–355. [Google Scholar] [CrossRef]
- Johansson, L.; Nikulin, J.; Juvonen, R.; Krogerus, K.; Magalhães, F.; Mikkelson, A.; Nuppunen-Puputti, M.; Sohlberg, E.; de Francesco, G.; Perretti, G.; et al. Sourdough Cultures as Reservoirs of Maltose-Negative Yeasts for Low-Alcohol Beer Brewing. Food Microbiol. 2021, 94, 103629. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, G.; Sannino, C.; Sileoni, V.; Marconi, O.; Filippucci, S.; Tasselli, G.; Turchetti, B. Mrakia gelida in Brewing Process: An Innovative Production of Low Alcohol Beer Using a Psychrophilic Yeast Strain. Food Microbiol. 2018, 76, 354–362. [Google Scholar] [CrossRef]
- Palanca, L.; Gaskett, A.C.; Günther, C.S.; Newcomb, R.D.; Goddard, M.R. Quantifying Variation in the Ability of Yeasts to Attract Drosophila melanogaster. PLoS ONE 2013, 8, e75332. [Google Scholar] [CrossRef] [Green Version]
- Salanță, L.C.; Coldea, T.E.; Ignat, M.V.; Pop, C.R.; Tofană, M.; Mudura, E.; Borșa, A.; Pasqualone, A.; Zhao, H. Non-Alcoholic and Craft Beer Production and Challenges. Processes 2020, 8, 1382. [Google Scholar] [CrossRef]
- Vilela, A. An Overview of CRISPR-Based Technologies in Wine Yeasts to Improve Wine Flavor and Safety. Fermentation 2021, 7, 5. [Google Scholar] [CrossRef]
- Navrátil, M.; Dömény, Z.; Šturdík, E.; Šmogrovičová, D.; Gemeiner, P. Production of Non-Alcoholic Beer Using Free and Immobilized Cells of Saccharomyces cerevisiae Deficient in the Tricarboxylic Acid Cycle. Biotechnol. Appl. Biochem. 2002, 35, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Selecký, R.; Šmogrovičová, D.; Šulák, M. Low-Alcoholic Beer Production Using Mutant Brewing Yeast. Kvas. Prum. 2005, 51, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Selecký, R.; Šmogrovičová, D.; Sulo, P. Beer with Reduced Ethanol Content Produced Using Saccharomyces cerevisiae Yeasts Deficient in Various Tricarboxylic Acid Cycle Enzymes. J. Inst. Brew. 2008, 114, 97–101. [Google Scholar] [CrossRef]
- Vaštík, P.; Šmogrovičová, D.; Kafková, V.; Sulo, P.; Furdíková, K.; Špánik, I. Production and Characterisation of Non-Alcoholic Beer Using Special Yeast. Kvas. Prum. 2020, 66, 336–344. [Google Scholar] [CrossRef]
- Molitor, R.W.; Roop, J.I.; Denby, C.M.; Depew, C.J.; Liu, D.S.; Stadulis, S.E.; Shellhammer, T.H. The Sensorial and Chemical Changes in Beer Brewed with Yeast Genetically Modified to Release Polyfunctional Thiols from Malt and Hops. Fermentation 2022, 8, 370. [Google Scholar] [CrossRef]
- Long, D.; Wilkinson, K.; Taylor, D.; Jiranek, V. Novel Wine Yeast for Improved Utilisation of Proline during Fermentation. Fermentation 2018, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; Gebbia, M.; Belotti, A.; Foschino, R.; Roth, F.P. CRISPR/Cas9 System as a Valuable Genome Editing Tool for Wine Yeasts with Application to Decrease Urea Production. Front. Microbiol. 2017, 8, 2194. [Google Scholar] [CrossRef] [PubMed]
- Bendixsen, D.P.; Frazão, J.G.; Stelkens, R. Saccharomyces Yeast Hybrids on the Rise. Yeast 2022, 39, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Krogerus, K.; Seppänen-Laakso, T.; Castillo, S.; Gibson, B. Inheritance of Brewing-Relevant Phenotypes in Constructed Saccharomyces cerevisiae × Saccharomyces eubayanus Hybrids. Microb. Cell Fact. 2017, 16, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catallo, M.; Iattici, F.; Randazzo, C.L.; Caggia, C.; Krogerus, K.; Magalhães, F.; Gibson, B.; Solieri, L. Hybridization of Saccharomyces cerevisiae Sourdough Strains with Cryotolerant Saccharomyces bayanus NBRC1948 as a Strategy to Increase Diversity of Strains Available for Lager Beer Fermentation. Microorganisms 2021, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Bellon, J.R.; Schmid, F.; Capone, D.L.; Dunn, B.L.; Chambers, P.J. Introducing a New Breed of Wine Yeast: Interspecific Hybridisation between a Commercial Saccharomyces cerevisiae Wine Yeast and Saccharomyces mikatae. PLoS ONE 2013, 8, e62053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruner, J.; Fox, G. Novel Non-cerevisiae Saccharomyces Yeast Species Used in Beer and Alcoholic Beverage Fermentations. Fermentation 2020, 6, 116. [Google Scholar] [CrossRef]
- Gyurchev, N.Y.; Coral-Medina, Á.; Weening, S.M.; Almayouf, S.; Kuijpers, N.G.A.; Nevoigt, E.; Louis, E.J. Beyond Saccharomyces pastorianus for modern lager brews: Exploring non-cerevisiae Saccharomyces hybrids with heterotic maltotriose consumption and novel aroma profile. Front. Microbiol. 2022, 13, 1–14. [Google Scholar] [CrossRef]
- Peris, D.; Ubbelohde, E.J.; Kuang, M.C.; Kominek, J.; Langdon, Q.K.; Adams, M.; Koshalek, J.A.; Hulfachor, A.B.; Opulente, D.A.; Hall, D.J.; et al. Macroevolutionary diversity of traits and genomes in the model yeast genus Saccharomyces. BioRxiv 2022. preprint. [Google Scholar] [CrossRef]
- Brachmann, C.B.; Davies, A.; Cost, G.J.; Caputo, E.; Li, J.; Hieter, P.; Boeke, J.D. Designer Deletion Strains Derived from Saccharomyces cerevisiae S288C: A Useful Set of Strains and Plasmids for PCR-Mediated Gene Disruption and Other Applications. Yeast 1998, 14, 115–132. [Google Scholar] [CrossRef]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Véronneau, S.; Dow, S.; Lucau-Danila, A.; Anderson, K.; André, B.; et al. Functional Profiling of the Saccharomyces cerevisiae Genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Philippsen, P.; Stotz, A.; Scherf, C. DNA of Saccharomyces cerevisiae. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1991; Volume 194, pp. 169–181. [Google Scholar]
- Defontaine, A.; Lecocq, F.M.; Hallet, J.N. A Rapid Miniprep Method for the Preparation of Yeast Mitochondrial DNA. Nucleic Acids Res. 1991, 19, 185. [Google Scholar] [CrossRef] [Green Version]
- Bágeľová, P.S.; Lichtner, Ž.; Szemes, T.; Smolejová, M.; Sulo, P. Mitochondrial DNA Duplication, Recombination, and Introgression during Interspecific Hybridization. Sci. Rep. 2021, 11, 12726. [Google Scholar] [CrossRef] [PubMed]
- Špírek, M.; Poláková, S.; Jatzová, K.; Sulo, P. Post-zygotic sterility and cytonuclear com-patibility limits in S. cerevisiae xenomitochondrial cybrids. Front. Genet. 2015, 5, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchez, A.; De Vuyst, L. Acetic Acid Bacteria in Sour Beer Production: Friend or Foe? Front. Microbiol. 2022, 13, 957167. [Google Scholar] [CrossRef]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe Domestication and the Identification of the Wild Genetic Stock of Lager-Brewing Yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef] [Green Version]
- Langdon, Q.K.; Peris, D.; Baker, E.P.; Opulente, D.A.; Nguyen, H.-V.; Bond, U.; Gonçalves, P.; Sampaio, J.P.; Libkind, D.; Hittinger, C.T. Fermentation Innovation through Complex Hybridization of Wild and Domesticated Yeasts. Nat. Ecol. Evol. 2019, 3, 1576–1586. [Google Scholar] [CrossRef] [PubMed]
- Sulo, P.; Szabóová, D.; Bielik, P.; Poláková, S.; Šoltys, K.; Jatzová, K.; Szemes, T. The Evolutionary History of Saccharomyces Species Inferred from Completed Mitochondrial Genomes and Revision in the ‘Yeast Mitochondrial Genetic Code’. DNA Res. 2017, 24, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Westermann, B. Mitochondrial Inheritance in Yeast. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2014, 1837, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- Dawes, I. The Molecular Biology of the Yeast Saccharomyces. Life Cycle and Inheritance. Biochem. Educ. 1983, 11, 81. [Google Scholar] [CrossRef]
- Briggs, D.; Boulton, C.; Brookes, P.; Stevens, R. Brewing: Science and Practice; CRC Press: Boca Raton, FL, USA, 2004; ISBN 978-0-8493-2547-2. [Google Scholar]
- Babazadeh, R.; Lahtvee, P.J.; Adiels, C.B.; Goksör, M.; Nielsen, J.B.; Hohmann, S. The Yeast Osmostress Response Is Carbon Source Dependent. Sci. Rep. 2017, 7, 990. [Google Scholar] [CrossRef] [Green Version]
- Bernardes, J.P.; Stelkens, R.B.; Greig, D. Heterosis in Hybrids within and between Yeast Species. J. Evol. Biol. 2017, 30, 538–548. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-López, F.N.; Pérez-Torrado, R.; Querol, A.; Barrio, E. Modulation of the Glycerol and Ethanol Syntheses in the Yeast Saccharomyces kudriavzevii Differs from That Exhibited by Saccharomyces cerevisiae and Their Hybrid. Food Microbiol. 2010, 27, 628–637. [Google Scholar] [CrossRef]
- Vaštík, P.; Rosenbergová, Z.; Furdíková, K.; Klempová, T.; Šišmiš, M.; Šmogrovičová, D. Potential of Non- Saccharomyces Yeast to Produce Non-Alcoholic Beer. FEMS Yeast Res. 2022, 22, foac039. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, F. Changes in Organic Acids During Beer Fermentation. J. Am. Soc. Brew. Chem. 2015, 73, 275–279. [Google Scholar] [CrossRef]
- Park, J.-M.; Shin, J.-A.; Lee, J.H.; Lee, K.-T. Development of a Quantitative Method for Organic Acid in Wine and Beer Using High Performance Liquid Chromatography. Food Sci. Biotechnol. 2017, 26, 349–355. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, S.; Zhang, W. Predicting Acetic Acid Content in the Final Beer Using Neural Networks and Support Vector Machine. J. Inst. Brew. 2012, 118, 361–367. [Google Scholar] [CrossRef]
- Dack, R.E.; Black, G.W.; Koutsidis, G.; Usher, S.J. The Effect of Maillard Reaction Products and Yeast Strain on the Synthesis of Key Higher Alcohols and Esters in Beer Fermentations. Food Chem. 2017, 232, 595–601. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters Affecting Ethyl Ester Production by Saccharomyces Cerevisiae during Fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Nešpor, J.; Andrés-Iglesias, C.; Karabín, M.; Montero, O.; Blanco, C.A.; Dostálek, P. Volatile Compound Profiling in Czech and Spanish Lager Beers in Relation to Used Production Technology. Food Anal. Methods 2019, 12, 2293–2305. [Google Scholar] [CrossRef]
- Krogerus, K.; Eurikäinen, R.; Aisala, H.; Gibson, B. Repurposing Brewery Contaminant Yeast as Production Strains for Low-alcohol Beer Fermentation. Yeast 2021, 39, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Giannakou, K.; Visinoni, F.; Zhang, P.; Nathoo, N.; Jones, P.; Cotterrell, M.; Vrhovsek, U.; Delneri, D. Biotechnological Exploitation of Saccharomyces jurei and Its Hybrids in Craft Beer Fermentation Uncovers New Aroma Combinations. Food Microbiol. 2021, 100, 103838. [Google Scholar] [CrossRef]
- Buiatti, S. Beer Composition: An Overview. In Beer in Health and Disease Prevention; Preedy, V.P., Ed.; Elsevier: Oxford, UK, 2009; pp. 213–225. ISBN 978-0-12-373891-2. [Google Scholar]
- Holt, S.; Miks, M.H.; de Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. The Molecular Biology of Fruity and Floral Aromas in Beer and Other Alcoholic Beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.H. Whisky Science: A Condensed Distillation; Springer International Publishing: Cham, Switzerland, 2019; ISBN 978-3-030-13731-1. [Google Scholar]
- Anderson, H.E.; Santos, I.C.; Hildenbrand, Z.L.; Schug, K.A. A Review of the Analytical Methods Used for Beer Ingredient and Finished Product Analysis and Quality Control. Anal. Chim. Acta 2019, 1085, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Senkarcinova, B.; Graça Dias, I.A.; Nespor, J.; Branyik, T. Probiotic Alcohol-Free Beer Made with Saccharomyces cerevisiae var. boulardii. LWT 2019, 100, 362–367. [Google Scholar] [CrossRef]
- Hughes, P. Identification of Taste- and Aroma-Active Components of Beer. In Beer in Health and Disease Prevention; Preedy, V.P., Ed.; Elsevier: Oxford, UK, 2009; pp. 227–238. ISBN 978-0-12-373891-2. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast | Genotype | Abbreviation (Type) |
|---|---|---|
| Saccharomyces mikatae CBS* 8839T | Wild type | Sm (parent) |
| Saccharomyces cerevisiae ΔACO1 G418R | MATα, his3Δ1, leu2Δ0, lys2Δ0, ura3Δ0, aco1::kanMX4 | ACO1 (parent) |
| Saccharomyces cerevisiae ΔKGD1 G418R | MATα, his3Δ1, leu2Δ0, lys2Δ0, ura3Δ0, kgd11::kanMX4 | KGD1 (parent) |
| Saccharomyces cerevisiae ΔCIT1 G418R | MATα, his3Δ1, leu2Δ0, lys2Δ0, ura3Δ0, cit1::kanMX4 | CIT1 (parent) |
| S. mikatae CBS 8839T × S. cerevisiae ΔACO1 G418R | Hyb1ACO1 (hybrid) | |
| S. mikatae CBS 8839T × S. cerevisiae ΔACO1 G418R | Hyb2ACO1 (hybrid) | |
| S. mikatae CBS 8839T × S. cerevisiae ΔKGD1 G418R | Hyb1KGD1 (hybrid) | |
| S. mikatae CBS 8839T × S. cerevisiae ΔKGD1 G418R | Hyb2KGD1 (hybrid) | |
| S. mikatae CBS 8839T × S. cerevisiae ΔCIT1 G418R | Hyb1CIT1 (hybrid) | |
| S. mikatae CBS 8839T × S. cerevisiae ΔCIT1 G418R | Hyb2CIT1 (hybrid) | |
| Saccharomyces cerevisiae K97 | K97 (top fermenting control) | |
| Saccharomyces pastorianus W34/70 | W34/70 (bottom fermenting control) |
| Direction | Name of Primer | Sequence 5’→3’ |
|---|---|---|
| Forward | D1/D2-NL1 | GCATATCAATAAGCGGAGGAAAAG |
| Reverse | D1/D2-NL4 | GGTCCGTGTTTCAAGACGG |
| Citric Acid | Malic Acid | Succinic Acid | Acetic Acid | |||||
|---|---|---|---|---|---|---|---|---|
| Producer | 8°P | 10°P | 8°P | 10°P | 8°P | 10°P | 8°P | 10°P |
| S. cerevisiae ACO1 | 0.32 ± 0.01 | 0.45 ± 0.04 | 0.54 ± 0.01 | 0.62 ± 0.02 | 0.29 ± 0.05 | 0.31 ± 0.01 | 0.22 ± 0.03 | 0.66 ± 0.03 |
| S. cerevisiae KGD1 | 0.22 ± 0.01 | 0.47 ± 0.04 | 0.53 ± 0.01 | 0.55 ± 0.02 | 0.32 ± 0.01 | 0.74 ± 0.02 | 0.23 ± 0.02 | 0.64 ± 0.03 |
| S. cerevisiae CIT1 | 0.37 ± 0.01 | 0.52 ± 0.03 | 0.63 ± 0.05 | 0.73 ± 0.02 | 0.11 ± 0.00 | 0.31 ± 0.01 | 0.21 ± 0.02 | 0.62 ± 0.02 |
| S. cerevisiae K97 | 0.07 ± 0.00 | 0.28 ± 0.01 | 0.04 ± 0.00 | 0.15 ± 0.00 | n.d. | n.d. | n.d. | 0.04 ± 0.00 |
| S. pastorianus W34/70 | 0.08 ± 0.00 | n.d. | 0.04 ± 0.00 | 0.16 ± 0.00 | n.d. | n.d. | n.d. | 0.06 ± 0.00 |
| Fermentation | ||||
|---|---|---|---|---|
| Yeast | Glucose | Maltose | Maltotriose | Sucrose |
| Hyb1ACO1 | + | − | − | + |
| Hyb2ACO1 | + | − | − | + |
| Hyb1KGD1 | + | − | − | + |
| Hyb2KGD1 | + | − | − | + |
| Hyb1CIT1 | + | − | − | + |
| Hyb2CIT1 | + | − | − | + |
| S. cerevisiae K97 | + | + | + | + |
| S. pastorianus W34/70 | + | + | + | + |
| Sample | Alcohol % (v/v) | Original Extract (°P) | Real Degree of Fermentation (%) | pH | Colour (EBC) |
|---|---|---|---|---|---|
| 7°P wort | 0.00 ± 0.00 | 7.19 ± 0.01 | 0.00 ± 0.00 | 6.05 ± 0.03 | 19.34 ± 0.35 |
| Sm | 0.03 ± 0.00 | 7.17 ± 0.01 | 0.68 ± 0.01 | 5.63 ± 0.02 | 16.69 ± 0.28 |
| CIT1 | 0.03 ± 0.00 | 7.18 ± 0.01 | 0.67 ± 0.07 | 5.59 ± 0.02 | 18.36 ± 0.35 |
| Hyb1CIT1 | 0.02 ± 0.00 | 7.17 ± 0.01 | 0.49 ± 0.04 | 5.72 ± 0.04 | 17.35 ± 0.27 |
| Organic Compound | 7°P Wort | Sm | CIT1 | Hyb1CIT1 |
|---|---|---|---|---|
| Glucose | 4.50 ± 0.07 | n.d. | n.d. | n.d. |
| Maltose | 34.65 ± 0.67 | 33.51 ± 0.98 | 34.34 ± 0.65 | 33.64 ± 0.76 |
| Maltotriose | 8.42 ± 0.14 | 8.37 ± 0.28 | 8.38 ± 0.24 | 8.04 ± 0.38 |
| Glycerol | n.d. | 0.62 ± 0.10 | 0.65 ± 0.07 | 0.63 ± 0.08 |
| Ethanol | n.d. | 0.23 ± 0.01 | 0.23 ± 0.01 | 0.16 ± 0.00 |
| Acetic acid | n.d. | n.d. | 0.21 ± 0.01 | n.d. |
| Citric acid | n.d. | 0.17 ± 0.01 | 0.24 ± 0.03 | 0.23 ± 0.01 |
| Malic acid | n.d. | 0.35 ± 0.03 | 0.52 ± 0.09 | 0.55 ± 0.02 |
| Succinic acid | n.d. | n.d. | n.d. | 0.04 ± 0.00 |
| Organic Compound (mg L−1) | Sm | CIT1 | Hyb1CIT1 |
|---|---|---|---|
| Esters | |||
| Ethyl acetate | 0.245 ± 0.079 | 0.369 ± 0.090 | 0.262 ± 0.039 |
| Propyl acetate | n.d. | n.d. | n.d. |
| Butyl acetate | 0.018 ± 0.005 | 0.022 ± 0.005 | 0.014 ± 0.006 |
| Isoamyl acetate | 0.003 ± 0.000 | 0.002 ± 0.000 | 0.002 ± 0.000 |
| Isobutyl acetate | 0.010 ± 0.006 | 0.012 ± 0.003 | 0.010 ± 0.005 |
| 2-Phenylethylacetate | 0.004 ± 0.001 | 0.002 ± 0.000 | 0.003 ± 0.001 |
| Ethyl butanoate | 0.029 ± 0.005 | 0.025 ± 0.003 | 0.026 ± 0.004 |
| Ethyl hexanoate | 0.274 ± 0.020 | 0.212 ± 0.018 | 0.208 ± 0.012 |
| Ethyl octanoate | 0.004 ± 0.001 | 0.003 ± 0.000 | 0.006 ± 0.002 |
| Ethyl decanoate | 0.014 ± 0.002 | 0.007 ± 0.001 | 0.025 ± 0.011 |
| Higher Alcohols | |||
| 3-Methyl-1-butanol | 0.714 ± 0.247 | 0.746 ± 0.132 | 0.884 ± 0.122 |
| 2-Methyl-1-propanol | 1.381 ± 0.403 | 1.531 ± 0.207 | 1.638 ± 0.247 |
| 2-Methyl-1-butanol | 0.371 ± 0.069 | 0.457 ± 0.067 | 0.485 ± 0.059 |
| 2-Phenylethanol | 0.533 ± 0.070 | 0.435 ± 0.054 | 0.469 ± 0.025 |
| Butane-2,3-diol | n.d. | n.d. | n.d. |
| Furfuryl alcohol | 0.072 ± 0.037 | 0.210 ± 0.073 | 0.145 ± 0.072 |
| Terpene alcohols | |||
| Linalool | 0.097 ± 0.011 | 0.088 ± 0.006 | 0.091 ± 0.003 |
| Phenols | |||
| 4-Vinylguaiacol | 0.466 ± 0.060 | 0.258 ± 0.051 | 0.425 ± 0.079 |
| Fatty acids | |||
| Hexanoic acid (caproic) | 0.202 ± 0.051 | 0.173 ± 0.045 | 0.130 ± 0.057 |
| Octanoic acid (caprylic) | 0.010 ± 0.002 | 0.011 ± 0.003 | 0.009 ± 0.002 |
| Decanoic acid (capric) | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.004 ± 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaštík, P.; Sulo, P.; Rosenbergová, Z.; Klempová, T.; Dostálek, P.; Šmogrovičová, D. Novel Saccharomyces cerevisiae × Saccharomyces mikatae Hybrids for Non-alcoholic Beer Production. Fermentation 2023, 9, 221. https://doi.org/10.3390/fermentation9030221
Vaštík P, Sulo P, Rosenbergová Z, Klempová T, Dostálek P, Šmogrovičová D. Novel Saccharomyces cerevisiae × Saccharomyces mikatae Hybrids for Non-alcoholic Beer Production. Fermentation. 2023; 9(3):221. https://doi.org/10.3390/fermentation9030221
Chicago/Turabian StyleVaštík, Peter, Pavol Sulo, Zuzana Rosenbergová, Tatiana Klempová, Pavel Dostálek, and Daniela Šmogrovičová. 2023. "Novel Saccharomyces cerevisiae × Saccharomyces mikatae Hybrids for Non-alcoholic Beer Production" Fermentation 9, no. 3: 221. https://doi.org/10.3390/fermentation9030221