

Enhancement of Bioactive Properties in Momordica charantia by Leuconostoc Fermentation


1
Department of Food Science and Nutrition, Dankook University, Cheonan 31116, Republic of Korea
2
Research Center for Industrialization of Natural Neutralization, Dankook University, Yongin 16890, Republic of Korea
3
Department of Clinical Laboratory Science, Semyung University, Jecheon 27136, Republic of Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Fermentation 2023, 9(6), 523; https://doi.org/10.3390/fermentation9060523
Submission received: 1 May 2023
/
Revised: 25 May 2023
/
Accepted: 25 May 2023
/
Published: 29 May 2023
(This article belongs to the Special Issue Health and Bioactive Compounds of Fermented Foods and By-Products)
Abstract
:Momordica charantia (MC; commonly known as bitter melon) was fermented with Leuconostoc mesenteroides MKJW, MKSR, and KCTC 3719 (LM), and Leuconosoc citreum KCTC 3526 (LC), and their anti-diabetic, anti-dementia, and antioxidant activities were evaluated. The fermentation was performed for 24 h at 30 °C, and non-fermented MCs (CON1 and CON2) were included for comparison. All fermented MCs produced lactic acids, mannitol, dextran, and oligosaccharides. The highest amount of mannitol (34.76 mg/mL) and lactic acids (10.42 mg/mL) were produced in MKSR-MC, and the highest amount of dextran (22.37%) was produced in MKJW-MC. MKSR-MC showed complete α-glucosidase inhibition (99.91%), but it did not show a significant change in α-amylase inhibition (24.43%) compared to non-fermented MC (20.14%) (p > 0.05). It was also high in acetylcholinesterase inhibition (55.24%) compared to other fermented MCs (31.21–44.46%). Fermentation increased butyrylcholinesterase inhibition, but no significant differences were observed among the groups. Therefore, our results indicated that MKSR-fermented MC might be used as a non-dairy probiotic plant extract to achieve multi-health functional activities such as anti-diabetic, anti-dementia, and antioxidant activities.
1. Introduction
Diabetes mellitus is the ninth leading cause of death, with an estimated 1.5 million mortalities directly caused by diabetes [1]. More than 90% of all diabetes cases are classified into type 2 diabetes mellitus (T2DM), which can be prevented and managed by lifestyle changes, such as diet and exercise, and medical treatment [2]. Epidemiological investigation has found that patients with T2DM are closely associated with dementia and cognitive impairment by some possible mechanisms such as insulin resistance, oxidative stress, and inflammatory cytokines [3,4,5]. Oxidative stress is also a crucial characteristic of T2DM and Alzheimer’s disease (AD), which is a type of dementia in pathology. Excessive oxidative stress promotes insulin resistance, β-amyloid plaque accumulation, and neurofibrillary entanglement in the brain, leading to occasional T2DM and AD [6,7]. Therefore, multi-targeted approaches, including anti-diabetic, anti-dementia, and antioxidant activity, might be necessary instead of treating each disease separately. In treating T2DM and AD, inhibitors against carbohydrate-hydrolyzing enzymes, and cholinesterase are individually used.
The probiotic administration to treat chronic diseases such as T2DM and AD have been promising according to several successive results such as improved hemoglobin A1C in patients with T2DM and increased serum kynurenine levels in patients with AD [8,9]. Previous studies have shown that Leuconostoc mesenteroides MKJW and Leuconostoc mesenteroides MKSR isolated from kimchi show probiotic effects with α-glucosidase inhibitory (AGI; 98% for MKJW and 87% for MKSR) and antioxidant (46% for MKJW and 82% for MKSR) or anti-dementia activity (51% for MKJW) [10,11]. Several studies have reported that fruits or plants fermented by probiotic strains activate various health functionalities such as the presence of GABA (γ-aminobutyric acid), high nutrition value, antioxidant activity, potential immunomodulatory properties related to proline content, and so on, in addition to their probiotic effects [12,13,14,15,16]. In a study of Demarinis et al. [17], the fermentation of legume-based water extract using probiotic Lactobacillus acidophuilus, Limosilactobacillus fermentum, or Lacticaseibacillus paracasei positively affected the nutrition value by increasing the total amino acid content and survivability of probiotic culture during cold storage, as well as the organoleptic profile. Tomato pastes fermented with Lactobacillus plantarum increased the total polyphenol and glutamine content and possessed antioxidant activity [15].
Momordica charantia L. (MC), which is commonly known as bitter melon, is a tropical and subtropical vine of the family Cucurbitaceae. It has been widely used as an edible or healthy functional fruit and oriental medicine. Its extracts using various solvents have been studied for health functionalities such as anti-diabetic, antioxidant, antibacterial, and anticancer activity due to various phytochemicals including phenolic compounds such as gallic acid, tannic acid, catechin, caffeic acid, flavonoids, glycoside, and carbohydrates [18,19,20]. In particular, it is known to be effective for diabetes because of its charantin content, which promotes insulin secretion and lowers blood sugar levels by activating the β-cells of the pancreas [21] However, it is a mixture of fat-soluble stigmasterol glucoside and beta-sitosterol glucoside, which is hardly soluble in water and ethanol. Therefore, the effective extraction of charantin from MC is sometimes challengeable for health functional food products [22]. Shu et al. [23] increased the charantin content by the fermentation of 10% soymilk-supplemented MC by Lactobacillus plantarum. They also reported that co-culture fermentation of MC using Lac. plantarum, Saccharomyces cerevisiae, and Gluconobacer for 6 days enhanced xanthin oxidase inhibition for potentially treating hyperuricemia. The fermentation of fresh MC juice by Lac. plantarum improved flavor profile by increasing alcohols and acids and reducing aldehydes and ketones, and it also enhanced antioxidant activity by increasing total phenolic and flavonoid content [24]. In a study of Gao et al. [25], Lactobacillus plantarum-fermented MC juice improved serum glucose and insulin and regulated lipid metabolism and gut microbiota in streptozotocin-induced diabetic rats. Wen et al. [26] reported that polysaccharides from Lactobacillus plantarum-fermented MC promoted the growth of beneficial bacteria such as Bifidobacterium spp. and Lactobacillus spp. but inhibited the growth of harmful bacteria such as Helicobacter.
Herein, a more acceptable fermentation strategy using water-extracted MC was attempted to develop a multi-target health functional product. MC was fermented by probiotic Leu. mesenteroides MKJW (MKJW) and Leu. mesenteroides MKSR (MKSR), and the distributed strains, Leu. mesenteroides KCTC 3719 (LM), and Leu. citreum KCTC 3526 (LC) for the comparison, as well as their anti-diabetic, anti-dementia, and antioxidant activities, were evaluated.
2. Materials and Methods
2.1. Raw Materials and Chemicals
The hot-water-extracted MC was provided by Jeju farm Agriculture Association Crop (Jeju, Korea). Briefly, MC was extracted with a MC/water ratio of 1:10 (w/v) for 2 h at 107 °C. After extraction, the extract was concentrated to 33.7° brix by heating at 60 °C. The crude MC extract was stored at –18 °C until use. Leu. mesenteroides MKJW KCTC 14459BP and Leu. mesenteroides MKSR KCTC 18665P were used from our laboratory stocks, and Leu. mesenteroides KCTC 3719 and Leu. citreum KCTC 3526 were distributed from KCTC (Jeongeup-si, Korea). Yeast extract and peptone were purchased from Becton, Dickison and Co. (Bergen County, NJ, USA).
2.2. Preparation of Fermentation
The stock solutions such as MC extract, sucrose, and maltose were separately sterilized at 121 °C and 15 psi for 15 min. Starter cultures such as MKJW, MKSR, LM, and LC were sub-cultured three times for 24 h at 30 °C and centrifuged to obtain the cells before fermentation. MC extract (5.0°brix) was mixed with 394 mmol of sucrose, 143 mmol of maltose in the fermentation medium containing 5 g·L−1 yeast extract, 5 g·L−1 peptone, 20 g·L−1 K2HPO4, 0.2 g·L−1 MgSO4, 0.13 g·L−1 CaCl2·2H2O, 0.01 g·L−1 FeH14O11S, 0.01 g·L−1 MnSO4·H2O, and 0.1 g·L−1 NaCl and fermented by 6.7 log CFU/mL of MKJW, MKSR, LM, and LC at 30 °C for 72 h. CON1 and CON2 were included to compare the fermented MCs. CON1 contains only MC extract, and CON2 contains MC extract solubilized in the fermentation medium same as the fermentation groups. Control groups were carried out without a starter.
2.3. Measurement of Free Sugar, Organic Acid, and Alcohol
A high-performance liquid chromatography system (Dionex Ultimate 3000, Thermo Fisher Scientific, Waltham, MA, USA) was used for the analysis of free sugar, organic acid, and alcohol according to the modified method of Ilaslan et al. [27] and da Costa et al. [28]. In measuring free sugar and mannitol, the sample was homogenized using distilled water and filtered through a 0.45 μm filter, and 10 μL of the sample was injected using a Waters Sugar-Pak (300 × 6.5 mm, 10 μm, Milford, MA, USA). Distilled water was used as a mobile phase, and peaks were detected using shodex RI-101 operated at 70 °C with a flow rate of 0.5 mL/min. When measuring organic acid contents and ethanol, the fermentation broth was homogenized using distilled water and filtered through a 0.45 μm filter, and 10 μL of the sample was injected into an Aminex 87H column (300 × 10 mm, 9 μm, Bio-Rad, Hercules, CA, USA). A total of 0.01 N sulfuric acid was used as the mobile phase, and the peak at 210 nm was detected using a refractive index detector (ERC Refracto MAX520, ERC Inc., Saitama, Japan) operated at 40 °C with a flow rate of 0.5 mL/min.
2.4. Measurement of Dextran Contents
In measuring dextran contents, the methods of Diana. et al. [29] were modified. The fermented MC was centrifuged at 6000 rpm and 4 °C for 10 min to collect the supernatant. The collected supernatant was diluted 1:1 with 99.5% ethanol in a cold state and incubated at 4 °C overnight. Then, the ethanol-diluted solution was centrifuged at 6000 rpm and 4 °C for 20 min to collect dextran. The collected dextran was freeze-dried and weighed, and dextran production was calculated by substituting the following equation.
Dextran contents (%) = {(W1/A) − (W2/A)} ×100
- W1: weight of dextran produced of fermented sample (g);
- W2: weight of dextran produced of non-fermented sample (g);
- A: amount of sucrose added to the medium (g).
2.5. Measurement of Carbohydrate Pattern
Thin-layer chromatography (TLC) was performed as described by Oshima et al. [30] and used for carbohydrate analysis of fermented MC. Samples (1 μL) fermented at 0, 12, and 24 h were loaded onto TLC silica gel (60 f254, Merck) at 1 cm intervals. The plate was developed by using 2:5:1.5 (v/v/v) nitromethane/1-propanol/water, and it was used three times for developing solvent. After development, drying was repeated three times. Then, the TLC plate was visualized by using a methanol solution consisting of 0.3% (w/v) N-(1-naphthyl) ethylenediamine and 5% (v/v) sulfuric acid and dried at 105 °C for 10 min.
2.6. Measurement of Carbohydrate-Hydrolyzing Enzyme Inhibitory Activity
2.6.1. α-Glucosidase Inhibitory Activity
The assay of the AGI activity was performed as described by Chen et al. [31] and Sarmadi et al. [32]. The substrate solution (ρ-nitrophenyl-α-d-glucopyranoside, 2 mM) was prepared in 0.02 M phosphate buffer (pH 6.8, to simulate an intestinal fluid model). A total of 50 μL of the sample was added to 50 μL of 0.02 M sodium phosphate buffer (pH 6.8), 200 μL of 2 mM pNPG (4-nitrophenyl α-D-glucopyranoside) solution, and 50 μL of α-glucosidase enzyme, followed by reaction at 37 °C for 40 min. Then, 1.5 mL of 0.1 M Na2CO3 was added, and absorbance was measured at 405 nm. In blank, distilled water was used, and dispensed instead of the sample and enzyme. It was also used instead of the sample for the control group, and acarbose was used instead of the sample for the positive control. α-Glucosidase inhibition activity was calculated using the following equation:
Inhibition (%) = {1 − (ABSsample − ABSsample blank)/(ABScontrol − ABSblank)} × 100
- ABSsample: absorbance of the experimental sample;
- ABSsample blank: absorbance of the sample blank;
- ABScontrol: absorbance of the control;
- ABSblank: absorbance of the blank.
2.6.2. α-Amylase Inhibitory Activity
α-Amylase inhibitory activity was measured using the method of Rana et al. [33] with slight modifications. After adding 50 µL of a sample to 150 µL of 0.02 M sodium phosphate buffer (pH 7.0), 100 µL of 1% starch solution, and 50 µL of the α-amylase enzyme, the mixture was reacted at 37 °C for 20 min. Then, 2 mL of 3,5-dinitrosalicylic acid reagent (DNS) was added, and the mixture was boiled at 95 °C for 5 min, and then 2 mL of distilled water was added to terminate the reaction. The sample after completing the reaction was calculated by measuring the absorbance at 540 nm and substituting it in the following equation:
Inhibition (%) = {1 − (ABSsample − ABSsample blank)/(ABScontrol − ABSblank)} × 100
2.7. Measurement of Acetylcholinesterase (AChE) Inhibitory Activity and Butyrylcholinesterase (BuChE) Inhibitory Activity
Acetylcholinesterase inhibitory (AChEI) activity and butyrylcholinesterase inhibitory (BuChEI) activity measurements were performed as described by Sharififar et al. [34]. In brief, when measuring the AChE inhibitory activity, 125 μL of the sample, 40 μL of sodium phosphate buffer (pH 7.7) containing 369 mg·L−1 of 5,5′-dithiobis (2-nitrobenzoic acid) (DTNB) and 150 mg·L−1 of sodium bicarbonate, and 5 μL of AChE (Sigma Aldrich, St. Louis, MO, USA) were mixed and left at room temperature for 5 min. Then, 7.5 μL of acetylthiocholine iodide was added. Afterward, absorbance was measured at 412 nm for 15 min. When measuring the BuChE inhibitory activity, BuChE and butyrylthiocholine iodide were used instead of AChE and acetylthiocholine iodide. As a positive control, galantamine was used instead of the sample. Enzymatic activity was calculated by substituting the slope of the absorbance value with time into the following equation.
AChE and BuChE inhibitory activity (%) = {1 − (Ssample/Scontrol)} × 100
- Ssample: Slope of the sample;
- Scontrol: Slope of the control.
2.8. Measurement of SOD-Like Activity
The SOD-like activity assay was measured by modifying the method of Lee and Kim [11]. A total of 100 μL of fivefold dilution sample, 100 μL of 7.2 mM pyrogallol solution, and 1.5 mL of 50 mM Tris-HCl buffer (pH 8.5) were mixed, and the mixture was left for 10 min to react at room temperature. After 10 min, 100 μL of 1 N hydrochloric acid was added to stop the reaction of the sample. As a control, 50 mM Tris-HCl buffer was used instead of the sample, and as a positive control, 1 mg/mL of ascorbic acid was used.
SOD-like activity (%) = {1 − (ABSsample − ABSsample blank)/ABScontrol} × 100
2.9. Correlation Analysis, Principal Component Analysis, and Statistical Analysis
Correlation analysis of various functions of fermented MC was measured using Minitab Pro 16.0. Principal component analysis (PCA) was performed using XLSTAT to preserve the distribution of the original data as much as possible by combining the existing variables and converting the data in the high-dimensional space to the low-dimensional space.
2.10. Statistical Analysis
The experiment was repeated three or more times, and the results were expressed as means ± standard deviation. For statistical analysis of each experimental result, one-way analysis of variance was performed to determine the significance among the experimental groups using Minitab Pro 16.0 (Minitab Inc., State College, PA, USA), and statistically significant differences (p < 0.05) were tested by using Tukey’s test.
3. Results
3.1. Contents of Free Sugar and Metabolites in Fermented MC
The contents of carbohydrates, organic acid, alcohol, and dextran are shown in Table 1. Initially added sugar such as sucrose and maltose was 189.80 mg/mL in CON2, which was non-fermented MC. It was higher than other fermented MCs, indicating that all Leuconostoc strains consumed most sucrose and maltose for their metabolism. All fermented MC produced lactic acid, mannitol, and ethanol, which were not detected in CON1 and CON2. Leu. mesenteroides strains produced higher lactic and acetic acids and mannitol content compared with Leu. citreum (LC) (p < 0.05). The production of lactic (10.42 mg/mL) and acetic (6.94 mg/mL) acids as well as mannitol (34.76 mg/mL) was the highest in MKSR-MC (p < 0.05). MKJW-MC showed the highest dextran content (22.37%), followed by MKSR-MC (18.28%), LM-MC (10.66%), LC-MC (1.99%), CON1 (1.54%), and CON2 (0.47%) at 24 h fermentation (p < 0.05).
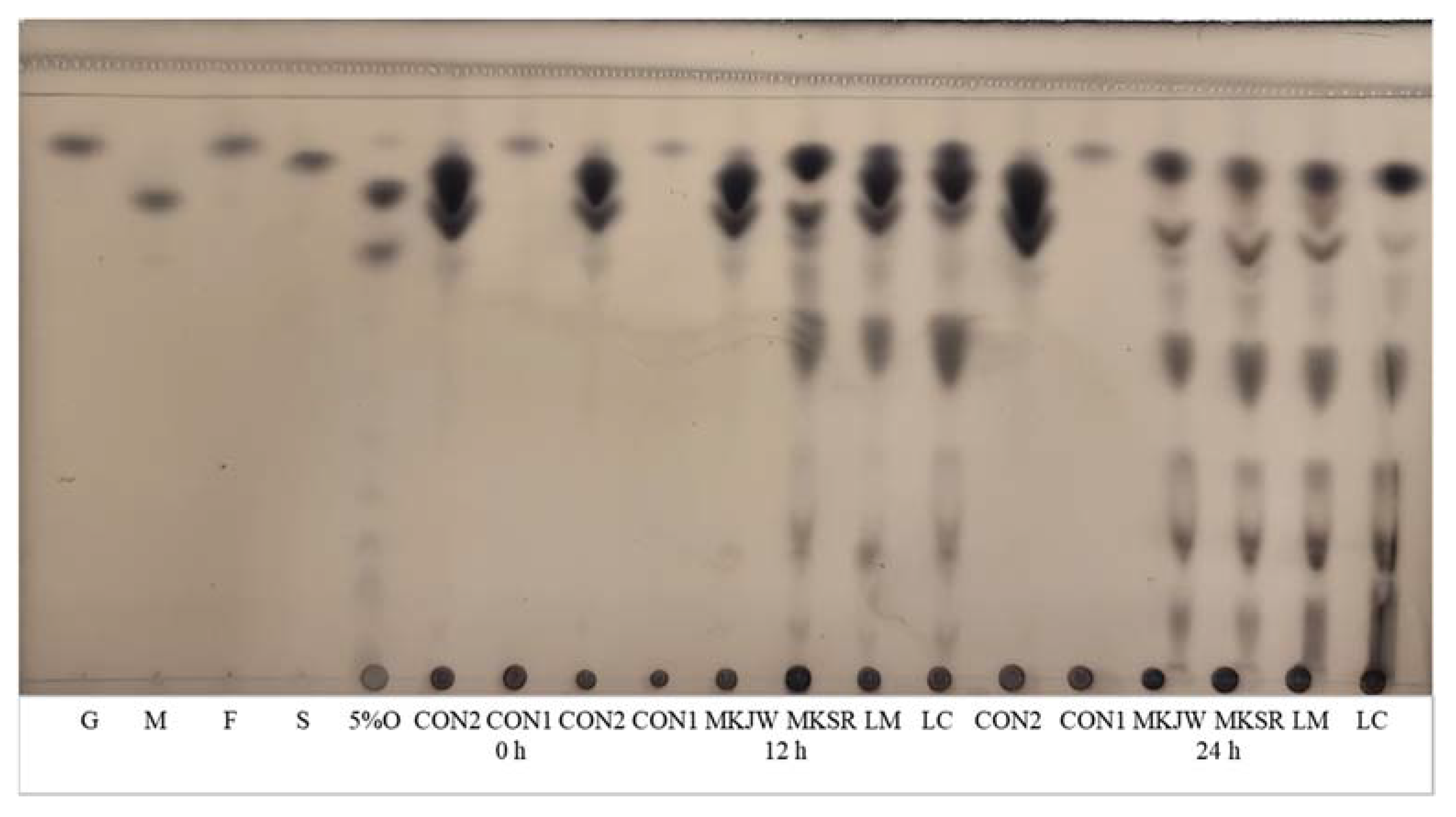
Figure 1 shows the carbohydrate pattern of all samples fermented for 24 h. Several spots showing oligosaccharides and dextran were observed, and the sucrose spot faded in all fermented MC, compared with CON2.
3.2. Carbohydrate-Hydrolyzing Enzyme Inhibitory Activity of Fermented MC
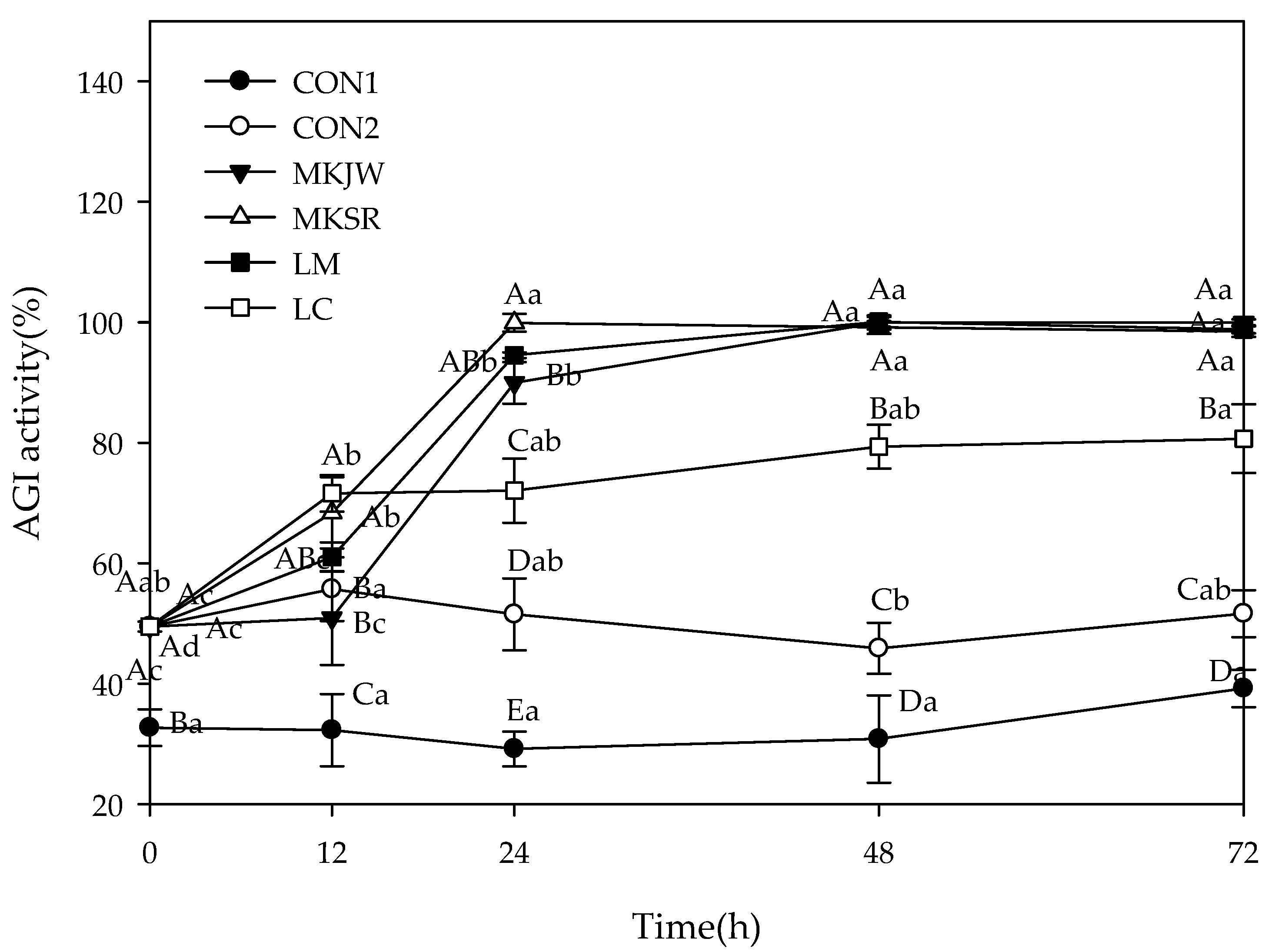
The results of AGI activity are described in Figure 2. The initial inhibition was 32.72% in CON1 and 49.48% in others, which increased with fermentation. After 24 h fermentation, the highest inhibition was shown in MKSR-MC (99.91%), followed by LM-MC (94.55%), MKJW-MC (89.96%), and LC-MC (72.05%). The maximal inhibition was maintained during 72 h fermentation.
Leu. mesenteroides strains showed nearly 100% inhibitory activity as a result of AGI activity after fermentation. By contrast, the fermentation did not change α-amylase inhibition in MKJW-MC and MKSR-MC (Table 2). However, a significant increase in α-amylase inhibition was observed in LM-MC and LC-MC. Our results indicated that MKSR and MKJW selectively inhibited α-glucosidase among carbohydrate-hydrolyzing enzymes.
3.3. Inhibitory Activity of Cholinesterase in MC Fermentation
AChEI and BuChEI of fermented MC are shown in Table 3. Before fermentation, AChEI was 19.40% in fermented groups and CON2, and 28.50% in CON1. After 24 h fermentation, AChEI increased in all fermented groups compared with that before fermentation. In particular, MKSR-MC showed a significantly high inhibitory activity compared with the other groups after fermentation, and Leu. mesenteroides strains showed relatively high AChEI compared with Leu. citreum (p < 0.05). In addition, a slight decrease was observed in CON1 and CON2. Fermentation groups had higher butyrylcholinesterase inhibitory activity than CON1 and CON2 after 24 h fermentation (p < 0.05). Before fermentation, butyrylcholinesterase inhibitory activity was 71.16% in fermented groups and CON2 and 73.74% in CON1. After 24 h fermentation, inhibition was shown in MKSR-MC (88.27%), followed by MKJW-MC (87.90%), LM-MC (87.86%), and LC-MC (85.28%).
3.4. Antioxidant Activity of MC Fermented by Leuconostoc Strains
Table 4 shows SOD-like activity results. The SOD-like activity of all groups except for CON1 before fermentation was 7.66%. However, after 24 h fermentation, the SOD-like activities of MKJW-MC and MKSR-MC were 41.68% and 47.51%, respectively, showing the highest SOD-like activity among all fermentation groups, whereas LM-MC and LC-MC only accounted for 39.79% and 36.81%, respectively. The fermentation group significantly increased the SOD-like activity after 24 h fermentation compared with control groups (p < 0.05).
3.5. Correlation Analysis of Enzyme Inhibitory Activities, Dextran Yield, Free Sugar, Organic Acid, and Antioxidant Activity of Fermented MC by Leuconostoc Strains
Table 5 shows the correlation of all metabolites (mannitol, ethanol, lactic, and acetic acid) and biological activity (AGI, AAI, AChEI, BuChEI, and SOD-like activity). Dextran content was correlated with mannitol (0.819). Lactic acid content was related to the production of mannitol (0.993) and acetic acid (0.997). This result indicated the increase in metabolites by fermentation. AGI activity was highly correlated with the content of dextran (0.917), mannitol (0.956), lactic acid (0.959), and acetic acid (0.959), and it was highly correlated with SOD-like activity (0.976). In addition, the AChEI result correlated with the production of mannitol (0.948), lactic acid (0.923), acetic acid (0.941), and BuChEI (0.895). Therefore, fermentation improved AChEI, and BuChEI similar to AGI.
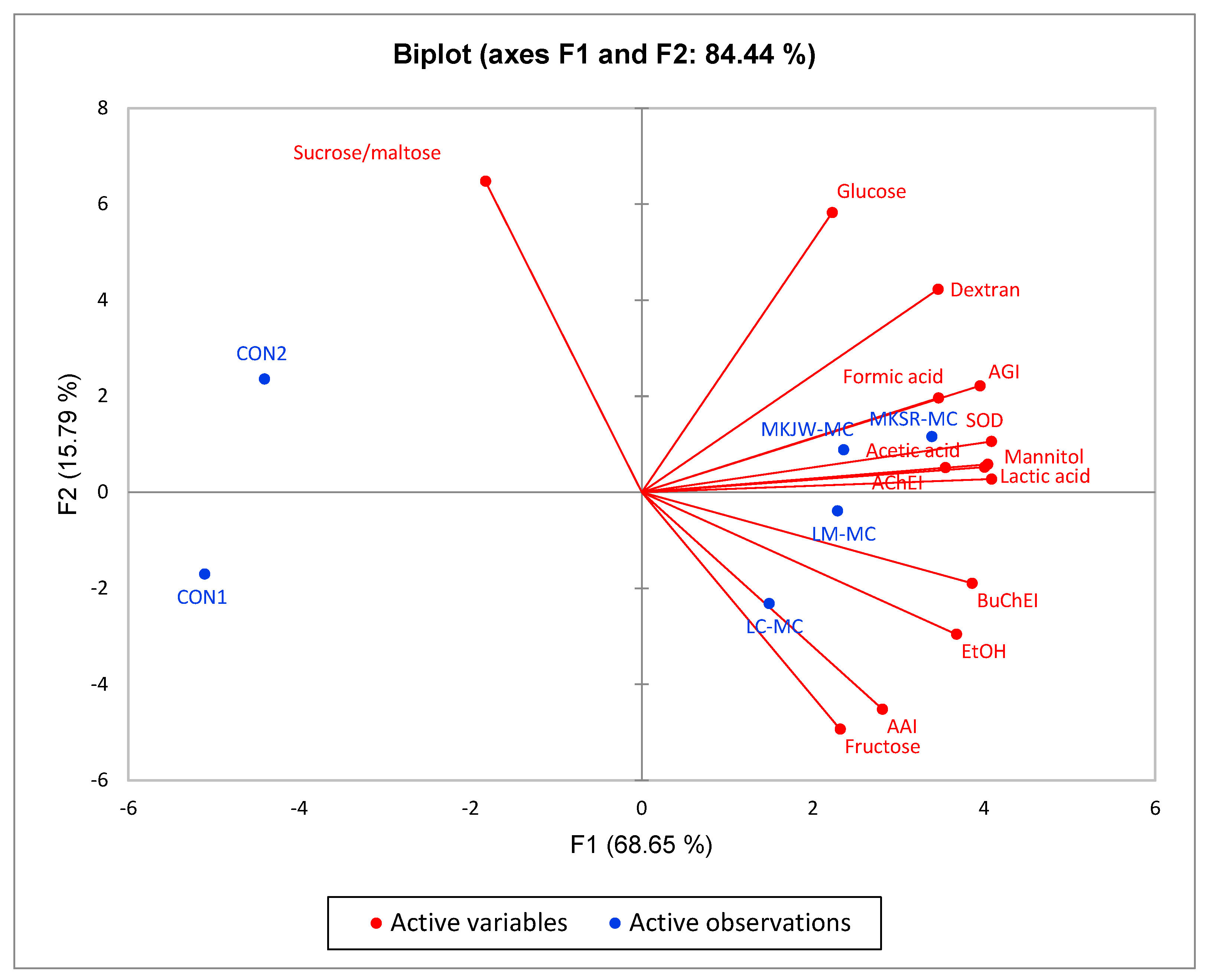
The key characteristics of each sample using all analyzed metabolites and biological functions were performed by PCA. The first principal component (F1) was 68.65%, and the second principal component (F2) was 15.79% (Figure 3). In addition, the total variance of F1 and F2 was 84.44%. Six experimental samples were clustered into four groups. CON1 was on the negative side of F1 and F2. By contrast, CON2 was on the negative side of F1 and positive side of F2. All analyzed values of both groups were not characterized. MKJW-MC and MKSR-MC were grouped on the positive side of F1 and F2. This group was highly correlated with the content of mannitol, acetic acid, lactic acid, and formic acid, as well as with SOD-like activity, AGI, and AChEI. LM-MC and LC-MC were grouped on the positive side of F1 and negative side of F2. This group was correlated with AAI, BuChEI, and ethanol production.
4. Discussion
Lactic acid bacteria (LAB) fermentation is primarily performed to supply beneficial bacteria for intestinal health in dairy products such as cheese or yogurt [35]. However, dairy products are sometimes avoided by humans who are suffering from lactose intolerance and hypercholesterolemia, as well as those who are vegetarian [36]. At present, LAB fermentation of plant materials has emerged as a good alternative for ingesting probiotic bacteria, improving taste and aroma of food, and effectively utilizing various physiologically active substances from plants [37].
MKJW and MKSR strains have been previously reported to possess probiotic, AGI, cholinesterase inhibitory (ChEI), antioxidant, and antibacterial activities [10,11]. Moreover, Yoon and Kim [14] verified that both strains were effective probiotic starters to ferment Artemisia capillaris, possessing multi-biological activities such as anti-diabetic, anticholinesterase inhibitory, and antibacterial activities.
In addition to the unique compounds of MC, fermented MC contained lactic and acetic acids, mannitol, ethanol, and dextran, which were typical metabolites of heterofermentative Leuconostoc spp., although the degree of production varies (Table 1). In particular, dextran was synthesized when Leuconostoc strains, possessing active dextransucrase, consumed sucrose. A series of oligosaccharides were also produced because both maltose and sucrose were present in the fermentation medium (Figure 1). When Leuconostoc spp. are grown on sucrose rich media, the production of dextransucrase is induced. Dextransucrase uses sucrose as a substrate to produce dextran and fructose. However, when they are grown on sucrose and maltose media, dextransucrase catalyzes the transfer of glucosyl residues from sucrose to maltose to produce a series of isomalto-oligosaccharides as well as the synthesis of dextran [11,14].
Bacterial metabolites such as lactic and acetic acids, mannitol, and dextran were highly correlated to α-glucosidase inhibition, but they were less associated with α-amylase (Table 5). The inhibition of α-glucosidase activities in the reaction solution might occur through competitive or non-competitive inhibition, where lactic and acetic acid, mannitol, or dextran might interfere with the active site of α-glucosidase or alter its structure. Changes in pH and ionic strength caused by lactic acid and acetic acid could also affect activities of the enzyme. Mannitol and dextran, being larger molecules, might hinder proper enzyme–substrate interactions. AGI was T2DM could be controlled by diet change. Controlling carbohydrate-hydrolyzing enzymes is important to regulate blood glucose levels in patients with diabetes. In particular, the selective inhibition of α-glucosidase among carbohydrate-hydrolyzing enzymes is essential to control blood glucose levels in patients with T2DM. The strong inhibition of α-glucosidase with low inhibitory α-amylase activity in MKSR-MC and MKJW-MC is effective for patients with T2DM because excessive inhibition of α-amylase inhibits the breakdown of carbohydrates into disaccharides, which can lead to digestive disorders such as abdominal distension, flatulence, and diarrhea [38].
AGI was closely associated with acetylcholinesterase inhibition (0.847) and butyrylcholinesterase inhibition (0.84). Cholinesterase inhibition was strongly correlated to bacterial metabolites such as mannitol and lactic and acetic acid (>0.92). The enzymatic inhibition could be due to competitive inhibition by lactic acid, acetic acid, or mannitol, which interfere with the enzyme’s active site. It is thought that the pH changes and alterations in ionic strength due to lactic and acetic acid also contribute to the inhibition, but when only lactic and acetic acid at the same concentration as the fermented MC were present, the inhibitory effect was not found. Further experimentation is necessary to ascertain the exact mechanism. ChEI is a potential strategy for the treatment of AD [39]. AD is a degenerative disease that affects cognitive functions, such as memory loss, speech loss, and judgment loss, because of the loss of cholinergic cells [34] and the presence of Aβ peptides (Aβ) and neurofibrillary tangles [40]. The loss of cholinergic cells leads to the loss of acetylcholine, a neurotransmitter [34]. In addition, acetylcholinesterase is an enzyme that hydrolyzes acetylcholine into acetyl and choline. When the activity of acetylcholinesterase increases, acetylcholine is hydrolyzed, and its levels are decreased, which contributes to cognitive impairment [41]. In treating these cognitive impairments, the amount of acetylcholine present in synapses between cholinergic neurons should be increased using cholinesterase inhibitors [36]. Enzymes that hydrolyze cholinergic neurotransmitters include acetylcholinesterase and butyrylcholinesterase [42]. Galantamine, donepezil, and rivastigmine have been used for the treatment of AD despite their reported side effects such as weight loss and anorexia [40]. Water-extracted MC showed 28.50% for AChEI and 73.74% for BuChEI (Table 3). Ethanol-extracted MCs contain AChEI (71–81%) [36,43]. Our fermentation increased the content of AChEI and BuChEI, indicating that fermented MC could be used as potential anti-dementia products. The highest AChEI was observed in MKSR-MC, but no significant difference in BuChEI was found among fermented MCs.
Various extracts from MC have been reported to show antioxidant activity [44]. Our water-extracted MC (CON1) did not show SOD-like activity (−18.40%), but fermentation increased SOD-like activity. SOD is an essential enzyme to defend cells against the toxic activity of oxygen free radicals. As shown in Table 5, SOD-like activity was highly correlated to fermentation metabolites such as mannitol (0.957), lactic acid (0.975), and acetic acid (0.968), as well as AGI. It has been reported that the fermentation of lactic acid bacteria (LAB) can contribute to SOD activity, but the degree of difference significantly varied depending on the type of LAB [45]. Additionally, the content of phenolic compounds and flavonoids that affect SOD-like activity may have increased after the fermentation. SOD-like activity was the highest in MKSR-MC, producing higher mannitol, lactic acid, and acetic acid among controls and fermentation groups. The higher activity was followed by MKJW-MC, LM-MC, and LC-MC.
Finally, six experimental samples were classified into four groups (Figure 3). In particular, MKSR-MC and MKJW-MC showed strong association with AGI, AChEI, and SOD-like activities.
5. Conclusions
The MC fermented by Leu. mesenteroides MKSR has superior bioactive properties due to strong α-glucosidase inhibition without affecting α-amylase, AChEI, and SOD-like activity compared to controls and other fermented MCs. It was highly related to metabolites such as mannitol, lactic acid, and dextran. The outcome indicates that utilizing MKSR fermentation of MC can be a favorable approach for controlling diabetes and other complicated diseases. Our findings provide a basis for further research and development of health functional products. However, it is crucial to emphasize that additional research, such as animal experiments and clinical trials, is warranted to validate and consolidate these findings.
Author Contributions
Conceptualization, S.Y., Y.J. and M.K.; methodology, J.K. and S.Y.; visualization, J.K. and S.Y.; formal analysis, data curation, and writing—original draft, J.K., S.Y. and M.K.; writing—review and editing, Y.J. and M.K.; project administration, Y.J. and M.K.; supervision, Y.J. and M.K.; funding acquisition, S.Y. and M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Small and Medium-sized Enterprises and Startups of Korea, S3093086, and by the National Research Foundation of Korea, 2020R1I1A3072127.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
This article is a part of Jiwoo Kim’s Master’s Thesis completed at the Department of Food Science and Nutrition, Dankook University, Korea, under the supervision of Yoonhwa Jeong.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization. Dementia. Available online: https://www.who.int/news-room/fact-sheets/detail/dementia (accessed on 15 March 2023).
- Goyal, R.; Jialal, I. Diabetes mellitus type 2. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Ahtiluoto, S.; Polvikoski, T.; Peltonen, M.; Solomon, A.; Tuomilehto, J.; Winblad, B.; Sulkava, R.; Kivipelto, M. Diabetes, Alzheimer disease, and vascular dementia: A population-based neuropathologic study. Neurology 2010, 75, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Feil, D.G.; Zhu, C.W.; Sultzer, D.L. The relationship between cognitive impairment and diabetes self-management in a population-based community sample of older adults with Type 2 diabetes. J. Behav. Med. 2012, 35, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, R.A.; Gunderson, E.P.; Quesenberry, C.P.; Zhou, J.; Yaffe, K. Body mass index in midlife and risk of Alzheimer disease and vascular dementia. Curr. Alzheimer Res. 2007, 4, 103–109. [Google Scholar] [CrossRef]
- Reddy, V.P.; Zhu, X.; Perry, G.; Smith, M.A. Oxidative stress in diabetes and Alzheimer’s disease. J. Alzheimer’s Dis. 2009, 16, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Rivest, S. Alzheimer’s disease: Microglia targets and their modulation to promote amyloid phagocytosis and mitigate neuroinflammation. Expert Opin. Ther. Targets 2020, 24, 331–344. [Google Scholar] [CrossRef]
- Leblhuber, F.; Steiner, K.; Schuetz, B.; Fuchs, D.; Gostner, J.M. Probiotic supplementation in patients with Alzheimer’s dementia-an explorative intervention study. Curr. Alzheimer Res. 2018, 15, 1106–1113. [Google Scholar] [CrossRef]
- Firouzi, S.; Majid, H.A.; Ismail, A.; Kamaruddin, N.A.; Barakatun-Nisak, M. Effect of multi-strain probiotics (multi-strain microbial cell preparation) on glycemic control and other diabetes-related outcomes in people with type 2 diabetes: A randomized controlled trial. Eur. J. Nutr. 2017, 56, 1535–1550. [Google Scholar] [CrossRef]
- Kim, M.; Yoon, J.; Seong, H.; Jeong, Y. Novel Leconostoc mesemteroides MKJW, and uses thereof. 10-2021-0014624, 2 February 2021. [Google Scholar]
- Lee, S.; Kim, M. Leuconostoc mesenteroides MKSR isolated from kimchi possesses α-glucosidase inhibitory activity, antioxidant activity, and cholesterol-lowering effects. LWT 2019, 116, 108570. [Google Scholar] [CrossRef]
- Campanella, D.; Rizzello, C.G.; Fasciano, C.; Gambacorta, G.; Pinto, D.; Marzani, B.; Scarano, N.; De Angelis, M.; Gobbetti, M. Exploitation of grape marc as functional substrate for lactic acid bacteria and bifidobacteria growth and enhanced antioxidant activity. Food Microbiol. 2017, 65, 25–35. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Yoon, J.; Kim, M. In vitro evaluation of antidiabetic, antidementia, and antioxidant activity of Artemisia capillaris fermented by Leuconostoc spp. LWT 2022, 172, 114163. [Google Scholar] [CrossRef]
- Oh, Y.J.; Kim, T.S.; Moon, H.W.; Lee, S.Y.; Lee, S.Y.; Ji, G.E.; Hwang, K.T. Lactobacillus plantarum PMO 08 as a probiotic starter culture for plant-based fermented beverages. Molecules 2020, 25, 5056. [Google Scholar] [CrossRef] [PubMed]
- Chavan, M.; Gat, Y.; Harmalkar, M.; Waghmare, R. Development of non-dairy fermented probiotic drink based on germinated and ungerminated cereals and legume. LWT 2018, 91, 339–344. [Google Scholar] [CrossRef]
- Demarinis, C.; Verni, M.; Pinto, L.; Rizzello, C.G.; Baruzzi, F. Use of Selected Lactic Acid Bacteria for the Fermentation of Legume-Based Water Extracts. Foods 2022, 11, 3346. [Google Scholar] [CrossRef]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 110, 881–890. [Google Scholar] [CrossRef]
- Yeo, Y.L.; Chia, Y.Y.; Lee, C.H.; Sow, H.S.; Yap, W.S. Effectiveness of Maceration Periods with Different Extraction Solvents on in-vitro Antimicrobial Activity from Fruit of Momordica charantia L. J. Appl. Pharm. Sci. 2014, 4, 016–023. [Google Scholar] [CrossRef]
- Bortolotti, M.; Mercatelli, D.; Polito, L. Momordica charantia, a nutraceutical approach for inflammatory related diseases. Front. Pharmacol. 2019, 10, 486. [Google Scholar] [CrossRef]
- Cho, I.S.; Yoon, H.Y.; Hong, K.W. Quality Characteristics of Teriyaki Sauce added with Bitter Melon (Momoridaica charantia L.) Powder. Culin. Sci. Hosp. Res. 2017, 23, 176–182. [Google Scholar]
- Desai, S.; Tatke, P. Charantin: An important lead compound from Momordica charantia for the treatment of diabetes. J. Pharmacogn. Phytochem. 2015, 3, 163–166. [Google Scholar]
- Shu, C.H.; Jaiswal, R.; Peng, Y.Y.; Liu, T.H. Improving bioactivities of Momordica charantia broth through fermentation using mixed cultures of Lactobacillus plantarum, Gluconacetobacter sp. and Saccharomyces cerevisiae. Process Biochem. 2022, 117, 142–152. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.J.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Nie, S.P.; Xiong, T.; Xie, M.Y. Momordica charantia juice with Lactobacillus plantarum fermentation: Chemical composition, antioxidant properties and aroma profile. Food Biosci. 2019, 29, 62–72. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.J.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Nie, S.P.; Xiong, T.; Xie, M.Y. Fermented Momordica charantia L. juice modulates hyperglycemia, lipid profile, and gut microbiota in type 2 diabetic rats. Food Res. Int. 2019, 121, 367–378. [Google Scholar] [CrossRef]
- Wen, J.J.; Li, M.Z.; Gao, H.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Zhang, Y.L.; Xie, M.Y.; Nie, S.P. Polysaccharides from fermented Momordica charantia L. with Lactobacillus plantarum NCU116 ameliorate metabolic disorders and gut microbiota change in obese rats. Food Funct. 2021, 12, 2617–2630. [Google Scholar] [CrossRef]
- Ilaslan, K.; Boyaci, I.H.; Topcu, A. Rapid analysis of glucose, fructose and sucrose contents of commercial soft drinks using Raman spectroscopy. Food Control 2015, 48, 56–61. [Google Scholar] [CrossRef]
- da Costa, M.P.; da Silva Frasao, B.; da Costa Lima, B.R.C.; Rodrigues, B.L.; Junior, C.A.C. Simultaneous analysis of carbohydrates and organic acids by HPLC-DAD-RI for monitoring goat’s milk yogurts fermentation. Talanta 2016, 152, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Diana, C.R.; Humberto, H.S.; Jorge, Y.F. Structural characterization and rheological properties of dextran produced by native strains isolated of Agave salmiana. Food Hydrocoll. 2019, 90, 1–8. [Google Scholar] [CrossRef]
- Oshima, N.; Saito, M.; Niino, M.; Hiraishi, Y.; Ueki, K.; Okoshi, K.; Hakamatsuka, T.; Hada, N. Elucidation of Chemical Interactions between Crude Drugs Using Quantitative Thin-Layer Chromatography Analysis. Molecules 2022, 27, 593. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Screening for potential new probiotic based on probiotic properties and α-glucosidase inhibitory activity. Food Control 2014, 35, 65–72. [Google Scholar] [CrossRef]
- Sarmadi, B.; Aminuddin, F.; Hamid, M.; Saari, N.; Abdul-Hamid, A.; Ismail, A. Hypoglycemic effects of cocoa (Theobroma cacao L.) autolysates. Food Chem. 2012, 134, 905–911. [Google Scholar] [CrossRef]
- Rana, Z.H.; Alam, M.K.; Akhtaruzzaman, M. Nutritional composition, total phenolic content, antioxidant and α-amylase inhibitory activities of different fractions of selected wild edible plants. Antioxidants 2019, 8, 203. [Google Scholar] [CrossRef]
- Sharififar, F.; Moshafi, M.H.; Shafazand, E.; Koohpayeh, A. Acetyl cholinesterase inhibitory, antioxidant and cytotoxic activity of three dietary medicinal plants. Food Chem. 2012, 130, 20–23. [Google Scholar] [CrossRef]
- Kandylis, P.; Pissaridi, K.; Bekatorou, A.; Kanellaki, M.; Koutinas, A.A. Dairy and non-dairy probiotic beverages. Curr. Opin. Food Sci. 2016, 7, 58–63. [Google Scholar] [CrossRef]
- Kumar, B.; Tharumasivam, S.V.; Boominathan, V.; Perumal, E.; Dhandapani, P.; Kaliyaperumal, K.; Arumugam, S.; Subramanian, K.; Renuga, P.S.; Shakthivel, V.; et al. A pilot study on nanotherapy of Momordica charantia against trimethyltin chloride-induced neurotoxicity in Danio rerio (zebrafish). J. Nanomater. 2021, 2021, 2180638. [Google Scholar] [CrossRef]
- Rice, T.; Sahin, A.W.; Heitmann, M.; Lynch, K.M.; Jacob, F.; Arendt, E.K.; Coffey, A. Application of mannitol producing Leuconostoc citreum TR116 to reduce sugar content of barley, oat and wheat malt-based worts. Food Microbiol. 2020, 90, 103464. [Google Scholar] [CrossRef]
- Paschou, S.A.; Papadopoulou-Marketou, N.; Chrousos, G.P.; Kanaka-Gantenbein, C. On type 1 diabetes mellitus pathogenesis. Endocr. Connect. 2018, 7, R38–R46. [Google Scholar] [CrossRef]
- Apostolidis, E.; Lee, C. In vitro potential of Ascophyllum nodosum phenolic antioxidant-mediated α-glucosidase and α-amylase inhibition. J. Food Sci. 2010, 75, H97–H102. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G. The biological activities of butyrylcholinesterase inhibitors. Biomed. Pharmacother. 2022, 146, 112556. [Google Scholar] [CrossRef]
- Ahmed, S.; Khan, S.T.; Zargaham, M.K.; Khan, A.U.; Khan, S.; Hussain, A.; Uddin, J.; Khan, A.; Al-Harrasi, A. Potential therapeutic natural products against Alzheimer’s disease with Reference of Acetylcholinesterase. Biomed. Pharmacother. 2021, 139, 111609. [Google Scholar] [CrossRef]
- Adewusi, E.A.; Moodley, N.; Steenkamp, V. Antioxidant and acetylcholinesterase inhibitory activity of selected southern African medicinal plants. S. Afr. J. Bot. 2011, 77, 638–644. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Kucukaydin, S.; Yeskaliyeva, B.; Ozturk, M.; Dinica, R.M. Non-alkaloid cholinesterase inhibitory compounds from natural sources. Molecules 2021, 26, 5582. [Google Scholar] [CrossRef]
- Nagarani, G.; Abirami, A.; Siddhuraju, P. A comparative study on antioxidant potentials, inhibitory activities against key enzymes related to metabolic syndrome, and anti-inflammatory activity of leaf extract from different Momordica species. Food Sci. Hum. Wellness 2014, 3, 36–46. [Google Scholar] [CrossRef]
- Perumal, V.; Khatib, A.; Ahmed, Q.U.; Uzir, B.F.; Abas, F.; Murugesu, S.; Saiman, M.Z.; Primaharinastiti, R.; El-Seedi, H. Correlation of the GC-MS-based metabolite profile of Momordica charantia fruit and its antioxidant activity. Int. Food Res. J. 2022, 29, 58–66. [Google Scholar] [CrossRef]
Figure 1.
TLC analysis of carbohydrates in fermented MC by Leuconostoc strains. G, 1% glucose; M, 1% maltose; F, 1% fructose; S, 1% sucrose; 5%O, 5% oligosaccharides; CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526.
Figure 1.
TLC analysis of carbohydrates in fermented MC by Leuconostoc strains. G, 1% glucose; M, 1% maltose; F, 1% fructose; S, 1% sucrose; 5%O, 5% oligosaccharides; CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526.

Figure 2.
Change of α-glucosidase inhibitory activity during 72 h fermentation. AGI, α-glucosidase inhibition; CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526. Different capital letters of the labels on the curve are significantly different according to the fermentation time within the same group (p < 0.05). Different capital letters are significantly different among the groups, and different small letters are significantly different according to the fermentation time (p < 0.05).
Figure 2.
Change of α-glucosidase inhibitory activity during 72 h fermentation. AGI, α-glucosidase inhibition; CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526. Different capital letters of the labels on the curve are significantly different according to the fermentation time within the same group (p < 0.05). Different capital letters are significantly different among the groups, and different small letters are significantly different according to the fermentation time (p < 0.05).
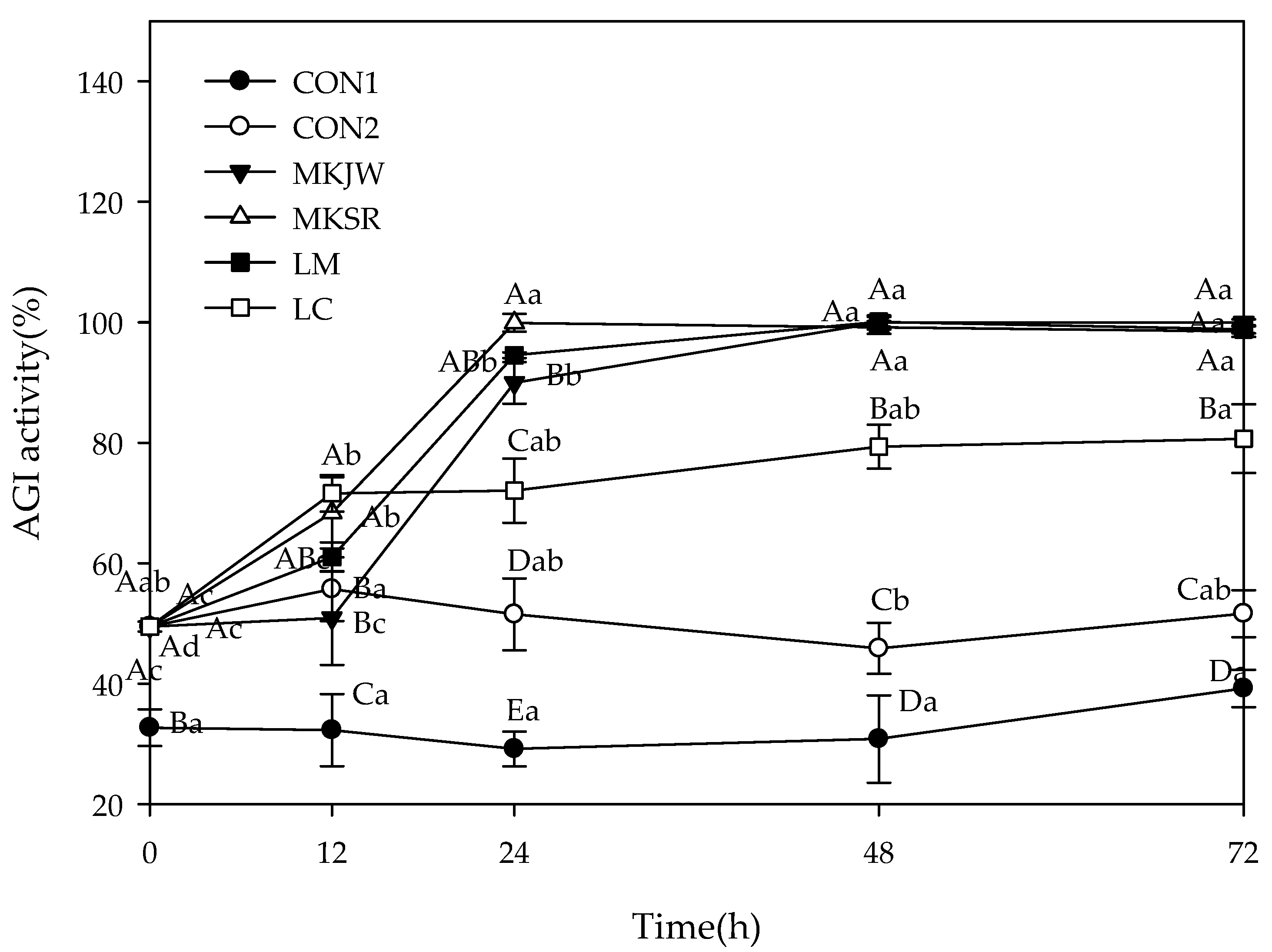
Figure 3.
Principal component analysis of metabolites and physiological properties of non-fermented and fermented MC. CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526; AGI, α-glucosidase inhibitory activity; AAI, α-amylase inhibitory activity; SOD, SOD-like activity; AChEI, acetylcholinesterase inhibitory activity; BuChEI, butyrylcholinesterase inhibitory activity.
Figure 3.
Principal component analysis of metabolites and physiological properties of non-fermented and fermented MC. CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526; AGI, α-glucosidase inhibitory activity; AAI, α-amylase inhibitory activity; SOD, SOD-like activity; AChEI, acetylcholinesterase inhibitory activity; BuChEI, butyrylcholinesterase inhibitory activity.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Free sugar, organic acid, alcohol, and dextran content of non-fermented and fermented MC.
| Free Sugar (mg/mL) | Organic Acid (mg/mL) | Alcohol (mg/mL) | Dextran (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sucrose or Maltose | Glucose | Fructose | Lactic Acid | Formic Acid | Acetic Acid | Mannitol | Ethanol | 0 h | 24 h | |
| CON1 (1) | 0 | 3.54 ± 0.68 BC | 2.28 ± 0.39 C | 0 | 0.11 ± 0.04 A | 0.11 ± 0.00 D | 0 | 0 | 0 | 1.54 ± 1.84 C |
| CON2 | 189.80 ± 22.08 A(2)(3) | 9.33 ± 0.99 B | 3.43 ± 0.48 C | 0 | 0.15 ± 0.08 A | 0.18 ± 0.02 D | 0 | 0 | 0 | 0.47 ± 0.50 C |
| MKJW-MC | 22.29 ± 3.02 BCD | 21.75 ± 3.11 A | 19.77 ± 1.72 B | 8.27 ± 0.06 B | 0.18 ± 0.08 A | 5.03 ± 0.19 B | 23.63 ± 3.05 B | 0.67 ± 0.04 B | 0 | 22.37 ± 0.77 A |
| MKSR-MC | 30.73 ± 7.09 BC | 21.81 ± 4.97 A | 8.86 ± 0.65 C | 10.42 ± 0.20 A | 0.18 ± 0.08 A | 6.94 ± 0.17 A | 34.76 ± 5.99 A | 0.68 ± 0.06 B | 0 | 18.28 ± 3.54 A |
| LM-MC | 32.81 ± 9.16 B | 8.74 ± 3.35 B | 26.85 ± 3.26 B | 8.55 ± 0.14 B | 0.17 ± 0.08 A | 5.38 ± 0.22 B | 27.77 ± 4.69 AB | 0.72 ± 0.08 B | 0 | 10.66 ± 1.56 B |
| LC-MC | 4.25 ± 0.71 CD | 0.51 ± 0.23 C | 48.43 ± 6.38 A | 6.43 ± 0.03 C | 0.16 ± 0.13 A | 3.85 ± 0.10 C | 18.74 ± 3.02 B | 1.00 ± 0.11 A | 0 | 1.99 ± 1.46 C |
(1) CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526. (2) Different capital letters within a column are significantly different (p < 0.05). (3) All values are mean ± SD (n = 3).
Table 2.
α-Amylase inhibitory activity of non-fermented and fermented MC (%).
| 0 h | 24 h | |
|---|---|---|
| CON1 (1) | 10.20 ± 1.22 Bb | 14.72 ± 1.66 Ac |
| CON2 | 20.14 ± 3.07 Aa | 15.26 ± 1.17 Ac |
| MKJW-MC | 20.14 ± 3.07 Aa | 22.82 ± 0.86 Abc |
| MKSR-MC | 20.14 ± 3.07 Aa | 24.43 ± 3.25 Abc |
| LM-MC | 20.14 ± 3.07 Ba | 29.93 ± 5.18 Aab |
| LC-MC | 20.14 ± 3.07 Ba | 36.61 ± 6.93 Aa |
All values are mean ± SD (n = 3). Different capital letters within a row and different small letters within a column are significantly different (p < 0.05). (1) CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526.
Table 3.
Acetylcholinesterase (AChE) inhibitory activity and butyrylcholinesterase (BuChE) inhibitory activity of non-fermented and fermented MC.
Table 3.
Acetylcholinesterase (AChE) inhibitory activity and butyrylcholinesterase (BuChE) inhibitory activity of non-fermented and fermented MC.
| AChE Inhibition (%) | BuChE Inhibition (%) | |||
|---|---|---|---|---|
| 0 h | 24 h | 0 h | 24 h | |
| CON1 (1) | 28.50 ± 3.47 Aa | 26.68 ± 2.29 Ac | 73.74 ± 0.95 Ba | 79.37 ± 4.23 Ab |
| CON2 | 19.40 ± 3.23 Ab | 17.79 ± 2.00 Ac | 71.16 ± 3.76 Aa | 74.10 ± 0.95 Ab |
| MKJW-MC | 19.40 ± 3.23 Bb | 40.59 ± 6.82 Ab | 71.16 ± 3.76 Ba | 87.90 ± 0.66 Aa |
| MKSR-MC | 19.40 ± 3.23 Bb | 55.24 ± 7.31 Aa | 71.16 ± 3.76 Ba | 88.27 ± 3.19 Aa |
| LM-MC | 19.40 ± 3.23 Bb | 44.46 ± 7.24 Aab | 71.16 ± 3.76 Ba | 87.86 ± 1.65 Aa |
| LC-MC | 19.40 ± 3.23 Bb | 31.21 ± 7.94 Abc | 71.16 ± 3.76 Ba | 85.28 ± 1.33 Aa |
| Galantamine (2) | 96.80 ± 0.59 | 96.84 ± 0.71 | ||
All values are mean ± SD (n = 4). Different capital letters within a row and different small letters within a column are significantly different (p < 0.05). (1) CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526. (2) Galantamine was used as the positive control at the concentration of 0.01 mg/mL for AChE inhibitory activity and 0.1 mg/mL for BuChE inhibitory activity.
Table 4.
Superoxide-dismutase-like activity of non-fermented and fermented MC.
| 0 h | 24 h | |
|---|---|---|
| CON1 (1) | −18.40 ± 2.20 Bb | 3.78 ± 2.73 Ad |
| CON2 | 7.66 ± 3.64 Aa | 14.46 ± 3.28 Ac |
| MKJW-MC | 7.66 ± 3.64 Ba | 41.68 ± 3.01 Aab |
| MKSR-MC | 7.66 ± 3.64 Ba | 47.51 ± 4.61 Aa |
| LM-MC | 7.66 ± 3.64 Ba | 39.79 ± 0.70 Aab |
| LC-MC | 7.66 ± 3.64 Ba | 36.81 ± 6.14 Ab |
All values are mean ± SD (n = 3). Different capital letters within a row and different small letters within a column are significantly different (p < 0.05). (1) CON1, MC extract; CON2, MC supplemented medium; MKJW-MC, fermented MC extract by Leu. mesenteroides MKJW; MKSR-MC, fermented MC extract by Leu. mesenteroides MKSR; LM-MC, fermented MC extract by Leu. mesenteroides KCTC 3719; LC-MC, fermented MC extract by Leu. citreum KCTC 3526.
Table 5.
Correlation between metabolites and physiological properties in non- and fermented MC.
| Dextran | Sucrose and Maltose | Glucose | Fructose | Mannitol | EtOH | Lactic Acid | Formic Acid | Acetic Acid | AGI | AAI | AChEI | BuChEI | SOD | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dextran | 1 | |||||||||||||
| Sucrose and maltose | −0.065 | 1 | ||||||||||||
| Glucose | 0.849 * | 0.026 | 1 | |||||||||||
| Fructose | 0.176 | −0.425 | −0.341 | 1 | ||||||||||
| Mannitol | 0.819 * | −0.432 | 0.609 | 0.396 | 1 | |||||||||
| EtOH (1) | 0.565 | −0.533 | 0.132 | 0.863 * | 0.787 | 1 | ||||||||
| Lactic acid | 0.841 * | −0.461 | 0.601 | 0.458 | 0.993 *** | 0.832 * | 1 | |||||||
| Formic acid | 0.795 | 0.01 | 0.4 | 0.627 | 0.736 | 0.815 * | 0.774 | 1 | ||||||
| Acetic acid | 0.84 * | −0.438 | 0.628 | 0.407 | 0.998 *** | 0.8 | 0.997 *** | 0.756 | 1 | |||||
| AGI | 0.917 * | −0.207 | 0.661 | 0.395 | 0.956 ** | 0.756 | 0.959 ** | 0.858 * | 0.959 ** | 1 | ||||
| AAI | 0.257 | −0.455 | −0.23 | 0.962 ** | 0.560 | 0.929 ** | 0.6 | 0.692 | 0.564 | 0.524 | 1 | |||
| AChEI | 0.734 | −0.514 | 0.68 | 0.155 | 0.948 ** | 0.604 | 0.923 ** | 0.498 | 0.941 ** | 0.847 * | 0.342 | 1 | ||
| BuChEI | 0.711 | −0.703 | 0.476 | 0.512 | 0.927 ** | 0.83 * | 0.947 ** | 0.609 | 0.932 ** | 0.84 * | 0.613 | 0.895 * | 1 | |
| SOD | 0.881 * | −0.314 | 0.587 | 0.521 | 0.957 ** | 0.861 * | 0.975 ** | 0.893 * | 0.968 ** | 0.976 ** | 0.646 | 0.828 | 0.876 * | 1 |
(1) EtOH, ethanol; AGI, α-glucosidase inhibition; AAI, α-amylase inhibition; AChEI, acetylcholinesterase inhibition; BuChEI, butyrylcholinesterase inhibition; SOD, superoxide dismutase-like activity. * Correlation is significantly different at p < 0.05. ** Correlation is significantly different at p < 0.01. *** Correlation is significantly different at p < 0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, J.; Yu, S.; Jeong, Y.; Kim, M. Enhancement of Bioactive Properties in Momordica charantia by Leuconostoc Fermentation. Fermentation 2023, 9, 523. https://doi.org/10.3390/fermentation9060523
AMA Style
Kim J, Yu S, Jeong Y, Kim M. Enhancement of Bioactive Properties in Momordica charantia by Leuconostoc Fermentation. Fermentation. 2023; 9(6):523. https://doi.org/10.3390/fermentation9060523
Chicago/Turabian StyleKim, Jiwoo, Sungryul Yu, Yoonhwa Jeong, and Misook Kim. 2023. "Enhancement of Bioactive Properties in Momordica charantia by Leuconostoc Fermentation" Fermentation 9, no. 6: 523. https://doi.org/10.3390/fermentation9060523
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.