

The Influence of Cabernet Sauvignon Ripeness, Healthy State and Maceration Time on Wine and Fermented Pomace Phenolic Profile
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals and Reagents
2.3. Winemaking
2.3.1. Different Ripening Stages
2.3.2. Healthy Grapes vs. Grapes Affected by Botrytis cinerea
2.3.3. Different Vinifications and the Dynamics of Phenolic Compound Extraction
2.4. Sample Preparation
2.5. LC-MS/MS Analysis
2.6. Determination of Titratable Acidity
2.7. Determination of Total Soluble Solids-Sugar Content
2.8. Total Phenolic Content (TPC)
2.9. Ferric Reducing Activity of Plasma (FRAP)
2.10. Trolox Equivalent Antioxidant Capacity (TEAC)
2.11. Statistical Analysis
3. Results and Discussion
3.1. Phenolic Profile and Antioxidant Properties of Wine and FP Obtained from Grapes of Different Ripeness
3.2. Influence of Botrytis cinerea on the Phenolic Profile of Wine
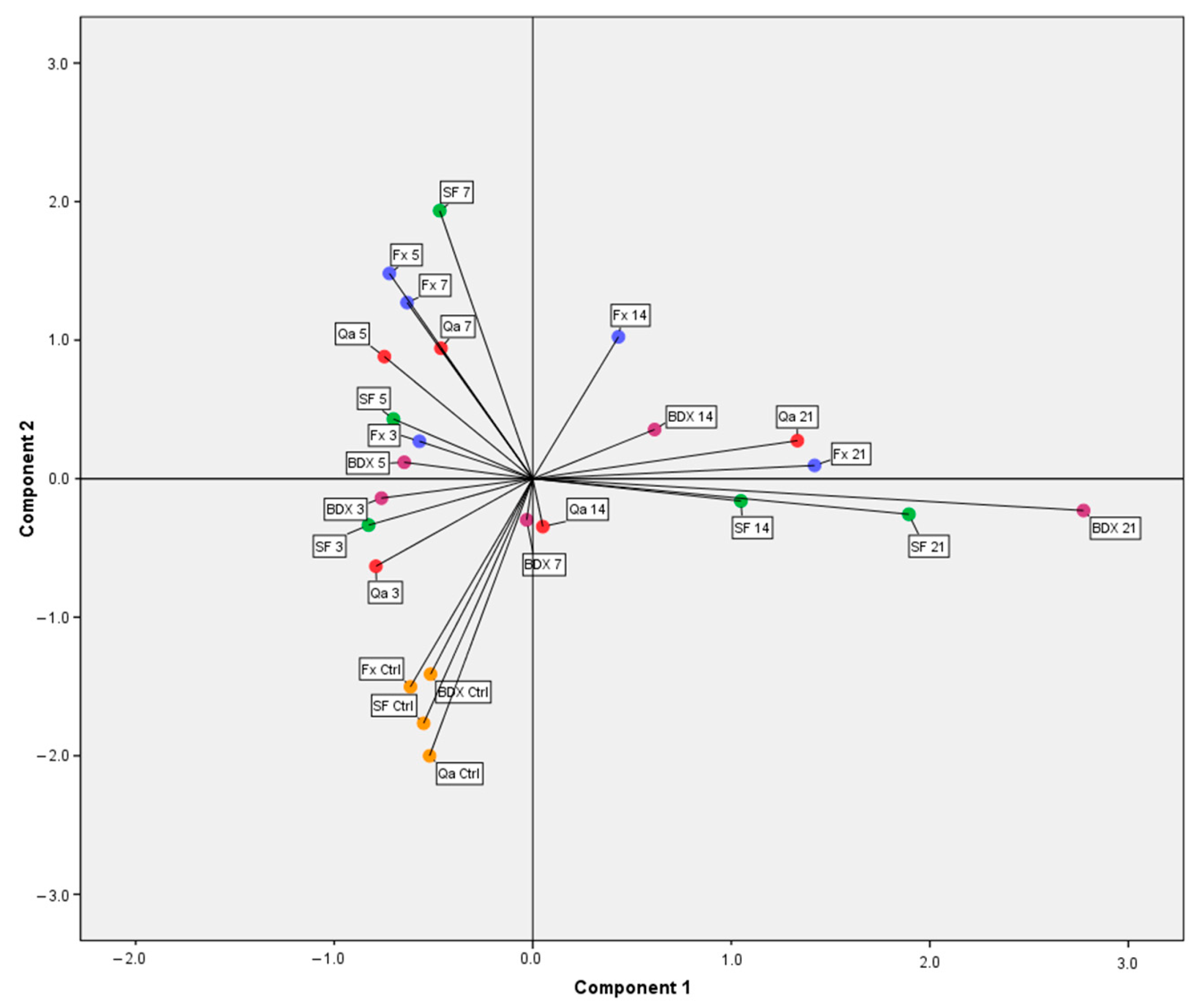
3.3. Dynamic of Phenolic Compounds Extraction during Inoculated and Spontaneous Fermentations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology: The Microbiology of Wine and Vinifications, 2nd ed.; Jonh Wiley & Sons: Chichester, UK, 2006; Volume 1, pp. 283–294. [Google Scholar]
- Generalić Mekinić, I.; Skračić, Ž.; Kokeza, A.; Soldo, B.; Ljubenkov, I.; Banović, M.; Skroza, D. Effect of winemaking on phenolic profile, colour components and antioxidants in Crljenak kaštelanski (sin. Zinfandel, Primitivo, Tribidrag) wine. J. Food Sci. Technol. 2019, 56, 1841–1853. [Google Scholar] [CrossRef] [PubMed]
- Francesca, N.; Romano, R.; Sannino, C.; Le Grottaglie, L.; Settanni, L.; Moschetti, G. Evolution of microbiological and chemical parameters during red wine making with extended post-fermentation maceration. Int. J. Food Microbiol. 2014, 171, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, A.; Adams, D.; Boss, P.; Heymann, H.; Solomon, P.; Trengove, R. Influence of geographic origin on the sensory characteristics and wine composition of Vitis vinifera cv. Cabernet sauvignon wines from Australia. Am. J. Enol. Vitic. 2012, 63, 467–476. [Google Scholar] [CrossRef]
- Radovanović, B.; Radovanović, A.; Souquet, J. Phenolic profile and free radical-scavenging activity of Cabernet Sauvignon wines of different geographical origins from the Balkan region. J. Sci. Food Agric. 2010, 90, 2455–2461. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Destrac-Irvine, A. Modified grape composition under climate change conditions requires adaptations in the vineyard. Oeno One 2017, 51, 147–154. [Google Scholar] [CrossRef]
- Madžgalj, V.; Petrović, A.; Čakar, U.; Maraš, V.; Sofrenić, I.; Tešević, V. The influence of different enzymatic preparations and skin contact time on the aromatic profile of wines produced from autochthonous grape varieties Krstač and Žižak. J. Serb. Chem. Soc. 2022, 88, 11–23. [Google Scholar] [CrossRef]
- Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan application in vineyards (Vitis vinifera L. cv. Tinto Cão) induces accumulation of anthocyanins and other phenolics in berries, mediated by modifications in the transcription of secondary metabolism genes. Int. J. Mol. Sci. 2020, 21, 306. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Chen, S.; Mi, Z.; Su, J.; Huang, R.; Song, Y.; Fang, Y.; Su, B. Identifying veraison process of coloured wine grapes in field conditions combining deep learning and image analysis. Comput. Electron. Agric. 2022, 200, 107268. [Google Scholar] [CrossRef]
- Medina-Plaza, C.; Meade, H.; Dokoozlian, N.; Ponangi, R.; Blair, T.; Block, D.E.; Oberholster, A. Investigating the Relation between Skin Cell Wall Composition and Phenolic Extractability in Cabernet Sauvignon Wines. Ferment 2022, 8, 401. [Google Scholar] [CrossRef]
- Steel, C.C.; Blackman, J.W.; Schmidtke, L.M. Grapevine bunch rots Impacts on wine composition, quality, and potential procedures for the removal of wine faults. J. Agric. Food Chem. 2013, 61, 5189–5206. [Google Scholar] [CrossRef]
- Blanco-Ulate, B.; Amrine, K.C.; Collins, T.S.; Rivero, R.M.; Vicente, A.R.; Morales-Cruz, A.; Doyle, C.L.; Ye, Z.; Greg, A.; Heymann, H.; et al. Developmental and metabolic plasticity of white-skinned grape berries in response to Botrytis cinerea during noble rot. Plant Physiol. 2015, 169, 2422–2443. [Google Scholar] [CrossRef] [Green Version]
- Quijada-Morin, N.; Garcia, F.; Lambert, K.; Walker, A.S.; Tiers, L.; Viaud, M.; Sauvage, F.X.; Hirtz, C.; Saucier, C. Strain effect on extracellular laccase activities from Botrytis cinerea. Aust. J. Grape Wine Res. 2018, 24, 241–251. [Google Scholar] [CrossRef]
- Lisov, N.; Petrovic, A.; Čakar, U.; Jadranin, M.; Tešević, V.; Bukarica-Gojković, L. Extraction kinetic of some phenolic compounds during Cabernet Sauvignon alcoholic fermentation and antioxidant properties of derived wines. Maced. J. Chem. Chem. 2020, 39, 185–196. [Google Scholar] [CrossRef]
- de Simón, B.F.; Hernández, T.; Estrella, I.; Gómez-Cordovés, C. Variation in phenol content in grapes during ripening: Low-molecular-weight phenols. Z. Lebensm. Unters. Forsch. 1992, 194, 351–354. [Google Scholar] [CrossRef]
- Peña-Neira, A.; Dueñas, M.; Duarte, A.; Hernandez, T.; Estrella, I.; Loyola, E. Effects of ripening stages and plant vegetative vigour on the phenolic composition of grapes (Vitis vinifera L.) cv. Cabernet sauvignon in the Maipo Valley (Chile). Vitis 2004, 43, 51–57. [Google Scholar] [CrossRef]
- Özcan, M.M.; Al Juhaimi, F.; Gülcü, M.; Uslu, N.; Geçgel, Ü.; Ghafoor, K.; Dursun, N. Effect of harvest time on physics-co-chemical properties and bioactive compounds of pulp and seeds of grape varieties. J. Food Sci. Technol. 2017, 54, 2230–2240. [Google Scholar] [CrossRef]
- Gil-Sánchez, I.; Cueva, C.; Sanz-Buenhombre, M.; Guadarrama, A.; Moreno-Arribas, M.V.; Bartolomé, B. Dynamic gastrointestinal digestion of grape pomace extracts: Bioaccessible phenolic metabolites and impact on human gut microbiota. J. Food Compos. 2018, 68, 41–52. [Google Scholar] [CrossRef]
- Jiang, Y.; Simonsen, J.; Zhao, Y. Compression-molded biocomposite boards from red and white wine grape pomaces. J. Appl. Polym. Sci. 2011, 119, 2834–2846. [Google Scholar] [CrossRef]
- Ivanova, V.; Vojnoski, B.; Stefova, M. Effect of winemaking treatment and wine ageing on phenolic content in Vranec wines. J. Food Sci. Technol. 2012, 49, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Kocabey, N.; Yilmaztekin, M.; Hayaloglu, A.A. Effect of maceration duration on physicochemical characteristics, organic acid, phenolic compounds and antioxidant activity of red wine from Vitis vinifera L. Karaoglan. J. Food Sci. Technol. 2016, 53, 3557–3565. [Google Scholar] [CrossRef] [Green Version]
- Anđelkovic, M.; Radovanović, B.; Radovanović, A.; Anđelkovic, A.M. Changes in polyphenolic content and antioxidant activity of grapes cv Vranac during ripening. South Afr. J. Enol. Vitic. 2013, 34, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Obreque-Slier, E.; Peña-Neira, Á.; López-Solís, R.; Cáceres-Mella, A.; Toledo-Araya, H.; López Rivera, A. Phenolic composition of skins from four Carmenet grape varieties (Vitis vinifera L.) during ripening. LWT-Food Sci. Technol. 2013, 54, 404–413. [Google Scholar] [CrossRef]
- Alencar, N.M.M.; Cazarin, C.B.B.; Corrêa, L.C.; Maróstica Junior, M.R.; Biasoto, A.C.T.; Behrens, J.H. Influence of maceration time on phenolic compounds and antioxidant activity of the Syrah must and wine. J. Food Biochem. 2017, 42, e12471. [Google Scholar] [CrossRef]
- Lingua, M.S.; Fabani, M.P.; Wunderlin, D.A.; Baroni, M.V. From grape to wine: Changes in phenolic composition and its influence on antioxidant activity. Food Chem. 2016, 208, 228–238. [Google Scholar] [CrossRef]
- Antoniolli, A.; Fontana, A.R.; Piccoli, P.; Bottini, R. Characterization of polyphenols and evaluation of antioxidant capacity in grape pomace of the cv. Malbec. Food Chem. 2015, 178, 172–178. [Google Scholar] [CrossRef]
- Ribeiro, L.F.; Ribani, R.H.; Francisco, T.M.G.; Soares, A.A.; Pontarolo, R.; Haminiuk, C.W.I. Profile of bioactive compounds from grape pomace (Vitis vinifera and Vitis labrusca) by spectrophotometric, chromatographic and spectral analyses. J. Chromatogr. B 2015, 1007, 72–80. [Google Scholar] [CrossRef]
- Ferreiro-González, M.; Carrera, C.; Ruiz-Rodríguez, A.; Barbero, G.F.; Ayuso, J.; Palma, M.; Barroso, C.G. A new solid phase extraction for the determination of anthocyanins in grapes. Molecules 2014, 19, 21398–21410. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Xiao, C.; Liu, L.; Hao, M.; Wang, J.; Liu, X. Simultaneous determination of 15 phenolic constituents of Chinese black rice wine by HPLC-MS/MS with SPE. J. Food Sci. 2014, 79, C1100-5. [Google Scholar] [CrossRef]
- Tanner, H.; Brunner, H.R. Gentranke-Analytik; Verlag Heller-Chemie und Verwaltunsgesellschaft mbH: Darmstadt, Germany, 1979. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolourization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Pérez-Magariño, S.; González-San José, M.L. Effect of ripening stage of grapes on the low molecular weight phenolic compounds of red wines. Eur. Food Res. Technol. 2005, 220, 597–606. [Google Scholar] [CrossRef]
- Casassa, L.F.; Sari, S.E.; Bolcato, E.A.; Fanzone, M.L. Microwave-assisted extraction applied to Merlot grapes with contrasting maturity levels: Effects on phenolic chemistry and wine colour. Fermentation 2019, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Nunes, M.A.; Pimentel, F.; Costa, A.S.; Alves, R.C.; Oliveira, M.B.P. Cardioprotective properties of grape seed proanthocyanidins: An update. Trends Food Sci. 2016, 57, 31–39. [Google Scholar] [CrossRef]
- Du, Y.; Li, X.; Xiong, X.; Cai, X.; Ren, X.; Kong, Q. An investigation on polyphenol composition and content in the skin of the grape (Vitis vinifera L. cv. Hutai No. 8) fruit during ripening by UHPLC-MS2 technology combined with multivariate statistical analysis. Food Biosci. 2021, 43, 101276. [Google Scholar] [CrossRef]
- Fernandes, A.C.F.; Martins, I.M.; Moreira, D.K.T.; Macedo, G.A. Use of agro-industrial residues as a potent antioxidant, antiglycation agents, and α-amylase and pancreatic lipase inhibitory activity. J. Food Process. Preserv. 2020, 44, e14397. [Google Scholar] [CrossRef]
- Muñoz-Bernal, Ó.A.; Coria-Oliveros, A.J.; Vazquez-Flores, A.A.; de la Rosa, L.A.; NúñezGastélum, J.A.; Rodrigo-García, J.; Ayala-Zavala, J.F.; Alvarez-Parrilla, E. Evolution of phenolic content, antioxidant capacity and phenolic profile during cold pre-fermentative maceration and subsequent fermentation of Cabernet sauvignon red wine. South Afr. J. Enol. Vitic. 2020, 41, 72–82. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Xing, K.; Zhang, X.X.; Wang, H.; Wang, Y.; Wang, F.; Li, J.M. Influence of freeze concentration technique on aromatic and phenolic compounds, colour attributes, and sensory properties of Cabernet Sauvignon wine. Molecules 2017, 22, 899. [Google Scholar] [CrossRef] [Green Version]
- Kahkeshani, N.; Farzaei, F.; Fotouhi, M.; Alavi, S.S.; Bahramsoltani, R.; Naseri, R.; Momtaz, S.; Abbasabadi, Z.; Rahimi, R.; Farzaei, M.H.; et al. Pharmacological effects of gallic acid in health and diseases: A mechanistic review. Iran. J. Basic Med. Sci. 2019, 22, 225. [Google Scholar] [CrossRef]
- Ekhtiar, M.; Ghasemi-Dehnoo, M.; Mirzaei, Y.; Azadegan-Dehkordi, F.; Amini-Khoei, H.; Lorigooini, Z.; Samiei-Sefat, A.; Bagheri, N. The coumaric acid and syringic acid ameliorate acetic acid-induced ulcerative colitis in rats via modulator of Nrf2/HO-1 and pro-inflammatory cytokines. Int. Immunopharmacol. 2023, 120, 110309. [Google Scholar] [CrossRef]
- Niu, S.; Hao, F.; Mo, H.; Jiang, J.; Wang, H.; Liu, C.; Fan, X.; Zhang, Y. Phenol profiles and antioxidant properties of white-skinned grapes and their coloured genotypes during growth. Biotechnol. Biotechnol. Equip. 2017, 31, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.R.; Choi, C.; Kim, M.J.; Mun, B.Y.; Ko, H.; Oh, K.N.; Jo, A.; Kim, J.Y.; Bae, D. Antidepressant effects of p-coumaric acid isolated from Vaccinium bracteatum leaves extract on chronic restraint stress mouse model and antagonism of serotonin 6 receptor in vitro. Phytomedicine 2023, 116, 154871. [Google Scholar] [CrossRef] [PubMed]
- Blancquaert, E.H.; Oberholster, A.; Ricardo-da-Silva, J.M.; Deloire, A.J. Grape flavonoid evolution and composition under altered light and temperature conditions in Cabernet Sauvignon (Vitis vinifera L.). Front. Plant Sci. 2019, 10, 1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, G.; Xiang, L.; Xiao, L. Quercetin alleviates atherosclerosis by suppressing oxidized LDL-induced senescence in plaque macrophage via inhibiting the p38MAPK/p16 pathway. J. Nutr. Biochem. 2023, 116, 109314. [Google Scholar] [CrossRef]
- Jediyi, H.; Naamani, K.; Elkoch, A.A.; Dihazi, A.; El Fels, A.E.A.; Arkize, W. First study on technological maturity and phenols composition during the ripeness of five Vitis vinifera L. grape varieties in Morocco. Sci. Hortic. 2019, 246, 390–397. [Google Scholar] [CrossRef]
- Kennedy, J. Understanding grape berry development. Pract. Winery Vineyard 2002, 4, 1–5. [Google Scholar]
- Bautista-Ortín, A.B.; Busse-Valverde, N.; Fernández-Fernández, J.I.; Gómez-Plaza, E.; Gil-Muñoz, R. The extraction kinetics of anthocyanins and proanthocyanidins from grape to wine in three different varieties. J. Int. Sci. Vigne. Vin. 2016, 50, 91–100. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Hernanz, D.; Escudero-Gilete, M.L.; Heredia, F.J. Antioxidant potential of white grape pomaces: Phenolic composition and antioxidant capacity measured by spectrophotometric and cyclic voltammetry methods. Food Res. Int. 2014, 66, 150–157. [Google Scholar] [CrossRef]
- Benbouguerra, N.; Richard, T.; Saucier, C.; Garcia, F. Voltammetric Behavior, Flavanol and Anthocyanin Contents, and Antioxidant Capacity of Grape Skins and Seeds during Ripening (Vitis vinifera var. Merlot, Tannat, and Syrah). Antioxidants 2020, 9, 800. [Google Scholar] [CrossRef]
- Prakash, O.; Supriya, A.; Kudachikar, V.B. Physicochemical Changes, Phenolic Profile and Antioxidant Capacities of Colored and White Grape (Vitis vinifera L.) Varieties during Berry Development and Maturity. Int. J. Fruit Sci. 2020, 20, 1773–1783. [Google Scholar] [CrossRef]
- Ky, I.; Lorrain, B.; Jourdes, M.; Pasquier, G.; Fermaud, M.; Gény, L.; Rey, P.; Donesche, B.; Teissedre, P.L. Assessment of grey mould (Botrytis cinerea) impact on phenolic and sensory quality of Bordeaux grapes, musts and wines for two consecutive vintages. Aust. J. Grape Wine Res. 2012, 18, 215–226. [Google Scholar] [CrossRef]
- Zinnai, A.; Venturi, F.; Sanmartin, C.; Quartacci, M.F.; Andrich, G. Chemical and laccase catalysed oxidation of gallic acid: Determination of kinetic parameters. Res. J. Biotechnol. 2013, 8, 62–65. [Google Scholar]
- Terrier, N.; Poncet-Legrand, C.; Cheynier, V. Flavanols, Flavonols and Dihydroflavonols. In Wine Chemistry and Biochemistry; Moreno-Arribas, V., Polo, C., Eds.; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Claus, H. Laccases of Botrytis cinerea. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 339–356. [Google Scholar]
- Clark, M.; Centner, A.M.; Ukhanov, V.; Nagpal, R.; Salazar, G. Gallic acid ameliorates atherosclerosis and vascular senescence and remodels the microbiome in a sex-dependent manner in ApoE−/− mice. J. Nutr. Biochem. 2022, 110, 109132. [Google Scholar] [CrossRef]
- Pal, S.; Ho, N.; Santos, C.; Dubois, P.; Mamo, J.; Croft, K.; Allister, E. Red wine polyphenolics increase LDL receptor expression and activity and suppress the secretion of ApoB100 from human HepG2 cells. J. Nutr. 2003, 133, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Ivanova-Petropulos, V.; Durakova, S.; Ricci, A.; Parpinello, G.P.; Versari, A. Extraction and evaluation of naturally occurring bioactive compounds and change in antioxidant activity during red winemaking. J. Food Sci. Technol. 2016, 53, 2634–2643. [Google Scholar] [CrossRef] [Green Version]
- Baleiras-Couto, M.M.; Guedes, R.; Duarte, F.L.; Fortes, A.M.; Serralheiro, M.L. Untargeted metabolomics discriminates grapes and wines from two Syrah vineyards located in the same wine region. Fermentation 2023, 9, 145. [Google Scholar] [CrossRef]
- Hernández-Maldonado, L.M.; Blancas-Benítez, F.J.; Zamora-Gasga, V.M.; Cárdenas-Castro, A.P.; Tovar, J.; Sáyago-Ayerdi, S.G. In Vitro Gastrointestinal Digestion and Colonic Fermentation of High Dietary Fiber and Antioxidant-Rich Mango (Mangifera indica L.) “Ataulfo”-Based Fruit Bars. Nutrients 2019, 11, 1564. [Google Scholar] [CrossRef] [Green Version]
- Artem, V.; Antoce, A.O.; Geana, E.I.; Ranca, A. Effect of grape yield and maceration time on the phenolic composition of ‘Fetească neagră’organic wine. Not. Bot. Horti Agrobot. 2021, 49, 12345. [Google Scholar] [CrossRef]
- Milenkovic, D.; Declerck, K.; Guttman, Y.; Kerem, Z.; Claude, S.; Weseler, A.R.; Bast, A.; Schroeter, H.; Morand, C.; Vanden Berghe, W. (−)-Epicatechin metabolites promote vascular health through epigenetic reprogramming of endothelial-immune cell signalling and reversing systemic low-grade inflammation. Biochem. Pharmacol. 2020, 173, 113699. [Google Scholar] [CrossRef]
- Patial, V.; Katoch, S.; Chhimwal, J.; Dadhich, G.; Sharma, V.; Rana, A.; Joshi, R.; Padwad, Y. Catechins prevent obesity-induced kidney damage by modulating PPARγ/CD36 pathway and gut-kidney axis in rats. Life Sci. 2023, 316, 121437. [Google Scholar] [CrossRef]
- Setford, P.C.; Jeffery, D.W.; Grbin, P.R.; Muhlack, R.A. Factors affecting extraction and evolution of phenolic compounds during red wine maceration and the role of process modelling. Trends Food Sci. Technol. 2017, 69, 106–117. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Colomo, B.; Suárez, J.A. Cell wall anthocyanin adsorption by different Saccharomyces strains during the fermentation of Vitis vinifera L. cv Graciano grapes. Eur. Food Res. Technol. 2005, 220, 341–346. [Google Scholar] [CrossRef]
- Oliva, E.; Mir-Cerdà, A.; Sergi, M.; Sentellas, S.; Saurina, J. Characterization of sparkling wine based on polyphenolic profiling by liquid chromatography coupled to mass spectrometry. Fermentation 2023, 9, 223. [Google Scholar] [CrossRef]
- de la Cerda-Carrasco, A.; López-Solís, R.; Nuñez-Kalasic, H.; Peña-Neira, Á.; Obreque-Slier, E. Phenolic composition and antioxidant capacity of pomaces from four grape varieties (Vitis vinifera L.). J. Sci. Food Agric. 2015, 95, 1521–1527. [Google Scholar] [CrossRef] [PubMed]
- Melo, P.S.; Massarioli, A.P.; Denny, C.; dos Santos, L.F.; Franchin, M.; Pereira, G.E.; Ferreira de Souza Vieira, T.M.; Rosalen, P.L.; de Alencar, S.M. Winery by-products: Extraction optimization, phenolic composition and cytotoxic evaluation to act as a new source of scavenging of reactive oxygen species. Food Chem. 2015, 181, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Costabile, G.; Vitale, M.; Luongo, D.; Naviglio, D.; Vetrani, C.; Ciciola, P.; Tura, A.; Castello, F.; Mena, P.; Del Rio, D.; et al. Grape pomace polyphenols improve insulin response to a standard meal in healthy individuals: A pilot study. Clin. Nutr. 2019, 38, 2727–2734. [Google Scholar] [CrossRef]
- Zhao, Y.; Dang, M.; Zhang, W.; Lei, Y.; Ramesh, T.; Priya Veeraraghavan, V.; Hou, X. Neuroprotective effects of Syringic acid against aluminium chloride-induced oxidative stress-mediated neuroinflammation in a rat model of Alzheimer’s disease. J. Funct. Foods 2020, 71, 104009. [Google Scholar] [CrossRef]
- Cheng, V.J.; Bekhit, A.E.D.A.; McConnell, M.; Mros, S.; Zhao, J. Effect of extraction solvent, waste fraction and grape variety on the antimicrobial and antioxidant activities of extracts from wine residue from cool climate. Food Chem. 2012, 134, 474–482. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Phenolic Compound | Molecular Formula | Molecular Mass (g/mol) | ESI Mode | MRM Transition | Cone Voltage (V) | Collision Energy (V) | tR (min) |
|---|---|---|---|---|---|---|---|
| Gallic acid | C7H6O5 | 170 | - | 169→125 | 166 | 10 | 3.90 |
| Catechin | C15H14O6 | 290 | - | 289→245 | 166 | 10 | 9.90 |
| Epicatechin | C15H14O6 | 290 | - | 289→245 | 166 | 10 | 11.52 |
| Syringic acid | C9H10O5 | 198 | - | 197→182 | 166 | 7 | 11.60 |
| p-Coumaric acid | C9H8O3 | 164 | - | 163→119 | 166 | 9 | 12.70 |
| Quercetin | C15H10O7 | 302 | - | 301→151 | 166 | 15 | 15.60 |
| Content (mg/L) | ANOVA | ||||
|---|---|---|---|---|---|
| Phenolic Compound | Véraison | Fully Ripe | Overripe | F | p-Value |
| Gallic acid | 5.18 ± 0.20 a | 4.22 ± 0.40 b | 1.59 ± 0.15 | 139.73 | 0.000 * |
| Catechin | 1.55 ± 0.10 a | 40.13 ± 3.25 b | 19.23 ± 1.40 | 267.841 | 0.000 * |
| Epicatechin | 0.73 ± 0.05 a | 22.29 ± 1.50 b | 12.35 ± 0.50 | 418.778 | 0.000 * |
| Syringic acid | 5.57 ± 0.40 a | 7.55 ± 0.50 | 8.26 ± 0.60 | 22.715 | 0.002 * |
| p-Coumaric acid | 4.19 ± 0.35 b | 3.88 ± 0.20 b | 2.33 ± 0.20 | 44.135 | 0.000 * |
| Quercetin | 0.09 ± 0.01 a | 0.78 ± 0.02 b | 0.00 | 3277.8 | 0.000 * |
| Content (mg/kg) | ANOVA | ||||
|---|---|---|---|---|---|
| Phenolic Compound | Véraison | Fully Ripe | Overripe | F | p-Value |
| Gallic acid | 0.86 ± 0.04 b | 0.82 ± 0.04 b | 0.67 ± 0.02 | 25.083 | 0.001 * |
| Catechin | 4.92 ± 0.15 a | 6.73 ± 0.11 b | 3.30 ± 0.09 | 620.564 | 0.000 * |
| Epicatechin | 2.70 ± 0.05 a | 3.37 ± 0.10 b | 1.76 ± 0.04 | 417.511 | 0.000 * |
| Syringic acid | 5.68 ± 0.10 a | 8.53 ± 0.15 b | 10.29 ± 0.25 | 512.719 | 0.000 * |
| p-Coumaric acid | 0.04 ± 0.004 c | 0.05 ± 0.002 b | 0.04 ± 0.002 | 12.500 | 0.007 * |
| Quercetin | 7.44 ± 0.10 a | 10.96 ± 0.14 b | 5.61 ± 0.15 | 440.549 | 0.000 * |
| Ripening Stage | Wine | FP | ||||
|---|---|---|---|---|---|---|
| FRAP (mmol Fe2+/L) | TEAC (mmol TE/L) | TPC (mg/L) | FRAP (mmol Fe2+/KG) | TEAC (mmol TE/KG) | TPC (mg/KG) | |
| Véraison | 19.50 ± 4.25 a | 8.74 ± 2.62 | 1000.0 ± 190.0 b | 16.22 ± 4.82 | 14.93 ± 3.62 | 1320.9 ± 290.0 c |
| Fully ripe | 20.10 ± 4.02 | 16.0 ± 3.20 | 1525.0 ± 210.0 | 26.31 ± 5.21 | 15.90 ± 3.18 | 2260.4 ± 325.0 |
| Overripe | 28.50 ± 2.85 | 15.34 ± 1.51 | 1550.0 ± 235.0 | 16.61 ± 1.58 | 12.43 ± 1.11 | 1629.5 ± 275.0 |
| Content (mg/L) | ANOVA | |||
|---|---|---|---|---|
| Phenolic Compound | Healthy Grape | Grape Affected by Botrytis cinerea | F | p-Value |
| Gallic acid | 2.80 ± 0.10 a | 2.00 ± 0.05 | 153.6 | 0.000 * |
| Catechin | 23.71 ± 0.50 a | 16.95 ± 0.33 | 381.98 | 0.000 * |
| Epicatechin | 10.46 ± 0.20 a | 9.32 ± 0.20 | 48.735 | 0.002 * |
| Syringic acid | 3.73 ± 0.15 a | 8.43 ± 0.40 | 363.123 | 0.000 * |
| p-Coumaric acid | 2.32 ± 0.10 | 2.19 ± 0.08 | 3.091 | 0.154 |
| Quercetin | 24.75 ± 0.50 a | 0.47 ± 0.02 | 7062.920 | 0.000 * |
| Wine (mg/L) | |||||||
|---|---|---|---|---|---|---|---|
| Type of Vinification | Maceration Time | Gallic Acid | Syringic Acid | p-Coumaric Acid | Catechin | Epicatechin | Quercetin |
| SF | 0 (control) | 0.09 ± 0.02 | 0.96 ± 0.20 | 0.73 ± 0.21 | 0.32 ± 0.09 | 0.03 ± 0.004 | 0.01 ± 0.001 |
| 3 | 0.16 ± 0.04 | 7.23 ± 1.98 a | 1.28 ± 0.38 | 0.73 ± 0.21 | 0.08 ± 0.02 | 0.38 ± 0.07 | |
| 5 | 1.02 ± 0.19 | 8.11 ± 1.55 a | 5.31 ± 1.59 a,b | 3.20 ± 0.96 | 0.59 ± 0.11 | 0.32 ± 0.06 | |
| 7 | 2.04 ± 0.31 a,b | 7.21 ± 1.70 a | 6.30 ± 1.89 a,b | 8.63 ± 2.00 | 1.68 ± 0.50 | 2.35 ± 0.47 a,b,c | |
| 14 | 1.82 ± 0.24 a,b | 5.43 ± 1.62 a | 4.03 ± 1.20 | 25.39 ± 5.61 a,b,c,d | 11.53 ± 2.90 a,b,c,d | 0.98 ± 0.20 a,b,c,d | |
| 21 | 3.44 ± 0.80 a,b,c,d,e | 6.58 ± 1.97 a | 5.07 ± 1.52 a,b | 29.72 ± 6.00 a,b,c,d | 16.72 ± 4.01 a,b,c,d | 0.46 ± 0.09 d | |
| BDX | 0 (control) | 0.57 ± 0.11 | 2.70 ± 0.35 | 0.97 ± 0.20 | 0.32 ± 0.07 | 0.06 ± 0.01 | 0.00 |
| 3 | 0.81 ± 0.16 | 8.78 ± 1.99 a | 2.35 ± 0.40 | 0.03 ± 0.001 | 0.00 | 0.00 | |
| 5 | 1.32 ± 0.30 | 6.28 ± 1.70 | 5.56 ± 1.60 a,b | 1.29 ± 0.30 | 0.26 ± 0.02 | 0.21 ± 0.04 | |
| 7 | 2.42 ± 0.70 a,b | 7.50 ± 1.70 a | 0.20 ± 0.04 c | 5.58 ± 1.40 | 1.50 ± 0.20 | 0.53 ± 0.10 a,b,c | |
| 14 | 2.42 ± 0.72 a,b | 7.74 ± 1.55 a | 5.54 ± 1.55 a,b,d | 19.35 ± 4.02 a,b,c,d | 7.60 ± 1.54 a,b,c,d | 0.57 ± 0.12 a,b,c | |
| 21 | 4.22 ± 0.90 a,b,c,d,e | 7.55 ± 2.01 a | 3.88 ± 0.80 a,d | 40.13 ± 5.50 a,b,c,d,e | 22.29 ± 4.00 a,b,c,d,e | 0.78 ± 0.20 a,b,c | |
| FX10 | 0 (control) | 0.11 ± 0.01 | 2.16 ± 0.40 | 1.05 ± 0.20 | 0.06 ± 0.01 | 0.01 ± 0.0001 | 0.02 ± 0.003 |
| 3 | 1.02 ± 0.18 a | 8.03 ± 1.80 a | 4.74 ± 0.75 a | 5.76 ± 1.45 | 0.79 ± 0.15 | 0.28 ± 0.04 | |
| 5 | 1.49 ± 0.30 a | 7.36 ± 1.55 a | 10.90 ± 1.02 a,b | 5.38 ± 1.40 | 1.29 ± 0.10 | 0.72 ± 0.21 a,b | |
| 7 | 1.77 ± 0.24 a,b | 6.52 ± 1.70 a | 8.01 ± 1.80 a | 4.22 ± 0.80 | 0.89 ± 0.16 | 1.19 ± 0.20 a,b,c | |
| 14 | 2.60 ± 0.25 a,b,c,d | 7.48 ± 1.50 a | 7.82 ± 1.50 a | 17.10 ± 3.20 a,b,c,d | 7.43 ± 1.40 a,b,c,d | 0.99 ± 0.11 a,b | |
| 21 | 2.68 ± 0.25 a,b,c,d | 6.77 ± 1.40 a | 6.47 ± 1.60 a,c | 29.14 ± 6.20 a,b,c,d,e | 13.67 ± 3.10 a,b,c,d,e | 0.54 ± 0.10 a,d,e | |
| Qa23 | 0 (control) | 0.04 ± 0.01 | 0.34 ± 0.05 | 0.00 | 0.00 | 0.00 | 0.00 |
| 3 | 0.21 ± 0.03 | 7.26 ± 1.80 a | 0.87 ± 0.17 | 0.00 | 0.00 | 0.05 ± 0.002 | |
| 5 | 1.65 ± 0.15 a,b | 8.15 ± 1.90 a | 7.61 ± 1.40 a,b | 1.11 ± 0.25 | 0.31 ± 0.05 | 0.34 ± 0.04 a,b | |
| 7 | 1.63 ± 0.20 a,b | 6.40 ± 1.20 a | 6.14 ± 1.40 a,b | 6.33 ± 1.05 a,b,c | 1.72 ± 0.25 a,b | 1.24 ± 0.20 a,b,c | |
| 14 | 3.70 ± 0.70 a,b,c,d | 8.80 ± 1.90 a | 0.00 c,d | 1.03 ± 0.30 d | 0.34 ± 0.04 | 0.02 ± 0.001 c,d | |
| 21 | 3.16 ± 0.60 a,b,c,d | 6.47 ± 1.40 a | 5.98 ± 1.45 a,b,e | 23.63 ± 3.20 a,b,c,d,e | 13.64 ± 1.25 a,b,c,d,e | 0.85 ± 0.15 a,b,c,d,e | |
| Fermented Pomace (mg/KG) | |||||||
|---|---|---|---|---|---|---|---|
| Type of Vinification | Maceration Time | Gallic Acid | Syringic Acid | p-Coumaric Acid | Catechin | Epicatechin | Quercetin |
| SF | 0 (control) * | 0.53 ± 0.05 | 2.11 ± 0.32 | 0.00 | 2.98 ± 0.40 | 2.13 ± 0.30 | 0.77 ± 0.20 |
| 3 | 0.49 ± 0.04 | 3.14 ± 0.40 | 0.05 ± 0.01 a | 1.41 ± 0.14 | 0.84 ± 0.15 | 1.70 ± 0.25 | |
| 5 | 0.69 ± 0.12 | 3.90 ± 0.30 | 0.06 ± 0.01 a | 8.61 ± 1.90 a,b | 4.48 ± 0.80 a,b | 6.53 ± 1.80 a,b | |
| 7 | 0.67 ± 0.10 | 4.47 ± 0.90 | 0.05 ± 0.01 a | 4.62 ± 0.90 b,c | 1.84 ± 0.25 c | 9.16 ± 2.05 a,b | |
| 14 | 0.84 ± 0.15 a,b | 7.40 ± 1.40 a,b,c,d | 0.05 ± 0.011 a | 5.06 ± 1.00 b,c | 2.26 ± 0.60 b,c | 9.99 ± 2.00 a,b | |
| 21 | 0.98 ± 0.15 a,b,d | 9.12 ± 1.90 a,b,c,d | 0.06 ± 0.02 a | 4.51 ± 0.50 b,c | 2.20 ± 0.50 b,c | 10.40 ± 1.95 a,b | |
| BDX | 3 | 0.55 ± 0.05 | 3.28 ± 0.40 | 0.05 ± 0.01 a | 5.06 ± 1.50 | 2.78 ± 0.40 | 2.62 ± 0.40 |
| 5 | 0.62 ± 0.18 | 4.77 ± 0.80 | 0.05 ± 0.01 a | 6.83 ± 1.70 | 2.92 ± 0.80 | 7.41 ± 1.70 a,b | |
| 7 | 0.73 ± 0.14 | 6.03 ± 1.70 a | 0.04 ± 0.005 a | 4.43 ± 0.80 | 1.70 ± 0.30 | 7.07 ± 1.50 a,b | |
| 14 | 0.73 ± 0.15 | 6.63 ± 1.50 a | 0.04 ± 0.01 a | 4.27 ± 0.90 | 1.92 ± 0.30 | 10.46 ± 1.99 a,b | |
| 21 | 0.82 ± 0.20 | 8.53 ± 2.10 a,b,c | 0.05 ± 0.01 a | 6.73 ± 1.00 | 3.37 ± 0.40 d,e | 10.96 ± 1.40 a,b,d | |
| FX | 3 | 0.58 ± 0.05 | 3.24 ± 0.25 | 0.06 ± 0.01 a | 2.37 ± 0.69 | 1.01 ± 0.25 | 3.00 ± 0.75 |
| 5 | 0.57 ± 0.11 | 4.62 ± 0.40 a | 0.04 ± 0.002 a | 7.83 ± 1.23 a,b | 3.42 ± 0.90 a,b | 3.54 ± 0.57 | |
| 7 | 0.59 ± 0.12 | 5.25 ± 0.60 a,b | 0.04 ± 0.001 a | 4.45 ± 0.90 c | 1.95 ± 0.42 c | 9.64 ± 1.63 a,b,c | |
| 14 | 0.68 ± 0.09 | 6.16 ± 0.50 a,b | 0.05 ± 0.01 a | 2.37 ± 0.80 c | 1.06 ± 0.20 c | 10.23 ± 2.04 a,b,c | |
| 21 | 0.72 ± 0.22 | 9.26 ± 1.32 a,b,c,d,e | 0.06 ± 0.02 a | 3.61 ± 1.34 c | 1.94 ± 0.20 c | 12.17 ± 2.13 a,b,c | |
| Qa | 3 | 0.73 ± 0.15 | 4.83 ± 0.60 | 0.08 ± 0.02 a | 10.59 ± 2.14 a | 5.95 ± 0.87 a | 3.83 ± 0.30 a |
| 5 | 1.00 ± 0.12 a | 6.83 ± 0.55 a | 0.09 ± 0.02 a | 28.30 ± 3.58 a,b | 15.08 ± 2.13 a,b | 10.31 ± 1.09 a,b | |
| 7 | 0.88 ± 0.16 | 6.56 ± 0.83 a | 0.06 ± 0.01 a | 17.42 ± 2.19 a,b,c | 7.79 ± 1.54 a,c | 11.30 ± 1.11 a,b | |
| 14 | 0.75 ± 0.14 | 10.45 ± 2.01 a,b | 0.05 ± 0.002 a,c | 3.70 ± 0.78 b,c,d | 2.05 ± 0.36 b,c,d | 10.59 ± 0.90 a,b | |
| 21 | 0.66 ± 0.16 | 10.24 ± 2.63 a,b | 0.06 ± 0.01 a | 4.21 ± 0.69 b,c,d | 2.70 ± 0.30 b,c,d | 10.99 ± 1.45 a,b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisov, N.; Čakar, U.; Milenković, D.; Čebela, M.; Vuković, G.; Despotović, S.; Petrović, A. The Influence of Cabernet Sauvignon Ripeness, Healthy State and Maceration Time on Wine and Fermented Pomace Phenolic Profile. Fermentation 2023, 9, 695. https://doi.org/10.3390/fermentation9070695
Lisov N, Čakar U, Milenković D, Čebela M, Vuković G, Despotović S, Petrović A. The Influence of Cabernet Sauvignon Ripeness, Healthy State and Maceration Time on Wine and Fermented Pomace Phenolic Profile. Fermentation. 2023; 9(7):695. https://doi.org/10.3390/fermentation9070695
Chicago/Turabian StyleLisov, Nikolina, Uroš Čakar, Danijela Milenković, Maria Čebela, Gorica Vuković, Saša Despotović, and Aleksandar Petrović. 2023. "The Influence of Cabernet Sauvignon Ripeness, Healthy State and Maceration Time on Wine and Fermented Pomace Phenolic Profile" Fermentation 9, no. 7: 695. https://doi.org/10.3390/fermentation9070695
APA StyleLisov, N., Čakar, U., Milenković, D., Čebela, M., Vuković, G., Despotović, S., & Petrović, A. (2023). The Influence of Cabernet Sauvignon Ripeness, Healthy State and Maceration Time on Wine and Fermented Pomace Phenolic Profile. Fermentation, 9(7), 695. https://doi.org/10.3390/fermentation9070695