
Screening of Ultraviolet-Induced Thermotolerant Yeast Mutants and Their Performance
1
State Key Laboratory of Pollution Control and Resource Reuse, College of Environmental Science and Engineering, Tongji University, Shanghai 200092, China
2
School of Environmental Science and Engineering, Shanghai Jiao Tong University, Shanghai 200240, China
3
College of Life and Environmental Science, Wenzhou University, Wenzhou 325035, China
4
Institute for Eco-Environmental Research of Sanyang Wetland, Wenzhou University, Wenzhou 325035, China
*
Author to whom correspondence should be addressed.
Fermentation 2023, 9(7), 608; https://doi.org/10.3390/fermentation9070608
Submission received: 16 May 2023
/
Revised: 19 June 2023
/
Accepted: 26 June 2023
/
Published: 28 June 2023
(This article belongs to the Topic Valorizing Waste through Thermal and Biological Processes for Sustainable Energy Production)
Abstract
:The simultaneous saccharification and fermentation (SSF) technique holds promise for the conversion of lignocellulose to ethanol. However, the optimal fermentation temperature of yeast is lower than the enzymatic hydrolysis temperature of the saccharification process, which leads to the temperature of the actual production process of SSF usually being lower than 38 °C. In this work, two ultraviolet (UV)-induced mutations were performed step by step using Saccharomyces cerevisiae BY4742 as the original strain to enable the yeast to perform well at higher temperatures. Thermotolerant strains obtained through mutagenesis and screening, YUV1-1 and YUV2-2, were utilized for fermentation and SSF at a targeted temperature of 40 °C. They obtained ethanol yields comparable to those at 38 °C in SSF, whereas the ethanol yields of the original strain at 40 °C decreased by about 10% compared to those at 38 °C. This study proves that thermotolerant strains adapted to elevated fermentation and SSF temperatures can be obtained through UV mutagenesis and screening, thereby increasing the stability of the fermentation and SSF processes and lowering the subsequent distillation costs.
1. Introduction
The world community largely recognizes the significance of energy security. Due to the decrease in traditional fossil energy sources, such as coal, oil, and natural gas, on which we are now largely dependent, and the carbon dioxide emissions from traditional fossil fuel use, it is vital and urgent to build a renewable energy supply system. In the past two decades, researchers have exerted considerable effort in the field of renewable energy. Biomass energy is an important renewable energy source and one of the research hotspots with non-toxic, biodegradable, and carbon-neutral characteristics [1]. By 2050, it is anticipated that 50% of the world’s energy consumption will be derived from biomass [2].
Lignocellulosic ethanol, a well-known second-generation biomass energy, is attracting increasing public attention [3,4,5,6,7]. In comparison to first-generation bioethanol, which utilizes corn or other food grains as raw materials, second-generation bioethanol technology based on agricultural residues such as wheat straw offers better environmental benefits with fewer greenhouse gas emissions. Moreover, using lignocellulose-rich crop straw to produce ethanol will reduce the environmental pollution caused by the improper treatment of straw such as direct incineration and discarding [8]. Substituting food crops with abundant lignocellulosic materials, which alleviates concerns about the impending food crisis [9,10,11,12,13], makes lignocellulosic ethanol a promising alternative to conventional fossil fuels [14,15,16,17]. It may potentially decrease our reliance on fossil fuels and address the global energy crisis.
The simultaneous saccharification and fermentation (SSF) technique for converting lignocellulose to ethanol would abate widespread microbial contamination (due to the presence of ethanol in the culture broth), reduce osmotic stress because glucose is fermented to ethanol instead of being allowed to accumulate, lessen the reducing sugar’s inhibition on the enzyme, minimize the potential loss of sugar, and shorten the entire processing cycle [10,18,19]. Despite the advantages of SSF, there exists a critical problem: the most suitable temperature for cellulase differs from that of fermentative microorganisms [20]. The optimal temperature for yeast fermentation is usually 28–35 °C, but 40–50 °C for enzymatic hydrolysis [9,21]. Reluctantly, SSF is usually carried out at 30–38 °C as a compromise [22,23,24,25,26,27]. It is difficult to reduce the optimum reaction temperature of cellulase through protein engineering. Therefore, the key to solving this problem is to elevate the fermentation temperature by obtaining thermotolerant ethanol-producing yeast. Furthermore, higher temperatures would help improve productivity while reducing cooling-related costs, contamination chances, and subsequent distillation costs during SSF, as well as the general fermentation process [28]. Given the magnitude of the worldwide bioethanol industry, even a minor improvement in ethanol production efficiency through the adoption of thermotolerant yeasts could have a significant economic impact [29].
Although research on non-conventional heat-resistant yeast is an idea to improve the temperature of SSF, certain non-traditional yeasts demonstrate poorer ethanol yields or recovery viability when compared to conventional Saccharomyces cerevisiae (S. cerevisiae) [30]. S. cerevisiae is widely utilized in the bioethanol sector and can readily be engineered to produce a variety of additional important products [12]. Therefore, from an industrialization standpoint, it is valuable to try to improve the heat resistance of conventional yeast. Currently, mutation breeding is frequently utilized to alter strains to satisfy production requirements. With the development of an increasing number of innovative mutation breeding techniques, such as high-energy particle flow, heavy-ion irradiation, atmospheric and room-temperature plasma breeding, and other techniques, it is possible to select thermotolerant yeast strains precisely and efficiently [30,31,32,33]. Among these, ultraviolet (UV)-induced mutation is a sensible and straightforward method for altering specific microbe traits. The simplicity of the equipment and the lack of extra chemical additives distinguish UV mutagenesis from other cutting-edge target-strain creation procedures. In comparison to adaptive evolution, which might take several months with equivalent benefits, UV mutagenesis and screening take substantially less time and can be finished in a few days [34]. UV radiation causes the pyrimidines of deoxyribose nucleic acid (DNA) molecules to polymerize into pyrimidine dimers, thereby affecting the replication and transcription of DNA, resulting in the reversion, frameshift, transversion, or deletion of gene sequence [35]. Screening and selection after UV mutagenesis treatment have traditionally been used to obtain the preferred yeast strain. Related studies have demonstrated the feasibility and effectiveness of enhancing yeast’s tolerance to certain restrictive circumstances. After UV radiation and massive screening, Sridhar et al. obtained the yeast mutants VS1 and VS3 strains, whose ethanol yields were 10.7% and 1.6% greater than those of the parent strain at 42 °C using 150 g/L glucose as the substrate [29]. However, additional research is needed to determine whether UV mutagenesis might help common S. cerevisiae perform better in SSF with more complicated environments and higher temperatures.
This research aims to obtain thermotolerant yeast through UV mutagenesis and screening using S. cerevisiae, the yeast most commonly used and industrially suitable for sustainable fuel and chemical production, as the original strain [36], to mitigate conflicts between the optimal temperature of the enzyme and fermentation strain, which is inconsistent in SSF. The mutants were evaluated in a sequencing batch reactor for glucose fermentation and straw SSF at 40 °C, which is within cellulase’s ideal working temperature range, to verify whether the mutants can achieve outstanding ethanol output at an elevated temperature.
2. Materials and Methods
2.1. Materials
2.1.1. Strain
Saccharomyces cerevisiae BY4742 (S. cerevisiae BY4742) was the original strain for mutagenesis. The pre-culture was conducted in a biochemical incubator (LRH-250, BLUE PARD, Shanghai, China) with a rocking device inside (WSZ-100A, BLUE PARD, Shanghai, China) at 37 °C and 180 rpm for 16 h and then used to inoculate 500 mL baffled shake-flasks containing 250 mL of the yeast extract peptone dextrose (YPD) medium with the same conditions to obtain sufficient yeast cells for the subsequent experiments. After recovery and cultivation, the yeast suspension was diluted to approximately 50 g/L with sterilized water.
2.1.2. Media
The yeast was recovered and pre-cultured in a YPD medium, which contained yeast extract (10 g/L), glucose (20 g/L), and peptone (20 g/L). The solid culture medium used for plate screening contained 10 g/L of agar in addition to the YPD medium. Before use, each culture medium was maintained at a pH between 4.0 and 4.5 and sterilized for 25 min at 121 °C. The components of the fermentation medium are detailed in Table S1.
2.2. Ultraviolet Mutagenesis
About 10 mL of a yeast suspension at a concentration of 50 g/L was stirred with a magnetic stirrer (MS5, LAB FISH, Huzhou, China) in a petri dish and exposed to a 15 W UV light at a distance of 35 cm for 2 min. Samples were withdrawn every 10 s and the cells’ survival rate was assessed using methylene blue to determine the exposure time of the cells to UV. All the operations were conducted on a super-clean worktable (JJ-CJ-1F, JINJING, Suzhou, China).
2.3. Survival Screening and Separation of the Thermotolerant Yeast
Spread plates were used to screen mutants that can grow well at elevated temperatures. The yeast suspension was coated in a standard solid YPD medium and cultivated for 5 d in a biochemical incubator (LRH-250, BLUE PARD, Shanghai, China). The culture temperatures of the first mutagenesis and second mutagenesis were 42 °C and 43.5 °C, respectively. The dominant independent colonies were chosen and cultured for subsequent fermentation screening. Purification took another 5 d at 42 °C or 43.5 °C in the biochemical incubator. All the plates were kept away from light during the culture process to prevent the genetic materials of the mutants from being repaired under illumination. For subsequent experiments, the yeast was grown in a YPD medium at 37 °C for 16–18 h, with shaking at 180 rpm until a sufficient amount of yeast cells were obtained for the fermentation assays.
2.4. Fermentation and SSF Process
For the fermentation, 2 g/L of yeast was added to the fermentation medium in a 500 mL automatic reactor (BIOTECH-8, BAOXING, Shanghai, China) containing 300 mL of fermentation reaction liquor. The process was conducted at a shaking speed of 180 rpm, pH of 4.0, and temperature of 40 °C or 42 °C. The fermentation duration was 60 h overall, with samples obtained from the reactor every 12 h and frozen at 4 °C. To describe the mutants’ thermotolerance properties in comparison to the original strain, cell growth, glucose intake, and ethanol production were examined. SSF was carried out in 100 mL DURAN glass bottles (SCHOTT, Mainz, Germany) containing a total liquid volume of 30 mL. Bottles were agitated at 180 rpm in a shaking incubator with a constant temperature. The substrate used for the SSF process was wheat straw that had been prepared with 1.0% NaOH, dried at 105 °C, crushed, and sieved through 80 mesh in accordance with Zhang’s procedures [9]. The initial solid content (dry weight of pretreated wheat straw) was set at 16.0% for all samples, and the pH was controlled with 50 mM of citrate buffer at 4.0. The temperature was set to 38 °C or 40 °C. The cellulase (purchased from Sigma-Aldrich, St. Louis, MO, USA) from Trichoderma reesei ATCC 26921 was a mixture of cellobiohydrolases, endoglucanases, and β-1,4 glucosidases, and its filter-paper activity (FPA) was 82.22 FPU/mL. The enzyme loading amount was determined based on our previous optimization experiments, set at 30 FPU/g substrate, which can ensure efficient substrate hydrolysis without causing enzyme waste [9]. The initial yeast concentration was set at 8.0 g/L. The total SSF time was 120 h, and samples were drawn from the reactor every 24 h and decanted into centrifugal tubs, then centrifuged at 4000 rpm for 10 min. The supernatant was filtered through 0.45 μm membrane filters and stored at 4 °C in 2 mL bottles. The analyses of the reducing sugars and ethanol concentrations were conducted to confirm the SSF effects of the strains.
2.5. Analytical Methods
The optical density (OD) at 600 nm was measured using a spectrophotometer (UV2600, SHUNYUHENGPING, Shanghai, China) to determine yeast growth. An inverted microscope (X71, OYMPUS, Kyoto, Japan) was used to examine cell morphology. The reducing sugars and ethanol concentrations of fermentation and SSF were evaluated using high-performance liquid chromatography (HPLC) (LC-10AD, SHIMADZU, Kyoto, Japan) equipped with a refractive index detector (RID-10A, SHIMADZU, Kyoto, Japan) according to Zhang and Pessani’s protocol [9,37].
2.6. Calculation
According to the stoichiometric relationship described by Equation (1), i.e., 100 g of glucose can create 51.1 g of ethanol, the maximum theoretical ethanol concentration from glucose was computed. The ethanol yield based on the total starting glucose concentration was determined according to Equation (2) as follows:
where stands for the actual ethanol yield (g/g, %), Cc for the concentration of glucose (g/L), and CE for the ethanol concentration (g/L).
C6H12O6→2CH3CH2OH+2CO2
Since, theoretically, 100 g of cellulose could produce 56.8 g of ethanol, the ethanol yield from the wheat straw’s initial cellulose was estimated using Equation (3)
where is the ethanol yield (g/g, %), is the initial dry wheat straw concentration (g/L), 0.568 is the transformation coefficient of ethanol per gram of cellulose, CE is the ethanol concentration of the SSF liquid (g/L), and PC is the percentage of cellulose in the initial wheat straw.
3. Results and Discussion
3.1. Ultraviolet-Induced Mutagenesis and Screening
As shown in Figure 1, UV mutagenesis and screening were gradually carried out twice. UV radiation was applied to the parent strains, and survival screening was carried out at specific elevated temperatures. The strains that passed the survival screening were selected for fermentation experiments, and the desired strains produced the best level of fermentation.
3.1.1. Determination of Mutagenesis Time
UV mutagenesis was performed twice in this study. The parent strains of the first and second mutagenesis processes were S. cerevisiae BY4742 and the first mutated strain with the best fermentation performance at 42 °C, respectively. The lethality rates, judged by the proportion of cells stained with methylene blue (an example of the staining picture is shown in Figure S1), of yeasts exposed to radiation during the two mutagenesis processes were basically the same. UV radiation lethality rates increased with exposure time. After 50 s of radiation, the lethality rates reached approximately 80%, and after 70 s, the lethality rates were 100% (Figure S2). When the lethality rate reaches 70–80%, the chance of mutation is relatively high. Therefore, 50 s was chosen as the UV mutagenesis period in this experiment, and the yeast suspension exposed to UV light for 50 s was employed for further screening.
3.1.2. Survival Screening of Thermotolerant Strains
In light of the benefits of higher operating temperatures, the objective of this study was to increase the suitable temperature for fermentation and the SSF process to 40 °C by enhancing yeast’s heat tolerance. In order to adapt the strains to higher temperatures, the survival screening temperatures were set at 42 °C and 43.5 °C in the first and second UV-induced mutagenesis processes, respectively, which is higher than the optimal temperature range of the original yeast strain S. cerevisiae BY4742 (below 37.5 °C) and the target temperature (40 °C).
After receiving 50 s of UV radiation, the S. cerevisiae BY4742 yeast solution was diluted 100 times and grown for 5 days at 42 °C in the first UV-induced mutagenesis process. Yeast colonies with a smooth surface, white hue, and clear boundary were obtained. It was demonstrated that mutant yeasts can thrive and reproduce effectively at a temperature of 42 °C. Five thermotolerant strains, designated YUV1-1, YUV1-2, YUV1-3, YUV1-4, and YUV1-5, were finally developed by streak-culturing colonies showing dominant growth at 42 °C. The parent strain of the second mutagenesis was YUV1-1, which obtained the highest ethanol yield in the subsequent fermentation screening. The second mutagenesis process followed the same procedure as the first, but the survival screening temperature was increased to 43.5 °C to help the strain better adapt to a higher temperature. As a result, five strains were obtained and designated YUV2-1, YUV2-2, YUV2-3, YUV2-4, and YUV2-5.
3.1.3. Fermentation Screening of Thermotolerant Strains
The first mutants, YUV1-1, YUV1-2, YUV1-3, YUV1-4, and YUV1-5, and the second mutants, YUV2-1, YUV2-2, YUV2-3, YUV2-4, and YUV2-5, as well as their parent strains, were subjected to more severe temperature fermentation at 42 °C to assess their fermentation performance. After diluting the NaOH pretreatment, the proportions of cellulose, hemicellulose, and lignin in our SSF substrate (wheat straw) were 58.8%, 22.9%, and 11.8%, respectively. As cellulose is essentially a large polysaccharide composed of a large number of glucose monomers, and our pretreated wheat straw contained more than half cellulose, it can be presumed that glucose would account for a large proportion of the SSF hydrolysis products. Our previous research on the optimization of enzymatic saccharification of wheat straw pre-treated with sodium hydroxide also supported this finding [38]. In order to better align with the SSF process, glucose was selected as the substrate for fermentation screening. The substrate concentration was set at 80 g/L, which, according to our previous research, would not inhibit the growth of the original strain S. cerevisiae BY4742 [39]. Variations in biomass, glucose consumption, and ethanol output during fermentation can indicate the yeast’s growth and fermentation activity.
For the first fermentation screening, the initial yeast concentration was 2.0 g/L. As depicted in Figure 2a, YUV1-1, YUV1-2, YUV1-3, and YUV1-4 grew well in the process of fermentation at 42 °C and achieved their maximum concentrations of 3.1, 2.5, 3.3, and 2.8 g/L after 60 h, 12 h, 12 h, and 36 h, respectively. In addition, the concentrations of YUV1-5 and the original strain S. cerevisiae BY4742 remained below 2.0 g/L throughout the fermentation process. YUV1-5 grew well on the YPD culture medium but could not replicate massively in a more complex fermentation environment. Except for heat tolerance, UV-induced mutagenesis may have altered other features of YUV1-5, resulting in its distinct developmental state during the fermentation process.
As depicted in Figure 2b, during the ongoing fermentation process, the glucose utilization rate of each strain exhibited a steady upward trend. YUV1-1 and YUV1-4 exhibited excellent glucose consumption, and the glucose utilization rate of these two mutated strains exceeded 60% at the end of the fermentation process. YUV1-2 and the original strain S. cerevisiae BY4742 had a low glucose utilization rate below 30% at the end of the fermentation process at 42 °C. YUV1-5 had a higher glucose consumption rate compared to S. cerevisiae BY4742 while having a similar cell concentration. However, YUV1-3, which had a substantially higher cell concentration compared to YUV1-5, did not have a significantly higher glucose utilization rate compared to YUV1-5. This demonstrates that the mutated strain’s cell concentration and glucose utilization rate did not always correspond positively. The reason for this could be that the various mutant strains have varied metabolic pathways and substrate assimilation abilities. There are precedents for mutagenesis altering yeast substrate utilization and product formation. Yi et al. obtained yeast mutants with the ability to ferment xylose by UV and γ-ray mutagenesis of S. cerevisiae CICC-1517RM [40]. Yao et al. created a superior isoprene synthase mutant ISPSLN through saturation mutagenesis, leading to an almost fourfold improvement in isoprene production [41]. In comparison, YUV1-3 exhibited stronger assimilation and YUV1-5 showed stronger alienation after the first mutagenesis in this study.
The steady accumulation of ethanol during fermentation is shown in Figure 2c. After 36 h of fermentation, the ethanol yield for the original strain of S. cerevisiae BY4742 stabilized, and the ultimate ethanol yield was under 20%. Other than YUV1-4, which reached its maximum ethanol yield of 31.9% after 48 h, all other mutant strains reached their maximum ethanol yields after 60 h of fermentation, with YUV1-1 having the highest ethanol yield at 42.6%. It is worth noting that YUV1-4 exhibited the highest glucose utilization rate among the five mutated strains, as shown in Figure 2b. However, its ethanol yield was not the highest and began to decrease after 48 h, and its cell concentration steadily declined after 36 h. The following reason may account for this phenomenon: UV increased the cell’s heat resistance while decreasing its tolerance to ethanol. As a result, the cell concentration started to decline, and the metabolic mode of living cells changed after the accumulation of ethanol to a certain concentration at the later stage of fermentation [36]. Cells growing on glucose initially switched from a fermentative metabolism that mostly relied on glycolysis to produce ethanol to a respiratory metabolism that utilized the tricarboxylic acid, glyoxylate, and mitochondrial electron transport chains to break down the ethanol produced during the earlier stages of growth [42]. All mutated strains produced more ethanol than the original strain at the end of fermentation, proving that the mutants were verified by fermentation at a higher temperature. However, their ethanol yield was still less than 45%.
To summarize, YUV1-1 maintained a high biomass throughout the fermentation process, particularly in the late fermentation stage, where its biomass and ethanol yield were the highest, showing that its heat resistance and ethanol tolerance were both good. Therefore, the original strain selected for the second UV mutagenesis was YUV1-1. The glucose utilization rate and ethanol yield of YUV1-2 were both poor, being only slightly higher than those of the original strain. Although YUV1-3 had a relatively high biomass, it did not stand out among the other four mutants in terms of its ability to use glucose or produce ethanol. The biomass of YUV1-5, however, was lower than 2 g/L and comparable to that of the original strain, whereas its ethanol output was higher than that of YUV1-3. The ethanol output, biomass, and glucose utilization rate of YUV1-4 were all quite high in the first 48 h of fermentation. However, because YUV1-4 had a low ethanol tolerance, ethanol inhibition during the late fermentation period caused a drop in the biomass and ethanol output. As can be seen, despite the strains after one mutation passing the high-temperature survival screening, they did not all have the same levels of stability in production due to variances in substrate tolerance, product tolerance, and substrate metabolic route.
The first mutagenesis yielded five strains that enhanced ethanol yield in the higher-temperature fermentation screening test. However, the ultimate ethanol yield was less than 45%, which is not ideal. So, YUV2-1, YUV2-2, YUV2-3, YUV2-4, and YUV2-5 were produced through secondary mutagenesis. Cell growth, glucose uptake, and ethanol production were still used to gauge their performance during the 42 °C fermentation process.
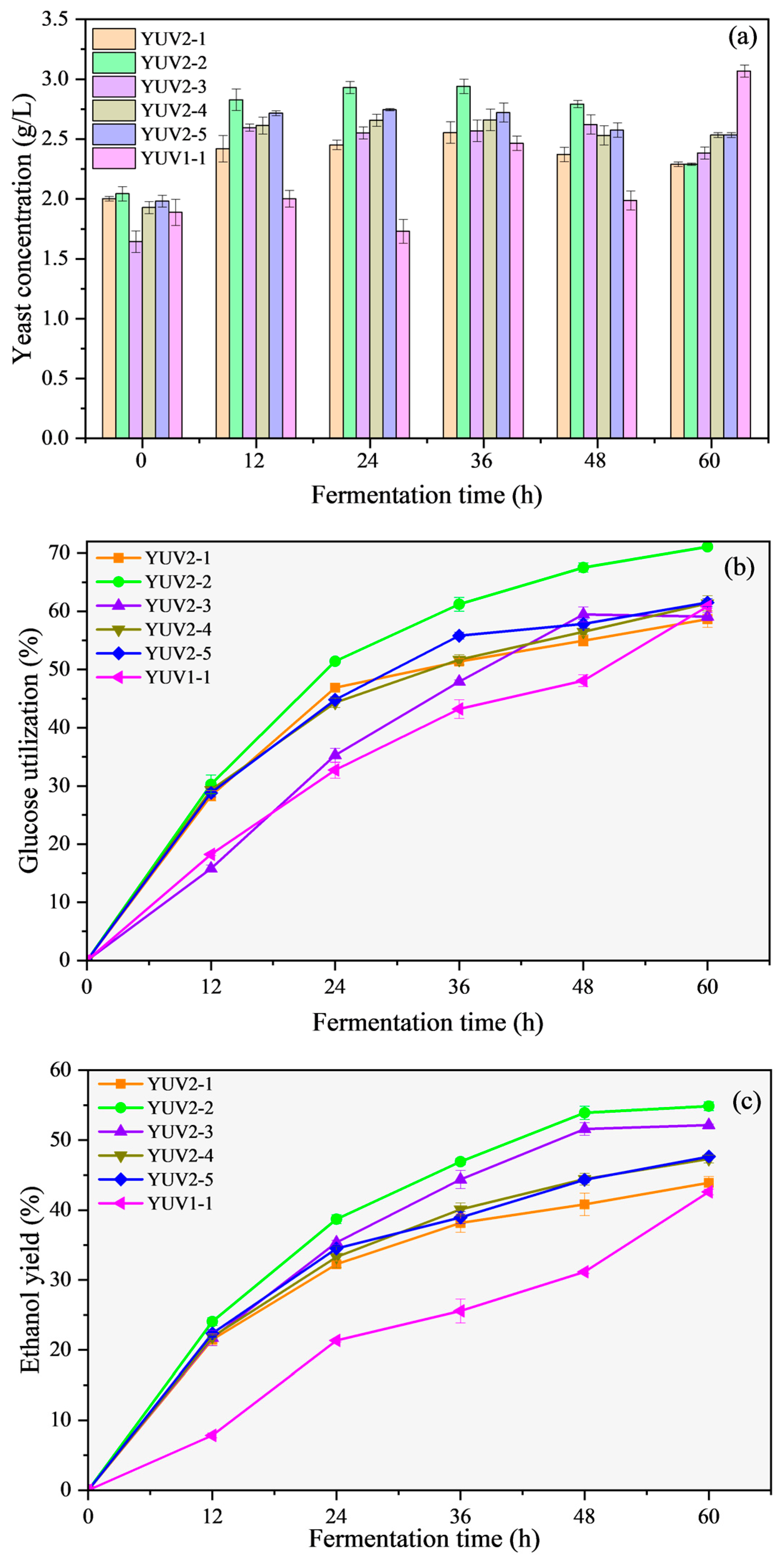
Figure 3a shows the concentration changes of the five strains obtained through the second mutagenesis process, as well as their parent strain YUV1-1 during fermentation at 42 °C. In the fermentation process, there was no significant difference in the cell concentrations among the five mutated strains. The cell concentrations of the second-mutated strains reached about 2.5 g/L after 12 h of fermentation, then remained relatively stable and only decreased slightly with substrate consumption and ethanol accumulation in the later period of fermentation, which was better than the general level of the first mutated strains. Among them, YUV2-2 has the highest biomass and maintained a cell concentration of about 3 g/L between 12–48 h of fermentation. The consumption of cells as a carbon source might account for the drop in cell concentrations in the later period of fermentation. In general, the second mutants screened at 43.5 °C had greater and more stable biomass compared to the first mutants screened at 42 °C during the fermentation phase.
Figure 3b and Figure 4c compare the glucose utilization rates and ethanol yields of the second-mutated strains, as well as their parent strain YUV1-1. Although the glucose intakes and ethanol yields of the five second-mutated strains varied slightly, they were all noticeably superior to their parent strain YUV1-1. YUV2-2 showed a high glucose utilization rate of more than 70%. In contrast to the original strain S. cerevisiae BY4742, whose ethanol yield was steady after 36 h (Figure 2c), the ethanol yields of the six mutant strains grew steadily throughout the 60 h fermentation. The original strain was increasingly difficult to maintain at a higher temperature; thus, the mutant strains that had survived higher-temperature survival screening had more obvious advantages in the later stages of higher-temperature fermentation screening. In addition to YUV2-1, the final ethanol yields of the other four strains exceeded 45%, of which YUV2-2 and YUV2-3 exceeded 50% (54.87% and 52.15%, respectively).
In addition, at 35 °C, neither a glucose concentration of 80 g/L nor an exogenous ethanol addition of 10 g/L hindered yeast growth in our prior work on substrate and product inhibition in ethanol fermentation [39]. In this study, when fermentation was carried out at 42 °C for 12 h, although the quantities of glucose and ethanol in the fermentation broth were lower than the inhibitory values, the parent strain showed a reduction in concentration, whereas the majority of the mutagenic strains showed an increase. This demonstrated that the screened strains’ improved fermentation performance was mainly due to their improved heat resistance rather than their improved ethanol or glucose tolerance.
The second-round mutated strains with YUV1-1, which had already exhibited certain heat tolerance, as the parent and survived screening at 43.5 °C, showed higher cell viability and more stable fermentation activity at 42 °C. This shows that step-by-step mutagenesis and the use of appropriate stringent screening conditions can aid in acquiring better and more stable performance when obtaining specific traits of S. cerevisiae through UV-induced mutagenesis.
3.2. Fermentation Performance of Mutated Strains
To compare their performance to that of the common strain S. cerevisiae BY4742, YUV1-1 obtained in the first mutagenesis and YUV2-2 obtained in the second mutagenesis, which had the highest ethanol yield in fermentation screening, were utilized for fermentation using glucose as the substrate at a target temperature of 40 °C. The whole fermentation process lasted 60 h. The final fermentation data, including the yeast cell and ethanol concentrations, calculated ethanol yields, and ethanol volumetric productivity, are presented in Table 1.
The initial cell concentrations were set at about 2 g/L. Figure 4a shows that at the initial stage of fermentation, the cell concentrations of the three strains, YUV1-1, YUV2-2, and S. cerevisiae BY4742, increased rapidly and reached the maximum values of 3.27, 3.41, and 3.07 g/L at 24 h, followed by a slight decrease. The cell concentrations finally maintained at about 3 g/L, which were all higher than those of the strains in the fermentation environment at 42 °C, indicating that the mutagenic strains could function at 42 °C but their activity was higher at a lower temperature. The consumption of the glucose substrate, whose concentrations below 40 g/L were insufficient to support a significant amount of yeast growth, may be the cause of the decline in cell concentrations in the late stage of the process. The yeast lived in a challenging environment with few accessible reducing sugars and a high concentration of ethanol during the late stages of fermentation. Cells entered the decay phase and used polysaccharides, lipids, poly-β-hydroxybutyrate, and other self-stored energy substances for endogenous respiratory metabolism to survive or potentially initiate cellular autolysis. The concentration of YUV2-2 was slightly higher than that of YUV1-1, and both mutants had higher concentrations compared to the original strain S. cerevisiae BY4742. Compared to the cell concentrations in the fermentation environment at 40 °C, the cell concentration of the original strain at 42 °C decreased by 1.62 g/L, whereas the cell concentrations of YUV1-1 and YUV2-2 at 42 °C decreased by 0.27 and 0.47 g/L, respectively. So, the mutated strains were more adaptable to a higher temperature compared to the original strain. Compared to the original strain, the mutant strains showed more obvious growth advantages at higher temperatures.
As shown in Figure 4b and Figure 5c, throughout the fermentation process, the yield of ethanol gradually increased with the consumption of the glucose substrate. For the three strains tested, most of the fermentation took place within the first 36 h, after which the fermentation speed slowed down considerably with the decrease in the glucose concentrations. In the last 48 h, the ethanol yield of the original strain S. cerevisiae BY4742 was stable, whereas the ethanol yields of the two mutated strains slowly increased. The ethanol yields of the two mutants were 55.84% and 57.09%, which were, respectively, 12.41% and 13.66% higher than that of the original strain. At the same time, the fermentation performance of the two mutated strains at 40 °C was better than that of their performance at 42 °C, with final ethanol yields of 42.60% and 54.87%, respectively. In addition, the comprehensive results of the fermentation experiments at 40 °C and the fermentation screening experiments at 42 °C mentioned above showed that the heat resistance of YUV2-2, which underwent the second mutagenesis and higher-temperature survival screening, was better than that of its parent strain YUV1-1.
3.3. Performance of Mutated Strains in Simultaneous Saccharification and Fermentation
The mutated strains exhibited a better fermentation effect than S. cerevisiae BY4742 during the glucose fermentation process at 40 °C. To further verify whether they could align perfectly with cellulase during the SSF process, YUV1-1, YUV2-2, and the original strain S. cerevisiae BY4742 as the control, were utilized in the SSF experiments for 120 h at a target temperature of 40 °C and a routine temperature of 38 °C. Wheat straw rich in cellulose content was used as the substrate for SSF after alkali pretreatment. The concentrations of cellulose hydrolysates, which consisted of two kinds of hexose (glucose, and cellobiose) that can be used by yeasts, and a kind of pentose (arabinose) that can scarcely be used by common yeasts, as well as ethanol, were detected every 24 h. The final ethanol concentrations, yields, and volumetric productivity after 120 h of SSF are listed in Table 1.
Figure 5 depicts the ethanol formation profiles and changes in carbohydrates during the SSF process using the two mutated strains compared to the original strain S. cerevisiae BY4742, at 40 °C and 38 °C. In all cases, more than half the fermentation was completed in the first 24 h, and then the ethanol yield increased slowly, reaching a stable level after 96 h and a maximum after 120 h. After 24 h, the concentration of hexose (glucose and cellobiose) decreased gradually, whereas the concentration of pentose (arabinose) remained relatively stable. During SSF at 38 °C, 2.6 to 3.1 g/L of xylose was detected and completely consumed during the later stages of fermentation. This may be due to the lack of easily available hexose in the later stages of fermentation, as the yeast needed to utilize a small amount of xylose. In the SSF at 40 °C, a concentration of 5.6 to 6.1 g/L of xylose was detected, which was higher than that at 38 °C. Furthermore, there was no significant decrease in the xylose concentration, indicating that higher temperatures did indeed increase the saccharification level of lignocellulose. Whether in ethanol production or reducing sugar consumption, the performance of the two mutants at 40 °C was slightly better than that at 38 °C, but the overall difference was not significant. At 38 °C, at the end of 120 h of fermentation, the ethanol yields of YUV1-1, YUV2-2, and S. cerevisiae BY4742 were very similar at 74.25%, 75.62%, and 71.73%, respectively, whereas the corresponding values at 40 °C were 77.30%, 78.01%, and 61.75%. At 40 °C, the ethanol yields of the two mutants were slightly higher than those at 38 °C. Regarding S. cerevisiae BY4742, the consumption of glucose and production of ethanol were slower at 40 °C, and the final ethanol yield at 40 °C was about 10% lower than that at 38 °C. The activity of S. cerevisiae BY4742 at 40 °C was lower than that at 38 °C.
For the two mutants YUV1-1 and YUV2-2, which successfully adapted to a temperature of 40 °C, the concentration of glucose and cellobiose that could be used decreased rapidly with the reaction, and the utilization rates of hexose by the two strains at 40 °C were slightly higher than that at 38 °C. At 40 °C, there was no glucose after 48 h because of the rapid consumption by the yeast. The increase in the ethanol yield after 48 h can be attributed to the hydrolysis of cellobiose. β-glucosidase was active at 40 °C and continued to decompose cellobiose into glucose, which was rapidly turned into ethanol by the mutated strains, so there was almost no cellobiose accumulation. This also explains why the final ethanol yield of the mutant at 40 °C was higher than that at 38 °C. The activity of the original strain at 40 °C was lower than that at 38 °C so the residual concentration of glucose at 40 °C was higher than that at 38 °C.
Despite the different pretreatment methods and parameters used, the final ethanol titers obtained in previous studies on the production of ethanol from wheat straw using the SSF process were mostly in the range of 18–52 g/L [43]. At a target temperature of 40 °C, at the end of SSF, the ethanol concentrations of YUV1-1 and YUV2-2 were 41.31 g/L and 41.69 g/L, respectively (detailed in Table 1). These values were 8.30 g/L and 8.68 g/L higher than that of S. cerevisiae BY4742 (33.01 g/L) and were at an upstream level in the studies of ethanol production from the same substrate. These phenomena can be explained by the following factors: the heat tolerance of the mutated yeast increased, and the enzyme activity of the mutated strains maintained a high level, thus increasing the concentration and production of ethanol. A higher fermentation broth and ethanol concentration can greatly reduce the distillation cost. Given the potentially huge scale of straw bioethanol, the economic cost savings would be substantial.
Heat stress has been shown to activate various stress-responsive genes, including those encoding heat-shock proteins (HSPs), enzymes involved in protein degradation such as ubiquitin ligase, and proteins involved in trehalose and glycogen metabolism in yeast [44]. The following research focuses on the expression of these genes to reveal the mechanism of improved heat resistance of mutated strains at the molecular-biology level.
Lignocellulose is the most important potential feedstock for bioethanol, and biologists estimate that the total amount of biomass growing on Earth each year amounts to several hundred billion tons. It serves as the material basis for biomass energy, which can be converted into biofuels equivalent to 34 billion to 160 billion barrels of crude oil. Although the production of lignocellulosic ethanol through the SSF process has many advantages, it also faces many challenges such as the high cost of cellulase and the high energy consumption per unit of product. Free cellulase is difficult to recover and its activity is highly sensitive to the external environment. Factors such as high temperature, high pressure, acidity, and alkalinity, as well as changes in the concentrations of reactants and products, can inhibit enzyme activity. Immobilization of cellulase can improve its stability, catalytic activity, selectivity, and resistance to inhibition, allowing it to be used repeatedly. The performance of yeast is strongly inhibited by the high concentrations of the substrate and end products, which restricts the generation of ethanol and the promotion of industrialized lignocellulose-based ethanol production. In situ ethanol removal and batch-feeding strategies can reduce the substrate and ethanol concentrations in the fermentation broth. When batch-feeding or in situ ethanol removal processes are used with SSF, much higher ethanol concentrations and cellulose conversion can be achieved with minimal enzyme dosage. Simultaneously, ethanol production can be enhanced more effectively [45,46,47,48].
4. Conclusions
This work proved that the performance of Saccharomyces cerevisiae in glucose fermentation and wheat straw SSF can be improved by UV mutagenesis and higher-temperature screening. Two thermotolerant strains, YUV1-1 and YUV2-2, were obtained through UV mutagenesis and screening of high-temperature cultures, and the validity of high-temperature fermentation experiments was confirmed. YUV1-1 and YUV2-2 achieved ethanol yields of 55.84% and 57.09%, respectively, during the process of fermentation, and 77.30% and 78.01% during the process of SSF at 40 °C. These yields were higher than those of the original yeast S. cerevisiae BY4742. A slight increase in the temperature of the ethanol production process can result in significant cost savings for distillation, given the rising demand for bioenergy. Thermotolerant strains have high application value in increasing the temperature of fermentation and SSF.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/fermentation9070608/s1. Figure S1: Yeasts stained with methylene blue after 20 s of UV radiation treatment; Figure S2: Yeast lethally rates under UV irradiation; Table S1: The components of the fermentation medium.
Author Contributions
Conceptualization, X.L. and Z.W.; Methodology, X.L.; Software, X.L.; Validation, X.L., Y.L. and Z.W.; Investigation, X.L.; Resources, Y.L. and H.K.; Data curation, X.L.; Writing—original draft, X.L.; Writing—review & editing, Y.L., H.K. and Z.W.; Visualization, X.L.; Supervision, Y.L. and H.K.; Project administration, Y.L.; Funding acquisition, Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Research on the Improvement of Water Environment Quality in Sanyang Wetland: Study on Ecological Coagulation Remediation Technology for High Turbidity Water (SY2022ZD-1002-03) and Research on the Hyperthermophilic Co-composting of Sludge and the Resource Utilization of Products (N20220014).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data supporting the findings of this study are included in this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Mathimani, T.; Mallick, N. A review on the hydrothermal processing of microalgal biomass to bio-oil-knowledge gaps and recent advances. J. Clean. Prod. 2019, 217, 69–84. [Google Scholar] [CrossRef]
- Zabed, H.; Sultana, S.; Sahu, J.N.; Qi, X. An overview on the application of ligninolytic microorganisms and enzymes for pretreatment of lignocellulosic biomass. In Recent Advancements in Biofuels and Bioenergy Utilization; Sarangi, P.K., Nanda, S., Mohanty, P., Eds.; Springer: Singapore, 2018; pp. 53–72. [Google Scholar]
- Toor, M.; Kumar, S.S.; Malyan, S.K.; Bishnoi, N.R.; Mathimani, T.; Rajendran, K.; Pugazhendhi, A. An overview on bioethanol production from lignocellulosic feedstocks. Chemosphere 2020, 242, 125080. [Google Scholar] [CrossRef]
- Liu, C.-G.; Xiao, Y.; Xia, X.-X.; Zhao, X.-Q.; Peng, L.; Srinophakun, P.; Bai, F.-W. Cellulosic ethanol production: Progress, challenges and strategies for solutions. Biotechnol. Adv. 2019, 37, 491–504. [Google Scholar] [CrossRef]
- Limayem, A.; Ricke, S.C. Lignocellulosic biomass for bioethanol production: Current perspectives, potential issues and future prospects. Prog. Energ. Combust. Sci. 2012, 38, 449–467. [Google Scholar] [CrossRef]
- Carrillo-Nieves, D.; Rostro Alanis, M.J.; de la Cruz Quiroz, R.; Ruiz, H.A.; Iqbal, H.M.N.; Parra-Saldivar, R. Current status and future trends of bioethanol production from agro-industrial wastes in Mexico. Renew. Sust. Energ. Rev. 2019, 102, 63–74. [Google Scholar] [CrossRef]
- Zhao, J.; Xia, L. Simultaneous saccharification and fermentation of alkaline-pretreated corn stover to ethanol using a recombinant yeast strain. Fuel Process. Technol. 2009, 90, 1193–1197. [Google Scholar] [CrossRef]
- Tan, H.; Yu, Y.; Zhu, Y.; Liu, T.; Miao, R.; Hu, R.; Peng, W.; Chen, J. Impacts of size reduction and alkaline-soaking pretreatments on microbial community and organic matter decomposition during wheat straw composting. Bioresour. Technol. 2022, 360, 127549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lin, Y.; Zhang, Q.; Wang, X.; Wu, D.; Kong, H. Optimisation of simultaneous saccharification and fermentation of wheat straw for ethanol production. Fuel 2013, 112, 331–337. [Google Scholar] [CrossRef]
- Rodionova, M.V.; Bozieva, A.M.; Zharmukhamedov, S.K.; Leong, Y.K.; Lan, J.C.-W.; Veziroglu, A.; Veziroglu, T.N.; Tomo, T.; Chang, J.-S.; Allakhverdiev, S.I. A comprehensive review on lignocellulosic biomass biorefinery for sustainable biofuel production. Int. J. Hydrog. Energ. 2022, 47, 1481–1498. [Google Scholar] [CrossRef]
- Zhao, F.; Fang, S.; Gao, Y.; Bi, J. Removal of aqueous pharmaceuticals by magnetically functionalized Zr-MOFs: Adsorption kinetics, isotherms, and regeneration. J. Colloid. Interf. Sci. 2022, 615, 876–886. [Google Scholar] [CrossRef]
- Hoffman, S.M.; Alvarez, M.; Alfassi, G.; Rein, D.M.; Garcia-Echauri, S.; Cohen, Y.; Avalos, J.L. Cellulosic biofuel production using emulsified simultaneous saccharification and fermentation (eSSF) with conventional and thermotolerant yeasts. Biotechnol. Biofuels 2021, 14, 1–17. [Google Scholar] [CrossRef]
- Saratale, R.G.; Cho, S.K.; Bharagava, R.N.; Patel, A.K.; Varjani, S.; Mulla, S.I.; Kim, D.S.; Bhatia, S.K.; Ferreira, L.F.R.; Shin, H.S.; et al. A critical review on biomass-based sustainable biorefineries using nanobiocatalysts: Opportunities, challenges, and future perspectives. Bioresour. Technol. 2022, 363, 127926. [Google Scholar] [CrossRef]
- Patel, A.; Patel, H.; Divecha, J.; Shah, A.R. Enhanced production of ethanol from enzymatic hydrolysate of microwave-treated wheat straw by statistical optimization and mass balance analysis of bioconversion process. Biofuels 2021, 12, 1251–1258. [Google Scholar] [CrossRef]
- Cheng, G.; Zhao, Y.; Pan, S.; Wang, X.; Dong, C. A comparative life cycle analysis of wheat straw utilization modes in China. Energy 2020, 194, 116914. [Google Scholar] [CrossRef]
- Xu, X.; Wang, X.; He, Y. The crucial problems and solutions on producing fuel ethanol by straw. Food Ferment. Ind. 2010, 36, 108–113. [Google Scholar]
- Arnoult, S.; Obeuf, A.; Bethencourt, L.; Mansard, M.-C.; Brancourt-Hulmel, M. Miscanthus clones for cellulosic bioethanol production: Relationships between biomass production, biomass production components, and biomass chemical composition. Ind. Crop Prod. 2015, 63, 316–328. [Google Scholar] [CrossRef]
- Olofsson, K.; Bertilsson, M.; Liden, G. A short review on SSF-an interesting process option for ethanol production from lignocellulosic feedstocks. Biotechnol. Biofuels 2008, 1, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Juneja, A.; Singh, V. Fermentation technology to improve productivity in dry grind corn process for bioethanol production. Fuel Process. Technol. 2018, 173, 66–74. [Google Scholar] [CrossRef]
- Park, I.; Kim, I.; Kang, K.; Sohn, H.; Rhee, I.; Jin, I.; Jang, H. Cellulose ethanol production from waste newsprint by simultaneous saccharification and fermentation using Saccharomyces cerevisiae KNU5377. Process. Biochem. 2010, 45, 487–492. [Google Scholar] [CrossRef]
- Menon, V.; Divate, R.; Rao, M. Bioethanol production from renewable polymer lichenan using lichenase from an alkalothermophilic Thermomonospora sp. and thermotolerant yeast. Fuel Process. Technol. 2011, 92, 401–406. [Google Scholar] [CrossRef]
- Liu, K.; Jiang, B.; Wang, Q.; Yang, L.; Lu, B.; Li, X.; Yuan, H. Study on the conditions of pretreating vinegar residue with sodium hydroxide for simultaneous saccharification and fermentation to produce alcohol and xylose. Food Sci. Technol. Res. 2020, 26, 381–388. [Google Scholar] [CrossRef]
- Carrillo-Nieves, D.; Ruiz, H.A.; Aguilar, C.N.; Ilyina, A.; Parra-Saldivar, R.; Antonio Torres, J.; Martinez Hernandez, J.L. Process alternatives for bioethanol production from mango stem bark residues. Bioresour. Technol. 2017, 239, 430–436. [Google Scholar] [CrossRef]
- Sassner, P.; Galbe, M.; Zacchi, G. Techno-economic evaluation of bioethanol production from three different lignocellulosic materials. Biomass Bioenerg. 2008, 32, 422–430. [Google Scholar] [CrossRef]
- Rudolf, A.; Alkasrawi, M.; Zacchi, G.; Liden, G. A comparison between batch and fed-batch simultaneous saccharification and fermentation of steam pretreated spruce. Enzym. Microb. Technol. 2005, 37, 195–204. [Google Scholar] [CrossRef]
- Claassen, P.A.M.; van Lier, J.B.; Contreras, A.M.L.; van Niel, E.W.J.; Sijtsma, L.; Stams, A.J.M.; de Vries, S.S.; Weusthuis, R.A. Utilisation of biomass for the supply of energy carriers. Appl. Microbiol. Biotechnol. 1999, 52, 741–755. [Google Scholar] [CrossRef]
- Eklund, R.; Zacchi, G. Simultaneous saccharification and fermentation of steam-pretreated willow. Enzym. Microb. Technol. 1995, 17, 255–259. [Google Scholar] [CrossRef]
- Hasunuma, T.; Kondo, A. Consolidated bioprocessing and simultaneous saccharification and fermentation of lignocellulose to ethanol with thermotolerant yeast strains. Process. Biochem. 2012, 47, 1287–1294. [Google Scholar] [CrossRef]
- Sridhar, M.; Sree, N.K.; Rao, L.V. Effect of UV radiation on thermotolerance, ethanol tolerance and osmotolerance of Saccharomyces cerevisiae VS1 and VS3 strains. Bioresour. Technol. 2002, 83, 199–202. [Google Scholar] [CrossRef]
- De Melo, A.H.F.; Lopes, A.M.M.; Dezotti, N.; Santos, I.L.; Teixeira, G.S.; Goldbeck, R. Evolutionary engineering of two robust brazilian industrial yeast strains for thermotolerance and second-generation biofuels. Ind. Biotechnol. 2020, 16, 91–98. [Google Scholar] [CrossRef]
- Li, Y.J.; Wei, H.B.; Wang, T.; Xu, Q.Y.; Zhang, C.L.; Fan, X.G.; Ma, Q.; Chen, N.; Xie, X.X. Current status on metabolic engineering for the production of L-aspartate family amino acids and derivatives. Bioresour. Technol. 2017, 245, 1588–1602. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, C.; Zhou, Q.Q.; Zhang, X.F.; Wang, L.Y.; Chang, H.B.; Li, H.P.; Oda, Y.; Xing, X.H. Quantitative evaluation of DNA damage and mutation rate by atmospheric and room-temperature plasma (ARTP) and conventional mutagenesis. Appl. Microbiol. Biotechnol. 2015, 99, 5639–5646. [Google Scholar] [CrossRef]
- Zheng, D.Q.; Zhang, K.; Gao, K.H.; Liu, Z.W.; Zhang, X.; Li, O.; Sun, J.G.; Zhang, X.Y.; Du, F.G.; Sun, P.Y.; et al. Construction of novel Saccharomyces cerevisiae strains for bioethanol active dry yeast (ADY) production. PLoS ONE 2013, 8, e85022. [Google Scholar] [CrossRef]
- Zhang, B.; Geberekidan, M.; Yan, Z.; Yi, X.; Bao, J. Very high thermotolerance of an adaptive evolved Saccharomyces cerevisiae in cellulosic ethanol fermentation. Fermentation 2023, 9, 393. [Google Scholar] [CrossRef]
- Dodd, M.C. Potential impacts of disinfection processes on elimination and deactivation of antibiotic resistance genes during water and wastewater treatment. J. Environ. Monit. 2012, 14, 1754–1771. [Google Scholar] [CrossRef]
- Boonchuay, P.; Techapun, C.; Leksawasdi, N.; Seesuriyachan, P.; Hanmoungjai, P.; Watanabe, M.; Srisupa, S.; Chaiyaso, T. Bioethanol production from cellulose-rich corncob residue by the thermotolerant Saccharomyces cerevisiae TC-5. J. Fungi 2021, 7, 547. [Google Scholar] [CrossRef] [PubMed]
- Pessani, N.K.; Atiyeh, H.K.; Wilkins, M.R.; Bellmer, D.D.; Banat, I.M. Simultaneous saccharification and fermentation of Kanlow switchgrass by thermotolerant Kluyveromyces marxianus IMB3: The effect of enzyme loading, temperature and higher solid loadings. Bioresour. Technol. 2011, 102, 10618–10624. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, S.; Fan, C.; Zheng, X.; Zhang, W.; Wu, D.; Wang, X.; Kong, H. Optimisation of enzymatic saccharification of wheat straw pre-treated with sodium hydroxide. Sci. Rep. 2021, 11, 23234. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, D.; Lin, Y.; Wang, X.; Kong, H.; Tanaka, S. Substrate and product inhibition on yeast performance in ethanol fermentation. Energy Fuels 2015, 29, 1019–1027. [Google Scholar] [CrossRef]
- Yi, Y.I. Mutation screening of Saccharomyces cerevisiae strains with alcohol tolerance. J. Anhui Agric. Sci. 2009, 37, 3. [Google Scholar]
- Yao, Z.; Zhou, P.; Su, B.; Su, S.; Ye, L.; Yu, H. Enhanced Isoprene production by reconstruction of metabolic balance between strengthened precursor supply and improved isoprene synthase in Saccharomyces cerevisiae. ACS Synth. Biol. 2018, 7, 2308–2316. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, W.; Li, C.; Sakakibara, K.; Tanaka, S.; Kong, H. Factors affecting ethanol fermentation using Saccharomyces cerevisiae BY4742. Biomass Bioenerg. 2012, 47, 395–401. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, B.; Liu, B.; Yi, Y.; Shan, Y.; Zhou, Y.; Wang, X.; Lu, X. Full components conversion of lignocellulose via a closed-circuit biorefinery process on a pilot scale. Environ. Res. 2022, 214 Pt 2, 113946. [Google Scholar] [CrossRef]
- Nuanpeng, S.; Thanonkeo, S.; Yamada, M.; Thanonkeo, P. Ethanol production from sweet sorghum juice at high temperatures using a newly isolated thermotolerant Yeast Saccharomyces cerevisiae DBKKU Y-53. Energies 2016, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.G.; Cheng, H.; Wu, J.L.; Chen, X.C.; Ying, H.J.; Zhou, P.; Chen, Y. Long-term production of fuel ethanol by immobilized yeast in repeated-batch simultaneous saccharification and fermentation of Cassava. Energy Fuels 2015, 29, 185–190. [Google Scholar] [CrossRef]
- Liu, K.; Lin, X.H.; Yue, J.; Li, X.Z.; Fang, X.; Zhu, M.T.; Lin, J.Q.; Qu, Y.B.; Xiao, L. High concentration ethanol production from corncob residues by fed-batch strategy. Bioresour. Technol. 2010, 101, 4952–4958. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.Y.; Liu, R.H.; Shen, F.; Wu, H.J. Optimization of fermentation conditions for the production of ethanol from stalk juice of sweet sorghum by immobilized yeast using response surface methodology. Energy Fuels 2009, 23, 487–491. [Google Scholar] [CrossRef]
- Sassner, P.; Galbe, M.; Zacchi, G. Bioethanol production based on simultaneous saccharification and fermentation of steam-pretreated Salix at high dry-matter content. Enzym. Microb. Technol. 2006, 39, 756–762. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of the screening process and obtained strains.

Figure 2.
Yeast concentrations (a), glucose utilization rates (b), and ethanol yields (c) of the first mutated strains during fermentation at 42 °C.
Figure 2.
Yeast concentrations (a), glucose utilization rates (b), and ethanol yields (c) of the first mutated strains during fermentation at 42 °C.

Figure 3.
Yeast concentrations (a), glucose utilization rates (b), and ethanol yields (c) of the second-mutated strains during the process of fermentation at 42 °C.
Figure 3.
Yeast concentrations (a), glucose utilization rates (b), and ethanol yields (c) of the second-mutated strains during the process of fermentation at 42 °C.

Figure 4.
Yeast concentrations (a), glucose utilization rates (b), and ethanol yields (c) of the mutated strains during the process of fermentation test at 40 °C.
Figure 4.
Yeast concentrations (a), glucose utilization rates (b), and ethanol yields (c) of the mutated strains during the process of fermentation test at 40 °C.

Figure 5.
SSF profile of YUV1-1 (a), YUV2-2 (b), and S. cerevisiae BY4742 (c) at 40 °C and 38 °C.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The final yeast cell mass concentration (CY), ethanol concentration (CE), calculated ethanol yields (YCE, the ratio of the final ethanol mass to the consumed substrate mass), and final ethanol volumetric productivity (VPE, the final ethanol concentration produced per unit time) at the end of the fermentation and SSF processes.
Table 1.
The final yeast cell mass concentration (CY), ethanol concentration (CE), calculated ethanol yields (YCE, the ratio of the final ethanol mass to the consumed substrate mass), and final ethanol volumetric productivity (VPE, the final ethanol concentration produced per unit time) at the end of the fermentation and SSF processes.
| Condition | Strain Name | CY, g/L | CE, g/L | YCE, g/g | VPE, g/L/h |
|---|---|---|---|---|---|
| Fermentation at 40 °C for 60 h | S. cerevisiae BY4742 | 2.52 | 16.82 | 0.34 | 0.28 |
| YUV1-1 | 3.07 | 22.85 | 0.39 | 0.38 | |
| YUV2-2 | 3.03 | 23.44 | 0.39 | 0.39 | |
| SSF at 40 °C for 120 h | S. cerevisiae BY4742 | - | 33.01 | 0.21 | 0.28 |
| YUV1-1 | - | 41.31 | 0.26 | 0.34 | |
| YUV2-2 | - | 41.69 | 0.26 | 0.35 | |
| SSF at 38 °C for 120 h | S. cerevisiae BY4742 | - | 38.33 | 0.24 | 0.32 |
| YUV1-1 | - | 39.68 | 0.25 | 0.33 | |
| YUV2-2 | - | 40.41 | 0.25 | 0.34 |
“-” indicates that no values are available.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, X.; Lin, Y.; Kong, H.; Wang, Z. Screening of Ultraviolet-Induced Thermotolerant Yeast Mutants and Their Performance. Fermentation 2023, 9, 608. https://doi.org/10.3390/fermentation9070608
AMA Style
Li X, Lin Y, Kong H, Wang Z. Screening of Ultraviolet-Induced Thermotolerant Yeast Mutants and Their Performance. Fermentation. 2023; 9(7):608. https://doi.org/10.3390/fermentation9070608
Chicago/Turabian StyleLi, Xiaodi, Yan Lin, Hainan Kong, and Zhiquan Wang. 2023. "Screening of Ultraviolet-Induced Thermotolerant Yeast Mutants and Their Performance" Fermentation 9, no. 7: 608. https://doi.org/10.3390/fermentation9070608
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.