
The Health of Vineyard Soils: Towards a Sustainable Viticulture

1
Department of Ecology and Global Change, Desertification Research Centre-CIDE (CSIC, UVEG, GVA), Carretera CV-315, km 10.7, 46113 Moncada, Spain
2
Department of Applied Chemistry and Physics, Faculty of Biology and Environmental Sciences, Universidad de León, Campus de Vegazana, 24071 León, Spain
3
Research Institute of Vine and Wine, Universidad de León, Avenida de Portugal, 41, 24071 León, Spain
*
Authors to whom correspondence should be addressed.
Horticulturae 2024, 10(2), 154; https://doi.org/10.3390/horticulturae10020154
Submission received: 27 December 2023
/
Revised: 1 February 2024
/
Accepted: 1 February 2024
/
Published: 6 February 2024
(This article belongs to the Special Issue Sustainable Viticulture: Soil Fertility, Plant Nutrition and Grape Quality)
Abstract
:Soil health encompasses the effects the uppermost part of the land have on human wellbeing in a broad sense, because soil is where most food ultimately comes from, and because it more inconspicuously fulfils other ecological functions, as important as feeding, for our planet’s welfare, which is ours. Viticulture exploits the soil’s resources from which wine, its most valuable produce, boasts to obtain some of its unique quality traits, which are wrapped within the terroir concept. However, using conventional methods, viticulture also has harsh impacts on the soil, thus jeopardizing its sustainability. How long will the terroir expression remain unchanged as vineyard soil degradation goes on? While this question is difficult to answer because of the complex nature of terroirs, it is undeniable that conventional soil management practices in viticulture leave, in general, ample room for improvement, in their impact on vineyards as much as on the environment. In response, viticulture must adopt practices that enable the long-lasting preservation of its grounds for both on-farm and off-farm benefits. In this regard, the increase in the soil’s organic matter alongside the enhancement of the soil’s biological community are key because they benefit many other soil properties of a physical, chemical, and biological nature, thus determining the soil’s healthy functioning, where the vines may thrive for a long time, whereas its surroundings remain minimally disturbed. In the present review, the importance of soil health as it relates to vineyards is discussed, the soil degradation factors and processes that threaten winegrowing areas are presented, successful soil-health enhancement practices are shown, and future research trends are identified for the benefit of researchers and stakeholders in this special agricultural industry.
1. Introduction
Grapevine growing is an important agricultural activity, fueling an international trade in fresh grapes and wine worth, respectively, G€9.4 and G€37.8 [1]. However, agricultural revenues come at an expense in terms of depletion of natural resources [2], with viticulture standing out in this regard because of its need for the intensive use of machinery, tillage, and chemicals under conventional management conditions [3]. These conventional practices decrease porosity, aeration, aggregation, and the organic matter of soils and increase their toxicity to soil organisms, thus directly and indirectly damaging the soil’s biological community, and eventually decreasing the availability of water, oxygen, nutrients, and mutualists to vine roots.
Since soils are living systems, the soil degradation vineyards experience because of conventional practices may be described as a health loss. Degraded soils are nonhealthy soils and, consequently, are unable to satisfactorily meet their functions both on-farm and off-farm. First, through conventional running practices, viticulture jeopardizes the health of the base, from which its most valued produce, i.e., wine, boasts to obtain a good deal of its distinctive traits, i.e., terroir. Moreover, through conventional practices, viticulture contributes to the jeopardizing of environmental health, which is a growing concern for humanity.
Vineyard management practices that preserve the soil health, and even help it recover, have been studied and continue to be investigated. In this article, we present some of these studies to address the concept of soil health as it relates to the more classical concepts of soil quality and soil fertility (Section 2). Then, we show how biological fertility is at the core of the concept of soil health, since soil organisms are what make soils alive (Section 3). In this regard, soils are healthy as long as they harbor a diverse, balanced, and therefore functional biological community. Vineyard soils can benefit from such healthy associations of organisms due to their ability to work and rework the organic matter continuum that starts from the vegetative production of vines, as well as from the primary production due to accompanying plants. This reworking of the organic matter continuum releases inorganic compounds, which are plant-absorbed and transformed again into vine tissues. Moreover, in this cycling process, the soil’s biological community influences the vines through biochemical signaling and develops the soil organic matter (SOM).
As SOM is tackled (Section 4), it will be shown that it is on one side of a coin that has the soil’s biological activity on the other for vine nutrient supply. However, they also complement each other for soil structure development in such a way that SOM keeps upright what soil organisms build. Then, as the soil’s physical fertility in vineyards is presented (Section 5), the relevance of keeping a sound soil structure will be put under the spotlight regarding soil compaction avoidance, erosion control, and the soil’s water-holding capacity for vines. Unfortunately, what mostly threatens soil structure in vineyards, and therefore primarily menaces soil and water conservation therein, is tillage, which will therefore be given particular attention (Section 6).
The threat of tillage to soil’s physical fertility is particularly severe in sloping vinelands under a Mediterranean climate, which leaves the soil highly vulnerable to water erosion. As for the soil’s biological fertility, the toxicity of phytosanitary products is a direct menace (Section 7), which also indirectly threatens its physical and chemical counterparts. Phytosanitary products in ample use in vineyards, e.g., copper (Cu) compounds, are xenobiotic, and thus they are toxic to soil organisms. On the contrary, fertilizers and other amendments are usually not so toxic. However, synthetic fertilizers and amendments may change the diversity and balance of the soil’s biological community, thus disrupting its functionality and activity, with impairing effects on soil health (Section 8). Given all these damaging results of conventional vineyard management on soil health, one may ask how long terroir will remain unchanged as soil degrades, and conversely, one may ask how terroir will evolve as soil heals (Section 9).
Another pertinent question is how all the viticulture stakeholders can check the health status of vineyard soils to know if they are degrading or healing, and hence how to take action. There are several options for this task, from the most traditional to the most modern, which will be addressed (Section 10). Finally, some conclusions will be drawn (Section 11).
2. Soil Health, Soil Quality, and Soil Fertility
The concept of health has been traditionally applied to living beings, and particularly humans, for describing a state of complete physical, mental, and social well-being, which goes beyond the absence of disease or infirmity [4]. As beings that require air for breathing, water for drinking, and food for eating from our environment, and furthermore, that relate to each other, our health depends on the state of our surrounding atmosphere, hydrosphere, biosphere, and societal networks. Therefore, the state of these spheres may be understood in terms of health as long as they affect ours. At this point, the concept of environmental health naturally arises, and since the environment comprises four interdependent compartments, air, water, land, and biota (Figure 1), one may speak in terms of their respective health. However, it is not only appropriate to speak in terms of soil health due to its effects on human health, it is also so because soils are living systems on their own [5].
A healthy soil is one with a continued capacity to function within its ecological boundaries to sustain productivity, maintain environmental quality, and promote plant and animal health [6,7], including human health [8]. Therefore, soils are healthy if they provide enough plant support for crops to yield adequate amounts of safe and nourishing food, and for woods to produce timber and other raw materials. However, soils are also healthy if they meet their other ecological functions: the protection of humans and the environment and, moreover, the constitution of a gene reservoir [9]. In this regard, the protection of humans and the environment is provided by the soil’s capacities for (i) water cycle regulation through precipitation collection and its transmission underground, (ii) buffering of chemicals through filtering and transformation reactions, and (iii) weather and climate regulation through matter and energy exchanges with the other environmental compartments. Moreover, as a gene reservoir, soil also contributes to human health since it may house more biodiversity than the above-ground biota and, therefore, it is a gene source for facing present and future microbial diseases [10,11].
Quality concerning soil could be understood in terms of fitness for the purpose of producing adequate amounts of safe and nourishing food, as well as materials for the industry, i.e., in terms of fertility. This was the original meaning; however, it has evolved, and soil quality is now understood, along with air and water quality, as contributing to the multicomponent interrelated and holistic concept of environmental quality [12]. Therefore, soil quality cannot be understood only in terms of soil fertility, but in terms of balancing the biomass production function with the ecological functions. Consequently, soil quality has been defined as the fitness of a specific kind of soil, to function within its capacity and within natural or managed ecosystem boundaries, to sustain plant and animal productivity, maintain or enhance water and air quality, and to support human health and habitation [13].
Soils that satisfactorily fulfil all their ecological functions score highly in soil-quality and soil-health standards. Therefore, both concepts almost overlap. On the contrary, the soil fertility [14] is clearly more restricted, since it is a plant support-centered concept, because humanity directly and indirectly depends on terrestrial plants for over 98% of calorie intake [15] as well as for many raw materials (Figure 2). Nevertheless, it is crucial to acknowledge that the nutritional quality of agricultural crops is intricately linked to the overall quality and health of soils [15]. Human health and quality of life cannot progress any further within an environment that does not meet minimum health and quality standards. Among these, soil is key, because it continuously exchanges inert matter and living matter, as well as different forms of energy, with the other three environmental compartments [16,17,18]. The great similitude between soil-health and soil-quality concepts is more obvious within the framework of the “One World—One Health” paradigm in which the tight linkages between human, animal, and ecosystem health are recognized [14]. The burst of the COVID-19 pandemic taught us this lesson the hard way [19].
3. The Soil Biological Fertility of Vineyards
The traditional concept of soil fertility has focused primarily on the chemical and physical fertility, while the soil biological fertility has been often sidestepped. Fortunately, this is changing as agricultural and forest systems are seen from a holistic point of view. In this regard, Abbot and Murphy [20] provided a comprehensive definition of soil biological fertility that underscores the direct connection between the soil biological community and the plant nutrient supply, thus recognizing the linkage of the soil biology with the soil chemistry and physics. In doing so, they showed how the physical, chemical, and biological soil fertility approaches may integrate the concept of soil health. Therefore, soil’s biological fertility is understood as the capacity of the organisms residing in the soil to (i) directly contribute to the nutritional requirements of crops for productivity and harvest quality, and (ii) indirectly, but concurrently, to contribute to the maintenance of all the biological processes that have a positive impact on the soil’s chemical- and physical conditions with on-farm and off-farm enhancement effects. Unfortunately, our understanding of the soil’s biological fertility remains relatively limited in two critical aspects: (i) how to optimize the advantages derived from soil’s biological processes, and (ii) whether it is economically or environmentally sustainable to take advantage of the benefits of the various biological processes unfolding in soils [20].
Although the soil organisms constitute only a small fraction of the total soil weight, and even a small fraction of the SOM itself, they play numerous pivotal roles in soil systems [21]. The immense diversity of life forms we can find in soils is responsible for several critical processes that support soil functions. These processes include: on the one hand, (i) the decomposition of organic debris, (ii) the transformation of chemical compounds between different forms, (iii) the facilitation of plant-nutrient acquisition through symbiotic associations, and (iv) the production of soil-stabilizing agents that promote the aggregation of mineral particles, while, on the other hand, they also include (v) the degradation of synthetic organic chemicals like pesticides and herbicides, and (vi) the production of antibiotics that can aid in suppressing soil-borne diseases [20]. Consequently, soils with a functional biological community provide both a nurturing and a healing environment for crops and other agroecosystem plant-beneficial organisms, thus making the soil healthy. As our understanding of the soil biology improves, we will gain insight into how to take advantage of all these biological processes to enhance yields and incomes for producers, as well as environmental health for everyone [22].
Unfortunately, the intensification of agriculture contributes to soil biodiversity losses, with vineyard terrains being one of the most harmed because they are very intensively managed and erosion-prone [23,24]. In this context, Karimi et al. [25], in their meta-analysis of the ecotoxicological impact of viticultural practices on soil biodiversity, demonstrated that soil microorganisms are more abundant in organic viticulture compared to conventional viticulture. This is due to the detrimental effects associated with conventional vineyard management, which involves (i) frequent tillage and use of weed-killers to keep soils bare both below and between vines, (ii) the application of mineral fertilizers, and (iii) the use of several pesticides, particularly against fungal pathogens. The adverse impacts on overall soil biodiversity lead to degradation of its biological fertility, with harmful consequences for the ecological services soils provide [26]. On the contrary, conservation tillage, organic fertilization, and cover cropping, among other practices, may increase the diversity of the soil’s biological community and hence boost their associated functions for developing more sustainable vineyards [3]. Over time, viticulturists are increasingly adopting these conservation agriculture practices (CAPs). However, concerns persist for winegrowers, as these practices, particularly cover cropping, if improperly carried out, may increase water and nutrient stresses on vines markedly enough to harm, at least, grape production objectives [27].
Fostering a suitable soil habitat is paramount to enhance its biological fertility [20]. Through the application of different CAPs, viticulturists can modulate microbial activity inasmuch as they directly or indirectly provide the adequate physical and chemical conditions for the microbial populations to thrive (Table 1) [20]. In this respect, a sufficient supply of organic debris to the soil is the foundation, though not the only method, for enhancing its biological fertility. At least, to have a non-compacted soil is equally important since the soil-pore network plays a critical role in determining the spatial and temporal distribution of soil biota and nutrients, facilitating flow paths for solutes and gases [20]. In this regard, further research is needed to unravel the impact of soil management practices on the soil’s microbial diversity, and on changes in the evenness of the soil’s microbial community.
There is a growing body of evidence indicating that a comprehensive understanding of soil requires the integration of relevant physical and biochemical approaches [32]. Therefore, incorporating a pedological perspective into soil biology becomes essential for assessing the significance of soil’s biological processes and identifying fundamental principles that can be universally applied across various soil types and environments [20]. In the context of viticultural soils, the impact of soil biodiversity on soil functions, encompassing changes in both chemical and physical soil fertility, as well as ecosystem services, holds significant importance. Additionally, it is crucial to acknowledge that grapevines are typically cultivated as scions grafted onto rootstocks, which are selected based on the prevailing and expected soil and climate conditions [33]. Consequently, the molecular and microbial interactions between the scion and the soil via the rootstock must be considered in order to enhance the sustainability and resilience of viticulture [34].
3.1. The Impact of Soil Organisms on Chemical Fertility
Maintaining a suitable soil habitat by fostering the supply to the soil organisms of adequate quantities of organic materials of enough quality, is crucial for soil microbial communities to efficiently release nutrients in root-absorbable forms and, therefore, available to plants [20]. In this regard, rhizodepositions from vines, but also from other accompanying plants in the agroecosystem, both spontaneous and seeded as cover crops, should be considered as organic materials of the highest quality that can effectively contribute to increased soil carbon (C), and maybe also nitrogen (N) [35].
In soil agroecosystems, most nutrient transformations involve a multitude of biochemical reactions that are part of the biota metabolisms [36]. Pertaining to this, key microbial processes associated with soil nutrient cycling encompass (i) nutrient release through mineralization of organic compounds and solubilization of nutrient-containing minerals, e.g., Bettenfeld et al. [37] established that some plant growth-promoting bacteria species like Pseudomonas, Enterobacter, Arthrobacter, and Bacillus improve grapevine nutrition, at least in part, by increasing the vine-available phosphorus (P) through the transformation of organic P into soluble phosphate ions, (ii) nutrient enrichment through symbiotic and non-symbiotic N2 fixation, etc., (iii) nutrient transformations through nitrification of NH4+ to NO3−, reduction of Fe3+ to Fe2+ and MnO2 to Mn2+, oxidation of S and S2− to SO42−, etc., and (iv) nutrient losses through denitrification of NO3− to N2O and N2 [20]. Consequently, the activity of soil microorganisms plays a pivotal role in regulating the amount and availability to grapevines of nutrients such as phosphate and nitrate and hence it controls chemical fertility [38].
As indicated, the soil biota affects the concentrations of nutrients in available forms; however, the other way round, the soil nutrient richness leads to intricate metabolic interactions within microbial communities, resulting in changes in the identity and relative abundances of soil taxa [39]. In this regard, it seems that P is the nutrient that mostly affects soil microbiome traits, probably because N is not applied in vineyards at high enough rates to clearly show up [40].
Since vineyard fertilization is complex and highly dependent on plant material, soil characteristics, and the production objectives [41], enhancement of grapevine nutrition by microorganisms is an interesting line of viticulture research.
3.2. The Impact of Soil Organisms on Physical Fertility
The profound impact soil fauna activities have on the soil’s physical properties include (i) excavation for food or living spaces, (ii) ingestion of soil materials, (iii) production of fecal pellets, (iv) utilization of excreta, mucus, or salivary secretions, and (v) collection of plant, animal, and fungal debris mixing with the soil mineral particles, with or without prior digestion [20]. The combined activities of soil fauna with those of plant roots, besides fungi and bacteria, result in the formation of biological aggregates, to which earthworms particularly contribute [42].
The soil aggregates thus formed serve as microhabitats that support the proliferation of soil microorganisms. These microorganisms further contribute to soil aggregate formation by producing specific exudates and by releasing lysates when they die [43]. While soil fauna strongly affects the distribution and organization of soil particles, the microbiota crucially contributes to their soil-structuring works through several direct and indirect processes. These include: (i) moving and aligning primary particles along cell or hyphal surfaces, (ii) adhering particles together through sticking compounds, e.g., exudates, involved in colony cohesion, (iii) enmeshing and binding aggregates through fungal hyphae, actinomycete filaments, and associated mycelia, and (iv) coating pore walls with hydrophobic compounds [36]. Bacteria contribute to aggregate formation by producing negatively charged polysaccharides, and polyuronic and amino acids that stick to soil particles with the aid of cations. Fungi, including saprophytic, ectomycorrhizal, and arbuscular mycorrhizal, also play essential roles in forming soil aggregates through physical mechanisms such as the formation of hyphae and mycelia that entangle soil particles, and through physicochemical mechanisms that glue soil particles together by means of exudates, which include polysaccharides like glomalin, mucilages, hydrophobins, and extracellular compounds from hyphae [43].
3.3. Key Interactions in the Rhizosphere on Soil Biological Fertility
The rhizosphere is the soil region closer to the plant roots and is, therefore, greatly influenced by the rhizodepositions, which encompass exudates, as well as detached root tissues and cells and the lysates they release when break up [44,45]. Accordingly, the rhizosphere is the primary interface for interaction between a plant and the soil environment [46], possessing distinct biological, physical, and chemical characteristics compared to the bulk soil (Table 2). The rizhosphere high contents of labile organic materials boost the microbial populations, which are jointly referred to as the rhizomicrobiome [45]. The rhizomicrobiome is particularly important in regulating plant growth and health, and makes the rhizosphere to become the most dynamic part of the soil, where multitude of biogeochemical processes take place. A comprehensive understanding of these processes is vital for sustaining the microorganisms that inhabit soils and preserve soil health [47].
The nature of the soil, environmental conditions, and plant genotype, particularly the characteristics of its root rhizodepositions, determine the microbial diversity of the rhizosphere [45]. It is quite likely that plants and their associated rhizomicrobiome have coevolved in natural ecosystems, leading to the mutual adaptation of each other for survival [54]. Furthermore, the availability of plant nutrients in the rhizosphere is influenced by a combination of soil properties, plant characteristics, and the interactions between plant roots and the rhizomicrobiome [20]. As a result, the rhizosphere and the characteristics of the rhizomicrobiome feature a delicate balance between stress and fitness, and could serve as valuable biological indicators of plant health status [34] and, therefore, soil health.
The interaction between the vine roots and the soil microbiota is essential for the control of soil phytopathogens and for the acquisition of the nutrients required for growth, and the rhizosphere is at the core of these interactions [34]. Indeed, the richness and diversity of bacterial and fungal communities in the root rhizosphere are significantly influenced by the grape variety [55], and also by the rootstock genotype as an important driver of the bacterial community composition [56]. Therefore, a new research approach named rhizosphere engineering is increasingly coming into focus to enhance grapevine growth and health, in which microbial communities are modified by adding specific fertilizers, nutrients or by bio-inoculation with certain bacteria and/or fungi, suggesting a potential for reaching a more sustainable development of pesticide-reduced viticulture in the future [57]. Another potential area of future research would be to investigate the grapevine rhizosphere as a microbial source of inoculum of grape berry microbiota since a hypothetical endophytic way of colonization has been proposed for the fermentative yeast Saccharomyces cerevisiae to be transported from the soil via roots and xylem [34].
3.4. The Impact of Mycorrhizae on Soil Health
The rhizomicrobiome of plants colonized by mycorrhizae exhibits significant differences compared to that of non-colonized plants, and thus deserves some depth of comment. Mycorrhizal plants have been shown to have higher proportions of organic acids, including amino acids, in their roots in contrast to non-mycorrhizal plants [20]. Mycorrhizal fungi are considered effective candidates for biofertilization programs due to several advantageous features, making them a valuable resource for soil health in the context of sustainable agriculture. These advantages include their ubiquity in nature, ease of use and production as carrier-based inoculants with a long shelf life, capacity to increase the surface area of plant roots, phosphate solubilization capability, improvement of micronutrient uptake, assistance in plant stress tolerance, and mitigation of biotic stress [58].
Arbuscular mycorrhizal fungi (AMF) benefits start from their ability to extend the volume of soil explored by grapevines, allowing an adequate uptake of nutrients such as N and P [41]. The fungal mycelium increases the exploitable soil volume, allowing access to soil pores that would be otherwise non-explorable by root hairs. Regarding the grapevine uptake of other nutrients, Schreiner [59] showed an enhanced Cu and sulfur (S) uptake by AMF-inoculated grapevine cuttings from two different soils, and suggested that the selection of appropriate inocula for specific functions would be the best strategy in biofertilizer procedures. Furthermore, in grapevines grafted onto rootstocks that are susceptible to iron chlorosis, AMF inoculation may be a way to alleviate its symptoms on calcareous soils, but further studies are needed [41]. Interestingly, the modulating effect of mycorrhization on vine nutrient absorption rate and selectivity seem to unfold through its effect on the bacterial microbiota in the root endosphere, where the populations of several beneficial taxa increase. Additionally, grapevines gain enhanced protection against pathogens when colonized by AMF, although the safeguarding effects of AMF may not apply uniformly to all pathogens [60] and should be further investigated.
In addition to vine nutrition and protection, as well as soil aggregation, AMF have other benefits for vines. In this regard, Gazioglu [61] also showed how AMF can mitigate the physiological and morphological changes induced by salt stress in grapevine cuttings, while Cardinale et al. [62] showed how mycorrhization can mitigate the impact of drought stress on plant survival and growth. The drought-ameliorating effect of AMF on grapevines seems to develop through various mechanisms, including higher osmolyte build-up and improved photosynthetic activity [41]. Therefore, AMF could form the basis of an effective management strategy to counteract drought effects on grapevines, and thus further investigations in this regard are of the utmost importance for enhancing many vineyards, particularly under Mediterranean and related semi-arid conditions, which are predicted to become dryer under the climate change scenario.
Counteracting the beneficial effects of AMF on crop plants is tillage, because this practice seems to effectively break up the mycelia networks, as the results of Lumini et al. [63] suggest. Indeed, these authors found lower AMF biodiversity in a tilled vineyard soil regarding another covered with spontaneous plants. In spite of the above, the question arises whether shallow tillage can affect AMF communities which grow relatively deeply in the soil [41]. Since vines’ roots can remarkably deepen, more research is needed in this regard.
To sum up, given the crucial role AMFs are likely to play in alleviating abiotic and biotic stress in grapevines, more studies are needed to identify the diversity of AMF species already present in vineyards across different edaphoclimatic conditions [60] and how their beneficial effects may be enhanced.
4. Soil Organic Matter as a Key Property of Soil Health in Vineyards
Soil biological activity and SOM are strongly interrelated and heavily depend on one another. The SOM comprises the part of the soil condensed material that features carbon in formal oxidation states under four (zC < +4). The SOM has gone through one to several cycles of biosynthesis that started from the plant or bacterial assimilation of simple inorganic compounds such as CO2, H2O, N2, HPO42−, and SO42−, and is eventually headed to release the same inorganic compounds to the environment [64,65]. The SOM often constitutes a little share of the soil condensed material featuring a worldwide average of 6.5% on a dry-weight base in the top 10 cm [66]. However, an ample range of SOM between 0.2 and 60% may be estimated for the topsoil, depending on soil and climate conditions [67]. Despite the general small portion the SOM represents regarding the soil’s minerals, the changes in SOM remarkably affect most soil properties, from physical to chemical and biological [68,69]. Additionally, despite its low share of soil weight, SOM down to 1 m depth also stores 1800 Gt of C, i.e., more than the atmosphere (570 Gt) and biota (740 Gt) combined [70,71], which might increase by 20–50 Gt if best CAPs were globally adopted [72]. Therefore, the SOM is key for soils to be healthy through at least (i) the maintenance and enhancement of the soil fertility and hence sustainability of cropping and farming systems; and (ii) the regulation of climate [73].
4.1. Soil Organic Matter and Soil-Health Attributes
From a physical perspective, SOM significantly increases the aggregation of the soil’s mineral particles [74,75,76,77]. Indeed, as the SOM increases, soil particles define a more organized and interconnected network of pores of variable sizes featuring, in general, lower bulk density [78]. This soil pore space is built and rebuilt time and again by plant roots [79,80] and soil fauna and fungi [81,82,83]. However, this dynamic soil pore space is kept from collapsing by the SOM ability to increase the magnitude of the non-directional Van der Waals attractive forces among the soil particles [84,85]. Through such an organized systems of pores, gases can continually diffuse in and out of the soil [86,87,88], water can infiltrate [89,90], and, moreover, be held in part against gravity and evaporation, and thus be kept available for plants and soil organisms [91].
From a chemical perspective, SOM is a reservoir of many essential plant nutrients. Indeed, as the SOM increases, the soil contents of the macronutrients N, P, and S, generally rise [92,93,94]. However, since N, P and S are covalent-bonded in the SOM in many different compounds [95,96,97], their availability to plants is constrained, and their release in the soil environment as plant-absorbable species is mediated by the soil organisms’ metabolism [94] as previously addressed (Section 3). This condition contrasts with that of the other macronutrients, i.e., potassium (K), magnesium (Mg), and calcium (Ca), whose plant-available contents increase more readily with SOM. This is because SOM is negatively charged at even the lowest soil pH values, whereas K, Mg, and Ca are always found as cations in the soil environment, i.e., K+, Mg2+ and Ca2+, and thus SOM is able to hold them readily exchangeable by other cations, including root-exudated protons [98,99].
Regarding the micronutrients, as the SOM increases, their availability may increase or decrease depending on the elements’ redox properties as well as on soil aeration, pH, and the fraction of SOM that is water-soluble [100]. Specifically, iron (Fe) is a nutrient whose available soil concentration seems to increase in general with SOM, because SOM decreases the soil redox potential (Eh), thus fostering the reduction of the (oxyhidr)oxide-immobilized Fe(III) to the water-soluble Fe(II) species [101,102,103]. Additionally, SOM also provides low-weight water-soluble organic ligands that further stabilize Fe(II) in solution and keep them plant-available [102,104]. For manganese (Mn), the same processes have been described [102] and, in general, it seems that Mn availability increases as SOM rises, but the effect seems weaker than with Fe [100]. Finally, zinc (Zn), Cu, boron (B), and molybdenum (Mo) present, in general, poorer redox chemistry in the soil environment [102]. Therefore, the availability of Zn, Cu, B, and Mo is not so controlled by the low Eh the SOM may foster, but rather by SOM adsorption abilities. Consequently, since SOM also forms surface complexes with Zn, Cu, B, and Mo, in general SOM decreases the availability of these elements to plants [102,105,106,107], which would thus be released at the pace the SOM is mineralized.
From a chemical perspective, in addition to behaving as a nutrient reservoir, SOM also behaves as a pollutant buffer by adsorbing xenobiotic organic compounds (XOC) [108,109,110]. In addition to adsorption, SOM may also foster XOC degradation through the enhancement of the activity of the soil organisms, hence behaving as a factor in environmental remediation [111]. In the case of heavy metals, the effects of SOM may or may not be beneficial, since SOM can immobilize metals as surface organic complexes, but it can also render them more available in the soil-pore water following reduction to more soluble chemical species and complexation with low-weight organic ligands [112,113]. Nevertheless, in general, upon passing through soil, the volatilization and, importantly, the leaching and plant uptake of many harmful chemicals is slowed down or even avoided if degradation is feasible and enough time passes by.
From a biological perspective, SOM is strongly and positively related to the functionality of the soil biological community. Indeed, as the SOM content increases, the abundance, activity, diversity, food web structure, and stability of the soil biological community are all enhanced [114,115,116]. A functional biological community is what distinguishes a healthy soil [6]. A functional biological community builds and rebuilds the soil’s pore space [81]. In doing this, the soil organisms feed on the plant debris, mostly dead roots, which are the primary source of SOM [117], resynthesize its compounds time and again, and eventually release and mix its remains with the mineral particles. The mixture of SOM with the mineral particles presents more cohesion than the separated organic or mineral particles. Therefore, such an arrangement of organic and inorganic materials is able to hold up the soil pore space against several forces that may act upon it: disruption by wetting, impact, overburden, shear, etc., thus showing in the increase in the stability of soil aggregates with SOM [118,119,120]. Moreover, as the organisms feed on SOM, some nutrients, which were bonded to it, are released for the benefit of other soil organisms and plants. In addition, some pollutants, remarkably XOCs, may be degraded by the organisms’ metabolism, or tightly adsorbed into the organic–mineral assemblages [111], and hence their volatilization, leaching, and plant uptake are diminished.
As presented, SOM is key to assure the ability of soils to sustain (i) plant growth and development, (ii) the biogeochemical cycles of water, as well as C, oxygen, N, etc., and (iii) the habitat structure of the soil biological community. The durable running of these ecological functions makes soils able to indefinitely keep the productivity and health-fostering properties of biological systems due to the quality preservation of agricultural produce, as well as of water and air, thus ultimately promoting the animal and human welfare [121,122].
4.2. Dynamics of Winegrowing Systems and Levels of Soil Organic Matter
Since the Neolithic age, the transformation of lands to agriculture has been accompanied by remarkable losses of SOM, whose C has been eventually oxidized to CO2 and released to the atmosphere [72]. In the case of viticultural systems, the SOM depletion has been especially severe [123,124] because vineyards have been overall traditionally developed on sloping lands subjected to sustained conventional tillage, thus becoming highly erodible [24,125,126,127,128]. Indeed, in areas under the Mediterranean climate, i.e., where a good deal of the World’s wine growing takes place, soil water erosion in vineyards attains high rates [129] due to the Mediterranean-characteristic intense rainfalls [130,131], evident in remarkable topsoil thinning [123] and even soil truncation [132]. Since SOM is usually highest in the topsoil and exponentially decreases with depth [133,134,135], topsoil thinning and soil truncation mean a severe loss of SOM. Therefore, it is not surprising that SOM levels in vineyards have decreased much more than under other land uses, including similar agricultural ones [123], and globally are estimated to range between 0.1% and 6.3% with a mean of 1.7% [136]. Such SOM levels are considered low in general for balanced textured soils [137], and therefore they jeopardize soil health in viticulture [3,138].
Water erosion causes SOM losses throughout the World [139], and in turn low SOM levels may increase water erosion rates because of the loss of topsoil aggregate stability [140]. Therefore, SOM loss and water erosion feed back to each other. As a way to counteract the harms to soil health in vineyards driven by SOM loss, several CAPs have been proposed [3,141]. The CAPs act by reducing the soil erosion and, consequently, by reducing or even reversing SOM loss, or by directly increasing the SOM content. The CAPs used in viticulture can be classified into the following four types: (i) soil mulching with organic materials like pruning residues and cereal straws, (ii) soil incorporation of foreign organic matters like composts, vermicomposts, biochars, and manures, (iii) soil inoculation of beneficial organisms like AMF, and (iv) the use of cover crops in vineyard alleys.
Straw mulches can foster SOM increase if timely applied, as well as adequately rated and replaced as indicated by Gómez de Barreda et al. [142], who found in a flat Eastern Spain vineyard that straw mulching increased SOM in the 0–12 cm layer up to 2.3%, i.e., a +32% increase as compared to bare soil. Interestingly, the average SOM increase during the three years of application was a linear function of the straw rate up to, at least, the maximum they tested, which was 50 Mg ha−1. Similarly, Qin et al. [143] found that, in a flat Eastern China vineyard, straw mulching applied at rates of between 7 and 21 Mg ha−1 significantly increased SOM in the 0–20 cm layer from 1.6 to 1.8%, i.e., a +10% increase as compared to bare soil. However, the linear increment of SOM with the straw rate was observed only for rates of 14 Mg ha−1 onwards during the first year of application, thus apparently remaining steady, and independent of the straw rate during the second one.
The application of organic amendments is another widely adopted method to enhance SOM in vineyards [144,145]. In this regard, Morlat and Chaussod [144] found in a flat Central France vineyard that SOM significantly increased up to almost saturation in the 0–30 cm layer from 1.5 to 2.2%, i.e., a +50% increase, in 22 years since the start of cattle manure and mushroom compost additions, at rates of, respectively, 20 Mg ha−1 yr−1 and 16 Mg ha−1 yr−1 (in terms of fresh weight). Besides, Gaiotti et al. [145] found in another flat NE Italy vineyard that SOM significantly increased in the 0–50 cm layer from 2.3 to 2.6% and from 2.3 to 2.9%, i.e., +14% and +26% increases, in 5 years since the starting of additions of, respectively, pruning wastes and cattle manure, at rates of 4 Mg ha−1 yr−1.
The inoculation of beneficial organisms in vineyards offers a promising method for SOM enhancement. Particularly, mycorrhizal fungi improve the relationships between vines and soil [146] with enhancing consequences on photosynthetic activity and C assimilation [147]. The allocation of vine photosynthates to the mycorrhizae, as well as their dampening of the rhizosphere-priming effect on SOM decomposition, might additionally contribute to higher SOM levels in vineyards [148]. However, there is limited knowledge of net changes in SOM levels as a consequence of plant mycorrhization in actual cropping conditions. Therefore, to what extent mycorrhizae may contribute to increase SOM in vineyards, if any at all, should be further investigated [149].
The use of cover crops has the potential to improve soil health by enhancing SOM and its dynamics through soil protection and the boosting of the functionality of the soil’s biological community [150]. Ruíz-Colmenero et al. [151] found, in a Central Spain vineyard, decreases of mean soil loss of 78% and 87% regarding traditional tillage due to two grass treatments, the former mown and the latter permanent. The soil conservation gains under both treatments were accompanied by a significant build-up of SOM from 1.2 to 1.6%, i.e., a +28% increase, in the 0–10 cm layer, after four years of cover crop establishment. Furthermore, Guzmán et al. [149] also found in a survey of 16 vineyards in a hilly area in Southern Spain that cover crops, either spontaneous or sown, significantly increased SOM in the 0–10 cm layer from 1.4 to 1.8%, i.e., a +29% increase, as compared to bare soil, after three years of cover crop establishment. Away from the sloping Mediterranean vineyards, Fleishman et al. [152] found in Pennsylvania a significant increase in SOM from 3.8 to 5.2%, i.e., a +37% increase, in the 0–20 cm layer, after four years of cover crop establishment [153,154].
The beneficial effect of cover crops for SOM enhancement regardless of vineyard slope is that cover crops are a source of fresh organic matter in the form or rhizodepositions [155,156]. However, the use of cover crops under arid to dry subhumid climates presents some drawbacks, since they decrease the availability of soil water for vines [157], thus generally increasing the water stress upon them. Conversely, mulches diminish the water stress on vines [158]. The effects of cover crops on vine water status under such dry climate conditions have consequences on vine vigor and also on grape yields under rain-fed water regimes [157]. However, the yield losses may be made up for by the increase in the grape must quality for winemaking [159]. The use of one management practice or the other to increase SOM levels, whereas preserving grape yields and quality, depends on the soil and climate conditions. The effects of all these CAPs on SOM, as well as on vine water status, and ultimately grape yield and quality, with the specific aim of increasing this latter, i.e., quality, should be further investigated.
5. Soil Physical Fertility in Vineyards
Soil organic matter affects soil’s physical properties, and a good physical condition of soil enhances SOM. A viticultural soil is considered to have good physical quality when it possesses the strength to not collapse against several forces that may act upon it, thus maintaining a sound structure that facilitates unhindered root growth, as well as the burrowing and settlement of soil fauna. Additionally, such a soil exhibits good water-transmission capacities and water-holding characteristics that allow for the proper distribution and storage of incoming rainfall and irrigation, its solutes, and gases. These qualities are essential both for optimal crop performance and for minimizing environmental degradation [160].
Maintaining soil structure is crucial for soil health as it governs the soil-water cycle and provides the medium for plant roots to anchor, respire and absorb water and nutrients [161]. Moreover, soil structure plays a pivotal role in regulating the physicochemical heterogeneity of the soil, thus influencing the distribution of microbial communities. The heterogeneity of soil aggregates promotes diverse biological niches with varying physicochemical properties, i.e., variation in aeration and hence Eh, pH, etc., thus microbial activities across different aggregate sizes differ [162]. In this context, biological activity plays a crucial role in resisting the physical degradation of soil since it importantly contributes to the formation and preservation of soil aggregates and pores, and thus soil structure [161].
Given that soil structure, as an abiotic factor, significantly influences microbial communities by creating distinct niches that support specific microbial adaptations, the interaction between soil structure and microbes emerges as a decisive factor shaping soil microbial diversity and, consequently, soil health [39]. It is evident, therefore, that soils suitable for grape production should possess well-developed structural integrity, stability against water and external mechanical stresses, and resilience, enabling them to recover their structure following disturbances [163].
5.1. Soil Compaction, Water Infiltration, and Aeration
Soil compaction, a significant concern for intensively used agricultural soils, is a physical process resulting in increased soil bulk density, and decreased soil porosity [164]. The ecological and economical damages associated to soil compaction are issues of global importance [165], which are particularly insidious in modern mechanized agricultural exploitations, particularly where soils are low in SOM, and high in clay and/or silt [166]. Unfortunately, soil compaction issues are anticipated to escalate due to the worsening weather patterns expected under climate change, including higher average temperatures and extended dry periods [167]. The effects due to higher temperatures are driven by the expected SOM losses as the climate warms up [168]. The effects due to longer droughts arise because the drier the soil, the more severe the compaction consequences on water infiltration, root development, etc. [169].
Soil compaction not only decreases porosity, it also changes pore length, average diameter, and tortuosity, thus adversely impacting ground penetrability by roots and soil fauna, as well as water infiltration, holding capacity, percolation, and drainage [166]. As a consequence, soil ecological, hydrological, and agronomic functions are harmed because of the following effects: (i) reduced overall plant productivity due to lack of water and nutrient availability to roots or either, because of water logging and scarce aeration; (ii) increased risk and severity of flooding and erosion due to decreased infiltration and increased surface runoff; (iii) elevated emissions of greenhouse gases like N2O and CH4 resulting from reduced soil aeration, but also more CO2 resulting from increased fuel consumption for tillage; (iv) decreased fertilizer efficiency due to the constrained mobility of their soluble species along with water; and (v) diminished rhizodepositions and hence less C input from plant roots due to heightened soil mechanical resistance to root penetration and impaired aeration [167].
The impact of compaction on the soil microbiome structure and activity varies considerably across different textures, compaction levels, and water contents. However, in general, the alterations in the soil-pore system characteristics induced by compaction create less favorable conditions for beneficial microorganisms [170]. Increasing soil compaction, and hence rising bulk density, constrains gas and water fluxes [170,171], eventually diminishing air-filled porosity, i.e., soil aeration. Anaerobic soil conditions lead to changes in the microbial community, causing most aerobic organisms to become quiescent or perish [172]. This favors microbial populations capable of tolerating anaerobic conditions. These involve both facultative and obligate anaerobic microorganisms including Fe- and sulfate-reducing microorganisms, which develop at the expense of fungal populations [162,172]. Moreover, under such low-aeration conditions soil C and N dynamics change and SOM concentration and availability of soil N also changes [173] thus impacting soil chemical fertility and plant nutrition.
5.2. Soil Erosion
Soil erosion by water is a critical threat to soil resources, particularly in Mediterranean regions, due to their specific climatic and geomorphologic conditions. Soil erosion rates in vineyards are higher than in other land uses due to both natural and anthropogenic influences [24]. Natural factors include rainfall intensity, slope steepness, rock fragment cover, and soil texture. Specifically, slope steepness, which characterizes many vineyards under the Mediterranean climate, is the factor that emerges, along with the late-summer and early-autumn intense rainfalls, as a key determinant of water-induced soil loss [126]. Indeed, soil erosion dramatically materializes during the intense rainfalls that feature the Mediterranean wet season’s inception, when plant cover is naturally at its minimum following the characteristic summer drought. Soil erosion is furthermore exacerbated in vineyard soils due to the use of conventional soil-management practices, including harsh tillage operations [174,175,176], frequent machinery traffic, and herbicide applications to get rid of inter-row and below-vine vegetation. Moreover, often young vineyards are particularly vulnerable, and therefore they experience unacceptably large soil erosion rates [177].
The conventional vineyard soil management practices significantly elevate erosion rates, potentially leading to severe and possibly irreversible situations [123,132]. For instance, a study in the conventionally managed steep Prosecco wine-producing area in Italy estimated erosion rates as high as 44 Mg ha−1 yr−1, which exceeds 31 times the maximum soil erosion threshold established for Europe [178]. Therefore, conventional vine management practices, particularly in areas with steep slopes, shallow soils, and many years of ploughing, such as the Prosecco area, negatively impact soil health. Herbicide application, which is used as an alternative to tillage for weed control in vineyards, possibly reduces erosion regarding tillage, but maintains it at non-negligible rates [179]. High soil-erosion rates in vineyards lead to increased nutrient losses [180] and less soil water availability and, therefore, to less vine vigor [181], as well as lower grape yield and berry quality, featuring higher total soluble solids and pH [182]. To counteract this soil fertility decline, viticulturists resort to greater external input use, ultimately compromising viticultural sustainability [181]. Climate change, with its higher-intensity rainfall events and interspersed droughts, threatens to increase soil erosion rates [183]. Therefore, encouraging viticultural practices that enhance soil and water conservation becomes increasingly important in this scenario.
Improved soil management practices aimed at increasing soil cover and SOM levels can bolster viticultural soil resistance to erosion, thereby improving their health [161]. To achieve sustainability in vineyards prone to soil erosion, it is imperative to adopt soil and water conservation practices. Particularly, terracing and the use of inter-row cover cropping can help. Terracing is a traditional practice that stands out as one of the most effective for erosion control, effectively protecting farms from soil loss. For instance, a study conducted by Pijl et al. [183] in 50 vineyards in Northern Italy found that three terracing methods (contour terracing, broad-base terracing, and oblique terracing) presented higher sediment flux mitigation ability than non-terraced practices. This was especially notable considering that terraced vineyards often occupy steeper slopes. Among the terracing methods, regular contour terracing consistently outperformed the others in terms of reducing uphill erosion, sediment flux, and downhill deposition. In spite of these beneficial effects, ineffective terrace establishment and management can lead to declines in soil fertility, as well as slope instability issues that may eventually result in terrace abandonment, amplified soil degradation, and even hydrogeological risks [184].
In addition to terracing, which is an engineering practice that tries to hold the soil in place by physical containment, cover crops can also be used to decrease soil erosion [184,185] and to improve water-holding capacity and overall soil condition, hence reducing reliance on industrial fertilizers [151,176,186,187], although not as effectively as terracing [188]. Pappalardo et al. [178] estimated that by using a 100% ground-covering inter-row cover crop, the erosion in the Prosecco wine-growing area could be curtailed by 67%. More specifically, a six-year experiment conducted by Stanchi et al. [179] in an Italian sloping vineyard revealed that erosion rates were almost negligible under permanent grass cover, whereas tractor passage significantly accelerated erosion showing up in an increased downhill accumulation of sediments high in SOM, N, and clay. These findings strongly point towards the conservation of soil fertility and health which results from the use of cover crops in vineyards.
In addition to behaving as a physical shield against raindrop impacts, the positive effects of vegetation covers that contribute to soil erosion control include increased aggregate stability and water infiltration rates because of the facilitation of soil aggregate formation [186], and enhancement of soil porosity due to gains in SOM and biological activity [126]. However, the use of cover crops in vineyards under dry climates may limit grape yields [187]. Notwithstanding, inter-row cover cropping in vineyards may also help improve grape quality, as Pou et al. [189] found. By precisely selecting the cover crop species and the time of sowing and reaping with regards to the local climate and vine phenology, the drawbacks of this soil management practice can be minimized and their benefits maximized in vineyards under arid to dry-subhumid climates [157].
Another soil management practice for soil conservation against water erosion in vineyards is the use of mulches. In this regard, Prosdocimi et al. [176] found in Eastern Spain vineyards an 80% decrease in median soil loss due to the protection effect of barley straw mulch rated at 750 kg ha−1. Furthermore, Mirás-Avalos et al. [190] found in a Western Spain vineyard a 57% decrease in average soil loss as a consequence of the protection effect of a mixture of straw and gorse residues. However, for soil erosion control, the issue with mulches, and straw mulching in particular, is the ease with which they may be dragged downhill by wind, e.g., by the mountain breeze, or rainfall.
6. Tillage and Soil Health in Vineyards
In agriculture, tillage has traditionally served as the primary means for weed control, playing a pivotal role in getting crops rid from the competition for water and nutrients of these “plants in the wrong place”. Therefore, tillage has been considered as an important means for achieving high crop production and economic profitability. In addition to weed control, tillage is usually viewed by agriculturists as an essential means for attaining several cultural targets, such as (i) grinding and soil mixing of leaves, pruning wood and other organic materials, (ii) promotion of water infiltration, (iii) boosting of SOM mineralization and nutrient release in plant available forms, (iv) thorough mixing of nutrients in the soil, (v) disruption of the habitats and the life cycle of soil-borne pests, and (vi) breakup of soil compacted layers that constrain root penetration and water movement throughout the soil depth [191].
Therefore, tillage has been considered as promoting several soil functions beneficial for cropping, i.e., the soil fertility in the short term. However, tillage comes with built-in disadvantages, as the soil disturbance it causes affects soil properties essential for the agroecosystem functioning in the mid- to long-run [192]. Regarding the soil physical fertility, tillage disrupts aggregates and ruins soil structure, paradoxically fostering the appearance of compacted soil layers [193], whose breakup demands more tillage operations, which are deeper and of higher intensity to make up for the low physical fertility status that results [194]. Tillage also exposes the soil to the rainfall drop impacts as well as to the runoff, hence increasing the soil vulnerability to water erosion. Moreover, although right after tillage the water infiltration capacity in vineyards may increase [158], tillage eventually eases water evaporation from soils [195], thus likely contributing to vine water-stress.
Tillage also influences soil chemical fertility but often not exactly in the way it was intended. Notably, it has been observed that topsoil total N, P, and K concentrations decrease, particularly under the harsher tillage operations in comparison to no tillage, as a meta-analysis by Nunes et al. has shown [196]. This effect on the primary nutrients is explained because tillage expedites SOM oxidation, thus boosting nutrient release in plant-available forms, which are, nevertheless, also more easily lost by leaching and gas emission [197] whether the adequate conditions are met, which it seems the most likely [196].
Regarding the soil’s biological fertility, the more intensive and frequent the tillage operations, the higher the destruction of the fungal hyphae and the soil fauna habitats. As a consequence, tillage eventually favors the development of bacterial communities over eukaryotic soil organisms, thus leading to imbalance in the soil biome [20] with harmful consequences on the functionality of the soil’s biological community and, consequently, on soil health. Conversely, minimum cultivation practices, like reduced tillage, decrease the destruction of soil fungi and fauna. Besides, minimum cultivation practices also result in less surface area contact between organic residues and soil microbial communities, which further boosts fungi development [20] and hence the balance among soil biological communities. As a consequence of all this, reducing tillage intensity significantly increases soil organic C (SOC) content and enhances biological activity in the topsoil and at greater soil depths, as shown by Nunes et al. [198] for US agricultural soils. Therefore, seeking more sustainable alternatives to tillage, such as cover cropping or mulching, not only decreases soil erosion but also promotes soil biodiversity. Moreover, in vineyard soils, these CAPs increase SOM content, enhance soil porosity, improve structural stability, and increase water holding capacity [23].
6.1. How Agricultural Traffic and Tillage Affect Soil Compaction
Tillage alters the physical characteristics of soil, significantly affecting its aeration, nutrient levels, and biological activity, but also water-holding capacity and hence temperature regulation. The effects on soil water content and flow are closely linked to the resulting compaction [199] caused by the forces exerted by the tractor wheels and tillage equipment, especially when they operate under a particular range of intermediate soil water contents, which render the soil highly susceptible to deformation [200,201]. Almost all types of tires result in increased soil compaction within the wheel tracks, although there may be differences depending on tire pressure [202]. Fortunately, as distance increases from the wheel tracks, soil compaction rapidly decreases, particularly in the subsoil [203].
Specifically for vineyards, studies have shown that subsoil compaction is predominantly caused by the machinery wheel load and not by the tillage operations themselves [204]. In addition to the soil condition during the pass, i.e., mainly the soil water content, factors such as the tire pressure and the number of passes over the same track are the primary drivers of local soil compaction [205]. Interestingly, high tire-inflation pressure is associated with greater soil compaction because it reduces the area contact between tire and soil [202,203,206]. Moreover, excessive numbers of machinery passes reinforce compaction and can spoil the positive effects of practices aiming at soil-structure enhancements, such as the use of organic amendments [203]. In intensive viticulture areas, where soil compaction due to tillage operations and machinery traffic is higher, there may be long-term effects on vineyard productivity [207]. Simply, soil compaction hinders root distribution both vertically and horizontally, thus interfering with water- and nutrient vine uptake, eventually hampering the achievement of yield and grape quality objectives, which may only be improved through regenerating soil management practices [208].
6.2. Cultivation Techniques to Avoid Soil Compaction
To avoid and mitigate soil compaction, the most suitable practices include minimal (or zero) tillage and machinery traffic. However, if machinery is used in vineyards, recognizing symptoms of soil compaction and developing methods to minimize its occurrence should be a top priority. The primary factors to bear in mind for preventing issues of soil compaction include kind, number, and places of soil tillage operations, as well as soil moisture and machinery weight and management in the field [199]. Therefore, in viticultural soils, compaction can be minimized through various strategies, including: (i) less passes over the vineyard; (ii) optimized weight equipment; (iii) adequate tire pressure; and (iv) use of cover crops and organic mulches.
To reduce soil compaction issues, simultaneous execution of multiple farm operations to minimize passes is recommended [203]. Additionally, controlled farming traffic, where machinery operations are confined to specific traffic lanes, using satellite guidance systems if needed, is an effective way to mitigate soil compaction [209]. Concerning tire inflation, avoiding excessively high pressures is important. The effective use of these techniques can stabilize soils against pore space collapse, and even be used alongside some reduced tillage operations to break up recalcitrant compacted soil layers with low probability of recompaction in the future, hence improving soil health and viticulture sustainability [210]. In this regard, the concomitant use of aggregating agents like organic amendments and gypsum may help to avoid recompaction [203].
The use of grassed vine inter-rows can be recommended as a corrective strategy to reduce soil compaction [207,211], as it lowers bulk density and root penetration resistance, increases soil porosity, and improves soil structure and infiltration rates [197]. However, it is important to note that the ameliorating effects of cover crops may take time to show up [212]. Therefore, cover crops with deep, strong roots capable of penetrating compacted soils should be preferred to combat soil compaction [203]. Additionally, the choice of the appropriate cover crops should be based on the specific characteristics of the vineyard, as well as others, as discussed in the next subsection.
6.3. Effects of Cover Crops and Mulching on Soil Health
Cover crops and living mulches, defined as any crop or crop mixture planted or allowed to spontaneously blossom into or after a main crop and then, respectively, killed or maintained alive, offer numerous advantages in grape production [213]. Despite their competition for water and nutrients, especially in the upper soil layers, where most of the annual vine roots are active [214], cover crops should be adopted, as soil and water conservation are becoming even more important in the face of climate change [215]. Regarding both synthetic and organic mulching, these have proven highly effective in conserving water around grapevine roots, a critical factor for viticulture in arid to dry sub-humid regions, thereby enhancing water use efficiency amid changing climate patterns [216,217].
The utilization of cover crops is closely linked to reduced tillage, demonstrating their joint effectiveness in increasing root biomass [218,219], and hence labile SOC, microbial biomass, and, eventually, SOC stocks [220]. Additionally, cover crops play an important role in improving soil N availability, thereby contributing to more efficient N uptake and utilization [221], reducing soil erodibility in terms of both lower surface-runoff rates and lower sediment load in runoff [176], controlling weed populations, maintaining grapevine yields, enhancing soil quality, and ultimately promoting the sustainability of grapevine growing [222].
The population and diversity of soil biota depends on the plant species that are chosen for the cover crop [20,217]. Indeed, increasing plant species richness may foster biodiversity across different trophic levels [223]. Remarkably, evidence from world-wide representative studies highlights that soil microbial diversity rises as a result of plant diversity in agroecosystems [224] with significant increasing effects on soil microbial biomass, bacterial biomass, fungal biomass, the fungi-to-bacteria ratio, and microbial respiration with eventual beneficial effects on soil nutrient cycling, particularly of C and N [225]. As for vineyards, Pingel et al. [40], in their research on cover crops in the German winegrowing region of Rhinehesse, observed a positive response in bacterial diversity to plant richness, with a positive but non-significant trend in fungal diversity. This increase in bacterial diversity may be attributed to the heterogeneous resources provided by cover crop plants, such as root exudates, leaf litter, and dead roots, as well as to the creation of diverse soil habitats by their root systems [224]. Importantly, these changes in the soil microbiome influenced by cover cropping practices may have implications for the grape microbiome and, consequently, wine quality [138].
Selecting the appropriate type of cover crops poses a challenge in vineyard soil management. Several characteristics should be considered when choosing cover crops [215]: (i) establishment capacity and resistance to repeated trampling; (ii) homogeneity and duration; (iii) ability for effective weed control; (iv) perennial habit to reduce the need for reseeding, that is to reduce costs; (v) reduced aerial development to minimize maintenance and vineyard interventions; (vi) seasonal growth pattern: featuring a lag in summer growth followed by autumn recovery; (vii) low evapotranspiration rates; (viii) deepening root growth patterns; and (ix) soil aggregate stability potential. Moreover, cover crop management, including the frequency and timing of maintenance operations and the choice of plant species, must be tailored to local climate and weather conditions, striking a balance between wine production, biodiversity, and ecosystem service provision [187]. Related to this, temporary plant cover, offering services like soil protection and water soil profile replenishment during winter, represents an attractive alternative to permanent ground cover, mitigating water and N competition with grapevines in dry years [220]. In this regard, the introduction of livestock in vineyards for grazing might help manage some cover crops, specifically considering that grazing animals can speed up the recycling of grass nutrients to benefit vines [226].
7. Impacts of Viticultural Phytosanitary Products on Soil Health
The profitable development of viticulture requires the use of crop protection techniques against several phytopathogens and parasites, which without control may severely harm grapevines and ultimately ruin the grape harvest [227]. Among these vine-damaging biotic agents, it is worth highlighting the fungi, which cause some well-known grapevine cryptogamic diseases such as downy mildew, powdery mildew, and grey mold. Additionally, there are many arthropods that particularly harm vines by herbivory or by being vectors of bacterial diseases, among which the leafhopper Scaphoideus titanus Ball, vector of the flavescence dorée disease, the grape berry moth Lobesia botrana, and the mealybug stand out [228,229,230]. In addition to these more vine-specific and widely spread fungi and arthropods, there are the weeds. Although weeds do not impair vines as much as the previous organisms, they are traditionally seen as competitors for water and nutrients, as hindering agricultural operations and, more grievously, as shelters for plagues of mollusks, bugs, viruses and other opportunists. Indeed, under uncontrolled circumstances, weeds can become highly detrimental for vine production, and thus viticulturists fight them with the aid of tillage operations and chemicals under conventional management conditions.
A ‘pesticide’ is something that prevents, destroys, or controls a harmful organism (‘pest’) or disease, or protects plants or plant products during production, storage, and transport [231]. However, often this something is of chemical nature and, specifically, of xenobiotic chemical nature. The use of chemicals for crop protection dates back to at least the third millennium B.C. [232]. However, since the onset of the Green Revolution, the use of agrochemicals for pest-fighting highly intensified and spread to all crops [233]. These kinds of agrochemicals are known as pesticides to be distinguished from fertilizers, although more recently, the nouns ‘phytosanitary’ and ‘plant-protection products’ have gained some ground. Whichever the name, the chemically based crop protection treatments showed some early failings in the form of the emergence of pest resistances, resurgence of primary pests, and upsurges of secondary pests [234,235]. Not least, the concerns about the effects of the xenobiotic chemicals that are used for crop protection on both the environment and human health have grown in recent decades [236,237]. Consequently, the techniques for crop protection have shifted from purely chemically based approaches to the combined use of these, along with biological and various cultural practices, under the umbrella of the integrated pest management concept [234]. However, in viticulture, despite the efforts to find alternatives to chemicals, the protection of vines still relies a lot on their use [238], even under organic production schemes [239,240].
Pesticides are classified into, mainly, fungicides, insecticides, bactericides, and herbicides, depending on their organism target [241]. The use of pesticides is particularly intensive in viticulture [242], with fungicides dominating the share of phytosanitary treatments [238]. Particularly in European agriculture, viticulture is by far the main user of fungicides with a market share of 86%, despite vines only spanning 3.3% of the cropland [243]. Among fungicides, of particular importance in viticulture are the Cu-based products. In addition to these, insecticides, mainly organo-phosphates, are also widely used in viticulture for the control of the leafhopper, the berry moth, and the mealybug.
7.1. Copper-Based Phytosanitary Products
Copper-based phytosanitary products in viticulture come in different compounds and formulations that range from the classical Bordeaux (CuSO4 and CaO in water) and Burgundy (CuSO4 and Na2CO3 in water) mixtures to the more modern Cu-oxychloride (3Cu(OH)2·CuCl2), CuSO4·3Cu(OH)2, Cu2O, Cu(OH)2, etc. The effectivity against the vine-typical fungal diseases, though particularly against downy mildew, of the Cu-based fungicides is difficult to match by whichever more environment-friendly strategy [239,240,244,245,246]. Therefore, Cu has been applied for vine protection since 1885 [247], and in rates that were as high as 50 kg Cu ha−1 yr−1 before legislations for their control were passed, e.g., the European Commission [231]. Because of the long-lasting and intensive Cu use in vineyards, its soil concentration has gradually increased to over 100 mg kg−1 [248]. Specifically in Europe, an estimated 15% of vine lands have exceeded the 100 mg kg−1 threshold [249] and, consequently, can be considered Cu-polluted according to some current standards [250,251].
Copper is an essential micronutrient for all organisms because its size and the reversibility and redox potential of the Cu2+/Cu+ pair make it irreplaceable as a cofactor of several oxidoreductases [252,253]. These Cu enzymes are mainly found in the electron transport chains of chloroplasts and mitochondria, in the detoxification of some reactive oxygen species (ROS), and, particularly in plants, in the lignification metabolism [254,255]. However, Cu in excess within cells freely catalyzes the generation of ROS [256,257] through the Fenton and Haber–Weiss reactions, which increase the cytoplasm concentration of the superoxide radical (O2−) and hydrogen peroxide (H2O2) species [258]. Then, the O2− and H2O2 generate other ROS, and all together oxidize proteins, nucleic acids, and lipids, thus destroying cell structures, particularly membrane systems [259].
In addition to the oxidative stress, Cu ions directly disrupt the structure of proteins in the cell membranes and in the cytosol, as well as the structure of nucleic acids, thus explaining how excessive Cu concentrations are biocidal [257]. Although plants can protect themselves better against excessive Cu than fungi or bacteria through, e.g., vacuole sequestration and restrained root-to-shoot transfer [258], the ROS generation boosted by excess Cu has also been described occurring in plant cells [260,261]. Copper-generated ROS damage in plants shows up as morphological and physiological impairments and as nutritional imbalances in most organs, particularly in roots [262,263].
Copper is applied to vines in formulations that prevent its absorption by the plant organs, hence trying to avoid the development of phytotoxicity and berry Cu build-up [247]. Whatever the case, vine Cu-treated leaves eventually fall and decompose and/or rainfall washes Cu down to the soil where it may accumulate [264], and hence be absorbed and excessively built up in vine organs including grapes [265]. The plant-availability of Cu depends on the free Cu2+ activity in the soil pore water, which depends, in turn, on soil pH and Cu’s resupplying ability from the soil solids [266]. In fact, since the solubility of Cu2+ minerals increases as pH decreases [267], the detrimental effects of excessive soil Cu concentrations upon plants develop, overall, in acidic soils [268,269,270].
In grapevines, toxic effects of Cu include oxidative stress in leaves [271], which may cause decreased photosynthetic and transpiration rates [272]. Moreover, excess Cu shows up as morphological changes in the roots: increased diameter, higher number of layers in the cortex, and rising vascular cylinder and total root areas [272], and less nitrate uptake [273]. Because of these metabolic disorders, impaired nutrient status, and root deformations, grapevines produce less dry matter [272,274]. In soils featuring pH values over 7, Cu2+ readily precipitates as solids like Cu2(OH)2CO3 (malachite) and Cu(OH)2 [275]. Additionally, Cu2+ activity in the soil solution steeply decreases from pH values of 6.5 upwards because of the formation of the CuHCO3+, CuCO3, and CuOH+ ion pairs and, accordingly, Cu toxicity severely decreases as soil pH raises [276]. Finally, soil solution Cu2+ concentration is remarkably decreased by rising levels of SOM since Cu2+ is tightly adsorbed by SOM as surface complexes [277,278,279].
In addition to vines, toxic effects of Cu have also been described on soil organisms, and for these, avoiding high soil Cu intake in soils with elevated Cu concentrations may be more difficult than for plants. Particularly, earthworms feed in the soil, and upon excessive Cu they decrease growth and survival rates [280]. Specifically, Eisenia fetida is affected by soil Cu concentrations from 28 mg kg−1 up [281], and it is, notwithstanding, more tolerant to Cu than other earthworms [282]. This 28 mg kg−1 Cu concentration could be overcome in the arable layer of vine soils (0–30 cm) in only 16 years, despite following the restrictive EC regulation 473/2002 [231], which establishes maximum rates of 8 kg Cu ha−1 yr−1 during the first 4 years of vineyard establishment and then 6 kg Cu ha−1 yr−1 onwards.
Zinc compounds are used as alternative to copper for fungal control in vineyards [283]. However, in general, viticulture cannot get by without Cu, and thus ways to adapt must be found, by means of which the Cu issue is minimized. Therefore, if soil Cu is rated to vines below 10 kg ha−1 yr−1, then concentrations over 200 mg kg−1, which severely impair the soil’s biological community functionality [284], should not be overcome. In acidic soils, the observation of these constraints is more important, even though under such circumstances the harmful effects of Cu on vines and soil organisms can be counteracted by liming to raise the soil pH [274]. In this regard, higher soil pH values not only decrease the solubility of Cu2+ minerals and the activity of Cu2+ in the soil solution, and therefore decrease the plant Cu availability and phytotoxicity, but also the Ca2+ ions play a beneficial role through competition with Cu2+ ions for plant uptake [276,285]. Additionally, the contribution of organic matters, such as farmyard manure, crop residues, and rhizodepositions from cover crops, may increase SOM contents and thus reinforce the protective effect of higher soil pH values. Moreover, aging decreases Cu availability [275], and to maximize the benefits from the aging factor, minimum soil disturbance, i.e., minimum tillage, may help.
Additionally, for mitigation of the Cu issue, cover crops have some potential in the remediation of vineyard soils [153]. Particularly, in non-calcareous soils, which are the most Cu-troublesome, cover crops featuring crimson clover seem the most promising since they have been found able to extract more than 0.1 kg Cu ha−1 for one year from soils containing between 80 and 160 mg Cu kg−1 [154]. Note that this range corresponds to between 20 and 70 kg Cu ha−1 if one takes the layer from 0 to 20 cm and a bulk density of 1.5 g cm−3. Other Cu-hyperaccumulator plants might be used as cover crops for Cu phytoremediation in vineyard soils, but more research is needed to provide practitioners with validated knowledge, and hence useful recommendations.
7.2. Organically Based Phytosanitary Products
In viticulture, herbicides and insecticides follow fungicides in this order as the most-used phytosanitary products by market share [15]. Being for the most part organically based, herbicides can be classified into phenoxy hormone compounds, triazines, amides, carbamates, dinitroanilines, urea derivates, sulfonyl ureas, bipiridils, and uracils [15], whereas insecticides can be classified into organochlorine, organophosphate, carbamate, pyrethroid, and neonicotinoid [286].
The herbicides are increasingly applied in vineyards worldwide with glyphosate, glufosinate, and flazasulfuron leading as the three most-used [287]. These three herbicides have been shown to decrease grapevine root mycorrhization and alter soil microorganisms, with consequences on the nutrient composition of grapevine roots, leaves, xylem sap, and grape juice [287]. Specifically regarding soil organisms, glyphosate has been shown to decrease the survival and reproduction rates of earthworms in vineyards [288], thus raising concerns about how it affects soil health. Even though its effects on weeds may vanish, glyphosate presence in the soil may last a long time as it is strongly adsorbed by SOM [289]. Moreover, from SOM, glyphosate may be gradually released to the underlying waters [290], thus polluting them.
A sustainable alternative for controlling weeds in vineyards is the integration of livestock such as sheep [226] and geese [291]. For obvious reasons, livestock introduction in vineyards may be particularly adequate during the vine-dormant stage, although new developments might extend it to the active stage [292] when, additionally, the shepherding activity might help prevent wild animals from feeding on the vines. A drawback is the likely Cu chronic poisoning of the livestock [291,293] because of the thorough application of Cu fungicides, as previously addressed. Due to all this, methods for the beneficial integration of grazing animals in vineyards should be widely investigated.
The organophosphate pesticides were developed as an alternative to organochlorine ones, with the advantage of being less stable, less lipophilic, and less volatile. However, they are more hydrosoluble and are, therefore, more readily dispersed in water until they are degraded, which spreads other, more chemically simple, but still xenobiotic compounds throughout the environment.
The organophosphates comprise many agrochemicals like malathion, parathion, chlorpyrifos, diazinon, fonofos, and disulfoton, which are formed by the esterification of phosphoric, phosphonic, phosphinic, or thiophosphoric acids with a variety of organic compounds such as those of the phenoxy, cyanide, and thiocyanate kinds [294]. The organophosphates act mainly by inhibiting esterases, among which the acetylcholinesterase stands out. The inhibition of acetylcholinesterase activity increases the levels of the neurotransmitter acetylcholine up to toxic levels, thus harming nerve communication in insects, but also in other organisms [295] including those in soil [296], with harmful effects [297] that negatively impact the survival and reproduction rates of earthworms [298].
In viticulture, organophosphates have been found to be effective enough against the mealybug Planococcus ficus, which is the most important insect pest affecting vines worldwide, including the Mediterranean basin [299]. Specifically, applications of 0.96 kg ha−1 of chlorpyrifos and of 0.48 kg ha−1 of methidathion have shown a mean efficacy of 70% against the sap-sucking mealybug in vineyards in Spain, Portugal, and Greece [300].
Although the organophosphates present advantageous characteristics compared with other pesticides, they still may accumulate in soils where they damage the soil’s biological community [298]. Particularly, chlorpyrifos has been found to be harmful to not only soil earthworms [297] but also bacteria [301,302] thus impairing soil health. Therefore, for the control of insects in grapevine growing, notably the mealybug, the combination of other more effective but at the same time more eco-friendly chemical insecticides such as the spirotetramat with other non-chemical strategies within an integrated pest management approach offers a promising way to enhance the soil health of vineyards [299].
7.3. Interrelation of Soil Health and Vine Health: Pest and Disease Pressure
Several pests and diseases pose a threat to grapevines, and boost the need for control over them, which is usually carried out with xenobiotic chemicals, as has been shown. The impacts of climate change are likely to alter the ecology of many harmful biological agents with, e.g., insects hitting crops harder in the temperate regions where most of grapevines are grown [303]. Therefore, adaptations in plant protection measures should unfold as we witness an increase in pest and disease pressure in vineyards [304]. Globally, the sustainable management of vineyard pests and diseases is considered a top research priority [238,305]. Soil-borne pests, in particular, can inflict significant losses on all crops, including vineyards, affecting both grape yield and quality [33]. These losses entail severe economic, environmental, and social implications and, therefore, to achieve soil pathogen suppressiveness is a desirable goal. However, the current focus also lies in preserving soil biological diversity, enhancing soil and plant health [306].
Therefore, a fundamental question is how to enhance the resilience of vineyards to pests and diseases while promoting sustainable productivity. A key solution lies in developing practices that promote general soil health and, consequently, enhancement of beneficial organisms and suppression of pests and diseases, with the eventual achievement of healthy vine growth and improvement of the environment. Specifically, the soil structure should be enhanced since it exerts a remarkable control on the soil Eh and pH, and the soil Eh-pH conditions are related to the susceptibility, tolerance, and resistance of plants to pests and pathogens across various stress conditions [307]. Indeed, it has been hypothesized that soil pathogen suppressiveness results from soil structure, leading to diverse Eh-pH niches that increase the variety of soil microorganisms. Soil structure can be enhanced through, among other measures, an adequate management of organic amendments. These amendments can be, additionally, customized to induce significant shifts in the soil microbiome, promoting favorable consortia that sustain a disease-suppressive viticulture system without the need for additional agrochemical inputs. However, it is worth noting that some organic amendments may, in certain cases, trigger other neutral or, even negative, effects [308], as will be in part addressed in the next section and, therefore, more research and developments are needed in this regard.
Soil-inhabiting microbes play pivotal roles in shaping grapevine-associated microbiota and are key factors in wine quality by means of their significant impact on vine health and productivity through various direct and indirect processes, including the triggering of vine immune responses [34]. Furthermore, the soil microbial community in the rhizosphere, which could serve as a new biological indicator of vineyard soil quality and health, is a part of alternative methods that show promise in protecting grapevines. These methods include biocontrol and microbe-assisted crop production to manage grapevine diseases and promote sustainability in viticulture [57].
According to the aforementioned facts, general practices to increase soil health for pest and disease suppressiveness should include the following [309]: (i) minimizing soil disturbance and reducing compaction, (ii) maximizing biodiversity in the soil and aboveground through mulching and cover crops or by applying various types of sanitized and unpolluted organic matter amendments, (iii) managing water to meet crop needs but also to facilitate timely fieldwork, (iv) maintaining soil Eh-pH values within adequate ranges, and (v) ensuring nutrient levels are sufficient for vines but never excessive.
8. Fertilizers and Soil Amendments for Soil Health in Viticulture
One of the primary challenges in the pursuit of agricultural sustainability and soil health lies in the effective management of fertilizers and soil amendments, both mineral and organic. The use of one and the other should be integrated, and vineyard soil fertilization strategies should be tailored to the diverse soil and climate situations encountered in actual practice. Particularly, the likely impact of vine nutrition choices on soil processes should be analyzed prior to their application to optimize soil health in vineyards [220]. In this regard, the use of models to simulate the nutrients’ transformations and fate in the soil–plant–water–atmosphere system is recommended.
In the quest for soil health, soil management strategies in viticulture should extend beyond merely optimizing grapevine yield and quality: they should also aim at sustaining soil biodiversity and overall soil functioning [40]. The application of inorganic fertilizers in excess of vine requirements is known to alter plant vigor, with unwanted effects on grape production and quality. Furthermore, the soil’s biological functionality is severely affected following excessive mineral fertilization, which shows up in lower essential soil enzymatic activities such as dehydrogenase, catalase, invertase, urease, casein protease, and arylsulphatase [58]. Long-term N fertilization can influence soil microbiome populations, thus reshaping vine and soil microbial communities [40]. Therefore, an excess of mineral fertilization, beyond crop requirements, harm plant and soil health. In addition, such mineral fertilizer overuse can result in secondary salinization issues and nutrient leakage from the soil–plant system into other environmental compartments through leaching and gaseous emissions, thereby polluting the environment and further compromising environmental health [129].
8.1. Mineral Nutrients Effects on Vines and Soil Health
Mineral nutrition plays a pivotal role in grape production, exerting influence over all aspects of vine growth, berry development and maturation, and ultimately, wine quality [310,311]. The primary macronutrients, i.e., N, P, and K, control critical grapevine metabolic processes, which influence grape quality and thus the eventual alcoholic fermentation process [46]. Therefore, soils deprived of essential nutrients lead to nutrient-deficient vines, resulting in low-quality grapes and imperfect wines [312,313]. To meet vine nutrition demands is imperative for achieving production and oenological objectives. However, while it is well-established that nutrients like N and K affect berry and wine quality, and despite recent advances on establishing N levels for maximizing berry quality [136], the precise levels of the remaining nutrients for attaining optimal grape quality must still be refined [59]. Therefore, to ensure a sufficient supply of nutrients, viticulturists often rate fertilizers in excess, thus eventually spoiling grape quality, as is known to occur with excessive N [136,314] and K [310].
Additionally, there are the environmental harms associated with over-fertilization, which show up as nutrient leakages to the atmosphere in the form of NH3 and N2O gas emissions, and to waters in the form of dissolved NO3− and colloidal organic P. Particularly, intensive viticulture can result in environmental pollution problems due to excessive nutrient inputs that surpass the soil–plant system’s capacity to absorb [161]. Complicating matters further is the impact of climate change on grapevine mineral nutrition. For instance, under the increasing warming conditions caused by climate change, K increasingly builds up in berries, thereby decreasing must acidity, and eventually harming overall wine quality [315]. Moreover, the improper use of mineral fertilizers, particularly nitrates and phosphates, not only jeopardize atmosphere and water health, but also the vineyard soil health in terms of its ecosystem services as shown by chemical indicators, and moreover by physical and biological ones [316]. Furthermore, the use of mineral fertilizers, which are directly salts featuring K+, NH4+, NO3−, and phosphate ions, in addition to, often, Na+, Ca2+, Mg2+, Cl−, and SO42−, or either indirectly have the ability to generate salts by reaction in the soil, can induce secondary salinization [317] resulting in a deterioration or loss of soil functions.
To mitigate the negative impact of fertilizers on vineyard soils, particularly the secondary salinization issue, excessive nutrient applications must be avoided, and high-purity, chloride-free, low-saline fertilizers should be selected [318]. Additionally, soil management practices to reduce the secondary salinization hazards due to fertilizer overuse include: (i) improving the efficiency of fertilizer applications through irrigation water (fertigation), (ii) improving the salt tolerance of the grapevine by the addition of different nutrients based on competition between ions, and (iii) ameliorating the deleterious effects of salt stress by the addition of humic substances or by the use of biofertilizers, which reduce the application of chemical fertilizers [318].
To mitigate over-fertilization issues in general, viticulturists will have to use several tools, such as the ones based on the current information and communications technologies. In this context, soil fertilization should be based on the spatial variability of the soil characteristics as well as on the vine nutritional status and requirements, as these change throughout the growing season [319]. Additionally, devising effective fertilization schedules for vineyards will benefit a lot from the use of models. Bespoke fertilization practices to enhance both macro- and micro-nutrient management, thereby supporting soil health, hinges on the integration of the processes governing nutrient availability, soil properties, and environmental conditions, along with vine nutrient requirements [320], and models offer an optimum way to perform such integration. Therefore, fertilizer recommendations in vineyards could be adapted to the varying nutrient uptake ability of the different Vitis genotypes and the influence of rootstocks on nutrient concentrations in the grafted scion [167]. As our understanding of the nutrient requirements of specific cultivar–rootstock combinations advances, site-specific fertilization management programs will be tailored to individual vineyards through the use of models. This will help reduce fertilization costs and environmental pollution, promoting vineyard sustainability [321]. For now, adequately validated models for the simulation of the nutrient dynamics in viticultural soil–plant systems are lacking and more investigation and innovation is needed in this regard.
8.2. Organic Approaches for Soil Health in Viticulture
Organic farming represents an alternative to conventional agriculture based on chemical supplies. Organic farming is based on holism principles and ecological interactions instead of reductionism postulates and artificial forcing. Therefore, organic farming does not fight nature; on the contrary, it takes advantage of the natural processes that already unfold in the agro-ecosystems so as to fulfil the objective of food- and raw materials production for human welfare. Such collaboration with nature fostered by organic farming logically implies that some agricultural produce is lost to it; specifically organic farming systems exhibit 20% lower yields than their conventional counterparts [322]. However, such loss is traded off by 34 to 53% less energy use, as well as 97% less pesticide use [322], hence enhancing environmental health.
Organic farming pays particular attention to soil health, and hence postulates the joint and integrated use of various nature-based strategies to enhance soil functions, among which the following may be cited, especially for vineyards: (i) the application of microbial and non-microbial biostimulants, (ii) fertilization with organic amendments, (iii) foliar fertilization with nature-based products, (iv) the use of cover crops, and (v) the integration of livestock [58,323]. Therefore, for organic grape production, success requires treating the vineyard as a balanced agro-ecosystem to maintain the trade-off between, on the one hand, grapevine productivity, provided vine health is enhanced and weed issues minimized, and on the other hand, soil ecosystem services [324]. Consequently, although there are variations in cultural practices in organic farming, they usually increase sustainability metrics such as soil species richness and abundance, overall soil fertility, and water infiltration and holding capacity [323].
Organic viticulture, by avoiding synthetic fertilizers, soil conditioners and pesticides as much as possible, can enhance the activity of beneficial soil organisms such as entomopathogenic nematodes and increase the presence of free-living nematodes [23]. Furthermore, considering that economic sustainability is also a concern for viticulturists, organic vineyard management offers a viable model to achieve economic goals while safeguarding the environment from issues like water pollution and depletion, greenhouse gas emissions, and soil degradation, ultimately promoting biodiversity [325]. Specifically, Tangolar et al. [326] observed in their study on cv. Black Magic grafted onto the Paulsen 1103 rootstock that microbial biomass C and enzyme activity, including dehydrogenase, urease, and phosphatase, were higher when compost was applied to a calcareous vineyard soil. Similarly, Di Giacinto et al. [327] found that organic viticultural soil management resulted in higher soil enzymatic activities, as well as a richer bacterial and fungal community, because of the use of cover crops and organic compost.
As well as composts, biochar, a stable C-rich material produced through biomass pyrolysis, i.e., burning under restricted oxygen access, represents another potential sustainable soil conditioner [328]. Biochar has been shown to continuously increase the saturated hydraulic conductivity and plant-available soil water content when applied up to rates of at least 80 Mg ha−1 [329,330], and to serve as an effective material for sequestration of potentially toxic elements [331]. In vineyards, the same effects on soil hydrological properties have been reported, entailing better water status and enhanced long-term photosynthetic activity of vines [332], which may ultimately increase vineyard production. Specifically, in their four-year study involving cv. Merlot grafted onto the Couderc 3309 rootstock, Genesio et al. [333] found that biochar augmented vine-water availability, thus increasing yield by 66% on average, with particular harvest-increasing effects during dry years, and no effects at all on berry quality. Interestingly, there are important variations in physicochemical characteristics resulting from differences in feedstock for biochar production and, in this regard, crop residues, as biochar raw materials, score higher in their soil-beneficial effects over manures, wood, and sludges [329].
To address soil nutrient limitations for vines and other physical and biological soil fertility constraints, the use of organic fertilizers is vital due to their potential for sustaining and restoring soil health. However, it is worth noting that organic fertilizers also entail hazards, heavy-metals concentrations being one of the most common [15]. The presence of heavy metals in significant concentrations in organic fertilizers, particularly sewage sludges, has been well-documented and is very variable [334]. Therefore, to avoid any soil pollution threat, the concentrations of heavy metals in organic materials should be known before use. Then, their rates can be conformed to safe standards, which usually also take account of key soil chemical properties, mainly pH, because in acid soils the mobility of many heavy metals increases [335].
Another point of caution in the use of organic fertilizers is the utilization of animal manures and the secondary soil salinization they may induce, which has been given much less attention in comparison with the heavy metal issue. Thus, successive applications of animal manures with high Na+, K+, Cl−, and SO42− concentrations, since these ions are commonly added into the animal diet, can also promote secondary salinization in vineyard soils [336], chiefly in arid and semi-arid regions.
To adequately rate organic materials in combination with mineral fertilizers, or alone, e.g., under an organic winegrowing scheme, the use of validated models of the soil–vine–water–atmosphere system that take into account the composition or the organic materials, including the different nutrients, heavy metals, and accompanying salts would be highly recommended.
8.3. Soil Acidity and Liming Effects on Soil Health in Vineyards
Soil acidity naturally develops due to various factors of soil formation, which include parent materials that are low in bases and humid climates that promote the dissolution of the alkali and alkali-earth minerals, including the aluminosilicates, followed by the leaching of their dissolution products [337]. However, productive agriculture also contributes to soil acidification, mainly through the use of acid fertilizers such as ammonium salts and hydrogen phosphates, resulting in adverse effects on soil health, which include the development of phytotoxicities due to hydrogen ions (H+), aluminum (Al), and Mn, and the limited plant availability of essential nutrients [338]. The phytotoxic effects of H+ and Al typically show up as damaged root systems in the first place, whereas those of Mn directly affect above-ground plant parts [338]. Moreover, in general, the mobility of heavy metals in the soil environment increases as pH drops [335], with likely consequences on vine health. However, vine damages by heavy metals, specifically decreases in plant growth and photochemical efficiencies, have only been studied for Cu and Zn [283], because of the particular importance of both in phytosanitary products for fungal control in viticulture, as previously addressed (Section 7.1). Therefore, more research in this regard is needed.
In addition to phytotoxic effects, soil acidity issues are characterized by the appearance of Ca deficiencies, and to a lesser extent, also of P, Mg, and Mo deficiencies [339]. Particularly, P deficiency in acid soils affects the vine reproduction more than the vegetative development [340], and is the result of high phosphate fixation by clay-sized Al and iron (Fe) oxides and hydroxides, as well as the formation of phosphate precipitates of Al, Fe, and Mn [341].
In addition to the effects of acidity on chemical fertility and plant nutrition, low soil pH values significantly influence the soil microbial community. Indeed, soil pH is currently considered the best predictor of microbiome diversity at the phylum level [162] and the primary soil driver for bacterial diversity [40]. The growth and activity of soil microorganisms in acidic soils are constrained by high H+ concentrations, which negatively impact most soil microbial processes, which show up in low rates of SOM turnover and nutrient cycling [338]. Furthermore, since soil fungi thrive in low pH environments, acid soils experience an imbalance between fungal and bacterial populations, with fungi often dominating. This can create a favorable environment for fungal pathogens, increasing the risk of infections, and reducing nutrient mineralization [162], and hence harming soil health.
For the long-term sustainability of viticulture agro-ecosystems and soil health, managing soil acidity is crucial. Traditional approaches to address soil acidity issues involve increasing soil pH by liming with materials rich in CaCO3, CaMg(CO3)2, and Ca(OH)2, which neutralize acidity and increase soil Ca. However, these strategies usually affect the topsoil only [337,342], whereas vine roots can pierce deeper. Therefore, preventing or minimizing subsoil acidification is as important as mitigating the topsoil acidification issue, being essential to understand the processes of subsoil acidification such as downward movement of soluble Al [343]. In this regard, a viable strategy to address soil acidity issues, which may be developed alongside the previous liming efforts, is grafting crop plants onto acidity-tolerant rootstocks. In viticulture, practitioners have for strongly acid soils (pH < 5.5) several available soil-acidity-tolerant rootstocks, such as Couderc 3306, Fercal, Gravesac, Paulsen 1103, Richter 99, Richter 110, and Ruggeri 140 [344].
9. The Expression of Terroir and Soil Health
Beyond grapevine cultivars, it is well known that the sensory attributes of wine depend on the soil and climate conditions in which the grapevines are cultivated, and in its simplest form, this constitutes the terroir concept [46,345]. This way, the ‘terroir’ idea has been anchored to phrases such as “the art of expressing the soil”. However, the wine industry, including researchers, professionals, and consumers, has collectively acknowledged that a specific terroir should not be an abstract notion like that: both wine producers and consumers should have clearer, more specific concepts [346]. Additionally, it is also recognized that cultural practices play an important role in contributing to terroir and, furthermore, the interactions among grapevine management, soil, climate, and grapevine cultivars and rootstocks. Consequently, terroir encapsulates all the biological, environmental, and cultural influences on grapevine cultivation and winemaking as well as their interactions, recognizing the complex multifaceted nature of wine composition, and hence its sensory attributes [347].
Therefore, the specific characteristics of one vineyard constitute its terroir, and consequently terroirs change as such characteristics vary. Then, the wine composition, and thus its organoleptic properties, expresses the terroir and consequently, the terroir expression changes following the vineyard characteristics throughout space and time. In the context of the soil-health loss unfolding in conventionally managed vineyards, as well as its regaining by means of improved management practices, there is a necessity to gain insight into how the spatial and temporal variations in grape and wine composition relate to the variability in soil and climate characteristics as well as management practices. This understanding will lead to the establishment of which spatial scales and resolutions are the most appropriate for managing the production of terroir wines within a specific region while preserving the environmental health [46]. In this context, the scientific community has been called upon to assist in identifying and defining the numerous interconnected aspects of terroir that collectively [347] contribute to shape the quality and characteristics of the wines resulting from one area [348].
Achieving a more profound comprehension of the connections between the vine and the environment where it thrives requires not only scientific expertise on winegrowing but also interdisciplinary approaches. Current technological capabilities in land zoning are instrumental in exploring the intricacies of terroir expression [349]. Therefore, terroir research has evolved from being predominantly descriptive and region-based, i.e., primarily focused on traditional soil and climate zoning, to becoming a more applied, technical field with two main development lines. These are based (i) on the use of metabolomics and metagenomics for assessing terroir characteristics, and (ii) on employing remote and proximal sensing to monitor soil and vine characteristics so as to give practitioners useful information to precisely manage the soil–vine system and attain predetermined oenological objectives [46].
9.1. The Factors That Determine the Terroir Expression
If focusing solely on one factor at a time, a multitude of physical and chemical soil and climate features can influence the terroir expression. For example, in rain-fed vineyards, the soil’s water-holding capacity largely determines the water supply to the grapevines during the ripening process, and thus it controls grape and wine quality remarkably [350]. However, in terms of the balance between vegetative and reproductive grapevine growth, which influences grape quality attributes, distinguishing the effects of increased water supply from increased nutrient supply is often challenging [59]. In many regions, it has been shown that climate change makes vine growth start earlier and progress faster with consequences on ripening profiles and wine styles [351].
To comprehend how soil and climate features operate on terroir expression, it is crucial not only to consider the factors one by one, but also the interactions among them [352]. Then, gaining a comprehensive understanding of the mechanistic connections between soil and climate and grapevines, mediated through the human interventions, which modulate the effects of soil and climate on the vines, can facilitate the analysis of the effects the soil health status have on wine terroir expression, and how to counteract them if needed. In this regard, since climate change affects grape and wine quality, and hence terroir, the identification of new suitable vineyard locations, and the precise vine management to achieve specific oenological objectives can be eased [349]. Specifically, our insight on the terroir expression significantly increases when considering the interactions among soil, climate, and human factors as revealed by, e.g., topography and fruit zone microclimate [353], and ground microbiome.
Particularly relevant is topography, which is shaped by the forces of tectonics, erosion, and land mass movements [354], often intermediated by human actions. Topography serves as an important nexus between soil and climate in the terroir expression, affecting not only the wine quality but also the feasibility of cultivating vines in climates that overall fail to meet grapevine requirements [353]. Another connection of soil and climate with terroir occurs at the level of the microclimatic variations in the bunch zone, which depend on the soil characteristics, singularly the albedo, and are also regulated by the canopy management [352].
Therefore, the human factor interacts with soil and climate and thus modulates the terroir expression through winegrowing practices that can be based on inherited culture and novel scientific knowledge. Even more, the decisions made by viticulturists regarding soil and overall vineyard management, directly or indirectly influence soil health and wine terroir. In this regard, examples exist of how wine growers’ practices that promote secondary soil salinization lead to outcomes in terroir expression [355], featuring soapy and salty tastes, as well as loss of fruitiness, and increases in sour, bitter, and oxidative perceptions in wine [356]. Therefore, through grape build-up of Na+ and Cl− ions, soil salinization becomes one of the clearest drivers of changes on terroir expression because of soil-health loss.
9.2. The Soil Environment and the Terroir Expression
When it comes to grape quality attributes, the significance of the soil’s physical and chemical properties cannot be overstated [319]. However, despite some exceptions, e.g., salts, the soil impact on wine terroir is often notably intricate because soil influences so many aspects of vine development, from mineral nutrition to water uptake, but also rooting depth and temperature within the root zone [349]. Therefore, despite years of research on how soil factors contribute to the terroir expression in wine, there are still fundamental questions that require more investigation. Specifically, two questions linger: (i) which are the primary soil factors that influence terroir expression, and (ii) how do these factors interact among each other and with the vine to influence grape and, ultimately, wine quality and terroir expression.
Soils differ in their capacity to retain water, and also in the quantity and composition of the mineral nutrients they contain, and the extent to which these nutrients are available for uptake by vine roots [167]. Thus, an understanding of the spatial variability of soil fertility and plant nutrition is crucial for planning and implementing site-specific vineyard management, making viable the adoption of differential management strategies [319]. The information regarding soil spatial variability is of paramount importance in the identification and classification of terroir [349].
Although the precise influences of specific soil characteristics on terroir are not yet fully understood [46], it seems that thermal properties, pH, salinity, macro- and micronutrients, and the biological activity of soil organisms all contribute to the terroir expression. Beyond the influence of specific soil components on the attributes of wine quality, which is still a topic of research, some ecophysiological mechanisms have been shown to be highly influential, particularly in relation to water supply and N nutrition during critical stages of the vegetative cycle [46].
It is widely accepted that the soil’s physical characteristics, particularly structure, exert significant control over the vine water supply [353], which strongly impacts N dynamics [46], as well as the qualitative response of the berries [349]. Specifically, the soil water availability has been found to influence the hormonal balance of each grapevine variety, subsequently regulating genotype expression [46]. Additionally, as with vine water status, soil N availability critically affects grape quality [136]. For example, in vineyard soils high in N, vines present excessive vigor, resulting in shaded canopies and subsequent poor grape quality [357]. Regarding other soil chemical properties, significant associations between them and wine’s organoleptic characteristics have been found [353]. For example, Tomasi et al. [358] reported the effect of high limestone content on obtaining higher aromatic wines, and Arnó et al. [319] found that high calcium carbonate and pH lead to Mn deficit and, consequently, grapes low in phenolic contents.
In addition to the physical and chemical properties, the soil microbial diversity associated with the vineyard location is a key aspect directly involved in vine cultivation and growth, as well as grape quality and winemaking [348]. However, the global and local variability of soil microbial communities and the biotic and abiotic factors that drive their differentiation are still poorly understood [359]. The richer the soil in life forms, the more capable it is of not only providing essential ecosystem services [354], but of influencing the terroir expression [360]. Emerging technologies such as metabolomics, with the ability to identify the chemical fingerprint of a particular cell, organ, organism, or tissue type, and metagenomics, with the ability to sequence the entire microbial genomes present in vineyard samples, hold promise for understanding the terroir formation with regards to this [46]. Additionally, although the debate on whether there is a microbiological component in the terroir expression is currently not very influential, researchers like Zarraonaindia and Gilbert [38] have highlighted the importance of the soil microbiome in shaping the microbiology of the vine, and possibly directly shaping the characteristics of wine.
Indeed, the recent advancements in omics techniques have already allowed for a more detailed exploration of soil functionality, microbial diversity, and vine-associated microorganisms [46]. For instance, Lazcano et al. [138] found that soil may serve as a reservoir for grape microorganisms that are carried over into fermentation, greatly influencing the organoleptic properties of wine. This microbial presence exhibits a strong biogeographical pattern that enables the differentiation of terroirs, leading several scientists to propose the concept of the “microbial terroir” [361,362]. In fact, the soil microbiome should be considered as an important variable in identifying viticultural sites for the definition of homogeneous functional zones such as basic terroir units [348]. Additionally, Vink et al. [363] demonstrated how specific rootstock–cultivar combinations influence bacterial diversity in grafted grapevines. This could affect the capacity of specific combinations to chemically attract specific bacteria associated with plant protection, performance, and productivity, through root exudates, as well as colonization in grapes, potentially affecting wine quality.
9.3. Terroir Expression and Soil Health in Zoning Procedures
Terroir forms the primary basis for representing regional wine quality. Even on a small vineyard, certain factors can introduce significant variations in local soil, resulting in remarkable variability in terroir characteristics [364]. Therefore, it is evident that a complex issue arises when maintaining terroirs unchanged is an objective in a scenario where soil health is changing alongside time and space, either worsening or improving [138]. Particularly, soil management, which significantly regulates overall soil-health status, is anticipated to affect terroir and therefore, wine quality attributes. This may be a drawback, but also an advantage. For example, because agrochemicals substantially influence the soil environment, alternatives like organic grape production represent an opportunity for grape growers and winemakers to develop wines with unique terroir, distinct from conventional wines [324]. However, several questions still remain: (i) how much each soil property contributes to terroir expression, (ii) how terroir expression changes as soil degrades, and (iii) how terroir expression changes as CAPs are introduced in vineyards and soil health is enhanced.
In this paper, we aim to establish the primary parameters to consider when the concept of soil health relates to vineyard soils. Given the evolving nature and scope of the terroir concept, identifying highly healthy soils may be one of the most crucial steps in vineyard zoning. However, considering that every winegrowing terroir can be defined as a spatial and temporal entity characterized by homogeneous or dominant attributes [365], and considering the legal-social framework associated with designations of origin and geographical indications aimed at ensuring wine origin for consumers, conflicts of interest and disputes seem likely. Perhaps the creation of new hierarchical scales of vineyard entities, incorporating soil-health parameters into quantitative and environmental/ecophysiological zoning criteria, could not only ensure a more comprehensive conflict resolution system between stakeholders but also promote the adoption of sustainability practices in grape production.
The wine industry has a fresh opportunity to play a vital role in promoting soil-health restoration and mitigating the impact of climate change through regenerative soil management practices. By minimizing soil disturbance and promoting biodiversity, it can create healthier and more resilient vineyards, potentially enhancing grape quality and the flavor of wine [366]. Consequently, managing for soil health becomes a strategy to enable the full expression of terroir [138].
10. Chemical, Physical, and Biological Indicators of Soil Health
As reviewed, to be considered as healthy, a soil should present the levels of its physical, chemical, and biological properties within the ranges that enable it to function within its ecological boundaries to sustain productivity, maintain environmental quality, and promote plant and animal health. However, the quantification of these ranges is not usually straightforward because soils are complex systems and hence all their properties are interrelated among each other, with the result that the evaluation of soil health depends on that interrelationship rather than on the particular values the attributes present [129,367]. Consequently, soil health and soil quality may be better assessed on the basis of indexes that make use of several selected soil properties [368,369,370,371]. However, alternatively, soil health may be assessed on the basis of indicators that make use of an integrative soil property, which is ideally a biological one [372,373,374], for which SOM or SOC are often advantageously used as proxies.
10.1. Soil Organic Carbon
Soil organic carbon (SOC) is recognized worldwide as having great potential to be used as an indicator of soil health because it correlates well with many attributes of soil’s biological functionality [375,376,377,378,379], and hence with soil-health indexes [380], and because it is easily measured through, e.g., soil sampling and ensuing classical titration or gravimetric methods in the laboratory [212]. However, the adoption of SOC as a soil-health indicator is not without drawbacks. First, there is no universal consensus on which levels of SOC have to be considered as healthy [68], because SOC levels depend on climate and soil conditions. Consequently, when soil health is evaluated on the SOC basis, soil health seems to increase as temperature drops, and rainfall rates and soil clay contents increase [381,382]. One option to sort out this issue would be the calculation of normalized SOC levels (NSOC). These NSOC levels would be calculated with regards to the potential SOC (PSOC) that represents the soil class and climate type to which one soil belongs by using equations such as the Equation (1):
where PSOC levels could be established by survey or by modelling or both.
NSOC = SOC/PSOC
In addition to normalization, some kind of concave score function that takes into account both the positive and the negative impacts of SOC on soil health should be adopted for use of SOC as soil-health indicator. These negative impacts mainly arise due to the effects high SOC levels have on the fate of the phytosanitary products in the soil environment. Indeed, the adsorption of many pesticides by SOM increases as the SOC level rises [383]. Consequently, as SOM increases the activity of pesticides against their target organisms decreases, and their loss to groundwater in association with the dissolved SOC fraction may increase, thus boosting the need for pesticide reapplication with on-farm as well as off-farm harmful effects on animal and human health [384].
10.2. Soil Aggregate Stability
Soil aggregate stability also has the potential to be used as indicator of soil health because it correlates well enough with many attributes of soil quality [381,385], and of susceptibility to water erosion [120,386]. Moreover, soil aggregate stability can be measured through easy-to-implement and cost-effective methods in the laboratory following field sampling [386,387]. However, since aggregate stability strongly depends on SOC [74,75], it will present the same drawbacks for soil-health assessment than SOC. Moreover, regarding SOC, soil aggregate stability presents two additional drawbacks. First, it is more variable and, second, its determination lacks a worldwide standard procedure and measurement benchmark. Therefore, the different methods in use, though correlated with one another, provide values of different magnitude that hinder comparisons [388,389].
10.3. Soil Bulk Density and Infiltration Capacity
Soil bulk density and infiltration capacity have also the potential to be used as indicators of soil health because both correlate well with many of its healthy attributes, though bulk density stands out in this regard [378]. Both soil characteristics are remarkably related to each other with soil infiltration capacity decreasing, in general, as bulk density increases [390,391]. However, bulk density is easier to measure than infiltration capacity and it is less variable, thus making bulk density preferable as an indicator of soil health over infiltration capacity. Additionally, bulk density presents the advantage of being directly traceable to the primary standards of weight and length thus making the interpretation of bulk density measurements straightforward, and even easier if its counterpart, porosity, is calculated and presented alongside. As drawbacks, bulk density is more variable than other likely indicators of soil health, e.g., SOC. Moreover, in general, bulk density is slowly responsive to soil management, which makes soil-health monitoring through bulk density unpractical in the short to medium term.
10.4. Soil Chemistry: pH, Salinity, Sodicity, Nutrients, and Pollutants
Classical soil chemistry measurements such as pH, electrical conductivity as surrogate of salinity, sodicity, and the contents of nutrients and pollutants are traditional soil fertility indicators. Therefore, they address only the soil fertility and partially the quality and plant- and animal-health aspects of soil health. They do not directly integrate ecological soil attributes and, therefore, they lack basis to be used as standalone soil-health indicators, even though they may form part of soil-health indexes [371].
10.5. Soil Biology
For soil assessment on the basis of the health paradigm, a range of promising methods are being devised featuring biological parameters. On this point, researchers like Burns et al. [392] have proposed to identify soil bacterial communities, related to soil resources and vineyard management, through 16S rDNA fingerprinting as a biological indicator to monitor soil-quality or health in vineyard soils. However, most of these new soil-health assessment methods require further validation and enhancement concerning their relevance, scientific validity, practicality, and adaptability to local conditions [268]. Particularly, a major challenge in the practical application to soil health assessment of these the new proposed biological and biochemical indicators is the absence of reference values for interpreting them, as well as the integration of these with the standard procedures to provide a comprehensive assessment of a particular soil-health status [393]. The challenge of soil-health evaluation may be addressed through innovative biological approaches; however, they should also integrate standardized physical and chemical techniques to be able to disclose the functioning of vineyard soils and be, therefore, readily interpretable.
11. Conclusions
Vine agro-ecosystems face several challenges, including (i) soil compaction and erosion, (ii) SOM decline, (iii) biological simplification, and (iv) soil and water pollution due to the use of chemicals [26,393]. All these processes constrain the soil’s ability to fulfil its ecological functions, with both on-farm and off-farm detrimental effects. Consequently, these degradation processes significantly affect soil health, thus jeopardizing the long-run sustainability of vine cropping [26,394]. For example, a notable issue in vineyard soils is the accumulation of Cu due to the application of Cu-based pesticides for vine protection against fungal pathogens (Section 7). The Cu accumulation in vineyard soils harms the functionality of the soil biological community, hence disrupting soil fauna burrowing activities and specific nutrient cycling pathways [395,396]. Such effects, in turn, may make vine growing more dependent on tillage, mineral fertilizers, and herbicides. In particular, high levels of soil Cu will hamper the integration of grazing livestock in vineyards for sustainable weed control and nutrient recycling [293]. Although instances of Cu-phytotoxicity symptoms on vines have been rare, mainly due to their deep rooting systems [397], it is abundantly clear that addressing this soil-health issue is highly needed because it may prevent the development of other sustainable vineyard management practices such as cover cropping and the integration of livestock. Therefore, vineyards require more sustainable ways of applying Cu, alternative plant-protection treatments and remediation procedures for vineyard Cu-polluted soils, which must be all investigated further.
Vineyard management is influenced by various factors, including climate, water availability for irrigation, soil type, grapevine variety, rootstock, agro-environmental policies, and, most importantly, the decisions and attitudes of viticulturists [187]. Note that distinctively from other agricultural activities, viticulture often prioritizes enhancing harvest quality over yield, specifically in winemaking, with the goal of obtaining terroir wines by means of highlighting specific flavors that please consumers and earn their loyalty. However, as support for planetary health expands, contemporary wine consumers are increasingly more concerned about the environmental impacts of their wine choices. In this regard, the continued ability of vineyard soils to function as living environments that support plant, animal, and human health, as encompassed by the soil-health concept, has been shown to go through the adoption of conservative agricultural management practices [3]. However, despite the good deal of research on the application of these sustainable soil management practices to viticulture, doubts persist in practitioners about how to manage vineyards to optimize both soil health and terroir expression [138]. Therefore, the key challenge in modern viticulture of enhancing sustainability in grape production processes without endangering productivity and, importantly, harvest quality and hence profitability, is a major research field [398,399]. Complicating matters further is global warming, which challenges traditional vine-management guidelines and demands their modernization to adapt to its consequences [399,400,401]. The adoption of the soil health framework is therefore even more necessary to cope with the significant effects climate change is having on viticulture [399].
The concept of soil health is gradually emerging, and research and vineyard operation are adopting this new paradigm. However, conventional agricultural soil management is still dominant, and revolves around the soil-nutrient status, often overlooking the significance of many soil-biological characteristics for plant health and agricultural sustainability [402]. Consequently, viticulturists still mostly rely on analytical chemical procedures to assess both soil fertility and quality, as the latter has been traditionally understood. As the soil-health concept gains ground, standard procedures will have to be adapted or replaced, thus offering research and development opportunities in this regard.
Advances in soil health are grounded in four principal development lines: soil biology understanding, the integrated interpretation of soil data, and improved in-field measurements [403], in addition to the modelling of the soil–vine–water–atmosphere system. These advances should collectively promote sustainable practices that contribute to significantly reduce the exhaustion viticulture causes on the natural resources it depends upon, thereby mitigating the environmental impact of their activities and especially sustaining soil health [404]. Particularly, given the substantial impact of soil biodiversity in vineyards by means of its effects on several regulating services like pest control and SOM decomposition, through which it supports nutrient cycling and availability, and its eventual influence on both grape yield and quality [26], it is likely that soil-health research will increasingly focus on this area. Nonetheless, it remains clear that soil health should be approached holistically and established as a scientific field open to contributions from various disciplines.
Therefore, researchers in viticulture, as well as in other agronomic fields, should embrace soil health as a guiding principle to which they can contribute knowledge, viewing it not merely as a property to measure but as a well-established scientific field that, remarkably, can benefit from insights from various disciplines [64,82]. In the pursuit of improving the soil health of winegrowing systems so that vineyard soils fulfil their ecosystem services, as well as the harvest amount and quality aims of viticulturists, collaboration at the landscape level will be essential. This cooperation should involve winegrowers, other agriculturalists, researchers from various disciplines, policymakers, consumers, and media. Importantly, however, such collaboration should be undertaken without undermining the sovereignty and independence of local viticulturists in their quest for producing wines that meet the standards they have committed to [405].
Author Contributions
Conceptualization, F.V. and M.Á.O.; methodology, F.V. and M.Á.O.; investigation, F.V., R.L. and M.Á.O.; resources, F.V.; data curation, F.V., R.L. and M.Á.O.; writing—original draft preparation, F.V., R.L. and M.Á.O.; writing—review and editing, F.V., R.L. and M.Á.O.; visualization, F.V., R.L. and M.Á.O.; supervision, F.V.; project administration, F.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- International Organisation of Vine and Wine. Annual Assessment of the World Vine and Wine Sector in 2022; International Organisation of Vine and Wine: Dijon, France, 2023. [Google Scholar]
- Poore, J.; Nemecek, T. Reducing Food’s Environmental Impacts through Producers and Consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef]
- Cataldo, E.; Fucile, M.; Mattii, G.B. A Review: Soil Management, Sustainable Strategies and Approaches to Improve the Quality of Modern Viticulture. Agronomy 2021, 11, 2359. [Google Scholar] [CrossRef]
- International Health Conference. Constitution of the World Health Organization. 1946. Bull. World Health Organ. 2002, 80, 983–984. [Google Scholar]
- Minami, K. Soil Is a Living Substance. Soil Sci. Plant Nutr. 2021, 67, 26–30. [Google Scholar] [CrossRef]
- Doran, J.W.; Zeiss, M.R. Soil Health and Sustainability: Managing the Biotic Component of Soil Quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Yang, T.; Siddique, K.H.M.; Liu, K. Cropping Systems in Agriculture and Their Impact on Soil Health—A Review. Glob. Ecol. Conserv. 2020, 23, e01118. [Google Scholar] [CrossRef]
- Doran, J.W.; Parkin, T.B. Defining and Assessing Soil Quality. In Defining Soil Quality for a Sustainable Environment; Doran, J.W., Coleman, D.C., Bezdicek, D.F., Stewart, B.A., Eds.; SSSA: Madison, WI, USA, 2015; pp. 3–21. ISBN 9780891189305. [Google Scholar]
- Blum, W.E.H. Functions of Soil for Society and the Environment. Rev. Environ. Sci. Biotechnol. 2005, 4, 75–79. [Google Scholar] [CrossRef]
- Riesenfeld, C.S.; Goodman, R.M.; Handelsman, J. Uncultured Soil Bacteria Are a Reservoir of New Antibiotic Resistance Genes. Environ. Microbiol. 2004, 6, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Van Goethem, M.W.; Pierneef, R.; Bezuidt, O.K.I.; Van De Peer, Y.; Cowan, D.A.; Makhalanyane, T.P. A Reservoir of “historical” Antibiotic Resistance Genes in Remote Pristine Antarctic Soils. Microbiome 2018, 6, 40. [Google Scholar] [CrossRef] [PubMed]
- van de Meent, D.; de Bruijn, J.H.M. A Modeling Procedure to Evaluate the Coherence of Independently Derived Environmental Quality Objectives for Air, Water and Soil. Environ. Toxicol. Chem. 1995, 14, 177–186. [Google Scholar] [CrossRef]
- Karlen, D.L.; Mausbach, M.J.; Doran, J.W.; Cline, R.G.; Harris, R.F.; Schuman, G.E. Soil Quality: A Concept, Definition, and Framework for Evaluation (A Guest Editorial). Soil Sci. Soc. Am. J. 1997, 61, 4–10. [Google Scholar] [CrossRef]
- Calistri, P.; Iannetti, S.; Danzetta, M.L.; Narcisi, V.; Cito, F.; Di Sabatino, D.; Bruno, R.; Sauro, F.; Atzeni, M.; Carvelli, A.; et al. The Components of ‘One World-One Health’ Approach. Transbound. Emerg. Dis. 2013, 60, 4–13. [Google Scholar] [CrossRef]
- United Nations. FAOSTAT. Available online: https://www.fao.org/faostat/en/#home (accessed on 6 December 2023).
- Ohno, T.; Hettiarachchi, G.M. Soil Chemistry and the One Health Initiative: Introduction to the Special Section. J. Environ. Qual. 2018, 47, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- Van Bruggen, A.H.C.; Goss, E.M.; Havelaar, A.; van Diepeningen, A.D.; Finckh, M.R.; Morris, J.G. One Health-Cycling of Diverse Microbial Communities as a Connecting Force for Soil, Plant, Animal, Human and Ecosystem Health. Sci. Total Environ. 2019, 664, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; van der Heijden, M.G.A. Soil Microbiomes and One Health. Nat. Rev. Microbiol. 2022, 21, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Aldana, D.K.; Dhama, K.; Rodriguez-Morales, A.J. Revisiting the One Health Approach in the Context of COVID-19: A Look into the Ecology of This Emerging Disease. Adv. Anim. Vet. Sci. 2020, 8, 234–237. [Google Scholar] [CrossRef]
- Abbott, L.; Murphy, D. Soil Biological Fertility: A Key to Sustainable Land Use in Agriculture; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Stirling, G.; Hayden, H.; Pattison, T.; Stirling, M. Soil Health, Soil Biology, Soilborne Diseases and Sustainable Agriculture; CSIRO: Canberra, Australia, 2016. [Google Scholar]
- Hatfield, J.L.; Walthall, C.L. Soil Biological Fertility: Foundation for the Next Revolution in Agriculture? Commun. Soil Sci. Plant Anal. 2015, 46, 753–762. [Google Scholar] [CrossRef]
- Blanco-Pérez, R.; Vicente-Díez, I.; Ramos-Sáez de Ojer, J.L.; Marco-Mancebón, V.S.; Pérez-Moreno, I.; Campos-Herrera, R. Organic Viticulture Enhanced the Activity of Native Entomopathogenic Nematodes in DOCa Rioja Soils (North of Spain). Agric. Ecosyst. Environ. 2022, 332, 107931. [Google Scholar] [CrossRef]
- Rodrigo-Comino, J. Five Decades of Soil Erosion Research in “Terroir”. The State-of-the-Art. Earth Sci. Rev. 2018, 179, 436–447. [Google Scholar] [CrossRef]
- Karimi, B.; Cahurel, J.Y.; Gontier, L.; Charlier, L.; Chovelon, M.; Mahé, H.; Ranjard, L. A Meta-Analysis of the Ecotoxicological Impact of Viticultural Practices on Soil Biodiversity. Environ. Chem. Lett. 2020, 18, 1947–1966. [Google Scholar] [CrossRef]
- Giffard, B.; Winter, S.; Guidoni, S.; Nicolai, A.; Castaldini, M.; Cluzeau, D.; Coll, P.; Cortet, J.; Le Cadre, E.; d’Errico, G.; et al. Vineyard Management and Its Impacts on Soil Biodiversity, Functions, and Ecosystem Services. Front. Ecol. Evol. 2022, 10, 850272. [Google Scholar] [CrossRef]
- Marques, M.J.; Bienes, R.; Cuadrado, J.; Ruiz-Colmenero, M.; Barbero-Sierra, C.; Velasco, A. Analysing Perceptions Attitudes and Responses of Winegrowers about Sustainable Land Management in Central Spain. Land Degrad. Dev. 2015, 26, 458–467. [Google Scholar] [CrossRef]
- Khmelevtsova, L.E.; Sazykin, I.S.; Azhogina, T.N.; Sazykina, M.A. Influence of Agricultural Practices on Bacterial Community of Cultivated Soils. Agriculture 2022, 12, 371. [Google Scholar] [CrossRef]
- Wu, T.; Milner, H.; Díaz-Pérez, J.C.; Ji, P. Effects of Soil Management Practices on Soil Microbial Communities and Development of Southern Blight in Vegetable Production. Appl. Soil Ecol. 2015, 91, 58–67. [Google Scholar] [CrossRef]
- García-Orenes, F.; Guerrero, C.; Roldán, A.; Mataix-Solera, J.; Cerdà, A.; Campoy, M.; Zornoza, R.; Bárcenas, G.; Caravaca, F. Soil Microbial Biomass and Activity under Different Agricultural Management Systems in a Semiarid Mediterranean Agroecosystem. Soil Tillage Res. 2010, 109, 110–115. [Google Scholar] [CrossRef]
- Pampulha, M.E.; Oliveira, A. Impact of an Herbicide Combination of Bromoxynil and Prosulfuron on Soil Microorganisms. Curr. Microbiol. 2006, 53, 238–243. [Google Scholar] [CrossRef]
- Young, I.M.; Crawford, J.W. Interactions and Self-Organization in the Soil-Microbe Complex. Science 2004, 304, 1634–1637. [Google Scholar] [CrossRef] [PubMed]
- Marín, D.; Armengol, J.; Carbonell-Bejerano, P.; Escalona, J.M.; Gramaje, D.; Hernández-Montes, E.; Intrigliolo, D.S.; Martínez-Zapater, J.M.; Medrano, H.; Mirás-Avalos, J.M.; et al. Challenges of Viticulture Adaptation to Global Change: Tackling the Issue from the Roots. Aust. J. Grape Wine Res. 2020, 27, 8–25. [Google Scholar] [CrossRef]
- Darriaut, R.; Lailheugue, V.; Masneuf-Pomarède, I.; Marguerit, E.; Martins, G.; Compant, S.; Ballestra, P.; Upton, S.; Ollat, N.; Lauvergeat, V. Grapevine Rootstock and Soil Microbiome Interactions: Keys for a Resilient Viticulture. Hortic. Res. 2022, 9, uhac019. [Google Scholar] [CrossRef] [PubMed]
- Agnelli, A.; Bol, R.; Trumbore, S.E.; Dixon, L.; Cocco, S.; Corti, G. Carbon and Nitrogen in Soil and Vine Roots in Harrowed and Grass-Covered Vineyards. Agric. Ecosyst. Environ. 2014, 193, 70–82. [Google Scholar] [CrossRef]
- Ritz, K.; Harris, J.; Murray, P. The Role of Soil Biota in Soil Fertility and Quality, and Approaches to Influencing Soil Communities to Enhance Delivery of These Functions; Department for Environment, Food and Rural Affairs: London, UK, 2010. [Google Scholar]
- Bettenfeld, P.; Cadena i Canals, J.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.E.; Trouvelot, S. The Microbiota of the Grapevine Holobiont: A Key Component of Plant Health. J. Adv. Res. 2022, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S.; et al. The Soil Microbiome Influences Grapevine-Associated Microbiota. mBio 2015, 6, e02527-14. [Google Scholar] [CrossRef] [PubMed]
- Custódio, V.; Gonin, M.; Stabl, G.; Bakhoum, N.; Oliveira, M.M.; Gutjahr, C.; Castrillo, G. Sculpting the Soil Microbiota. Plant J. 2022, 109, 508–522. [Google Scholar] [CrossRef]
- Pingel, M.; Reineke, A.; Leyer, I. A 30-Years Vineyard Trial: Plant Communities, Soil Microbial Communities and Litter Decomposition Respond More to Soil Treatment than to N Fertilization. Agric. Ecosyst. Environ. 2019, 272, 114–125. [Google Scholar] [CrossRef]
- Trouvelot, S.; Bonneau, L.; Redecker, D.; van Tuinen, D.; Adrian, M.; Wipf, D. Arbuscular Mycorrhiza Symbiosis in Viticulture: A Review. Agron. Sustain. Dev. 2015, 35, 1449–1467. [Google Scholar] [CrossRef]
- Lemtiri, A.; Colinet, G.; Alabi, T.; Cluzeau, D.; Zirbes, L.; Haubruge, É.; Francis, F. Impacts of Earthworms on Soil Components and Dynamics. A Review. Biotechnol. Agron. Soc. Environ. 2014, 18, 121–133. [Google Scholar]
- Neemisha. Role of Soil Organisms in Maintaining Soil Health, Ecosystem Functioning, and Sustaining Agricultural Production. In Soil Health; Springer: Berlin/Heidelberg, Germany, 2020; pp. 313–335. [Google Scholar]
- Tian, T.; Reverdy, A.; She, Q.; Sun, B.; Chai, Y. The Role of Rhizodeposits in Shaping Rhizomicrobiome. Environ. Microbiol. Rep. 2019, 12, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, M.P.; Santhoshkannada, A.N. Role of Rhizomicrobiome in Maintaining Soil Fertility and Crop Production. In Soil Health; Springer: Berlin/Heidelberg, Germany, 2020; pp. 373–401. [Google Scholar]
- Vaudour, E.; Costantini, E.; Jones, G.V.; Mocali, S. An Overview of the Recent Approaches to Terroir Functional Modelling, Footprinting and Zoning. Soil 2015, 1, 287–312. [Google Scholar] [CrossRef]
- Mcnear, D., Jr. The Rhizosphere-Roots, Soil and Everything in Between. Nat. Educ. Knowl. 2013, 4, 1. [Google Scholar]
- Whalley, W.R.; Riseley, B.; Leeds-Harrison, P.B.; Bird, N.R.A.; Leech, P.K.; Adderley, W.P. Structural Differences between Bulk and Rhizosphere Soil. Eur. J. Soil Sci. 2005, 56, 353–360. [Google Scholar] [CrossRef]
- Youssef, R.A.; Chino, M. Root-Induced Changes in the Rhizosphere of Plants. II. Distribution of Heavy Metals across the Rhizosphere in Soils. Soil Sci. Plant Nutr. 1989, 35, 609–621. [Google Scholar] [CrossRef]
- Youssef, R.A.; Chino, M. Root-Induced Changes in the Rhizosphere of Plants. I. PH Changes in Relation to the Bulk Soil. Soil Sci. Plant Nutr. 1989, 35, 461–468. [Google Scholar] [CrossRef]
- Wang, L.; Rengel, Z.; Zhang, K.; Jin, K.; Lyu, Y.; Zhang, L.; Cheng, L.; Zhang, F.; Shen, J. Ensuring Future Food Security and Resource Sustainability: Insights into the Rhizosphere. iScience 2022, 25, 104168. [Google Scholar] [CrossRef] [PubMed]
- Gianfreda, L. Enzymes of Importance to Rhizosphere Processes. J. Soil Sci. Plant Nutr. 2015, 15, 283–306. [Google Scholar] [CrossRef]
- Custos, J.-M.; Moyne, C.; Sterckeman, T. How Root Nutrient Uptake Affects Rhizosphere PH: A Modelling Study. Geoderma 2020, 369, 114314. [Google Scholar] [CrossRef]
- Afridi, M.S.; Fakhar, A.; Kumar, A.; Ali, S.; Medeiros, F.H.V.; Muneer, M.A.; Ali, H.; Saleem, M. Harnessing Microbial Multitrophic Interactions for Rhizosphere Microbiome Engineering. Microbiol. Res. 2022, 265, 127199. [Google Scholar] [CrossRef]
- Bao, L.; Sun, B.; Wei, Y.; Xu, N.; Zhang, S.; Gu, L.; Bai, Z. Grape Cultivar Features Differentiate the Grape Rhizosphere Microbiota. Plants 2022, 11, 1111. [Google Scholar] [CrossRef]
- Dries, L.; Bussotti, S.; Pozzi, C.; Kunz, R.; Schnell, S.; Löhnertz, O.; Vortkamp, A. Rootstocks Shape Their Microbiome—Bacterial Communities in the Rhizosphere of Different Grapevine Rootstocks. Microorganisms 2021, 9, 822. [Google Scholar] [CrossRef]
- Dries, L.; Hendgen, M.; Schnell, S.; Löhnertz, O.; Vortkamp, A. Rhizosphere Engineering: Leading towards a Sustainable Viticulture? OENO One 2021, 55, 353–363. [Google Scholar] [CrossRef]
- Baweja, P.; Kumar, S.; Kumar, G. Fertilizers and Pesticides: Their Impact on Soil Health and Environment. In Soil Health; Springer: Berlin/Heidelberg, Germany, 2020; pp. 265–285. [Google Scholar]
- Schreiner, R.P. Spatial and Temporal Variation of Roots, Arbuscular Mycorrhizal Fungi, and Plant and Soil Nutrients in a Mature Pinot Noir (Vitis vinifera L.) Vineyard in Oregon, USA. Plant Soil 2005, 276, 219–234. [Google Scholar] [CrossRef]
- Aguilera, P.; Ortiz, N.; Becerra, N.; Turrini, A.; Gaínza-Cortés, F.; Silva-Flores, P.; Aguilar-Paredes, A.; Romero, J.K.; Jorquera-Fontena, E.; Mora, M.d.L.L.; et al. Application of Arbuscular Mycorrhizal Fungi in Vineyards: Water and Biotic Stress Under a Climate Change Scenario: New Challenge for Chilean Grapevine Crop. Front. Microbiol. 2022, 13, 826571. [Google Scholar] [CrossRef]
- Gazioglu Sensoy, R. Effects of Arbuscular Mycorrhizal Fungus (AMF) and Whey Applications on the Grapevine (Vitis Vinifera L.) Cutting Exposed to Salt Stress. J. Elem. 2022, 27, 507–519. [Google Scholar] [CrossRef]
- Cardinale, M.; Minervini, F.; De Angelis, M.; Papadia, P.; Migoni, D.; Dimaglie, M.; Dinu, D.G.; Quarta, C.; Selleri, F.; Caccioppola, A.; et al. Vineyard Establishment under Exacerbated Summer Stress: Effects of Mycorrhization on Rootstock Agronomical Parameters, Leaf Element Composition and Root-Associated Bacterial Microbiota. Plant Soil 2022, 478, 613–634. [Google Scholar] [CrossRef]
- Lumini, E.; Orgiazzi, A.; Borriello, R.; Bonfante, P.; Bianciotto, V. Disclosing Arbuscular Mycorrhizal Fungal Biodiversity in Soil through a Land-use Gradient Using a Pyrosequencing Approach. Environ. Microbiol. 2010, 12, 2165–2179. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The Contentious Nature of Soil Organic Matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Kögel-Knabner, I. The Macromolecular Organic Composition of Plant and Microbial Residues as Inputs to Soil Organic Matter: Fourteen Years On. Soil Biol. Biochem. 2017, 105, A3–A8. [Google Scholar] [CrossRef]
- Stockmann, U.; Padarian, J.; McBratney, A.; Minasny, B.; de Brogniez, D.; Montanarella, L.; Hong, S.Y.; Rawlins, B.G.; Field, D.J. Global Soil Organic Carbon Assessment. Glob. Food Secur. 2015, 6, 9–16. [Google Scholar] [CrossRef]
- Hengl, T.; Mendes de Jesus, J.; Heuvelink, G.B.M.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global Gridded Soil Information Based on Machine Learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed]
- Loveland, P.; Webb, J. Is There a Critical Level of Organic Matter in the Agricultural Soils of Temperate Regions: A Review. Soil Tillage Res. 2003, 70, 1–18. [Google Scholar] [CrossRef]
- Murphy, B.W. Impact of Soil Organic Matter on Soil Properties—A Review with Emphasis on Australian Soils. Soil Res. 2015, 53, 605. [Google Scholar] [CrossRef]
- Foley, J.A. The Sensitivity of the Terrestrial Biosphere to Climatic Change: A Simulation of the Middle Holocene. Global Biogeochem. Cycles 1994, 8, 505–525. [Google Scholar] [CrossRef]
- Ruddiman, W.F. The Anthropogenic Greenhouse Era Began Thousands of Years Ago. Clim. Change 2003, 61, 261–293. [Google Scholar] [CrossRef]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Feller, C.; Manlay, R.J.; Swift, M.J.; Bernoux, M. Functions, Services and Value of Soil Organic Matter for Human Societies and the Environment: A Historical Perspective. Geol. Soc. Lond. Spec. Publ. 2006, 266, 9–22. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Organic Matter and Water-stable Aggregates in Soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Kong, A.Y.Y.; Six, J.; Bryant, D.C.; Denison, R.F.; van Kessel, C. The Relationship between Carbon Input, Aggregation, and Soil Organic Carbon Stabilization in Sustainable Cropping Systems. Soil Sci. Soc. Am. J. 2005, 69, 1078–1085. [Google Scholar] [CrossRef]
- Chen, H.; Hou, R.; Gong, Y.; Li, H.; Fan, M.; Kuzyakov, Y. Effects of 11 Years of Conservation Tillage on Soil Organic Matter Fractions in Wheat Monoculture in Loess Plateau of China. Soil Tillage Res. 2009, 106, 85–94. [Google Scholar] [CrossRef]
- Mustafa, A.; Minggang, X.; Ali Shah, S.A.; Abrar, M.M.; Nan, S.; Baoren, W.; Zejiang, C.; Saeed, Q.; Naveed, M.; Mehmood, K.; et al. Soil Aggregation and Soil Aggregate Stability Regulate Organic Carbon and Nitrogen Storage in a Red Soil of Southern China. J. Environ. Manag. 2020, 270, 110894. [Google Scholar] [CrossRef]
- Xu, N.; Amgain, N.R.; Rabbany, A.; Capasso, J.; Korus, K.; Swanson, S.; Bhadha, J.H. Interaction of Soil Health Indicators to Different Regenerative Farming Practices on Mineral Soils. Agrosystems Geosci. Environ. 2022, 5, e20243. [Google Scholar] [CrossRef]
- Cresswell, H.P.; Kirkegaard, J.A. Subsoil Amelioration by Plant-Roots—The Process and the Evidence. Aust. J. Soil Res. 1995, 33, 221. [Google Scholar] [CrossRef]
- Yunusa, I.A.M.; Newton, P.J. Plants for Amelioration of Subsoil Constraints and Hydrological Control: The Primer-Plant Concept. Plant Soil 2003, 257, 261–281. [Google Scholar] [CrossRef]
- Lavelle, P.; Spain, A.; Blouin, M.; Brown, G.; Decaëns, T.; Grimaldi, M.; Jiménez, J.J.; McKey, D.; Mathieu, J.; Velasquez, E.; et al. Ecosystem Engineers in a Self-Organized Soil: A Review of Concepts and Future Research Questions. Soil Sci. 2016, 181, 91–109. [Google Scholar] [CrossRef]
- Lehmann, A.; Leifheit, E.F.; Rillig, M.C. Mycorrhizas and Soil Aggregation. In Mycorrhizal Mediation of Soil; Elsevier: Amsterdam, The Netherlands, 2017; pp. 241–262. [Google Scholar]
- Wilpiszeski, R.L.; Aufrecht, J.A.; Retterer, S.T.; Sullivan, M.B.; Graham, D.E.; Pierce, E.M.; Zablocki, O.D.; Palumbo, A.V.; Elias, D.A. Soil Aggregate Microbial Communities: Towards Understanding Microbiome Interactions at Biologically Relevant Scales. Appl. Environ. Microbiol. 2019, 85, e00324-19. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, J.; Zhang, C.; Xin, X.; Li, H. The Coupling Effects of Soil Organic Matter and Particle Interaction Forces on Soil Aggregate Stability. Soil Tillage Res. 2017, 174, 251–260. [Google Scholar] [CrossRef]
- Li, S.; Wang, B.; Zhang, X.; Wang, H.; Yi, Y.; Huang, X.; Gao, X.; Zhu, P.; Han, W. Soil Particle Aggregation and Aggregate Stability Associated with Ion Specificity and Organic Matter Content. Geoderma 2023, 429, 116285. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, X.; Liu, Y. Pore-Scale Simulation of Gas Diffusion in Unsaturated Soil Aggregates: Accuracy of the Dusty-Gas Model and the Impact of Saturation. Geoderma 2017, 303, 196–203. [Google Scholar] [CrossRef]
- Hu, W.; Jiang, Y.; Chen, D.; Lin, Y.; Han, Q.; Cui, Y. Impact of Pore Geometry and Water Saturation on Gas Effective Diffusion Coefficient in Soil. Appl. Sci. 2018, 8, 2097. [Google Scholar] [CrossRef]
- Prifling, B.; Weber, M.; Ray, N.; Prechtel, A.; Phalempin, M.; Schlüter, S.; Vetterlein, D.; Schmidt, V. Quantifying the Impact of 3D Pore Space Morphology on Soil Gas Diffusion in Loam and Sand. Transp. Porous Media 2023, 149, 501–527. [Google Scholar] [CrossRef]
- Ehlers, W. Observations on Earthworm Channels and Infiltration on Tilled and Untilled Loess Soil. Soil Sci. 1975, 119, 242–249. [Google Scholar] [CrossRef]
- Franzluebbers, A.J. Water Infiltration and Soil Structure Related to Organic Matter and Its Stratification with Depth. Soil Tillage Res. 2002, 66, 197–205. [Google Scholar] [CrossRef]
- Lipiec, J.; Kuś, J.; Słowińska-Jurkiewicz, A.; Nosalewicz, A. Soil Porosity and Water Infiltration as Influenced by Tillage Methods. Soil Tillage Res. 2006, 89, 210–220. [Google Scholar] [CrossRef]
- Kirkby, C.A.; Kirkegaard, J.A.; Richardson, A.E.; Wade, L.J.; Blanchard, C.; Batten, G. Stable Soil Organic Matter: A Comparison of C:N:P:S Ratios in Australian and Other World Soils. Geoderma 2011, 163, 197–208. [Google Scholar] [CrossRef]
- Tipping, E.; Somerville, C.J.; Luster, J. The C:N:P:S Stoichiometry of Soil Organic Matter. Biogeochemistry 2016, 130, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Coonan, E.C.; Kirkby, C.A.; Kirkegaard, J.A.; Amidy, M.R.; Strong, C.L.; Richardson, A.E. Microorganisms and Nutrient Stoichiometry as Mediators of Soil Organic Matter Dynamics. Nutr. Cycl. Agroecosyst. 2020, 117, 273–298. [Google Scholar] [CrossRef]
- Schulten, H.-R.; Schnitzer, M. The Chemistry of Soil Organic Nitrogen: A Review. Biol. Fertil. Soils 1997, 26, 1–15. [Google Scholar] [CrossRef]
- Schroth, A.W.; Bostick, B.C.; Graham, M.; Kaste, J.M.; Mitchell, M.J.; Friedland, A.J. Sulfur Species Behavior in Soil Organic Matter during Decomposition. J. Geophys. Res. Biogeosci. 2007, 112, G04011. [Google Scholar] [CrossRef]
- Spohn, M. Increasing the Organic Carbon Stocks in Mineral Soils Sequesters Large Amounts of Phosphorus. Glob. Change Biol. 2020, 26, 4169–4177. [Google Scholar] [CrossRef] [PubMed]
- Parfitt, R.L.; Giltrap, D.J.; Whitton, J.S. Contribution of Organic Matter and Clay Minerals to the Cation Exchange Capacity of Soils. Commun. Soil Sci. Plant Anal. 1995, 26, 1343–1355. [Google Scholar] [CrossRef]
- Oorts, K.; Vanlauwe, B.; Merckx, R. Cation Exchange Capacities of Soil Organic Matter Fractions in a Ferric Lixisol with Different Organic Matter Inputs. Agric. Ecosyst. Environ. 2003, 100, 161–171. [Google Scholar] [CrossRef]
- Fageria, N.; Baligar, V.; Clark, R. Micronutrients in Crop Production. Adv. Agron. 2002, 77, 185–268. [Google Scholar]
- Lindsay, W.L. Iron Oxide Solubilization by Organic Matter and Its Effect on Iron Availability. Plant Soil 1991, 130, 27–34. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Naresh, R.K.; Mandal, A.; Singh, R.; Dhaliwal, M.K. Dynamics and Transformations of Micronutrients in Agricultural Soils as Influenced by Organic Matter Build-up: A Review. Environ. Sustain. Indic. 2019, 1–2, 100007. [Google Scholar] [CrossRef]
- Merino, C.; Kuzyakov, Y.; Godoy, K.; Jofré, I.; Nájera, F.; Matus, F. Iron-Reducing Bacteria Decompose Lignin by Electron Transfer from Soil Organic Matter. Sci. Total Environ. 2021, 761, 143194. [Google Scholar] [CrossRef]
- Jones, D.L. Organic Acids in the Rhizosphere-A Critical Review. Plant Soil 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Goldberg, S. Reactions of Boron with Soils. Plant Soil 1997, 193, 35–48. [Google Scholar] [CrossRef]
- Sauvé, S.; Hendershot, W.; Allen, H.E. Solid-Solution Partitioning of Metals in Contaminated Soils: Dependence on PH, Total Metal Burden, and Organic Matter. Environ. Sci. Technol. 2000, 34, 1125–1131. [Google Scholar] [CrossRef]
- Wichard, T.; Mishra, B.; Myneni, S.C.B.; Bellenger, J.-P.; Kraepiel, A.M.L. Storage and Bioavailability of Molybdenum in Soils Increased by Organic Matter Complexation. Nat. Geosci. 2009, 2, 625–629. [Google Scholar] [CrossRef]
- Tolls, J. Sorption of Veterinary Pharmaceuticals in Soils: A Review. Environ. Sci. Technol. 2001, 35, 3397–3406. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.J.; Gustafsson, O.; Kurt-Karakus, P.; Breivik, K.; Steinnes, E.; Jones, K.C. Relationships between Organic Matter, Black Carbon and Persistent Organic Pollutants in European Background Soils: Implications for Sources and Environmental Fate. Environ. Pollut. 2008, 156, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Chien, S.-W.C.; Chen, S.-H.; Li, C.-J. Effect of Soil PH and Organic Matter on the Adsorption and Desorption of Pentachlorophenol. Environ. Sci. Pollut. Res. 2017, 25, 5269–5279. [Google Scholar] [CrossRef]
- Cui, S.; Qi, Y.; Zhu, Q.; Wang, C.; Sun, H. A Review of the Influence of Soil Minerals and Organic Matter on the Migration and Transformation of Sulfonamides. Sci. Total Environ. 2023, 861, 160584. [Google Scholar] [CrossRef]
- Lo, K.S.L.; Yang, W.F.; Lin, Y.C. Effects of Organic Matter on the Specific Adsorption of Heavy Metals by Soil. Toxicol. Environ. Chem. 1992, 34, 139–153. [Google Scholar] [CrossRef]
- Salati, S.; Quadri, G.; Tambone, F.; Adani, F. Fresh Organic Matter of Municipal Solid Waste Enhances Phytoextraction of Heavy Metals from Contaminated Soil. Environ. Pollut. 2010, 158, 1899–1906. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Yang, X.; Zhang, J.; Lin, Z.; Zhao, B. Microbial Community Structure and Functional Metabolic Diversity Are Associated with Organic Carbon Availability in an Agricultural Soil. J. Integr. Agric. 2015, 14, 2500–2511. [Google Scholar] [CrossRef]
- Zhang, L.; Yan, H.; Ren, T.; Li, S.; Yang, Y.; Peng, G.; Yu, J.; Liu, G. Effects of Organic Matter on Nutrient, Enzyme Activity and Functional Diversity of Microbial Community in Tobacco Planting Soil. Acta Tabacaria Sin. 2019, 25, 55–62. [Google Scholar]
- Johnson, D.C.; Teague, R.; Apfelbaum, S.; Thompson, R.; Byck, P. Adaptive Multi-Paddock Grazing Management’s Influence on Soil Food Web Community Structure for: Increasing Pasture Forage Production, Soil Organic Carbon, and Reducing Soil Respiration Rates in Southeastern USA Ranches. PeerJ 2022, 10, e13750. [Google Scholar] [CrossRef] [PubMed]
- Sokol, N.W.; Kuebbing, S.E.; Karlsen-Ayala, E.; Bradford, M.A. Evidence for the Primacy of Living Root Inputs, Not Root or Shoot Litter, in Forming Soil Organic Carbon. New Phytol. 2018, 221, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Chaney, K.; Swift, R.S. The Influence of Organic Matter on Aggregate Stability in Some British Soils. J. Soil Sci. 1984, 35, 223–230. [Google Scholar] [CrossRef]
- Chenu, C.; Le Bissonnais, Y.; Arrouays, D. Organic Matter Influence on Clay Wettability and Soil Aggregate Stability. Soil Sci. Soc. Am. J. 2000, 64, 1479–1486. [Google Scholar] [CrossRef]
- Le Bissonnais, Y.; Blavet, D.; De Noni, G.; Laurent, J.-Y.; Asseline, J.; Chenu, C. Erodibility of Mediterranean Vineyard Soils: Relevant Aggregate Stability Methods and Significant Soil Variables. Eur. J. Soil Sci. 2007, 58, 188–195. [Google Scholar] [CrossRef]
- Burras, C.; Nyasimi, M.; Butler, L. Soils, Human Health, and Wealth: A Complicated Relationship. In Soils and Human Health; CRC Press: Boca Raton, FL, USA, 2013; pp. 215–226. [Google Scholar]
- Gu, B.; Chen, D.; Yang, Y.; Vitousek, P.; Zhu, Y.-G. Soil-Food-Environment-Health Nexus for Sustainable Development. Research 2021, 2021, 9804807. [Google Scholar] [CrossRef]
- Parras-Alcántara, L.; Martín-Carrillo, M.; Lozano-García, B. Impacts of Land Use Change in Soil Carbon and Nitrogen in a Mediterranean Agricultural Area (Southern Spain). Solid Earth 2013, 4, 167–177. [Google Scholar] [CrossRef]
- Jakšić, S.; Ninkov, J.; Milić, S.; Vasin, J.; Banjac, D.; Jakšić, D.; Živanov, M. The State of Soil Organic Carbon in Vineyards as Affected by Soil Types and Fertilization Strategies (Tri Morave Region, Serbia). Agronomy 2020, 11, 9. [Google Scholar] [CrossRef]
- Maetens, W.; Vanmaercke, M.; Poesen, J.; Jankauskas, B.; Jankauskiene, G.; Ionita, I. Effects of Land Use on Annual Runoff and Soil Loss in Europe and the Mediterranean: A Meta-Analysis of Plot Data. Prog. Phys. Geogr. Earth Environ. 2012, 36, 599–653. [Google Scholar] [CrossRef]
- Prosdocimi, M.; Cerdà, A.; Tarolli, P. Soil Water Erosion on Mediterranean Vineyards: A Review. Catena 2016, 141, 1–21. [Google Scholar] [CrossRef]
- Rodrigo Comino, J.; Senciales, J.M.; Ramos, M.C.; Martínez-Casasnovas, J.A.; Lasanta, T.; Brevik, E.C.; Ries, J.B.; Ruiz Sinoga, J.D. Understanding Soil Erosion Processes in Mediterranean Sloping Vineyards (Montes de Málaga, Spain). Geoderma 2017, 296, 47–59. [Google Scholar] [CrossRef]
- Gristina, L.; Novara, A.; Minacapilli, M. Rethinking Vineyard Ground Management to Counter Soil Tillage Erosion. Soil Tillage Res. 2022, 217, 105275. [Google Scholar] [CrossRef]
- Kibblewhite, M.G.; Ritz, K.; Swift, M.J. Soil Health in Agricultural Systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 685–701. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, J.M. The Effects of Land Uses on Soil Erosion in Spain: A Review. Catena 2010, 81, 1–11. [Google Scholar] [CrossRef]
- Lozano-Parra, J.; Maldonado-Clavelle, C.F.; Caballero-Calvo, A.; Pulido-Fernández, M.; Gyasi-Agyei, Y.; Rodrigo-Comino, J. Estimation of Soil Mobilization Rates by a Rainy Period and Intense Tillage Practices in Vineyards—A Case Study in the Maule Region (Chile). Hydrol. Process. 2023, 37, e14829. [Google Scholar] [CrossRef]
- Martínez-Casasnovas, J.A.; Ramos, M.C. Soil Alteration Due to Erosion, Ploughing and Levelling of Vineyards in North East Spain. Soil Use Manag. 2009, 25, 183–192. [Google Scholar] [CrossRef]
- Mishra, U.; Lal, R.; Slater, B.; Calhoun, F.; Liu, D.; Van Meirvenne, M. Predicting Soil Organic Carbon Stock Using Profile Depth Distribution Functions and Ordinary Kriging. Soil Sci. Soc. Am. J. 2009, 73, 614–621. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, G.; Zhao, Q.; Lu, Q.; Jia, J.; Cui, B.; Liu, X. Depth-Distribution Patterns and Control of Soil Organic Carbon in Coastal Salt Marshes with Different Plant Covers. Sci. Rep. 2016, 6, 34835. [Google Scholar] [CrossRef]
- Murphy, B.W.; Wilson, B.R.; Koen, T. Mathematical Functions to Model the Depth Distribution of Soil Organic Carbon in a Range of Soils from New South Wales, Australia under Different Land Uses. Soil Syst. 2019, 3, 46. [Google Scholar] [CrossRef]
- Visconti, F.; Intrigliolo, D.S.; Mirás-Avalos, J.M. Effects of the Annual Nitrogen Fertilization Rate on Vine Performance and Grape Quality for Winemaking: Insights from a Meta-Analysis. Aust. J. Grape Wine Res. 2023, 2023, 7989254. [Google Scholar] [CrossRef]
- Baldock, J.; Skjemstad, J. Soil Organic Carbon/Soil Organic Matter. In Soil Analysis: An Interpretation Manual; Peverill, K., Sparrow, L., Reuter, D., Eds.; CSIRO Publishing: Melbourne, Australia, 1999; pp. 159–170. [Google Scholar]
- Lazcano, C.; Decock, C.; Wilson, S.G. Defining and Managing for Healthy Vineyard Soils, Intersections With the Concept of Terroir. Front. Environ. Sci. 2020, 8, 68. [Google Scholar] [CrossRef]
- Müller-Nedebock, D.; Chaplot, V. Soil Carbon Losses by Sheet Erosion: A Potentially Critical Contribution to the Global Carbon Cycle. Earth Surf. Process Landf. 2015, 40, 1803–1813. [Google Scholar] [CrossRef]
- Li, H.; Liu, G.; Gu, J.; Chen, H.; Shi, H.; Abd Elbasit, M.A.M.; Hu, F. Response of Soil Aggregate Disintegration to the Different Content of Organic Carbon and Its Fractions during Splash Erosion. Hydrol. Process. 2021, 35, e14060. [Google Scholar] [CrossRef]
- Vicente-Vicente, J.L.; García-Ruiz, R.; Francaviglia, R.; Aguilera, E.; Smith, P. Soil Carbon Sequestration Rates under Mediterranean Woody Crops Using Recommended Management Practices: A Meta-Analysis. Agric. Ecosyst. Environ. 2016, 235, 204–214. [Google Scholar] [CrossRef]
- Gómez de Barreda, D.; Bautista, I.; Castell, V.; Lidón, A. Rice Straw Mulch Installation in a Vineyard Improves Weed Control and Modifies Soil Characteristics. Agronomy 2023, 13, 3068. [Google Scholar] [CrossRef]
- Qin, T.; Wang, L.; Zhao, J.; Zhou, G.; Li, C.; Guo, L.; Jiang, G. Effects of Straw Mulching Thickness on the Soil Health in a Temperate Organic Vineyard. Agriculture 2022, 12, 1751. [Google Scholar] [CrossRef]
- Morlat, R.; Chaussod, R. Long-Term Additions of Organic Amendments in a Loire Valley Vineyard. I. Effects on Properties of a Calcareous Sandy Soil. Am. J. Enol. Vitic. 2008, 59, 353–363. [Google Scholar] [CrossRef]
- Gaiotti, F.; Marcuzzo, P.; Belfiore, N.; Lovat, L.; Fornasier, F.; Tomasi, D. Influence of Compost Addition on Soil Properties, Root Growth and Vine Performances of Vitis Vinifera Cv Cabernet Sauvignon. Sci. Hortic. 2017, 225, 88–95. [Google Scholar] [CrossRef]
- Torres, N.; Yu, R.; Kurtural, S.K. Inoculation with Mycorrhizal Fungi and Irrigation Management Shape the Bacterial and Fungal Communities and Networks in Vineyard Soils. Microorganisms 2021, 9, 1273. [Google Scholar] [CrossRef] [PubMed]
- Betancur-Agudelo, M.; Meyer, E.; Lovato, P.E. Growth, Heavy Metal Uptake, and Photosynthesis in “Paulsen 1103” (Vitis Berlandieri x Rupestris) Grapevine Rootstocks Inoculated with Arbuscular Mycorrhizal Fungi from Vineyard Soils with High Copper Contents. VITIS—J. Grapevine Res. 2020, 59, 169–180. [Google Scholar] [CrossRef]
- Zhou, J.; Zang, H.; Loeppmann, S.; Gube, M.; Kuzyakov, Y.; Pausch, J. Arbuscular Mycorrhiza Enhances Rhizodeposition and Reduces the Rhizosphere Priming Effect on the Decomposition of Soil Organic Matter. Soil Biol. Biochem. 2020, 140, 107641. [Google Scholar] [CrossRef]
- Guzmán, G.; Cabezas, J.M.; Sánchez-Cuesta, R.; Lora, Á.; Bauer, T.; Strauss, P.; Winter, S.; Zaller, J.G.; Gómez, J.A. A Field Evaluation of the Impact of Temporary Cover Crops on Soil Properties and Vegetation Communities in Southern Spain Vineyards. Agric. Ecosyst. Environ. 2019, 272, 135–145. [Google Scholar] [CrossRef]
- Steenwerth, K.; Belina, K.M. Cover Crops Enhance Soil Organic Matter, Carbon Dynamics and Microbiological Function in a Vineyard Agroecosystem. Appl. Soil Ecol. 2008, 40, 359–369. [Google Scholar] [CrossRef]
- Ruiz-Colmenero, M.; Bienes, R.; Marques, M.J. Soil and Water Conservation Dilemmas Associated with the Use of Green Cover in Steep Vineyards. Soil Tillage Res. 2011, 117, 211–223. [Google Scholar] [CrossRef]
- Fleishman, S.M.; Bock, H.W.; Eissenstat, D.M.; Centinari, M. Undervine Groundcover Substantially Increases Shallow but Not Deep Soil Carbon in a Temperate Vineyard. Agric. Ecosyst. Environ. 2021, 313, 107362. [Google Scholar] [CrossRef]
- Ortega, P.; Sánchez, E.; Gil, E.; Matamoros, V. Use of Cover Crops in Vineyards to Prevent Groundwater Pollution by Copper and Organic Fungicides. Soil Column Studies. Chemosphere 2022, 303, 134975. [Google Scholar] [CrossRef]
- Eon, P.; Robert, T.; Goutouly, J.-P.; Aurelle, V.; Cornu, J.-Y. Cover Crop Response to Increased Concentrations of Copper in Vineyard Soils: Implications for Copper Phytoextraction. Chemosphere 2023, 329, 138604. [Google Scholar] [CrossRef] [PubMed]
- Austin, E.E.; Wickings, K.; McDaniel, M.D.; Robertson, G.P.; Grandy, A.S. Cover Crop Root Contributions to Soil Carbon in a No-till Corn Bioenergy Cropping System. GCB Bioenergy 2017, 9, 1252–1263. [Google Scholar] [CrossRef]
- Mortensen, E.Ø.; De Notaris, C.; Peixoto, L.; Olesen, J.E.; Rasmussen, J. Short-Term Cover Crop Carbon Inputs to Soil as Affected by Long-Term Cropping System Management and Soil Fertility. Agric. Ecosyst. Environ. 2021, 311, 107339. [Google Scholar] [CrossRef]
- Novara, A.; Cerda, A.; Barone, E.; Gristina, L. Cover Crop Management and Water Conservation in Vineyard and Olive Orchards. Soil Tillage Res. 2021, 208, 104896. [Google Scholar] [CrossRef]
- Buesa, I.; Mirás-Avalos, J.M.; De Paz, J.M.; Visconti, F.; Sanz, F.; Yeves, A.; Guerra, D.; Intrigliolo, D.S. Soil Management in Semi-Arid Vineyards: Combined Effects of Organic Mulching and No-Tillage under Different Water Regimes. Eur. J. Agron. 2021, 123, 126198. [Google Scholar] [CrossRef]
- Beslic, Z.; Pantelic, M.; Dabic, D.; Todic, S.; Natic, M.; Tesic, Z. Effect of Vineyard Floor Management on Water Regime, Growth Response, Yield and Fruit Quality in Cabernet Sauvignon. Sci. Hortic. 2015, 197, 650–656. [Google Scholar] [CrossRef]
- Reynolds, W.D.; Bowman, B.T.; Drury, C.F.; Tan, C.S.; Lu, X. Indicators of Good Soil Physical Quality: Density and Storage Parameters. Geoderma 2002, 110, 131–146. [Google Scholar] [CrossRef]
- Kumar, K.S.A.; Karthika, K.S. Abiotic and Biotic Factors Influencing Soil Health and/or Soil Degradation. In Soil Health; Springer: Berlin/Heidelberg, Germany, 2020; pp. 145–161. [Google Scholar]
- Manda, R.R.; Addanki, V.A.; Giabardo, A.; Benjamin, J.; Hossain, M.J.; Khanna, S.; Gaddam, M.; Kumar, R.; Srivastava, S. Soil Health Management and Microorganisms: Recent Development. In Detection, Diagnosis and Management of Soil-Borne Phytopathogens; Springer Nature Singapore: Singapore, 2023; pp. 437–493. [Google Scholar]
- Oliver, D.P.; Bramley, R.G.V.; Riches, D.; Porter, I.; Edwards, J. Review: Soil Physical and Chemical Properties as Indicators of Soil Quality in Australian Viticulture. Aust. J. Grape Wine Res. 2013, 19, 129–139. [Google Scholar] [CrossRef]
- Saljnikov, E.; Mueller, L.; Lavrishchev, A.; Eulenstein, F. Advances in Understanding Soil Degradation Innovations in Landscape Research; Springer: Müncheberg, Germany, 2022. [Google Scholar]
- Schjønning, P. Subsoil Compaction—A Threat to Sustainable Food Production and Soil Ecosystem Services; RECARE Policy Brief; RECARE: Berlin, Germany, 2018. [Google Scholar]
- Batey, T. Soil Compaction and Soil Management—A Review. Soil Use Manag. 2009, 25, 335–345. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines, 3rd ed.; Maragioglio, N., Ed.; Elsevier: Nueva Delhi, India, 2020. [Google Scholar]
- Kirschbaum, M.U.F. The Temperature Dependence of Soil Organic Matter Decomposition, and the Effect of Global Warming on Soil Organic C Storage. Soil Biol. Biochem. 1995, 27, 753–760. [Google Scholar] [CrossRef]
- Keller, T.; Sandin, M.; Colombi, T.; Horn, R.; Or, D. Historical Increase in Agricultural Machinery Weights Enhanced Soil Stress Levels and Adversely Affected Soil Functioning. Soil Tillage Res. 2019, 194, 104293. [Google Scholar] [CrossRef]
- Horn, R.; Schrader, S.; Mordhorst, A.; Fleige, H.; Schroeder, R. Soil Health and Biodiversity: Interactions with Physical Processes and Functions. In Sustainable Soil Management as a Key to Preserve Soil Biodiversity and Stop Its Degradation; Reyes-Sánchez, L., Horn, R., Costantini, E., Eds.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022; pp. 314–330. [Google Scholar]
- Bogunovic, I.; Bilandzija, D.; Andabaka, Z.; Stupic, D.; Rodrigo Comino, J.; Cacic, M.; Brezinscak, L.; Maletic, E.; Pereira, P. Soil Compaction under Different Management Practices in a Croatian Vineyard. Arab. J. Geosci. 2017, 10, 340. [Google Scholar] [CrossRef]
- Inglett, P.; Reddy, K.; Corstanje, R. Anaerobic Soils. In Encyclopedia of Soils in the Environment; Elsevier: Amsterdam, The Netherlands, 1998; pp. 72–78. [Google Scholar]
- Tan, X.; Chang, S.X. Soil Compaction and Forest Litter Amendment Affect Carbon and Net Nitrogen Mineralization in a Boreal Forest Soil. Soil Tillage Res. 2007, 93, 77–86. [Google Scholar] [CrossRef]
- Tropeano, D. Rate of Soil Erosion Processes on Vineyards in Central Piedmont (NW Italy). Earth Surf. Process Landf. 1984, 9, 253–266. [Google Scholar] [CrossRef]
- Vršič, S. Soil Erosion and Earthworm Population Responses to Soil Management Systems in Steep-Slope Vineyards. Plant Soil Environ. 2011, 57, 258–263. [Google Scholar] [CrossRef]
- Prosdocimi, M.; Jordán, A.; Tarolli, P.; Keesstra, S.; Novara, A.; Cerdà, A. The Immediate Effectiveness of Barley Straw Mulch in Reducing Soil Erodibility and Surface Runoff Generation in Mediterranean Vineyards. Sci. Total Environ. 2016, 547, 323–330. [Google Scholar] [CrossRef]
- Cerdà, A.; Keesstra, S.D.; Rodrigo-Comino, J.; Novara, A.; Pereira, P.; Brevik, E.; Giménez-Morera, A.; Fernández-Raga, M.; Pulido, M.; di Prima, S.; et al. Runoff Initiation, Soil Detachment and Connectivity Are Enhanced as a Consequence of Vineyards Plantations. J. Environ. Manag. 2017, 202, 268–275. [Google Scholar] [CrossRef]
- Pappalardo, S.E.; Gislimberti, L.; Ferrarese, F.; De Marchi, M.; Mozzi, P. Estimation of Potential Soil Erosion in the Prosecco DOCG Area (NE Italy), toward a Soil Footprint of Bottled Sparkling Wine Production in Different Land-Management Scenarios. PLoS ONE 2019, 14, e0210922. [Google Scholar] [CrossRef]
- Stanchi, S.; Zecca, O.; Hudek, C.; Pintaldi, E.; Viglietti, D.; D’Amico, M.E.; Colombo, N.; Goslino, D.; Letey, M.; Freppaz, M. Effect of Soil Management on Erosion in Mountain Vineyards (N-W Italy). Sustainability 2021, 13, 1991. [Google Scholar] [CrossRef]
- Alewell, C.; Ringeval, B.; Ballabio, C.; Robinson, D.A.; Panagos, P.; Borrelli, P. Global Phosphorus Shortage Will Be Aggravated by Soil Erosion. Nat. Commun. 2020, 11, 4546. [Google Scholar] [CrossRef]
- Novara, A.; Pisciotta, A.; Minacapilli, M.; Maltese, A.; Capodici, F.; Cerdà, A.; Gristina, L. The Impact of Soil Erosion on Soil Fertility and Vine Vigor. A Multidisciplinary Approach Based on Field, Laboratory and Remote Sensing Approaches. Sci. Total Environ. 2018, 622–623, 474–480. [Google Scholar] [CrossRef]
- Costantini, E.A.C.; Castaldini, M.; Diago, M.P.; Giffard, B.; Lagomarsino, A.; Schroers, H.-J.; Priori, S.; Valboa, G.; Agnelli, A.E.; Akça, E.; et al. Effects of Soil Erosion on Agro-Ecosystem Services and Soil Functions: A Multidisciplinary Study in Nineteen Organically Farmed European and Turkish Vineyards. J. Environ. Manag. 2018, 223, 614–624. [Google Scholar] [CrossRef]
- Pijl, A.; Wang, W.; Straffelini, E.; Tarolli, P. Soil and Water Conservation in Terraced and Non-Terraced Cultivations: An Extensive Comparison of 50 Vineyards. Land Degrad. Dev. 2022, 33, 596–610. [Google Scholar] [CrossRef]
- Biddoccu, M.; Ferraris, S.; Opsi, F.; Cavallo, E. Long-Term Monitoring of Soil Management Effects on Runoff and Soil Erosion in Sloping Vineyards in Alto Monferrato (North–West Italy). Soil Tillage Res. 2016, 155, 176–189. [Google Scholar] [CrossRef]
- Dezső, J.; Lóczy, D.; Rezsek, M.; Hüppi, R.; Werner, J.; Horváth, L. Crop Growth, Carbon Sequestration and Soil Erosion in an Organic Vineyard of the Villány Wine District, Southwest Hungary. Hung. Geogr. Bull. 2020, 69, 281–298. [Google Scholar] [CrossRef]
- Ruiz-Colmenero, M.; Bienes, R.; Eldridge, D.J.; Marques, M.J. Vegetation Cover Reduces Erosion and Enhances Soil Organic Carbon in a Vineyard in the Central Spain. Catena 2013, 104, 153–160. [Google Scholar] [CrossRef]
- Winter, S.; Bauer, T.; Strauss, P.; Kratschmer, S.; Paredes, D.; Popescu, D.; Landa, B.; Guzmán, G.; Gómez, J.A.; Guernion, M.; et al. Effects of Vegetation Management Intensity on Biodiversity and Ecosystem Services in Vineyards: A Meta-Analysis. J. Appl. Ecol. 2018, 55, 2484–2495. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.H.; Wang, L.; Wan, X.G.; Peng, Q.Z.; Huang, Q.; Shi, Z.H. A Systematic Review of Soil Erosion in Citrus Orchards Worldwide. Catena 2021, 206, 105558. [Google Scholar] [CrossRef]
- Pou, A.; Gulías, J.; Moreno, M.; Tomàs, M.; Medrano, H.; Cifre, J. Cover Cropping in Vitis vinifera L. Cv. Manto Negro Vineyards under Mediterranean Conditions: Effects on Plant Vigour, Yield and Grape Quality. OENO One 2011, 45, 223. [Google Scholar] [CrossRef]
- Mirás-Avalos, J.M.; Ramírez-Cuesta, J.M.; Fandiño, M.; Cancela, J.J.; Intrigliolo, D.S. Agronomic Practices for Reducing Soil Erosion in Hillside Vineyards under Atlantic Climatic Conditions (Galicia, Spain). Soil Syst. 2020, 4, 19. [Google Scholar] [CrossRef]
- Triplett, G.B.; Dick, W.A. No-Tillage Crop Production: A Revolution in Agriculture! Agron. J. 2008, 100, S-153. [Google Scholar] [CrossRef]
- Osterholz, W.R.; Culman, S.W.; Herms, C.; Joaquim de Oliveira, F.; Robinson, A.; Doohan, D. Knowledge Gaps in Organic Research: Understanding Interactions of Cover Crops and Tillage for Weed Control and Soil Health. Org. Agric. 2021, 11, 13–25. [Google Scholar] [CrossRef]
- Laudicina, V.A.; Palazzolo, E.; Catania, P.; Vallone, M.; García, A.D.; Badalucco, L. Soil Quality Indicators as Affected by Shallow Tillage in a Vineyard Grown in a Semiarid Mediterranean Environment. Land Degrad. Dev. 2017, 28, 1038–1046. [Google Scholar] [CrossRef]
- Shaheb, M.R.; Venkatesh, R.; Shearer, S.A. A Review on the Effect of Soil Compaction and Its Management for Sustainable Crop Production. J. Biosyst. Eng. 2021, 46, 417–439. [Google Scholar] [CrossRef]
- Schwartz, R.C.; Baumhardt, R.L.; Evett, S.R. Tillage Effects on Soil Water Redistribution and Bare Soil Evaporation throughout a Season. Soil Tillage Res. 2010, 110, 221–229. [Google Scholar] [CrossRef]
- Nunes, M.R.; Karlen, D.L.; Moorman, T.B.; Cambardella, C.A. How Does Tillage Intensity Affect Chemical Soil Health Indicators? A United States Meta-Analysis. Agrosyst. Geosci. Environ. 2020, 3, e20083. [Google Scholar] [CrossRef]
- Linares, R.; de la Fuente, M.; Junquera, P.; Lissarrague, J.R.; Baeza, P. Effects of Soil Management in Vineyard on Soil Physical and Chemical Characteristics. BIO Web Conf. 2014, 3, 01008. [Google Scholar] [CrossRef]
- Nunes, M.R.; Karlen, D.L.; Veum, K.S.; Moorman, T.B.; Cambardella, C.A. Biological Soil Health Indicators Respond to Tillage Intensity: A US Meta-Analysis. Geoderma 2020, 369, 114335. [Google Scholar] [CrossRef]
- Badalíková, B. Influence of Soil Tillage on Soil Compaction. In Soil Biology; Dedousis, A., Bartzanas, T., Eds.; Springer: Volos, Greece, 2010; pp. 19–30. [Google Scholar]
- Saffih-Hdadi, K.; Défossez, P.; Richard, G.; Cui, Y.-J.; Tang, A.-M.; Chaplain, V. A Method for Predicting Soil Susceptibility to the Compaction of Surface Layers as a Function of Water Content and Bulk Density. Soil Tillage Res. 2009, 105, 96–103. [Google Scholar] [CrossRef]
- Ahmadi, I.; Ghaur, H. Effects of Soil Moisture Content and Tractor Wheeling Intensity on Traffic-Induced Soil Compaction. J. Cent. Eur. Agric. 2015, 16, 489–502. [Google Scholar] [CrossRef]
- Schjønning, P.; Lamandé, M.; Tøgersen, F.A.; Arvidsson, J.; Keller, T. Modelling Effects of Tyre Inflation Pressure on the Stress Distribution near the Soil–Tyre Interface. Biosyst. Eng. 2008, 99, 119–133. [Google Scholar] [CrossRef]
- Hamza, M.A.; Anderson, W.K. Soil Compaction in Cropping Systems: A Review of the Nature, Causes and Possible Solutions. Soil Tillage Res. 2005, 82, 121–145. [Google Scholar] [CrossRef]
- Van Dijck, S.J.E.; van Asch, T.W.J. Compaction of Loamy Soils Due to Tractor Traffic in Vineyards and Orchards and Its Effect on Infiltration in Southern France. Soil Tillage Res. 2002, 63, 141–153. [Google Scholar] [CrossRef]
- Pessina, D.; Galli, L.E.; Santoro, S.; Facchinetti, D. Sustainability of Machinery Traffic in Vineyard. Sustainability 2021, 13, 24745. [Google Scholar] [CrossRef]
- Elaoud, A.; Chehaibi, S. Soil Compaction Due to Tractor Traffic. J. Fail. Anal. Prev. 2011, 11, 539–545. [Google Scholar] [CrossRef]
- Lagacherie, P.; Coulouma, G.; Ariagno, P.; Virat, P.; Boizard, H.; Richard, G. Spatial Variability of Soil Compaction over a Vineyard Region in Relation with Soils and Cultivation Operations. Geoderma 2006, 134, 207–216. [Google Scholar] [CrossRef]
- Hunter, J.J.; Archer, E.; Van Schalkwyk, D.; Strever, A.E.; Volschenk, C.G. Grapevine Roots: Interaction with Natural Factors and Agronomic Practices. In Acta Horticulturae, Proceedings of the I International Symposium on Grapevine Roots, Chania, Greece, 30 July 2016; International Society for Horticultural Science: Leuven, Belgium, 2016; Volume 1136, pp. 63–80. [Google Scholar]
- Marinello, F.; Pezzuolo, A.; Cillis, D.; Chiumenti, A.; Sartori, L. Traffic Effects on Soil Compaction and Sugar Beet (Beta Vulgaris L.) Taproot Quality Parameters. Span. J. Agric. Res. 2017, 15, e0201. [Google Scholar] [CrossRef]
- Döring, J.; Friedel, M.; Hendgen, M.; Stoll, M.; Kauer, R. Soil Management in Sustainable Viticultural Systems. In Improving Sustainable Viticulture and Winemaking Practices; Elsevier: Amsterdam, The Netherlands, 2022; pp. 85–103. [Google Scholar]
- Burg, P.; Masan, V.; Zemanek, P. Evaluation of Effect of Vegetation Cover on Soil Compaction. In Proceedings of the Engineering for Rural Development, Jelgava, Latvia, 23–25 May 2017; Latvia University of Agriculture: Jelgava, Latvia, 2017; Volume 16, pp. 1346–1353. [Google Scholar]
- Visconti, F.; Jiménez, M.G.; de Paz, J.M. How Do the Chemical Characteristics of Organic Matter Explain Differences among Its Determinations in Calcareous Soils? Geoderma 2022, 406, 115454. [Google Scholar] [CrossRef]
- Hartwig, N.L.; Ammon, H.U. Cover Crops and Living Mulches. Weed Sci. 2002, 50, 688–699. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Litskas, V.; Stavrinides, M.; Heyman, L.; Demeestere, K.; Höfte, M.; Tzortzakis, N. Assessing the Impact of Drought Stress and Soil Cultivation in Chardonnay and Xynisteri Grape Cultivars. Agronomy 2020, 10, 670. [Google Scholar] [CrossRef]
- Capri, C.; Gatti, M.; Fiorini, A.; Ardenti, F.; Tabaglio, V.; Poni, S. A Comparative Study of Fifteen Cover Crop Species for Orchard Soil Management: Water Uptake, Root Density Traits and Soil Aggregate Stability. Sci. Rep. 2023, 13, 721. [Google Scholar] [CrossRef]
- Zengin, H.; Sabir, A. Physiological and Growth Responses of Grapevine Rootstocks (Vitis Spp.) to Organic and Synthetic Mulch Application in Arid Ecology under the Effect of Climate Change. J. Cent. Eur. Agric. 2022, 23, 655–664. [Google Scholar] [CrossRef]
- Cabrera-Pérez, C.; Llorens, J.; Escolà, A.; Royo-Esnal, A.; Recasens, J. Organic Mulches as an Alternative for Under-Vine Weed Management in Mediterranean Irrigated Vineyards: Impact on Agronomic Performance. Eur. J. Agron. 2023, 145, 126798. [Google Scholar] [CrossRef]
- Marks, J.N.J.; Lines, T.E.P.; Penfold, C.; Cavagnaro, T.R. Cover Crops and Carbon Stocks: How under-Vine Management Influences SOC Inputs and Turnover in Two Vineyards. Sci. Total Environ. 2022, 831, 154800. [Google Scholar] [CrossRef] [PubMed]
- Gattullo, C.E.; Mezzapesa, G.N.; Stellacci, A.M.; Ferrara, G.; Occhiogrosso, G.; Petrelli, G.; Castellini, M.; Spagnuolo, M. Cover Crop for a Sustainable Viticulture: Effects on Soil Properties and Table Grape Production. Agronomy 2020, 10, 1334. [Google Scholar] [CrossRef]
- Salomé, C.; Coll, P.; Lardo, E.; Metay, A.; Villenave, C.; Marsden, C.; Blanchart, E.; Hinsinger, P.; Le Cadre, E. The Soil Quality Concept as a Framework to Assess Management Practices in Vulnerable Agroecosystems: A Case Study in Mediterranean Vineyards. Ecol. Indic. 2016, 61, 456–465. [Google Scholar] [CrossRef]
- Ferrara, G.; Nigro, D.; Torres, R.; Gadaleta, A.; Fidelibus, M.W.; Mazzeo, A. Cover Crops in the Inter-Row of a Table Grape Vineyard Managed with Irrigation Sensors: Effects on Yield, Quality and Glutamine Synthetase Activity in Leaves. Sci. Hortic. 2021, 281, 109963. [Google Scholar] [CrossRef]
- DeVetter, L.W.; Dilley, C.A.; Nonnecke, G.R. Mulches Reduce Weeds, Maintain Yield, and Promote Soil Quality in a Continental-Climate Vineyard. Am. J. Enol. Vitic. 2015, 66, 54–64. [Google Scholar] [CrossRef]
- Mania, E.; Isocrono, D.; Pedullà, M.L.; Guidoni, S. Plant Diversity in an Intensively Cultivated Vineyard Agro-Ecosystem (Langhe, North-West Italy). S. Afr. J. Enol. Vitic. 2015, 36, 378–388. [Google Scholar] [CrossRef]
- Eisenhauer, N. Plant Diversity Effects on Soil Microorganisms: Spatial and Temporal Heterogeneity of Plant Inputs Increase Soil Biodiversity. Pedobiologia 2016, 59, 175–177. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.Y.H.; Chen, X.; Huang, Z. Meta-Analysis Shows Positive Effects of Plant Diversity on Microbial Biomass and Respiration. Nat. Commun. 2019, 10, 1332. [Google Scholar] [CrossRef] [PubMed]
- Niles, M.T.; Garrett, R.D.; Walsh, D. Ecological and Economic Benefits of Integrating Sheep into Viticulture Production. Agron. Sustain. Dev. 2018, 38, 1. [Google Scholar] [CrossRef]
- Fermaud, M.; Smits, N.; Merot, A.; Roudet, J.; Thiéry, D.; Wery, J.; Delbac, L. New Multipest Damage Indicator to Assess Protection Strategies in Grapevine Cropping Systems. Aust. J. Grape Wine Res. 2016, 22, 450–461. [Google Scholar] [CrossRef]
- Daane, K.M.; Cooper, M.L.; Triapitsyn, S.V.; Walton, V.M.; Yokota, G.Y.; Haviland, D.R.; Bentley, W.J.; Godfrey, K.E.; Wunderlich, L.R. Vineyard Managers and Researchers Seek Sustainable Solutions for Mealybugs, a Changing Pest Complex. Calif. Agric. 2008, 62, 167–176. [Google Scholar] [CrossRef]
- Reineke, A.; Thiéry, D. Grapevine Insect Pests and Their Natural Enemies in the Age of Global Warming. J. Pest Sci. 2016, 89, 313–328. [Google Scholar] [CrossRef]
- Imperatore, G.; Ghirardelli, A.; Strinna, L.; Baldoin, C.; Pozzebon, A.; Zanin, G.; Otto, S. Evaluation of a Fixed Spraying System for Phytosanitary Treatments in Heroic Viticulture in North-Eastern Italy. Agriculture 2021, 11, 833. [Google Scholar] [CrossRef]
- European Commission Commission Regulation (EC). No 473/2002 of 15 March 2002 Amending Annexes I, II and VI to Council Regulation (EEC) No 2092/91 on Organic Production of Agricultural Products and Indications Referring Thereto on Agricultural Products and Foodstuffs, and Laying Down Detailed Rules as Regards the Transmission of Information on the Use of Copper Compounds. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32002R0473 (accessed on 6 December 2023).
- Unsworth, J. History of Pesticide Use; International Union of Pure and Applied Chemistry: Durham, NC, USA, 2010. [Google Scholar]
- Vocke, G. The Green Revolution for Wheat in Developing Countries; US Department of Agriculture: New York, NY, USA, 1986.
- Kogan, M. Integrated Pest Management: Historical Perspectives and Contemporary Developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight Principles of Integrated Pest Management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Gregoire, C.; Payraudeau, S.; Domange, N. Use and Fate of 17 Pesticides Applied on a Vineyard Catchment. Int. J. Environ. Anal. Chem. 2010, 90, 406–420. [Google Scholar] [CrossRef]
- Cabras, P.; Meloni, M.; Pirisi, F.M. Pesticide Fate from Vine to Wine. Rev. Environ. Contam. Toxicol. 1987, 99, 83–117. [Google Scholar] [CrossRef] [PubMed]
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; Hoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A Critical Review of Plant Protection Tools for Reducing Pesticide Use on Grapevine and New Perspectives for the Implementation of IPM in Viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar] [CrossRef]
- Kullaj, E.; Shahini, S.; Varaku, S.; Çakalli, M. Evaluation of the Efficacy for Reducing Copper Use against Downy Mildew Control in Organic Mediterranean Viticulture. Int. J. Pest. Manag. 2016, 63, 3–9. [Google Scholar] [CrossRef]
- Kraus, C.; Abou-Ammar, R.; Schubert, A.; Fischer, M. Warburgia Ugandensis Leaf and Bark Extracts: An Alternative to Copper as Fungicide against Downy Mildew in Organic Viticulture? Plants 2021, 10, 2765. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Bernal, R.; Rodríguez-Miranda, E.; Herrera-Pérez, G.; Vargas-Bernal, R.; Rodríguez-Miranda, E.; Herrera-Pérez, G. Evolution and Expectations of Enzymatic Biosensors for Pesticides. In Pesticides-Advances in Chemical and Botanical Pesticides; IntechOpen: London, UK, 2012; ISBN 978-953-51-0680-7. [Google Scholar]
- Urruty, N.; Deveaud, T.; Guyomard, H.; Boiffin, J. Impacts of Agricultural Land Use Changes on Pesticide Use in French Agriculture. Eur. J. Agron. 2016, 80, 113–123. [Google Scholar] [CrossRef]
- Droz, B.; Payraudeau, S.; Rodríguez Martín, J.A.; Tóth, G.; Panagos, P.; Montanarella, L.; Borrelli, P.; Imfeld, G. Copper Content and Export in European Vineyard Soils Influenced by Climate and Soil Properties. Environ. Sci. Technol. 2021, 55, 7327–7334. [Google Scholar] [CrossRef] [PubMed]
- Dagostin, S.; Schärer, H.-J.; Pertot, I.; Tamm, L. Are There Alternatives to Copper for Controlling Grapevine Downy Mildew in Organic Viticulture? Crop Prot. 2011, 30, 776–788. [Google Scholar] [CrossRef]
- Lo Scalzo, R.; Fibiani, M.; Pietromarchi, P.; Mandalà, C.; La Torre, A. Effects of Different Fungicide Treatments on Grape, Must and Wine Quality. Commun. Agric. Appl. Biol. Sci. 2012, 77, 151–161. [Google Scholar]
- Thuerig, B.; James, E.E.; Schärer, H.; Langat, M.K.; Mulholland, D.A.; Treutwein, J.; Kleeberg, I.; Ludwig, M.; Jayarajah, P.; Giovannini, O.; et al. Reducing Copper Use in the Environment: The Use of Larixol and Larixyl Acetate to Treat Downy Mildew Caused by Plasmopara Viticola in Viticulture. Pest Manag. Sci. 2017, 74, 477–488. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.-N. Thirteen Decades of Antimicrobial Copper Compounds Applied in Agriculture. A Review. Agron. Sustain. Dev. 2018, 38, 28. [Google Scholar] [CrossRef]
- Widmer, J.; Norgrove, L. Identifying Candidates for the Phytoremediation of Copper in Viticultural Soils: A Systematic Review. Environ. Res. 2023, 216, 114518. [Google Scholar] [CrossRef]
- Ballabio, C.; Panagos, P.; Lugato, E.; Huang, J.-H.; Orgiazzi, A.; Jones, A.; Fernández-Ugalde, O.; Borrelli, P.; Montanarella, L. Copper Distribution in European Topsoils: An Assessment Based on LUCAS Soil Survey. Sci. Total Environ. 2018, 636, 282–298. [Google Scholar] [CrossRef] [PubMed]
- New South Wales Environment Protection Agency. Environmental Management Guidelines—Use and Disposal of Biosolids Products; New South Wales Environment Protection Agency: Parramatta, Australia, 1997.
- Voluntary Risk Assessment Reports-Copper and Copper Compounds-ECHA. Available online: https://echa.europa.eu/es/copper-voluntary-risk-assessment-reports (accessed on 6 December 2023).
- Sandmann, G.; Böger, P. The Enzymatological Function of Heavy Metals and Their Role in Electron Transfer Processes of Plants. In Encyclopedia of Plant Physiology; Lauchli, A., Bieleski, R.L., Eds.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 563–596. [Google Scholar]
- Solomon, E.I.; Heppner, D.E.; Johnston, E.M.; Ginsbach, J.W.; Cirera, J.; Qayyum, M.; Kieber-Emmons, M.T.; Kjaergaard, C.H.; Hadt, R.G.; Tian, L. Copper Active Sites in Biology. Chem. Rev. 2014, 114, 3659–3853. [Google Scholar] [CrossRef]
- Yruela, I. Copper in Plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients: Micronutrients. In Marschner’s Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 191–248. [Google Scholar] [CrossRef]
- Gaetke, L. Copper Toxicity, Oxidative Stress, and Antioxidant Nutrients. Toxicology 2003, 189, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Borkow, G.; Gabbay, J. Copper, An Ancient Remedy Returning to Fight Microbial, Fungal and Viral Infections. Curr. Chem. Biol. 2009, 3, 272–278. [Google Scholar] [CrossRef]
- Wairich, A.; De Conti, L.; Lamb, T.I.; Keil, R.; Neves, L.O.; Brunetto, G.; Sperotto, R.A.; Ricachenevsky, F.K. Throwing Copper Around: How Plants Control Uptake, Distribution, and Accumulation of Copper. Agronomy 2022, 12, 994. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Henriques, F.S. Biochemical, Physiological, and Structural Effects of Excess Copper in Plants. Bot. Rev. 1991, 57, 246–273. [Google Scholar] [CrossRef]
- Babu, T.S.; Akhtar, T.A.; Lampi, M.A.; Tripuranthakam, S.; Dixon, D.G.; Greenberg, B.M. Similar Stress Responses Are Elicited by Copper and Ultraviolet Radiation in the Aquatic Plant Lemna Gibba: Implication of Reactive Oxygen Species as Common Signals. Plant Cell Physiol. 2003, 44, 1320–1329. [Google Scholar] [CrossRef]
- Sheldon, A.R.; Menzies, N.W. The Effect of Copper Toxicity on the Growth and Root Morphology of Rhodes Grass (Chloris gayana Knuth.) in Resin Buffered Solution Culture. Plant Soil 2005, 278, 341–349. [Google Scholar] [CrossRef]
- Lequeux, H.; Hermans, C.; Lutts, S.; Verbruggen, N. Response to Copper Excess in Arabidopsis thaliana: Impact on the Root System Architecture, Hormone Distribution, Lignin Accumulation and Mineral Profile. Plant Physiol. Biochem. 2010, 48, 673–682. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, P.; Soto-Gómez, D.; De La Calle, I.; López-Periago, J.E.; Paradelo, M. Rainfall-Induced Removal of Copper-Based Spray Residues from Vines. Ecotoxicol. Environ. Saf. 2016, 132, 304–310. [Google Scholar] [CrossRef]
- Hummes, A.P.; Bortoluzzi, E.C.; Tonini, V.; da Silva, L.P.; Petry, C. Transfer of Copper and Zinc from Soil to Grapevine-Derived Products in Young and Centenarian Vineyards. Water Air Soil Pollut. 2019, 230, 150. [Google Scholar] [CrossRef]
- Zhao, F.; Rooney, C.P.; Zhang, H.; McGrath, S.P. Comparison of Soil Solution Speciation and Diffusive Gradients in Thin-films Measurement as an Indicator of Copper Bioavailability to Plants. Environ. Toxicol. Chem. 2006, 25, 733–742. [Google Scholar] [CrossRef]
- Li, L.; Wu, H.; van Gestel, C.A.M.; Peijnenburg, W.J.G.M.; Allen, H.E. Soil Acidification Increases Metal Extractability and Bioavailability in Old Orchard Soils of Northeast Jiaodong Peninsula in China. Environ. Pollut. 2014, 188, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Zuo, Y.B.; Wang, B.R.; Li, J.M.; Ma, Y.B. Toxicity and Accumulation of Copper and Nickel in Maize Plants Cropped on Calcareous and Acidic Field Soils. Plant Soil 2010, 333, 365–373. [Google Scholar] [CrossRef]
- Huang, J.-S.; Wei, D.-P.; Guo, X.-Y.; Ma, Y.-B. Toxicity and Accumulation of Copper and Nickel in Wheat Plants Cropped on Alkaline and Acidic Field Soils. Huan Jing Ke Xue/Environ. Sci. 2012, 33, 1369–1375. [Google Scholar]
- Trentin, E.; Cesco, S.; Pii, Y.; Valentinuzzi, F.; Celletti, S.; Feil, S.B.; Zuluaga, M.Y.A.; Ferreira, P.A.A.; Ricachenevsky, F.K.; Stefanello, L.O.; et al. Plant Species and PH Dependent Responses to Copper Toxicity. Environ. Exp. Bot. 2022, 196, 104791. [Google Scholar] [CrossRef]
- Miotto, A.; Ceretta, C.A.; Brunetto, G.; Nicoloso, F.T.; Girotto, E.; Farias, J.G.; Tiecher, T.L.; De Conti, L.; Trentin, G. Copper Uptake, Accumulation and Physiological Changes in Adult Grapevines in Response to Excess Copper in Soil. Plant Soil 2013, 374, 593–610. [Google Scholar] [CrossRef]
- Ambrosini, V.G.; Rosa, D.J.; Bastos de Melo, G.W.; Zalamena, J.; Cella, C.; Simão, D.G.; Souza da Silva, L.; Pessoa dos Santos, H.; Toselli, M.; Tiecher, T.L.; et al. High Copper Content in Vineyard Soils Promotes Modifications in Photosynthetic Parameters and Morphological Changes in the Root System of ‘Red Niagara’ Plantlets. Plant Physiol. Biochem. 2018, 128, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Feil, S.B.; Zuluaga, M.Y.A.; Cesco, S.; Pii, Y. Copper Toxicity Compromises Root Acquisition of Nitrate in the High Affinity Range. Front. Plant Sci. 2023, 13, 1034425. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.J.; Ambrosini, V.G.; Kokkoris, V.; Brunetto, G.; Hart, M.; Ricachenevsky, F.; Pescador, R. Lime Protection for Young Vines Exposed to Copper Toxicity. Water Air Soil Pollut. 2020, 231, 296. [Google Scholar] [CrossRef]
- Ma, Y.; Lombi, E.; Oliver, I.W.; Nolan, A.L.; McLaughlin, M.J. Long-Term Aging of Copper Added to Soils. Environ. Sci. Technol. 2006, 40, 6310–6317. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hua, L.; Ma, Y. A Biotic Ligand Model Predicting Acute Copper Toxicity for Barley (Hordeum Vulgare): Influence of Calcium, Magnesium, Sodium, Potassium and PH. Chemosphere 2012, 89, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, F.J.; Fitch, A. Reactions with Organic Matter. In Copper in Soils and Plants; Loneragan, J.F., Robson, A.D., Graham, R.D., Eds.; Academic Press: Sydney, Australia, 1981; pp. 265–285. [Google Scholar]
- Fernández-Calviño, D.; Nóvoa-Muñoz, J.C.; López-Periago, E.; Arias-Estévez, M. Changes in Copper Content and Distribution in Young, Old and Abandoned Vineyard Acid Soils Due to Land Use Changes. Land. Degrad. Dev. 2007, 19, 165–177. [Google Scholar] [CrossRef]
- Lagomarsino, A.; Marabottini, S.; Grego, S.; Stazi, S.R. Copper Distribution among Physical and Chemical Fractions in a Former Vineyard Soil. Agrochimica 2010, LIV, 1–12. [Google Scholar]
- Eijsackers, H.; Beneke, P.; Maboeta, M.; Louw, J.P.E.; Reinecke, A.J. The Implications of Copper Fungicide Usage in Vineyards for Earthworm Activity and Resulting Sustainable Soil Quality. Ecotoxicol. Environ. Saf. 2005, 62, 99–111. [Google Scholar] [CrossRef]
- Duan, X.; Xu, M.; Zhou, Y.; Yan, Z.; Du, Y.; Zhang, L.; Zhang, C.; Bai, L.; Nie, J.; Chen, G.; et al. Effects of Soil Properties on Copper Toxicity to Earthworm Eisenia Fetida in 15 Chinese Soils. Chemosphere 2016, 145, 185–192. [Google Scholar] [CrossRef]
- Qiu, H.; Vijver, M.G.; He, E.; Peijnenburg, W.J.G.M. Predicting Copper Toxicity to Different Earthworm Species Using a Multicomponent Freundlich Model. Environ. Sci. Technol. 2013, 47, 4796–4803. [Google Scholar] [CrossRef] [PubMed]
- Tiecher, T.L.; Tiecher, T.; Ceretta, C.A.; Ferreira, P.A.A.; Nicoloso, F.T.; Soriani, H.H.; De Conti, L.; Kulmann, M.S.S.; Schneider, R.O.; Brunetto, G. Tolerance and Translocation of Heavy Metals in Young Grapevine (Vitis Vinifera) Grown in Sandy Acidic Soil with Interaction of High Doses of Copper and Zinc. Sci. Hortic. 2017, 222, 203–212. [Google Scholar] [CrossRef]
- Shaw, J.L.A.; Ernakovich, J.G.; Judy, J.D.; Farrell, M.; Whatmuff, M.; Kirby, J. Long-Term Effects of Copper Exposure to Agricultural Soil Function and Microbial Community Structure at a Controlled and Experimental Field Site. Environ. Pollut. 2020, 263, 114411. [Google Scholar] [CrossRef] [PubMed]
- Ben Massoud, M.; Kharbech, O.; Sakouhi, L.; Ben Hassine, S.; Zhu, Y.; Chaoui, A.; Sheehan, D.; Djebali, W. Calcium and Citrate Protect Pisum Sativum Roots against Copper Toxicity by Regulating the Cellular Redox Status. J. Soil Sci. Plant Nutr. 2021, 22, 345–358. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Neonicotinoid Insecticides: Highlights of a Symposium on Strategic Molecular Designs. J. Agric. Food Chem. 2010, 59, 2883–2886. [Google Scholar] [CrossRef]
- Zaller, J.G.; Cantelmo, C.; Dos Santos, G.; Muther, S.; Gruber, E.; Pallua, P.; Mandl, K.; Friedrich, B.; Hofstetter, I.; Schmuckenschlager, B.; et al. Herbicides in Vineyards Reduce Grapevine Root Mycorrhization and Alter Soil Microorganisms and the Nutrient Composition in Grapevine Roots, Leaves, Xylem Sap and Grape Juice. Environ. Sci. Pollut. Res. Int. 2018, 25, 23215–23226. [Google Scholar] [CrossRef] [PubMed]
- Stellin, F.; Gavinelli, F.; Stevanato, P.; Concheri, G.; Squartini, A.; Paoletti, M.G. Effects of Different Concentrations of Glyphosate (Roundup 360®) on Earthworms (Octodrilus Complanatus, Lumbricus Terrestris and Aporrectodea Caliginosa) in Vineyards in the North-East of Italy. Appl. Soil Ecol. 2018, 123, 802–808. [Google Scholar] [CrossRef]
- Albers, C.N.; Banta, G.T.; Hansen, P.E.; Jacobsen, O.S. The Influence of Organic Matter on Sorption and Fate of Glyphosate in Soil–Comparing Different Soils and Humic Substances. Environ. Pollut. 2009, 157, 2865–2870. [Google Scholar] [CrossRef]
- Gros, P.; Ahmed, A.; Kühn, O.; Leinweber, P. Glyphosate Binding in Soil as Revealed by Sorption Experiments and Quantum-Chemical Modeling. Sci. Total Environ. 2017, 586, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Massaccesi, L.; Cartoni Mancinelli, A.; Mattioli, S.; De Feudis, M.; Castellini, C.; Dal Bosco, A.; Marongiu, M.L.; Agnelli, A. Geese Reared in Vineyard: Soil, Grass and Animals Interaction. Animals 2019, 9, 179. [Google Scholar] [CrossRef]
- Gonçalves, P.; Nóbrega, L.; Monteiro, A.; Pedreiras, P.; Rodrigues, P.; Esteves, F. SheepIT, an E-Shepherd System for Weed Control in Vineyards: Experimental Results and Lessons Learned. Animals 2021, 11, 2625. [Google Scholar] [CrossRef]
- Trouillard, M.; Lèbre, A.; Heckendorn, F. Grazing Sheep in Organic Vineyards: An On-Farm Study about Risk of Chronic Copper Poisoning. Sustainability 2021, 13, 12860. [Google Scholar] [CrossRef]
- O’Brien, R.D. Toxic Phosphorous Esters: Chemistry, Metabolism, and Biological Effects; Elsevier: New York, NY, USA, 2016. [Google Scholar]
- Kushwaha, M.; Verma, S.; Chatterjee, S. Profenofos, an Acetylcholinesterase-Inhibiting Organophosphorus Pesticide: A Short Review of Its Usage, Toxicity, and Biodegradation. J. Environ. Qual. 2016, 45, 1478–1489. [Google Scholar] [CrossRef]
- Booth, L.H.; Hodge, S.; O’Halloran, K. Use of Biomarkers in Earthworms to Detect Use and Abuse of Field Applications of a Model Organophosphate Pesticide. Bull. Environ. Contam. Toxicol. 2001, 67, 633–640. [Google Scholar] [CrossRef]
- Reinecke, S.A.; Reinecke, A.J. The Impact of Organophosphate Pesticides in Orchards on Earthworms in the Western Cape, South Africa. Ecotoxicol. Environ. Saf. 2007, 66, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, C.; Barot, S.; Capowiez, Y.; Hedde, M.; Vandenbulcke, F. Pesticides and Earthworms. A Review. Agron. Sustain. Dev. 2013, 34, 199–228. [Google Scholar] [CrossRef]
- Mansour, R.; Belzunces, L.P.; Suma, P.; Zappalà, L.; Mazzeo, G.; Grissa-Lebdi, K.; Russo, A.; Biondi, A. Vine and Citrus Mealybug Pest Control Based on Synthetic Chemicals. A Review. Agron. Sustain. Dev. 2018, 38, 37. [Google Scholar] [CrossRef]
- Brück, E.; Elbert, A.; Fischer, R.; Krueger, S.; Kühnhold, J.; Klueken, A.M.; Nauen, R.; Niebes, J.-F.; Reckmann, U.; Schnorbach, H.-J.; et al. Movento®, an Innovative Ambimobile Insecticide for Sucking Insect Pest Control in Agriculture: Biological Profile and Field Performance. Crop Prot. 2009, 28, 838–844. [Google Scholar] [CrossRef]
- Pandey, S.; Singh, D.K. Total Bacterial and Fungal Population after Chlorpyrifos and Quinalphos Treatments in Groundnut (Arachis Hypogaea L.) Soil. Chemosphere 2004, 55, 197–205. [Google Scholar] [CrossRef]
- Chu, X.; Fang, H.; Pan, X.; Wang, X.; Shan, M.; Feng, B.; Yu, Y. Degradation of Chlorpyrifos Alone and in Combination with Chlorothalonil and Their Effects on Soil Microbial Populations. J. Environ. Sci. 2008, 20, 464–469. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in Crop Losses to Insect Pests in a Warming Climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef]
- Bois, B.; Zito, S.; Calonnec, A.; Ollat, N. Climate vs Grapevine Pests and Diseases Worldwide: The First Results of a Global Survey. J. Int. Sci. Vigne Vin 2017, 51, 133–139. [Google Scholar]
- Daane, K.M.; Vincent, C.; Isaacs, R.; Ioriatti, C. Entomological Opportunities and Challenges for Sustainable Viticulture in a Global Market. Annu. Rev. Entomol. 2018, 63, 193–214. [Google Scholar] [CrossRef]
- Lodovica Gullino, M.; Garibaldi, A.; Gamliel, A.; Katan, J. Soil Disinfestation: From Soil Treatment to Soil and Plant Health. Plant Dis. 2022, 106, 1541–1554. [Google Scholar] [CrossRef]
- Husson, O.; Sarthou, J.-P.; Bousset, L.; Ratnadass, A.; Schmidt, H.-P.; Kempf, J.; Husson, B.; Tingry, S.; Aubertot, J.-N.; Deguine, J.-P.; et al. Soil and Plant Health in Relation to Dynamic Sustainment of Eh and PH Homeostasis: A Review. Plant Soil 2021, 466, 391–447. [Google Scholar] [CrossRef]
- Akanmu, A.O.; Babalola, O.O.; Venturi, V.; Ayilara, M.S.; Adeleke, B.S.; Amoo, A.E.; Sobowale, A.A.; Fadiji, A.E.; Glick, B.R. Plant Disease Management: Leveraging on the Plant-Microbe-Soil Interface in the Biorational Use of Organic Amendments. Front. Plant Sci. 2021, 12, 700507. [Google Scholar] [CrossRef] [PubMed]
- Davis, L. Soil Health, Plant Health and Pests. In Building Soils for Better Crops; Magdoff, F., Es, H., Eds.; Sustainable Agriculture Research and Education (SARE): New York, NY, USA, 2021; pp. 103–116. [Google Scholar]
- Mpelasoka, B.; Schachtman, D.; Treeby, M.; Thomas, M. A Review of Potassium Nutrition in Grapevines with Special Emphasis on Berry Accumulation. Aust. J. Grape Wine Res. 2003, 9, 154–168. [Google Scholar] [CrossRef]
- James, A.; Mahinda, A.; Mwamahonje, A.; Rweyemamu, E.W.; Mrema, E.; Aloys, K.; Swai, E.; Mpore, F.J.; Massawe, C. A Review on the Influence of Fertilizers Application on Grape Yield and Quality in the Tropics. J. Plant Nutr. 2022, 46, 2936–2957. [Google Scholar] [CrossRef]
- Skinner, P.W.; Ishii, R.; O’Mahony, M.; Matthews, M. Sensory Attributes of Wines Made from Vines of Differing Phosphorus Status. OENO One 2019, 53, 347–361. [Google Scholar] [CrossRef]
- Spayd, S.E.; Stevens, R.G.; Wample, R.L.; Evans, R.G.; Edwards, C.G.; Webster, D. Impact of Nitrogen Fertilization on Vine Performance and Juice and Wine Composition of “Riesling” Grapes (Vitis Vinifera L.) in Washington State. Acta Hortic. 2000, 512, 65–76. [Google Scholar] [CrossRef]
- Bell, S.-J.; Henschke, P.A. Implications of Nitrogen Nutrition for Grapes, Fermentation and Wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Monder, H.; Maillard, M.; Chérel, I.; Zimmermann, S.D.; Paris, N.; Cuéllar, T.; Gaillard, I. Adjustment of K(+) Fluxes and Grapevine Defense in the Face of Climate Change. Int. J. Mol. Sci. 2021, 22, 10398. [Google Scholar] [CrossRef] [PubMed]
- Krasilnikov, P.; Taboada, M.A. Amanullah Fertilizer Use, Soil Health and Agricultural Sustainability. Agriculture 2022, 12, 462. [Google Scholar] [CrossRef]
- Montoroi, J.-P. Soil Degradation by Salinization. In Ecology and Environmental Science: Reduction of Water Stress and Adaptation to Aridity: Proceedings of the Third Ecoscience Workshop; Gawronski, K., Ziernicka-Wojtaszek, A., Ksiazek, L., Nizinski, G., Eds.; Wydawnictwo Uniwersytetu Rolniczego im. H. Kołłątaja: Cracovie, Poland, 2017; pp. 104–106. [Google Scholar]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Arnó, J.; Rosell, J.R.; Blanco, R.; Ramos, M.C.; Martínez-Casasnovas, J.A. Spatial Variability in Grape Yield and Quality Influenced by Soil and Crop Nutrition Characteristics. Precis. Agric. 2011, 13, 393–410. [Google Scholar] [CrossRef]
- Lacombe, B. Increasing Our Knowledge on Grapevines Physiology to Increase Yield, Quality and Sustainably. Physiol. Plant 2022, 174, e13664. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.-J.; Anderson, M.M.; Wolpert, J.A. Vineyard Nutrient Needs Vary with Rootstocks and Soils. Calif. Agric. 2008, 62, 202–207. [Google Scholar] [CrossRef]
- Maeder, P.; Fliessbach, A.; Dubois, D.; Gunst, L.; Fried, P.; Niggli, U. Soil Fertility and Biodiversity in Organic Farming. Science 2002, 296, 1694–1697. [Google Scholar] [CrossRef]
- Tully, K.L.; McAskill, C. Promoting Soil Health in Organically Managed Systems: A Review. Org. Agric. 2020, 10, 339–358. [Google Scholar] [CrossRef]
- Provost, C.; Pedneault, K. The Organic Vineyard as a Balanced Ecosystem: Improved Organic Grape Management and Impacts on Wine Quality. Sci. Hortic. 2016, 208, 43–56. [Google Scholar] [CrossRef]
- Borsato, E.; Zucchinelli, M.; D’Ammaro, D.; Giubilato, E.; Zabeo, A.; Criscione, P.; Pizzol, L.; Cohen, Y.; Tarolli, P.; Lamastra, L.; et al. Use of Multiple Indicators to Compare Sustainability Performance of Organic vs Conventional Vineyard Management. Sci. Total Environ. 2020, 711, 135081. [Google Scholar] [CrossRef]
- Tangolar, S.; Tangolar, S.; Torun, A.A.; Ada, M.; Göçmez, S. Influence of Supplementation of Vineyard Soil with Organic Substances on Nutritional Status, Yield and Quality of “Black Magic” Grape (Vitis Vinifera L.) and Soil Microbiological and Biochemical Characteristics. OENO One 2020, 54, 1143–1157. [Google Scholar] [CrossRef]
- Di Giacinto, S.; Friedel, M.; Poll, C.; Döring, J.; Kunz, R.; Kauer, R. Vineyard Management System Affects Soil Microbiological Properties. OENO One 2020, 54, 131–143. [Google Scholar] [CrossRef]
- Cernansky, R. Agriculture: State-of-the-Art Soil. Nature 2015, 517, 258–260. [Google Scholar] [CrossRef]
- Omondi, M.O.; Xia, X.; Nahayo, A.; Liu, X.; Korai, P.K.; Pan, G. Quantification of Biochar Effects on Soil Hydrological Properties Using Meta-Analysis of Literature Data. Geoderma 2016, 274, 28–34. [Google Scholar] [CrossRef]
- Razzaghi, F.; Obour, P.B.; Arthur, E. Does Biochar Improve Soil Water Retention? A Systematic Review and Meta-Analysis. Geoderma 2020, 361, 114055. [Google Scholar] [CrossRef]
- Shaheen, S.M.; Mosa, A.; Natasha; Arockiam Jeyasundar, P.G.S.; Hassan, N.E.E.; Yang, X.; Antoniadis, V.; Li, R.; Wang, J.; Zhang, T.; et al. Pros and Cons of Biochar to Soil Potentially Toxic Element Mobilization and Phytoavailability: Environmental Implications. Earth Syst. Environ. 2023, 7, 321–345. [Google Scholar] [CrossRef]
- Baronti, S.; Magno, R.; Maienza, A.; Montagnoli, A.; Ungaro, F.; Vaccari, F.P. Long Term Effect of Biochar on Soil Plant Water Relation and Fine Roots: Results after 10 Years of Vineyard Experiment. Sci. Total Environ. 2022, 851, 158225. [Google Scholar] [CrossRef]
- Genesio, L.; Miglietta, F.; Baronti, S.; Vaccari, F.P. Biochar Increases Vineyard Productivity without Affecting Grape Quality: Results from a Four Years Field Experiment in Tuscany. Agric. Ecosyst. Environ. 2015, 201, 20–25. [Google Scholar] [CrossRef]
- Mortvedt, J.J. Heavy Metal Contaminants in Inorganic and Organic Fertilizers; Springer: Cham, Switzerland, 1996; Volume 43. [Google Scholar]
- Roberts, D.; Nachtegaal, M.; Sparks, D.L. Speciation of Metals in Soils. In Chemical Processes in Soils; Tabatabai, M.A., Sparks, D.L., Eds.; Soil Science Society of America: Madison, WI, USA, 2005; pp. 619–654. [Google Scholar]
- Yao, L.-X.; Li, G.-L.; Tu, S.-H.; Gavin, S.; He, Z.-H. Salinity of Animal Manure and Potential Risk of Secondary Soil Salinization through Successive Manure Application. Sci. Total Environ. 2007, 383, 106–114. [Google Scholar] [CrossRef]
- Olego, M.A.; Visconti, F.; Quiroga, M.J.; de Paz, J.M.; Garzón-Jimeno, E. Assessing the Effects of Soil Liming with Dolomitic Limestone and Sugar Foam on Soil Acidity, Leaf Nutrient Contents, Grape Yield and Must Quality in a Mediterranean Vineyard. Span. J. Agric. Res. 2016, 14, e1102. [Google Scholar] [CrossRef]
- Foy, C.D. Physiological Effects of Hydrogen, Aluminum, and Manganese Toxicities in Acid Soil. Soil Acidity Liming 2015, 12, 57–97. [Google Scholar] [CrossRef]
- Hue, N. Soil Acidity: Development, Impacts, and Management. In Structure and Functions of Pedosphere; Springer Nature: Cham, Switzerland, 2022; pp. 103–131. ISBN 9789811687709. [Google Scholar]
- Skinner, P.W.; Matthews, M.A. Reproductive Development in Grape (Vitis Vinifera L.) under Phosphorus-Limited Conditions. Sci. Hortic. 1989, 38, 49–60. [Google Scholar] [CrossRef]
- Čoga, L.; Slunjski, S.; Herak Ćustić, M.; Maslać, J.; Petek, M.; Ćosić, T.; Pavlović, I. Influence of Soil Reaction on Phosphorus, Potassium, Calcium and Magnesium Dynamics in Grapevine (Vitis Vinifera L.). Agric. Conspec. Sci. 2009, 74, 39–43. [Google Scholar]
- Quiroga, M.J.; Olego, M.Á.; Sánchez-García, M.; Medina, J.E.; Visconti, F.; Coque, J.J.R.; Garzón Jimeno, J.E. Effects of Liming on Soil Properties, Leaf Tissue Cation Composition and Grape Yield in a Moderately Acid Vineyard Soil. Influence on Must and Wine Quality. OENO One 2017, 51, 343. [Google Scholar] [CrossRef]
- Tang, C. Causes and Management of Subsoil Acidity. In Proceedings of the SuperSoil 2004: 3rd Australian New Zealand Soils Conference, Camperdown, NSW, Australia, 5 December 2004; University of Sydney: Camperdown, Sydney, NSW, Australia, 2004; pp. 1–7. [Google Scholar]
- Grapevine Rootstock Selector Tool—Rootstock Selector. Available online: https://grapevinerootstock.com/ (accessed on 31 January 2024).
- Van Leeuwen, C.; de Rességuier, L. Major Soil-Related Factors in Terroir Expression and Vineyard Siting. Elements 2018, 14, 159–165. [Google Scholar] [CrossRef]
- Ferretti, C.G. Relationship between the Geology, Soil Assessment, and Terroir of Gewürtztraminer Vineyards: A Case Study in the Dolomites of Northern Italy. Catena 2019, 179, 74–84. [Google Scholar] [CrossRef]
- Jones, G.V. Grapevines in a Changing Environment. In Grapevine in a Changing Environment; Gerós, H., Chaves, M., Medrano Gil, H., Delrot, S., Eds.; Wiley: Hoboken, NJ, USA, 2015; pp. 1–17. [Google Scholar]
- Gobbi, A.; Acedo, A.; Imam, N.; Santini, R.G.; Ortiz-Álvarez, R.; Ellegaard-Jensen, L.; Belda, I.; Hansen, L.H. A Global Microbiome Survey of Vineyard Soils Highlights the Microbial Dimension of Viticultural Terroirs. Commun. Biol. 2022, 5, 241. [Google Scholar] [CrossRef]
- Bonfante, A.; Brillante, L. Terroir Analysis and Its Complexity. OENO One 2022, 56, 375–388. [Google Scholar] [CrossRef]
- Reynard, J.-S.; Zufferey, V.; Nicol, G.-C.; Murisier, F. Vine Water Status as a Parameter of the « terroir » Effect under the Non-Irrigated Conditions of the Vaud Viticultural Area (Switzerland). OENO One 2011, 45, 139. [Google Scholar] [CrossRef]
- Jones, G.V. The Climate Component of Terroir. Elements 2018, 14, 167–172. [Google Scholar] [CrossRef]
- Van Leeuwen, C. Soils and Terroir Expression in Wines. In Soil and Culture; Springer Netherlands: Dordrecht, The Netherlands, 2010; pp. 453–465. [Google Scholar]
- Bohmrich, R. Terroir: Competing Perspectives on the Roles of Soil, Climate and People. J. Wine Res. 1996, 7, 33–46. [Google Scholar] [CrossRef]
- Certini, G.; Scalenghe, R. The Crucial Interactions between Climate and Soil. Sci. Total Environ. 2023, 856, 159169. [Google Scholar] [CrossRef]
- Coli, M.S.; Rangel, A.G.P.; Souza, E.S.; Oliveira, M.F.; Chiaradia, A.C.N. Chloride Concentration in Red Wines: Influence of Terroir and Grape Type. Food Sci. Technol. 2015, 35, 95–99. [Google Scholar] [CrossRef]
- Zhou-Tsang, A.; Wu, Y.; Henderson, S.W.; Walker, A.R.; Borneman, A.R.; Walker, R.R.; Gilliham, M. Grapevine Salt Tolerance. Aust. J. Grape Wine Res. 2021, 27, 149–168. [Google Scholar] [CrossRef]
- Wheeler, S.; Pickering, G. Effects of Soil Management Techniques on Grape and Wine Quality. In Fruits: Growth, Nutrition and Quality; Dris, R., Ed.; Wfl: Helsinki, Finland, 2005; pp. 195–208. [Google Scholar]
- Tomasi, D.; Marcuzzo, P.; Nardi, T.; Lonardi, A.; Lovat, L.; Flamini, R.; Mian, G. Influence of Soil Chemical Features on Aromatic Profile of V. vinifera Cv. Corvina Grapes and Wines: A Study-Case in Valpolicella Area (Italy) in a Calcareous and Non-Calcareous Soil. Agriculture 2022, 12, 1980. [Google Scholar] [CrossRef]
- Coller, E.; Cestaro, A.; Zanzotti, R.; Bertoldi, D.; Pindo, M.; Larger, S.; Albanese, D.; Mescalchin, E.; Donati, C. Microbiome of Vineyard Soils Is Shaped by Geography and Management. Microbiome 2019, 7, 140. [Google Scholar] [CrossRef] [PubMed]
- Nanetti, E.; Palladino, G.; Scicchitano, D.; Trapella, G.; Cinti, N.; Fabbrini, M.; Cozzi, A.; Accetta, G.; Tassini, C.; Iannaccone, L.; et al. Composition and Biodiversity of Soil and Root-Associated Microbiome in Vitis vinifera Cultivar Lambrusco Distinguish the Microbial Terroir of the Lambrusco DOC Protected Designation of Origin Area on a Local Scale. Front. Microbiol. 2023, 14, 1108036. [Google Scholar] [CrossRef]
- Gilbert, J.A.; van der Lelie, D.; Zarraonaindia, I. Microbial Terroir for Wine Grapes. Proc. Natl. Acad. Sci. USA 2014, 111, 5–6. [Google Scholar] [CrossRef]
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial Terroir and Food Innovation: The Case of Yeast Biodiversity in Wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef]
- Vink, S.N.; Dini-Andreote, F.; Höfle, R.; Kicherer, A.; Salles, J.F. Interactive Effects of Scion and Rootstock Genotypes on the Root Microbiome of Grapevines (Vitis Spp. L.). Appl. Sci. 2021, 11, 1615. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, R.; Qin, Q.; Sun, Q. Soil Affected the Variations in Grape and Wine Properties along the Eastern Foot of Helan Mountain, China. Acta Agric. Scand. B Soil Plant Sci. 2019, 69, 494–502. [Google Scholar] [CrossRef]
- Vaudour, E.; Shaw, A. A Worldwide Perspective on Viticultural Zoning. S. Afr. J. Enol. Vitic. 2005, 26, 106–116. [Google Scholar] [CrossRef]
- Villat, J. The Roots of a Better Future: Regenerative Viticulture. Available online: https://portoprotocol.com/the-roots-of-a-better-future-regenerative-viticulture/ (accessed on 11 December 2023).
- Bezuidenhout, C.N.; van Antwerpen, R.; Berry, S.D. An Application of Principal Component Analyses and Correlation Graphs to Assess Multivariate Soil Health Properties. Soil Sci. 2012, 177, 498–505. [Google Scholar] [CrossRef]
- Smith, J.L.; Halvorson, J.J.; Papendick, R.I. Using Multiple-Variable Indicator Kriging for Evaluating Soil Quality. Soil Sci. Soc. Am. J. 1993, 57, 743–749. [Google Scholar] [CrossRef]
- Andrews, S.S.; Karlen, D.L.; Cambardella, C.A. The Soil Management Assessment Framework. Soil Sci. Soc. Am. J. 2004, 68, 1945–1962. [Google Scholar] [CrossRef]
- Zornoza, R.; Acosta, J.A.; Bastida, F.; Domínguez, S.G.; Toledo, D.M.; Faz, A. Identification of Sensitive Indicators to Assess the Interrelationship between Soil Quality, Management Practices and Human Health. Soil 2015, 1, 173–185. [Google Scholar] [CrossRef]
- Moebius-Clune, B.N.; Moebius-Clune, D.J.; Gugino, B.K.; Idowu, O.J.; Schindelbeck, R.R.; Ristow, A.J.; Van Es, H.M.; Thies, J.E.; Shayler, H.A.; Mcbride, M.B.; et al. Comprehensive Assessment of Soil Health—The Cornell Framework; 3.2; Cornell University: Geneva, NY, USA, 2016; ISBN 0967650763. [Google Scholar]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil Enzyme Activities as Biological Indicators of Soil Health. Rev. Environ. Health 2003, 18, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, S.J.; Ezernieks, V.; Yen, A.L. NMR-Based Metabolomics Using Earthworms as Potential Indicators for Soil Health. Metabolomics 2008, 5, 95–107. [Google Scholar] [CrossRef]
- Chae, Y.; Cui, R.; Woong Kim, S.; An, G.; Jeong, S.-W.; An, Y.-J. Exoenzyme Activity in Contaminated Soils before and after Soil Washing: SS-Glucosidase Activity as a Biological Indicator of Soil Health. Ecotoxicol. Environ. Saf. 2017, 135, 368–374. [Google Scholar] [CrossRef]
- Madejon, E.; Moreno, F.; Murillo, J.; Pelegrin, F. Soil Biochemical Response to Long-Term Conservation Tillage under Semi-Arid Mediterranean Conditions. Soil Tillage Res. 2007, 94, 346–352. [Google Scholar] [CrossRef]
- Gong, W.; Yan, X.; Wang, J.; Hu, T.; Gong, Y. Long-Term Manure and Fertilizer Effects on Soil Organic Matter Fractions and Microbes under a Wheat–Maize Cropping System in Northern China. Geoderma 2009, 149, 318–324. [Google Scholar] [CrossRef]
- Niemeyer, J.C.; Lolata, G.B.; de Carvalho, G.M.; Da Silva, E.M.; Sousa, J.P.; Nogueira, M.A. Microbial Indicators of Soil Health as Tools for Ecological Risk Assessment of a Metal Contaminated Site in Brazil. Appl. Soil Ecol. 2012, 59, 96–105. [Google Scholar] [CrossRef]
- Norkaew, S.; Miles, R.J.; Brandt, D.K.; Anderson, S.H. Effects of 130 Years of Selected Cropping Management Systems on Soil Health Properties for Sanborn Field. Soil Sci. Soc. Am. J. 2019, 83, 1479–1490. [Google Scholar] [CrossRef]
- Caudle, C.; Osmond, D.; Heitman, J.; Ricker, M.; Miller, G.; Wills, S. Comparison of Soil Health Metrics for a Cecil Soil in the North Carolina Piedmont. Soil Sci. Soc. Am. J. 2020, 84, 978–993. [Google Scholar] [CrossRef]
- Ye, R.; Parajuli, B.; Szogi, A.A.; Sigua, G.C.; Ducey, T.F. Soil Health Assessment after 40 Years of Conservation and Conventional Tillage Management in Southeastern Coastal Plain Soils. Soil Sci. Soc. Am. J. 2021, 85, 1214–1225. [Google Scholar] [CrossRef]
- Fine, A.K.; van Es, H.M.; Schindelbeck, R.R. Statistics, Scoring Functions, and Regional Analysis of a Comprehensive Soil Health Database. Soil Sci. Soc. Am. J. 2017, 81, 589–601. [Google Scholar] [CrossRef]
- Liptzin, D.; Norris, C.E.; Cappellazzi, S.B.; Bean, G.M.; Cope, M.; Greub, K.L.H.; Rieke, E.L.; Tracy, P.W.; Aberle, E.; Ashworth, A.; et al. An Evaluation of Carbon Indicators of Soil Health in Long-Term Agricultural Experiments. Soil Biol. Biochem. 2022, 172, 108708. [Google Scholar] [CrossRef]
- Katayama, A.; Bhula, R.; Burns, G.R.; Carazo, E.; Felsot, A.; Hamilton, D.; Harris, C.; Kim, Y.-H.; Kleter, G.; Koedel, W.; et al. Bioavailability of Xenobiotics in the Soil Environment. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–86. [Google Scholar]
- Sojka, R.E.; Upchurch, D.R. Reservations Regarding the Soil Quality Concept. Soil Sci. Soc. Am. J. 1999, 63, 1039–1054. [Google Scholar] [CrossRef]
- Chamberlain, L.A.; Aguayo, T.; Zerega, N.J.C.; Dybzinski, R.; Egerton-Warburton, L.M. Rapid Improvement in Soil Health Following the Conversion of Abandoned Farm Fields to Annual or Perennial Agroecosystems. Front. Sustain. Food Syst. 2022, 6, 1010298. [Google Scholar] [CrossRef]
- Barthès, B.; Roose, E. Aggregate Stability as an Indicator of Soil Susceptibility to Runoff and Erosion; Validation at Several Levels. Catena 2002, 47, 133–149. [Google Scholar] [CrossRef]
- Kemper, W.D.; Rosenau, R.C. Aggregate Stability and Size Distribution. Methods of Soil Analysis, Part 1. In Physical and Mineralogical Methods; Klute, A., Ed.; SSSA: Madison, WI, USA, 2018; pp. 425–442. [Google Scholar]
- Amézketa, E. Soil Aggregate Stability: A Review. J. Sustain. Agric. 1999, 14, 83–151. [Google Scholar] [CrossRef]
- Rieke, E.L.; Bagnall, D.K.; Morgan, C.L.S.; Flynn, K.D.; Howe, J.A.; Greub, K.L.H.; Mac Bean, G.; Cappellazzi, S.B.; Cope, M.; Liptzin, D.; et al. Evaluation of Aggregate Stability Methods for Soil Health. Geoderma 2022, 428, 116156. [Google Scholar] [CrossRef]
- Suleiman, A.A.; Ritchie, J.T. Estimating Saturated Hydraulic Conductivity from Soil Porosity. Trans. ASAE 2001, 44, 235. [Google Scholar] [CrossRef]
- Li, Z.; Wu, P.; Feng, H.; Zhao, X.; Huang, J.; Zhuang, W. Simulated Experiment on Effect of Soil Bulk Density on Soil Infiltration Capacity. Nongye Gongcheng Xuebao/Trans. Chin. Soc. Agric. Eng. 2009, 25, 40–45. [Google Scholar]
- Burns, K.N.; Bokulich, N.A.; Cantu, D.; Greenhut, R.F.; Kluepfel, D.A.; O’Geen, A.T.; Strauss, S.L.; Steenwerth, K.L. Vineyard Soil Bacterial Diversity and Composition Revealed by 16S RRNA Genes: Differentiation by Vineyard Management. Soil Biol. Biochem. 2016, 103, 337–348. [Google Scholar] [CrossRef]
- Viers, J.H.; Williams, J.N.; Nicholas, K.A.; Barbosa, O.; Kotzé, I.; Spence, L.; Webb, L.B.; Merenlender, A.; Reynolds, M. Vinecology: Pairing Wine with Nature. Conserv. Lett. 2013, 6, 287–299. [Google Scholar] [CrossRef]
- Stoleru, V.; Liliana, R.; Mihai, M. Fertilization with Green Manure on Chasselas Dore Grape Vine as an Alternative for Sustainable Agriculture. J. Food Agric. Environ. 2011, 9, 8. [Google Scholar]
- Cesco, S.; Pii, Y.; Borruso, L.; Orzes, G.; Lugli, P.; Mazzetto, F.; Genova, G.; Signorini, M.; Brunetto, G.; Terzano, R.; et al. A Smart and Sustainable Future for Viticulture Is Rooted in Soil: How to Face Cu Toxicity. Appl. Sci. 2021, 11, 907. [Google Scholar] [CrossRef]
- MacKie, K.A.; Müller, T.; Kandeler, E. Remediation of Copper in Vineyards—A Mini Review. Environ. Pollut. 2012, 167, 16–26. [Google Scholar] [CrossRef]
- Komárek, M.; Čadková, E.; Chrastný, V.; Bordas, F.; Bollinger, J.C. Contamination of Vineyard Soils with Fungicides: A Review of Environmental and Toxicological Aspects. Environ. Int. 2010, 36, 138–151. [Google Scholar] [CrossRef]
- Casson, A.; Ortuani, B.; Giovenzana, V.; Brancadoro, L.; Corsi, S.; Gharsallah, O.; Guidetti, R.; Facchi, A. A Multidisciplinary Approach to Assess Environmental and Economic Impact of Conventional and Innovative Vineyards Management Systems in Northern Italy. Sci. Total Environ. 2022, 838, 156181. [Google Scholar] [CrossRef]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate Change and Global Wine Quality. Clim. Change 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Webb, L.B.; Whetton, P.H.; Barlow, E.W.R. Climate Change and Winegrape Quality in Australia. Clim. Res. 2008, 36, 99–111. [Google Scholar] [CrossRef]
- Hall, A.; Mathews, A.J.; Holzapfel, B.P. Potential Effect of Atmospheric Warming on Grapevine Phenology and Post-Harvest Heat Accumulation across a Range of Climates. Int. J. Biometeorol. 2016, 60, 1405–1422. [Google Scholar] [CrossRef]
- Lehmann, J.; Bossio, D.A.; Kögel-Knabner, I.; Rillig, M.C. The Concept and Future Prospects of Soil Health. Nat. Rev. Earth Environ. 2020, 1, 544–553. [Google Scholar] [CrossRef]
- Karlen, D.L.; Veum, K.S.; Sudduth, K.A.; Obrycki, J.F.; Nunes, M.R. Soil Health Assessment: Past Accomplishments, Current Activities, and Future Opportunities. Soil Tillage Res. 2019, 195, 104365. [Google Scholar] [CrossRef]
- Montalvo-Falcón, J.V.; Sánchez-García, E.; Marco-Lajara, B.; Martínez-Falcó, J. Sustainability Research in the Wine Industry: A Bibliometric Approach. Agronomy 2023, 13, 871. [Google Scholar] [CrossRef]
- Villat, J. Down to Earth: Identifying and Promoting Regenerative Viticulture Practices for Soil and Human Health. Ph.D. Thesis, Harvard University, Cambridge, MA, USA, 2021. [Google Scholar]
Figure 1.
Energy and matter exchange relationships between the four environmental compartments and the places soil and humanity occupy within, respectively, land and biota. Circles and sectors sizes are not at scale.
Figure 1.
Energy and matter exchange relationships between the four environmental compartments and the places soil and humanity occupy within, respectively, land and biota. Circles and sectors sizes are not at scale.

Figure 2.
The concepts of soil health, soil quality, and soil fertility as defined by the soil ecological functions they totally or partially embrace.
Figure 2.
The concepts of soil health, soil quality, and soil fertility as defined by the soil ecological functions they totally or partially embrace.


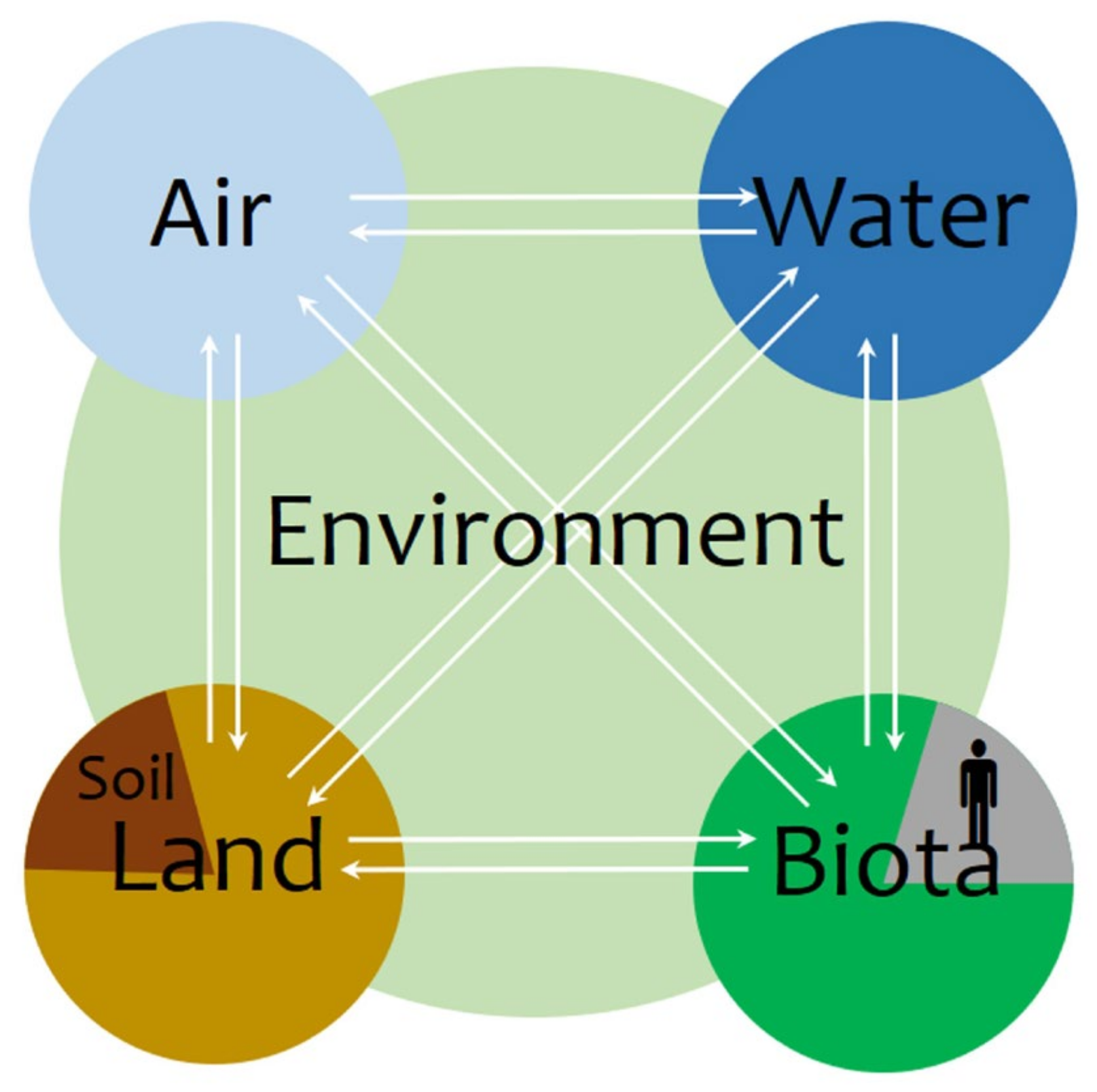{kind=link}
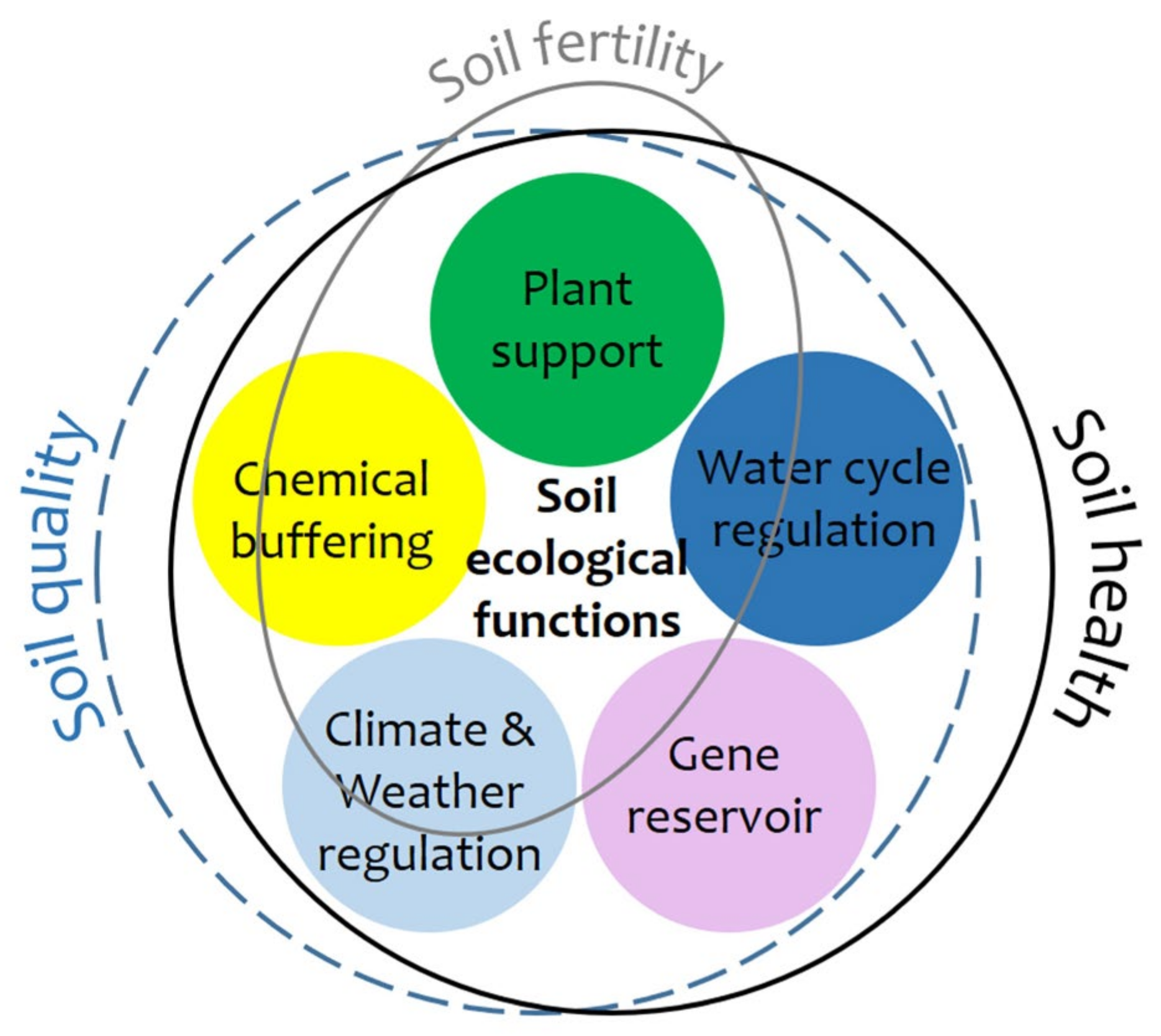{kind=link}
Table 1.
Impact of soil agricultural management practices on the microbial biomass (↑ increase in population; ↓ decrease in population; ↑↓ controversial results) [20,28,29,30,31].
| Soil Practice | Effect on Soil Microbial Population |
|---|---|
| Retention of crop residues | ↑ |
| Application of organic amendments to soil | ↑↓ |
| Minimum tillage practices or absence of ploughing | ↑↓ |
| Irrigation of crops in rain-limited ecosystems | ↑ |
| Application of fungicides to soil | ↓ |
| Application of herbicides to soil | ↑↓ |
| Liming of soil to raise pH on acidic soils | ↑ |
| Unbalanced fertilization | ↓ |
Table 2.
Distinctive characteristics between rhizosphere and bulk soil (↑ this soil characteristic is increased; ↓ this soil characteristic is decreased) [20,48,49,50,51,52,53].
| Characteristic | Rhizosphere | Bulk Soil |
|---|---|---|
| Concentrations of easily degradable substances (sugars, organic acids, and amino acids) | ↑ | ↓ |
| Diversity of microorganisms | ↓ | ↑ |
| Abundance of specific microbial taxa | ↑ | ↓ |
| Phytotoxicity of heavy metals | ↓ | ↑ |
| Enzyme activities | ↑ | ↓ |
| Nutrient availability | ↑ | ↓ |
| Soil pH | ↑ (alkalinisation) | ↓ |
| Water retention because of changes in soil structure | ↑ | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Visconti, F.; López, R.; Olego, M.Á. The Health of Vineyard Soils: Towards a Sustainable Viticulture. Horticulturae 2024, 10, 154. https://doi.org/10.3390/horticulturae10020154
AMA Style
Visconti F, López R, Olego MÁ. The Health of Vineyard Soils: Towards a Sustainable Viticulture. Horticulturae. 2024; 10(2):154. https://doi.org/10.3390/horticulturae10020154
Chicago/Turabian StyleVisconti, Fernando, Roberto López, and Miguel Ángel Olego. 2024. "The Health of Vineyard Soils: Towards a Sustainable Viticulture" Horticulturae 10, no. 2: 154. https://doi.org/10.3390/horticulturae10020154
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.