
Rotational Strip Bean and Celery Intercropping Alters the Microbial Community to Improve Crop Yield and Soil Nutrients

1
Department of Horticulture, Northeast Agricultural University, Harbin 150030, China
2
Key Laboratory of Cold Area Vegetable Biology, Northeast Agricultural University, Harbin 150030, China
*
Author to whom correspondence should be addressed.
Horticulturae 2024, 10(5), 432; https://doi.org/10.3390/horticulturae10050432
Submission received: 19 March 2024
/
Revised: 17 April 2024
/
Accepted: 21 April 2024
/
Published: 24 April 2024
(This article belongs to the Section Protected Culture)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Rotational strip intercropping involves the strategic combination of stubble intercropping and the rotational placement of intercropped strips, which can improve crop yield. Here, we evaluated the effects of rotational strip bean (Phaseolus vulgaris L.) and celery (Apium graveolens L.) intercropping with bed ratios of 1:1 and 2:2 on crop yield, soil microbial community and nutrition. The 1:1 ratio of rotational strip bean and celery intercropping had a higher yield than the 2:2 ratio. The 1:1 rotational strip intercropping markedly improved the nitrate nitrogen and available phosphorus contents of the beans, and the nitrate nitrogen and available potassium contents of the celery. The 2:2 rotational strip intercropping reduced the bacterial α diversity of the beans, and the 1:1 and 2:2 rotational strip intercropping enhanced the fungal α diversity of both the beans and the celery. Both planting patterns also altered the microbial communities found on the bean and celery plants. Microorganisms enriched in the rhizosphere of the bean and celery plants significantly increased the soil nutrient content and yield in the 1:1 rotational strip intercropping, and included the genera Achromobacter, Luteibacter, Stenotrophomonas, Fusarium, Flavobacterium and Preussia in the bean rhizosphere, and Alternaria, Arthrographis and Pluteus, Sphingopyxis and Clostridium_sensu_stricto_1 in the celery rhizosphere.
1. Introduction
Plant diversity plays a crucial role in the stability of ecosystems [1,2]. In agricultural ecosystems, plant diversity is widely utilized to address a series of challenges posed by continuous cropping. Thus, crop rotation, intercropping and the use of cover crops are common practices in agricultural systems [3,4,5]. In rotation and intercropping systems, some plants can inhibit plant pests and diseases and improve crop yield and growth by increasing soil nutrients and improving the soil microbial environment. In these rotational and intercropping systems, legumes play a role in increasing the yield of other cash crops and inhibiting disease, such as in wheat, corn and cotton [6,7,8,9,10,11]. In order to combine the advantages of intercropping and rotation, a new cultivation model was proposed: rotational strip intercropping, in which two crops are intercropped on a wide strip, and their planting positions are alternated on the same land each season or year [12]. When choosing the crop combination for rotational strip intercropping, it should be matched according to the growth habit of the crop and the role of growth. Some rotational strip intercropping patterns, such as maize and peanuts, peanuts and cotton and cotton and soybeans rotational strip intercropping, have been successful [11,13,14,15,16]. However, the application value of rotational strip bean and celery intercropping in agricultural production has not yet been determined.
Rhizosphere microorganisms play an important role in crop growth and yield, and rotational and intercropping systems increase agriculture ecosystem diversity and alter the soil microbial environment. Compared with the continuous cropping of peanuts, the bacterial diversity and richness of the rhizosphere in an intercropping system using maize and peanut were significantly increased after rotation. The main bacterial phyla affected were Proteobacteria, Acidobacteria, Firmicutes, Bacteroidetes and actinobacteria [17,18]. Compared with cucumber monoculture, rotation increased rhizosphere bacterial diversity, but decreased fungal diversity and abundance, but also significantly increased the relative abundance of bacterial genes representing Flavobacterium and Ohtaekwangia and the fungal genes Monographella and Operculomyces in the cucumber rhizosphere [19]. Compared with melon monoculture, rotation increased the relative diversity and abundance of fungi and bacteria found in the melon plants, and decreased the relative abundance of Fusarium [20]. Intercropping of potato onion and tomato increased the diversity and richness of bacteria and fungi, increased the relative abundance of Acidobacteria-Subgroup-6, Arthrobacter, Bacillus and Pseudomonas and decreased the relative abundance of Cladosporium [21]. Intercropping of wheat and cucumber significantly increased the diversity of cucumber rhizosphere bacteria, and the relative abundance of potentially beneficial plant bacteria (Pseudomonas, Haliangium and Archangium) was significantly improved [22]. Compared with maize and peanut monoculture, rotational strip intercropping significantly increased both bacterial richness and diversity [16]. Rotational strip peanut and cotton intercropping increased the bacterial richness and diversity of the plants, and increased the relative abundance of Nitrospirae and Bryobacter in the peanut rhizosphere, and the relative abundance of Bacillus, Sediminibacterium and Pseudomonas in the cotton rhizosphere [11].
Soil microbial communities, as major drivers of the ecosystem’s function, are regulated by plant diversity through changes in resource availability or environmental conditions [23,24,25]. Soil microorganisms are one of the largest reservoirs of biodiversity and participate in various ecological processes in terrestrial ecosystems [26,27,28,29]. For example, microorganisms perform soil decomposition and mediate C, N, S and P biogeochemical cycles [26,28]. Plant–microbial interactions can also have an impact on soil properties, which can vary depending on crop species and cultivation patterns. It had been reported that the relative abundance of Fimbriimonadia is an important variable in predicting the levels of KCl, available phosphorus (AP) and organic matter contents, while the relative abundance of Cytophagia and Phycisphaerae are important variables in predicting available potassium (AK) contents. Additionally, the relative abundance of Bacilli was an important variable in predicting AP contents in maize soil. The relative abundance of Clostridia and Bacilli were the most important variables in predicting NH4+−N contents, while the relative abundance of Anaerolineae and Ktedonobacteria were the predictors of AK and AP contents, respectively [29]. It had been reported that Arthrobacter is positively correlated with NO3−−N, AK and total nitrogen (TN) contents in cover crops [30]. Agromyces, Arthrobacter, Bacillus, Lysobacter and Paenibacillus play an important role in nitrogen fixation, while Gemmatimonas, Heliobacillus, Natronocella and Sorangiumincrease increase the nitrogen-fixing capacity of nitrogen-fixing bacteria through indirect means in an intercropping pattern using faba bean and maize [31]. The abundance of Sphingomonas was positively correlated with available phosphorus (AP), total phosphorus (TP), total potassium (TK) and soil organic carbon (SOC) contents, while the abundance of Gemmatimonas was positively correlated with AP and TP contents in rotational strip maize and peanut intercropping [16]. However, there are few studies on the effects of microorganisms on soil nutrient cycling in the rotational strip intercropping system.
In this study, we aimed to address the following questions: (1) Which rotational strip intercropping pattern, 1:1 or 2:2, is more suitable for beans (Triticum aestivum L.) and celery (Apium graveolens L.) in intensive agricultural production? (2) How do two cultivation patterns affect soil microbes and nutrients? To answer these questions, we evaluated the crop yield, soil microbial community diversity and nutrient contents in a rotational strip bean and celery intercropping system. We first studied the effects of rotational strip intercropping on bean and celery yields and soil nutrients. The effects of rotational strip intercropping on soil bacterial and fungal community diversity were studied by high-throughput amplification sequencing. We formed the following hypotheses: (1) the crop yield can be increased by rotational strip intercropping of bean and celery, with the 1:1 ratio being more effective than the 2:2 ratio in rotational strip bean and celery intercropping; (2) soil microbial diversity will be affected by rotational strip intercropping of bean and celery; (3) there will be a significant correlation between soil microorganisms, soil nutrients and yield in the rotational strip intercropping of bean and celery.
2. Materials and Methods
2.1. Experimental Design and Treatments
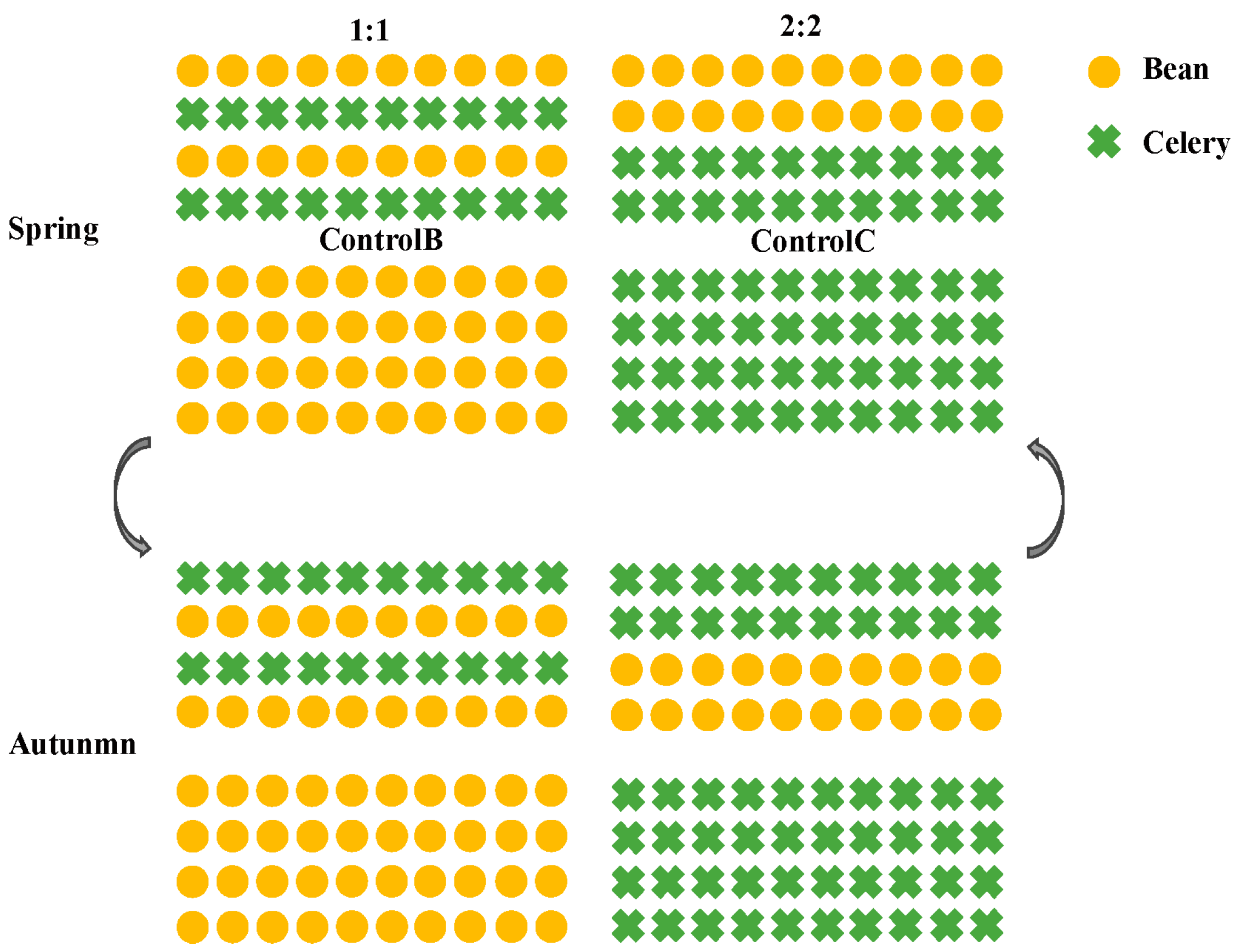
Field experiments were conducted from 2019 to 2021 at Northeast Agricultural University, Harbin, China (45°419 N, 126°379 E). The experiment was conducted twice a year in a greenhouse (spring crop: 25 April~15 July; autumn crop: 25 July~20 October). The experiments followed a randomized block design with three replications. The ratios of the number of beds between the beans and celery in the rotational strip intercropping were 1:1 and 2:2. The treatments were as follows: the number of beds split between beans and celery is 1:1; that is, 1 bed of beans and 1 bed of celery intercropped (Bean1:1 and Celery1:1). And the number of beds split between beans and celery is 2:2; that is, 2 beds of beans and 2 beds of celery intercropped (Bean2:2 and Celery2:2) (Figure 1). Continuously monocropped beans and celery were used as controls (ControlB and ControlC). Both crop species were planted in a south–north bed direction. Each experimental plot had an area of 4.5 m2 (5 m × 0.9 m) and the border beds were not used for sampling. Four rows of celery and two rows of beans, respectively, were planted in each bed. Beans were planted where celery was last grown, and celery was planted where beans were last grown; the sole bean and sole celery plots did not change. The row spacing for beans was 60 cm, and plant spacing was 30–35 cm. The row spacing for celery was 15 cm, and plant spacing was 10–12 cm.
2.2. Bean and Celery Yields
Bean and celery yields were measured in fall 2020 and spring and fall 2021. Beans and celery were planted twice a year in the spring and autumn, respectively. Spring planting: celery and beans were planted in late April and harvested in mid-July. Autumn planting: celery and beans were planted in late July and harvested in late October. In spring planting, celery with more consistent growth was selected in mid-June to measure fresh weight and the yield of beans was measured in early July every day, respectively, and 5 plants were selected for each replication. In autumn planting, the fresh weight of celery and the yield of beans were measured in mid-September and early October, respectively. The total yield of each plot was measured and converted to kg·ha−1 to standardize yield measurements.
2.3. Photosynthesis Property
The illuminance and net photosynthetic rate (Pn) were measured on a sunny day during the growth periods of the beans and celery. Illuminance and Pn were measured once at 10 a.m. Illuminance was measured using the Illuminometer TES-1355. The net photosynthetic rate was measured with a Photosynthesizer Li-6400 (Li-Cor Inc., Lincolin, NE, USA).
2.4. Soil Sampling
In the autumn planting of 2021, celery bulk soils and rhizosphere soils were collected in mid-September, while bean bulk soils and rhizosphere soils were collected in early October. Five soil cores (8 cm diameter, 20 cm deep) were randomly collected from each plot, and these five soil cores were mixed to create a composite sample. Thus, there were three composite samples for each treatment. Briefly, bean or celery roots were gently removed from the soil, and soil loosely attached to the bean or celery roots were carefully removed by manual shaking was considered as bulk soil. Then, soil tightly adhering to the roots was removed from the root surface with a sterile brush and considered as rhizosphere soil. Soils were sieved (2 mm), and large stone and plant debris were removed. Bulk soils were stored at 4 °C for soil chemical property analysis, while rhizosphere soils were stored at −80 °C for DNA extraction.
2.5. Soil Nutrients
2.6. Soil DNA Extraction and Quantitative PCR
Total soil DNA was extracted by the Power Soil DNA isolation Kit according to the manufacturer’s instructions (MO BIO laboratories, Garlsbad, CA, USA). We required 0.25 g of soil for each sample. A spectrophotometer (NanoDrop2000, Thermo, Waltham, MA, USA) was used to evaluate the quantity and purity of the DNA. Three separate extractions were carried out for each of the composites of every soil sample and then combined to form a single composite sample of extracted DNA for each soil sample (three composite DNA samples per treatment). Abundances of total bacterial and fungal communities were estimated using SYBR Green quantitative PCR assays conducted with an IQ5 Real-time PCR system (Bio-Rad Lab, Hercules, CA, USA). For the total bacterial community, the primer set 338 F/518 R [33] was used to amplify the partial bacterial 16S rRNA gene. The primer set ITS1 F/ITS4 R [34] was used to amplify the ITS regions of the rRNA gene for the total fungal community, and this process was repeated three times. Purified water was used as a negative control. Sequences of plasmids (pMD18-T) containing the target genes of bacteria and fungi for the standard curves were created with tenfold dilution, respectively. To determine the copy number of the target gene, the threshold cycle values obtained for each sample were compared with the standard curve.
2.7. High-Throughput Amplicon Sequencing and Data Processing
High-throughput amplicon sequencing for the determination of soil bacterial and fungal community structures were conducted. The V3–V4 regions of the bacterial 16 S rRNA gene and the ITS1 regions of the fungal rRNA gene were amplified using the primer sets F338/R806 and ITS1F/ITS2, respectively [35]. Similarly, both forward and reverse primers were tagged with a 6 bp unique barcode to differentiate each sample. Every combination soil DNA sample had three exactly replicated PCR responses and then the PCR products were pooled and purified using an Agarose Gel DNA purification kit (TaKaRa, Dalian, China). The purified amplicons were quantified using a TBS-380 micro fluorometer (Invitrogen, Waltham, MA, USA) and then mixed to achieve equal concentrations in the final mixture. The concluding mixture was then paired-end sequenced (2 × 300) on an Illumina Miseq platform (Majorbio Bio-Pharm Technology Co. Ltd., Shanghai, China).
Raw sequence reads were de-multiplexed, quality-filtered and processed using FLASH [36]. Chimeric structures were recognized and removed by USEARCH11 in QIIME. Operational taxonomic units (OTUs) were collected at 97% resemblance with UPARSE by an agglomerative collecting algorithm [37]. The actual structure of each OTU was taxonomically categorized using the SILVA database issue 128 (bacteria) and Unite databases (fungi) [38,39]. The proposal Assent Numbers PRJNA859279 for bacteria and PRJNA859295 for fungi were obtained from the statistical data in the NCBI-sequence delivered record.
2.8. Statistical Analysis
For all statistical analyses, R software (version 4.2) was used. The Shannon diversity indices, relative abundances and bacterial and fungal alpha diversities were measured. To assess the differences in the structures of bacterial and fungal communities (R vegan package), Principal Coordinates Analysis (PCoA) based on the Bray–Curtis distance variation was performed using the vegan package in R. To examine the effect of rotational strip intercropping ratio on bacterial community structures, Permutational Multivariate Analysis of Variance (PERMANOVA) with Bray–Curtis distance and 9999 random permutations was performed. Sensitive OTUs stimulated by rotational strip intercropping were identified using the indicator species study and likelihood ratio test with the Benjamini–Hochberg (BH) p value adjustment, respectively (R indicspecies and edgeR packages) [40]. The relationships between environmental factors and significantly altered bacterial taxa were establish on the Spearman correlation coefficient (R heat map package). All data were tested for normality and homogeneity of variances to meet the assumptions of normality before statistical analysis. Analysis of variance (ANOVA) followed by Tukey’s HSD test was used to assess the differences between treatments, with p < 0.05 considered statistically significant.
3. Results
3.1. Yields of Bean and Celery under Rotational Strip Intercropping
Treatments of Bean 1:1 and Bean 2:2 significantly improved the bean yield during all three assessments (2020 autumn, 2021 spring and 2021 autumn) compared with ControlB (p < 0.05) (Figure 2a). The yield of Bean 1:1 was higher than that of Bean 2:2 in autumn 2020, and there was no difference between the yields of Bean 1:1 and Bean 2:2 in 2021 (p < 0.05) (Figure 2a).
3.2. Illuminance and Net Photosynthetic Rate of Rotational Strip Bean and Celery Intercropping
The Pns of the Bean1:1 and Bean2:2 treatments were significantly higher than that of the ControlB monoculture in the autumn of 2020 and the spring of 2021. Additionally, the Pn of the Bean1:1 treatment was higher than that of the Bean2:2 treatment (Table S1). Compared with ControlC treatment, the Pns of the Celery1:1 and Celery2:2 treatments were higher. The Pn of Celery 1:1 treatment was higher than that of the Celery2:2 treatment in the autumn of 2021 (Table S1).
The illuminance of the Bean 1:1 treatment was significantly higher than the ControlB and Bean2:2 treatments in the spring of 2021. The illuminance of the Bean1:1 treatment was significantly higher than that of the Bean2:2 treatment in 2020 and 2021, and the Bean2:2 treatment did not show a significant difference compared to the ControlB treatment in 2021 (Table S1). The illuminances of the Celelry1:1 and Celery2:2 treatments were lower than the ControlC treatment, and the illuminance of the Celery1:1 treatment was lower compared to that of the Celery2:2 treatment in 2020 and 2021 (Table S1).
3.3. Soil Nutrients of Rotational Strip Bean and Celery Intercropping
Compared with the ControlB, the Bean1:1 and Bean2:2 treatments significantly increased the AP content in the yields from the autumn of 2020 and 2021, and the AP content of the crops following the Bean1:1 treatment was higher than that of those after the Bean2:2 treatment (p < 0.05). The Bean1:1 treatment significantly increased the content of NO3−-N relative to the bean monoculture during all assessments, and the NO3−-N content of the Bean1:1 treated crops was also higher than that of the Bean2:2 crops (p < 0.05). The Bean2:2 treatment prominently increased the NH4+-N content in 2021 (p < 0.05) (Figure 3a).
NO3−-N and AP contents were higher in the crops treated with Celery1:1 and Celery2:2 than ControlC at the Autumn 2020 and 2021 assessments (p < 0.05) (Figure 3b). Compared to the monoculture of celery, the AK content was significantly reduced in the Celery2:2 group. The NO3−-N and AK contents were higher in the Celery1:1 group compared to Celery2:2 (p < 0.05) (Figure 3b). The NH4+-N content did not show a significant difference between treatments during any of the assessments (Figure 3b).
3.4. Soil Microbial Community Abundances and Diversity
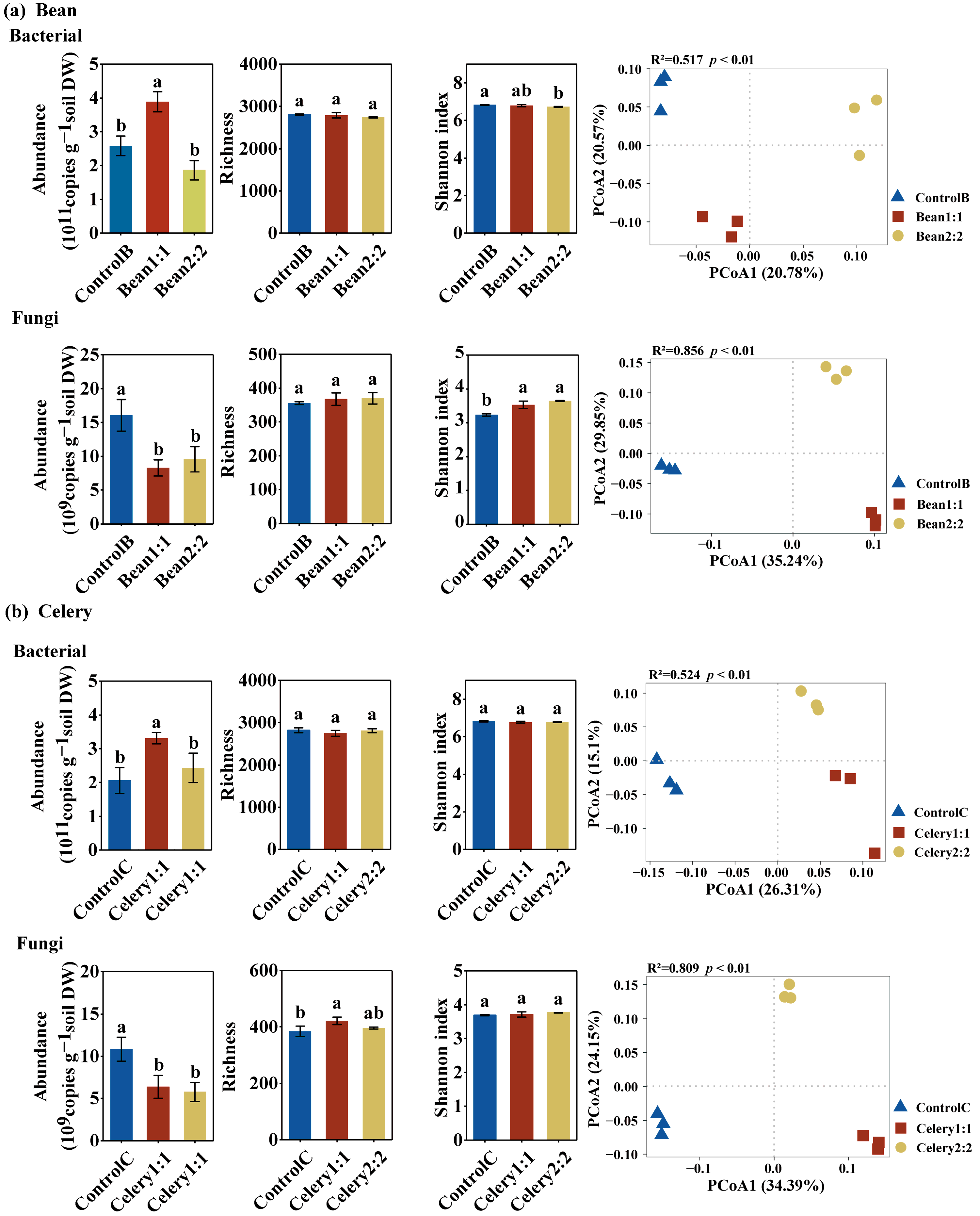
Compared with the monoculture, the 1:1 rotational strip intercropping increased the bacterial abundances of both the bean and celery plants, while the 2:2 rotational strip intercropping was not different. Thus, the bacterial abundance of the 1:1 rotational strip intercropping was higher than that of the 2:2 rotational strip intercropping (p < 0.05) (Figure 4a,b). The fungal abundance was lower in the 1:1 and 2:2 rotational strip intercropping compared to the single culture, but there was no difference between the two rotational treatments (p < 0.05) (Figure 4a,b). In the bacterial community, the Bean2:2 treatment significantly reduced the Shannon index of the bacterial community compared to ControlB, while the Shannon index of the bacterial community in the Bean1:1 crops did not show any significant differences compared to ControlB. (p < 0.05) (Figure 4a). However, the 1:1 and 2:2 rotational strip intercropping had no effect on the alpha diversity of the celery bacteria (Shannon index and richness) (Figure 4b). In terms of the fungal community, compared with the monoculture, both the 1:1 and 2:2 rotational strip intercropping treatments significantly increased the fungal Shannon index of the bean crops, but there was no difference between them (p < 0.05) (Figure 4a,b). Additionally, the 1:1 rotational strip intercropping significantly increased fungal richness. The PERMANOVA analysis revealed that the proportions of rotational strip bean and celery intercropping had significant effects on both bacterial and fungal community beta diversities (Figure 4a,b).
3.5. Bacterial Community Composition in Rotational Strip Intercropping
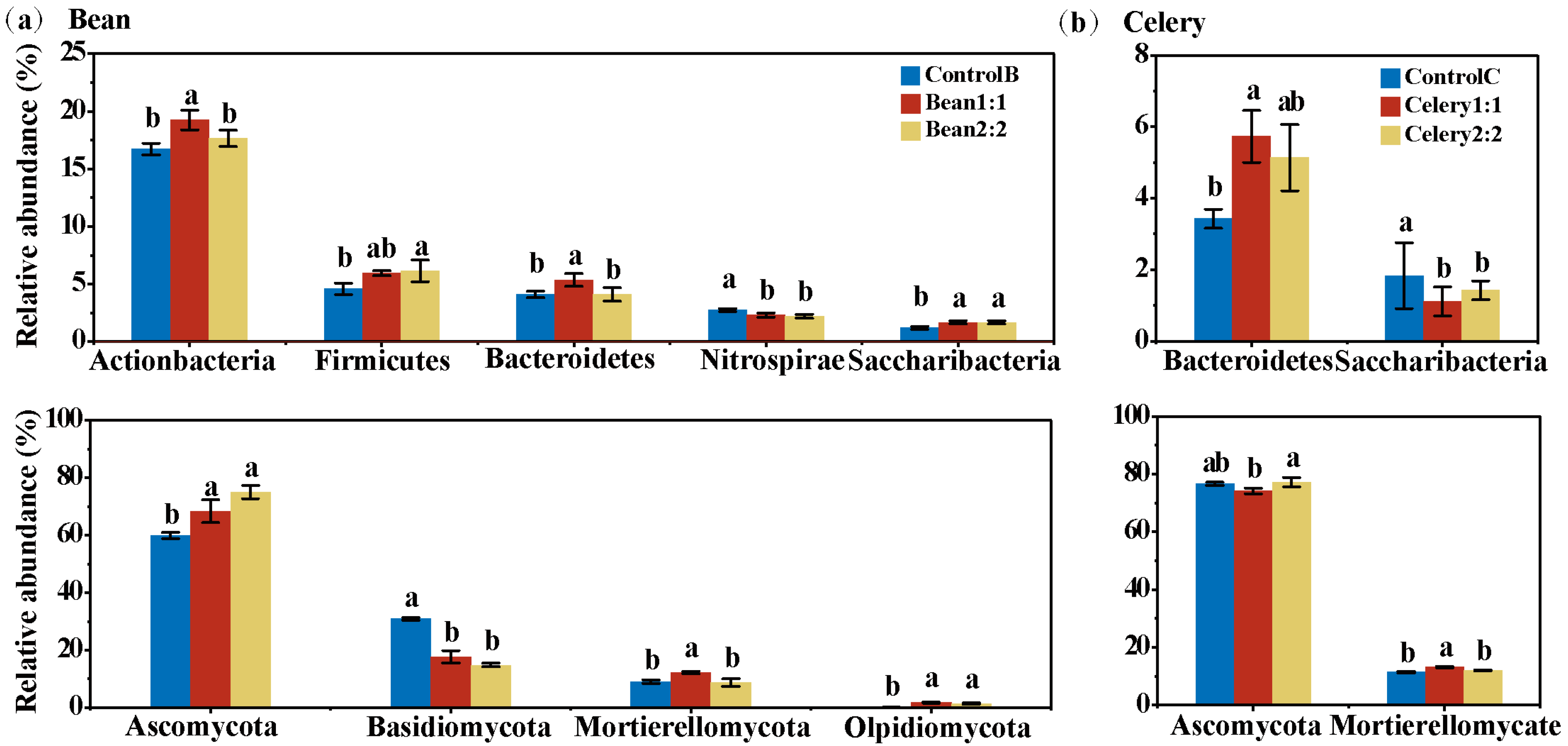
A total of 41 bacterial phyla were observed in the bean and celery samples. The dominant phyla were Proteobacteria, Actinobacteria, Acidobacteria and Chloroflexi, which together accounted for more than 75% and 70% of the bacterial sequences in the bean and celery plants, respectively. Firmicutes, Gemmatimonadetes, Bacteroidetes, Nitrospirae, Verrucomicrobia and Saccharibacteria were also detected at relatively high abundances in the beans and celery (average relative abundance > 1% at in all treatments) (Figure S1a). Compared with the bean monoculture treatment, both Bean1:1 and Bean2:2 had higher relative abundances of Saccharibacteria and had lower relative abundances of Nitrospirae, but there was no difference between the two treatments (Figure 5a). Moreover, the abundance of Actinobacteria and Bacteroidetes in Bean1:1 was significantly higher than that in ControlB and Bean2:2, while the relative abundance of Firmicutes was higher in Bean2:2 than in ControlB (p < 0.05) (Figure 5a). Compared with the celery monoculture treatment, Celery1:1 showed higher relative abundances of Bacteroidetes. The relative abundance of Saccharibacteria was lower in Celery1:1 and Celery2:2 than in celery monoculture (p < 0.05) (Figure 5b).
Taxonomical classification at the genus level revealed that more than 480 bacterial genera were detected in the bean treatments and more than 496 bacterial genera were detected in the celery treatments.
Compared to the bean monoculture treatment, bean intercropping showed a lower relative abundance of Nitrospira and a higher relative abundance of Clostridium_sensu_stricto_1, Rhizobium and Aeromicrobium (p < 0.05). Additionally, RB41 and Streptomyces exhibited significantly higher relative abundances in the rhizosphere of bean under 1:1 rotational strip intercropping (p < 0.05). The relative abundance of Steroidobacter was significantly higher, while the abundances of Gaiella and Lysobacter were significantly lower in the bean rhizosphere under the 2:2 rotational strip intercropping treatment compared to ControlB. The relative abundances of Streptomyces and Gaiella were higher in Bean1:1 than in Bean2:2 (p < 0.05) (Figure S2a). Compared to the celery monoculture treatment, celery intercropping led to higher relative abundances of Clostridium_sensu_stricto_1 and Flavobacterium, and lower relative abundances of Rhizobium, Streptomyces, Burkholderia-Paraburkholderia, Pseudomonas and Pseudarthrobacter (p < 0.05). Furthermore, Celery1:1 exhibited higher relative abundances of Haliangium and Streptomyces compared to Celery2:2 (p < 0.05). Meanwhile, compared with the celery monoculture, the relative abundances of Gaiella and Lysobacter were significantly higher, while the relative abundances of RB41 were significantly lower in rhizosphere of celery in the 1:1 rotational strip intercropping (p < 0.05). Additionally, the relative abundance of Sphingomonas was significantly higher in the rhizosphere of celery in the 2:2 rotational strip intercropping (p < 0.05) (Figure S2b).
3.6. Fungi Community Composition of in Rotational Strip Intercropping
In total, 10 fungi phyla were observed across all of the bean samples. Ascomycota, Basidiomycota and Mortierellomycota, accounted for more than 98% of the sequences, and Olpidiomycota was a minor phyla with a relatively lower abundance (Figure S1b). Compared with the bean monoculture treatment, Bean1:1 and Bean2:2 had higher relative abundances of Ascomycota and Olpidiomycota, and had lower relative abundances of Basidiomycota. In addition, the relative abundance of Mortierellomycota was higher in Bean1:1 than in Bean2:2 (Figure 5b). Eleven fungi phyla were observed across all of the celery samples. The dominant phyla were Ascomycota, Mortierellomycota and Basidiomycota, and these phyla accounted for more than 99% of the fungal sequences (Figure S1b). Compared with celery monoculture treatment, Celery1:1 and Celery2:2 had higher relative abundances of Mortierellomycota (Figure 5b).
Taxonomical classification at the genus level revealed that more than 216 fungal genera were detected across all of the bean treatments. Compared with the bean monoculture treatment, Bean1:1 and Bean2:2 had higher relative abundances of Lophotrichus, Wardomyces, Plectosphaerella and Olpidium. They also had lower relative abundances of Solicoccozyma, Tausonia and Cephaliophora. Moreover, the relative abundances of Mortierella, Fusarium and Aphanoascus were higher in Bean1:1 than in the bean monoculture. Additionally, the relative abundances of Acaulium, Alternaria, Sodiomyces, Acremonium and Thelebolus were higher in Bean2:2 than in the bean monoculture. In addition, the relative abundances of Lophotrichus, Mortierella and Fusarium were higher in Bean1:1 compared to Bean2:2. Conversely, the relative abundances of Acaulium, Alternaria and Acremonium were lower in Bean1:1 than Bean2:2 (Figure S3a).
In the celery treatments, more than 241 bacterial genera were detected. Compared with the celery monoculture treatment, Celery1:1 and Celery2:2 had higher relative abundances of Mortierella, Alternaria and Idriella, and lower relative abundances of Acaulium, Botryotrichum, Plectosphaerella, Thelebolus and Penicillium. Moreover, the relative abundances of Lophotrichus and Tausonia were higher, while the relative abundance of Cephaliophora was lower in Celery1:1 than in the celery monoculture. The relative abundances of Sodiomyces, Chrysosporium and Scutellinia were higher, while the relative abundance of Acremonium was lower in Celery2:2 compared to the celery monoculture. Additionally, the Celery1:1 treatment had higher relative abundances of Mortierella, Alternaria, Tausonia, Acremonium and Idriella, while the relative abundances of Acaulium, Botryotrichum, Sodiomyces, Chrysosporium and Scutellinia were lower compared to Celery2:2 (Figure S3b).
3.7. Correlation Analysis of Sensitive OTUs with Soil Nutrients and Yield
The indicator species analysis and likelihood ratio test identified 66 and 98 sensitive OTUs of the bacteria found in the bean and celery soils, respectively, within the rotational strip intercropping system using beans and celery (Figure S4a,b).
In the 1:1 rotational strip intercropping, we found eight OTUs and sixteen OTUs that were enriched in bean and celery, respectively (Figure S5a,b). In the bacterial-sensitive OTUs in the bean soils, Methylobacillus sp. OTU4257 was significantly positively correlated with NO3−-N, AP, AK and yield. Achromobacter sp. OTU5462, Luteibacter sp. OTU1712 and Stenotrophomonas sp. OTU6012 were significantly positively correlated with AP. Achromobacter sp. 5462 and Flavobacterium sp. OTU5429 was significantly positively correlated with NO3−-N, AP and yield, but Haliangium sp. OTU3576 and Gaiella sp. OTU4143 were significantly negative correlated with NH4+-N (Figure S6a). In the bacterial-sensitive OTUs of the celery, Bryobacter sp. OTU4361 was significantly positively correlated with NO3−-N and AP. Flavisolibacter sp. OTU5084 was significantly positively correlated with NO3−-N and yield. Fluviicola sp. OTU5254 was significantly positively correlated with AK. Sphingopyxis sp. OTU3971 and Flavobacterium sp. OTU5429 were significantly positively correlated with AP and yield (Figure S6b).
In the 2:2 rotational strip intercropping, we found 28 OTUs and 22 OTUs, respectively, that were enriched in the bean and celery soils (Figure S5a,b). In bacterial-sensitive OTUs in the bean soils, Saccharomonospora sp. OTU4006, Rhodococcus sp. OTU4867, Asticcacaulis sp. OTU203, Caulobacter sp. 3018, Brevundimonas sp. OTU654 and Massilia sp. OTU2063 were significantly positively correlated with NH4+-N. Bryobacter sp. OTU5786, Agromyces sp. OTU3978, Archangium sp. OTU5416 and Fluviicola sp. OTU5148 were significantly positively correlated with AK. Steroidobacter sp. OTU5723 was significantly positively correlated with AK and yield. Rhizobium sp. OTU5934 was significantly positively correlated with NH4+-N, AK and yield (Figure S6a). In the bacterial-sensitive OTUs in the celery soil, Stenotrophobacter sp. OTU4138, Streptosporangium sp. OTU4352 and Escherichia-Shigella sp. OTU5167 were significantly positively correlated with AP. Flavobacterium sp. OTU4700 was significantly positively correlated with yield. Nocardioides sp. OTU4929, Flavobacterium sp. OTU4173, Gemmatimonas sp. OTU4002, Pseudoxanthomonas sp. OTU5878 and Nocardioides sp. OTU4929 were significantly positively correlated with AP and yield. Sphingomonas sp. OTU4339 was significantly positively correlated with NH4+-N, AP and yield (Figure S6b).
The indicator species analysis and likelihood ratio test, respectively, identified 33 and 46 sensitive OTUs in the fungi found in the bean and celery soils under the rotational strip intercropping system (Figure S4c,d).
In the rotational strip intercropping system between bean and celery in the 1:1 ratio, six OTUs were enriched in rhizosphere of the beans, and eleven OTUs were enriched in the rhizosphere of celery (Figure 6a,b). In the fungal-sensitive OTUs in the bean soils, Fusarium sp. OTU174 was significantly positively correlated with NO3−-N and AP. Lophotrichus sp. OTU1527 was significantly positively correlated with AP and yield. Ilyonectria sp. OTU52 and Preussia sp. OTU97 were significantly positively correlated with NO3−-N, AP and yield. Stachybotrys sp. OTU122, Acaulium sp. OTU20 and Podospora sp. OTU111 were significantly negative correlated with NH4+-N (Figure 7a). In fungal sensitive OTUs of celery, Alternaria sp. OTU12, Chaetomium sp. OTU70, Idriella sp. OTU19, Mortierella sp. OTU6 and Pluteus sp. OTU390 were significantly positively correlated with NO3−-N. Podospora sp. OTU111 and Tetracladium sp. OTU160 were significantly positively correlated with AK. Arthrographis sp. OTU88 and Periconia sp. OTU297 were significantly positively correlated with NO3−-N and yield. Retroconis sp. OUT 68 was significantly positively correlated with AP and yield. Penicillium sp. OTU233 was significantly positively correlated with NH4+-N, NO3−-N and yield (Figure 7b).
In the rotational strip intercropping system between bean and celery in the 2:2 ratio, 14 OTUs were enriched in rhizosphere of bean, and 15 OTUs were enriched in the rhizosphere of celery (Figure 6a,b). In fungal-sensitive OTUs in the bean soils, Pseudotaeniolina sp. OTU210, Purpureocillium sp. OTU443 and Alternaria sp. OTU12 were significantly positively correlated with AK (Figure 7a). Penicillium sp.162 and Acaulium sp. OTU4 were significantly positively correlated with NH4+-N. Wardomyces sp. OTU8 was significantly positively correlated with AK and yield. In the fungal-sensitive OTUs in the celery soil, Gymnascella sp. OTU211, Chrysosporium sp. OTU21 and OTU1561 and Scutellinia sp. OTU30 were significantly positively correlated with AP. Pithoascus sp. OTU279, Acaulium sp. OTU509, Duddingtonia sp. OTU332 and Sodiomyces sp. OTU18 were significantly negatively correlated with AK. Stachybotrys sp. OTU122, Sarocladium sp. OTU465, Gymnoascus sp. OTU139 and Schizothecium sp. OTU248 were significantly positively correlated with AP and yield (Figure 7b).
4. Discussion
Crop rotation and intercropping are common ways to increase crop yield in agricultural cultivation [41,42]. Rotational strip intercropping has the dual advantages of intercropping and rotation cropping, and is more widely used in agricultural production. It was found that rotational strip intercropping led to an increase in the crop yields of both beans and celery compared to monoculture practices (Figure 2), which was corroborated by the findings of Zou et al. [16]. This supports our first hypothesis. However, the yield from the 1:1 rotational strip intercropping was higher than that from the 2:2 rotational strip intercropping. Additionally, compared with the 2:2 rotational strip intercropping, the photosynthesis of the beans was stronger, while that of the celery was weaker in the 1:1 rotational strip intercropping system (Table S1). Therefore, the 1:1 rotational strip bean and celery intercropping pattern is feasible for facility-based agricultural production.
The relationship between plant diversity and soil chemical properties is of great significance in understanding the function of agricultural ecosystem [43]. Soil chemical properties can be used to evaluate soil heath, and the growth and development of crops will adapt in response to changes in the soil chemical properties [44]. Similarly, variations in surface vegetation can also lead to changes in the soil chemical properties. In this study, 1:1 rotation strip intercropping significantly increased the contents of AP and NO3−-N in the bean rhizosphere, as well as those of NO3−-N and AK in the celery rhizosphere (Figure 3), which was consistent with previous research’s results [16]. Further investigations can explore the effects of reducing fertilizer input on crops in rotational strip bean and celery intercropping. Diverse planting patterns create complex relationships within the plant–soil–microbial complex [45], and Nitrogen, which is an essential element for plant growth and development, plays a crucial role in these interactions. Reducing nitrogen fertilizer application can reduce minimize nitrogen loss, enhance nitrogen recovery efficiency, up-regulate the abundance of nitrogen cycling-related genes in soil, and strengthen the activity of nitrogen conversion cycles [46,47,48]; in addition, nutrient optimization may be associated with selective nutrient uptake by crops or microbial driven nutrient morphological transformations [49]. Further research on nutrient optimization mechanisms is essential to improve nutrient management in diversified cropping systems.
Plant diversity research focuses on interspecific interactions and their legacy effects on soil [50,51,52]. In this study, rotational strip intercropping of bean and celery altered the rhizosphere microbial community composition and diversity of bean and celery (Figure 4), these results were consistent with previous observations, supporting the second hypothesis in this study [53,54]. These results suggest that fungal and bacterial communities play different roles in rotational strip intercropping. Rotational strip intercropping altered the abundance of microorganisms in soil bacterial and fungal communities. In this study, rotational strip intercropping improved the relative abundances of Aeromicrobium, Rhizobium, Clostridium_sensu_stricto_1 and Lophotrichus in the bean rhizosphere, and the relative abundances of Flavobacterium, Clostridium_sensu_stricto_1, Mortierella and Alternaria in the celery rhizosphere (Figure S2). Previous studies have found that Aeromicrobium, Clostridium_sensu_stricto_1, Flavobacterium, Lophotrichus and Mortierella spp. participate in lignocellulose degradation, releasing nutrients that can be absorbed by plants and promoting plant growth, and some of these microbes also inhibit plant diseases [32,55,56,57,58,59]. Rotational strip intercropping increased the relative abundance of Rhizobium in the bean rhizosphere (Figure S2). Legumes and Rhizobium establish a symbiotic relationship to enhance nitrogen fixation, which helps to improve soil fertility [60]. Additionally, rotational strip intercropping decreased the relative abundance of potential pathogenic microorganisms like Botryotrichum in the celery rhizosphere (Figure S2) [61]. Compared with the 2:2 rotational strip intercropping system, the relative abundances of bacteria such as Gaiella and Streptomyces, and fungi such as Mortierella, Fusarium and Lophotrichus, were increased in the bean rhizosphere, and the relative abundances of the bacteria Haliangium and Streptomyces and the fungi Mortierella, Alternaria, Tausonia, Acremonium and Idriella were increased in the celery rhizosphere following the 1:1 rotational strip intercropping (Figures S2, S3 and S5). Some microorganisms increased the decomposition effect on organic matter under the 1:1 rotational strip intercropping system. Gaiella is commonly involved in the decomposition of organic matter, breaking down complex organic matter into simpler compounds. This facilitates the circulation of organic matter and the release of nutrients from the soil, and increases nutrient availability and assimilation to promote plant growth [62]. Haliangium is considered to be an important nitrogen-fixing microorganism [63], which has a potential role in denitrification and phosphorus enhancement [64]. Additionally, Streptomyces is known for their antagonistic abilities and ability to inhibit a variety of plant diseases [65,66]. Therefore, we suggest that the 1:1 rotational strip bean and celery intercropping alter specific rhizosphere microbial groups, and these decomposition-related microorganisms release nutrients from plant litter. At the same time, the input of exogenous substrates such as plant litter may stimulate the activity of specific microbial assemblages, thus decomposing soil organic matter and releasing nutrients that become available to plants in the rotational strip intercropping system.
In agroecosystems, microbial communities play a crucial role in soil nutrient cycling under diverse cropping patterns, and soil microbes as decomposers provide nutrients that can be absorbed by plants [67,68]. In the rotational strip bean and celery intercropping, certain microbial taxa were stimulated, which were significantly correlated with soil nutrient content and yield, and these results support our third hypothesis. We found that the yield and soil nutrient content of the 1:1 rotational strip intercropping were higher than the 2:2 rotational strip intercropping, which may be related to the presence of some beneficial microorganisms. In the 1:1 rotational strip intercropping, some microorganisms were enriched in the rhizosphere of the beans and the rhizosphere of the celery, and there was a significantly positive correlation between these microorganisms and yield and nutrients. Stenotrophomonas were significantly positively correlated with AP, and Flavobacterium was significantly positively correlated with AP and yield in the bean and/or celery rhizosphere (Figure S6). Stenotrophomonas may convey benefits to plant growth through multiple mechanisms, such as the production of siderophores and IAA, the enhancement of soil alkaline phosphatase activity and phosphate solubilization, and the promotion of N, P and K uptake by plants [69,70,71]. It is possible to provide essential nutrients for plant growth through mechanisms such as dissolving inorganic phosphorus from non-mobile quartzes in the soil or fixing nitrogen from the atmosphere [72]. Fusarium and Preussia were significantly positively correlated with NO3−-N, AP and yield in both the beans and celery (Figure 7). Fusarium and Preussia are known for their ability to degrade recalcitrant compounds [73,74]. They may degrade lignocellulose by breaking the glycosidic bond in the cell wall of litter in the rotational strip bean and celery intercropping system, thereby increasing the nutrients in the soil and the yield [75]. However, further experiments are necessary, and isolating and testing the litter-decomposing abilities of these stimulated microbial taxa are necessary to improve our understanding of the role of microbial decomposer community in the rotational strip intercropping.
5. Conclusions
We found that rotational strip bean and celery intercropping altered the diversity and compositions of decomposition-related microorganisms, and increased the relative abundances of certain taxa that serve as potential decomposers. Moreover, the yield from the 1:1 rotational strip intercropping was more than that from the 2:2 rotational strip intercropping. The 1:1 rotational strip intercropping also increased soil nitrogen and phosphorus contents. The 1:1 rotational strip bean and celery intercropping pattern is feasible in facility-based agricultural production.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae10050432/s1, Figure S1: Taxonomic profiles of bacterial (a) and fungal (b) communities at phylum level. Bacterial and fungal phyla with relative abundances lower than 1% were summarized with ‘other’; Figure S2: Bacterial genus with average relative abundances > 0.5% altered by rotational strip intercropping of bean (a) and celery (b). Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test); Figure S3: Fungal genus with average relative abundances > 0.5% altered by rotational strip intercropping of bean (a) and celery (b). Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test); Figure S4: Defining cropping bacterial and fungal sensitive OTUs in bean (a,b) and celery (c,d) samples; Table S1: Effects of the rotational strip bean and celery intercropping on Pn and Illuminance. Figure S5: Bacterial taxa sensitive OTUs to the rotational strip bean (a) and celery (b) intercropping by both indicator species analysis and likelihood ratio test. Figure S6: Spearman correlation heat map of bacterial sensitive OTUs of bean (a) and celery (b) soil samples by rotational strip intercropping.
Author Contributions
S.L.: Data curation, Investigation, Formal analysis, Writing—original draft, Writing—review and editing. T.Y.: Investigation, Software. M.I.: Investigation, Software. F.W.: Conceptualization, Funding acquisition, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program (2018YFD1000800).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tilman, D.; Reich, P.B.; Knops, J.M.H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; De Mazancourt, C.; Duffy, E. Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecol. Lett. 2013, 16, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Bowles, T.M.; Mooshammer, M.; Socolar, Y.; Calderón, F.; Cavigelli, M.A.; Culman, S.W.; Deen, W.; Drury, C.F.; Garcia, A.G.Y.; Gaudin, A.C.M.; et al. Long-Term Evidence Shows that Crop-Rotation Diversification Increases Agricultural Resilience to Adverse Growing Conditions in North America. One Earth 2020, 2, 284–293. [Google Scholar] [CrossRef]
- Garland, G.; Edlinger, A.; Banerjee, S.; Degrune, F.; Garcia-Palacios, P.; Pescador, D.S.; Herzog, C.; Romdhane, S.; Saghai, A.; Spor, A.; et al. Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nat. Food 2021, 2, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Chadfield, V.G.A.; Hartley, S.E.; Redeker, K.R. Associational resistance through intercropping reduces yield losses to soil-borne pests and diseases. New Phytol. 2022, 235, 2393–2405. [Google Scholar] [CrossRef]
- Wahbi, S.; Prin, Y.; Thioulouse, J.; Sanguin, H.; Baudoin, E.; Maghraoui, T.; Oufdou, K.; Le Roux, C.; Galiana, A.; Hafidi, M.; et al. Impact of Wheat/Faba Bean Mixed Cropping or Rotation Systems on Soil Microbial Functionalities. Front. Plant Sci. 2016, 7, 1364. [Google Scholar] [CrossRef]
- Luo, C.; Lv, J.; Guo, Z.; Dong, Y. Intercropping of Faba Bean with Wheat Under Different Nitrogen Levels Reduces Faba Bean Rust and Consequent Yield Loss. Plant Dis. 2022, 106, 2370–2379. [Google Scholar] [CrossRef]
- Ma, H.; Yu, X.; Yu, Q.; Wu, H.; Zhang, H.; Pang, J.; Gao, Y. Maize/alfalfa intercropping enhances yield and phosphorus acquisition. Field Crops Res. 2023, 303, 109136. [Google Scholar] [CrossRef]
- Mesfin, S.; Gebresamuel, G.; Haile, M.; Zenebe, A. Potentials of legumes rotation on yield and nitrogen uptake of subsequent wheat crop in northern Ethiopia. Heliyon 2023, 9, e16126. [Google Scholar] [CrossRef]
- Yu, T.; Nie, J.; Zang, H.; Zeng, Z.; Yang, Y. Peanut-based Rotation Stabilized Diazotrophic Communities and Increased Subsequent Wheat Yield. Microb. Ecol. 2023, 86, 2447–2460. [Google Scholar] [CrossRef]
- Lu, J.; Liu, Y.; Zou, X.; Zhang, X.; Yu, X.; Wang, Y.; Si, T. Rotational strip peanut/cotton intercropping improves agricultural production through modulating plant growth, root exudates, and soil microbial communities. Agric. Ecosyst. Environ. 2024, 359, 108767. [Google Scholar] [CrossRef]
- Zou, X.X.; Shi, P.X.; Zhang, C.J.; Si, T.; Wang, Y.F.; Zhang, X.J.; Yu, X.N.; Wang, H.X.; Wang, M.L. Rotational strip intercropping of maize and peanuts has multiple benefits for agricultural production in the northern agropastoral ecotone region of China. Eur. J. Agron. 2021, 129, 126304. [Google Scholar] [CrossRef]
- Chi, B.; Zhang, Y.; Zhang, D.; Zhang, X.; Dai, J.; Dong, H. Wide-strip intercropping of cotton and peanut combined with strip rotation increases crop productivity and economic returns. Field Crops Res. 2019, 243, 107617. [Google Scholar] [CrossRef]
- Han, Y.; Dong, Q.; Zhang, K.; Sha, D.; Jiang, C.; Yang, X.; Liu, X.; Zhang, H.; Wang, X.; Guo, F.; et al. Maize-peanut rotational strip intercropping improves peanut growth and soil properties by optimizing microbial community diversity. Peerj 2022, 10, e13777. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Chi, B.; He, N.; Zhang, D.; Dai, J.; Zhang, Y.; Dong, H. Cotton-Based Rotation, Intercropping, and Alternate Intercropping Increase Yields by Improving Root-Shoot Relations. Agronomy 2023, 13, 413. [Google Scholar] [CrossRef]
- Zou, X.; Liu, Y.; Huang, M.; Li, F.; Si, T.; Wang, Y.; Yu, X.; Zhang, X.; Wang, H.; Shi, P. Rotational strip intercropping of maize and peanut enhances productivity by improving crop photosynthetic production and optimizing soil nutrients and bacterial communities. Field Crops Res. 2023, 291, 108770. [Google Scholar] [CrossRef]
- Sudini, H.; Liles, M.R.; Arias, C.R.; Bowen, K.L.; Huettel, R.N. Exploring Soil Bacterial Communities in Different Peanut-Cropping Sequences Using Multiple Molecular Approaches. Phytopathology 2011, 101, 819–827. [Google Scholar] [CrossRef]
- Guo, F.; Wang, M.; Si, T.; Wang, Y.; Zhao, H.; Zhang, X.; Yu, X.; Wan, S.; Zou, X. Maize-peanut intercropping led to an optimization of soil from the perspective of soil microorganism. Arch. Agron. Soil Sci. 2021, 67, 1986–1999. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, J.; Wu, F. Soil microbial communities in cucumber monoculture and rotation systems and their feedback effects on cucumber seedling growth. Plant Soil 2017, 415, 507–520. [Google Scholar] [CrossRef]
- Liu, X.; Ren, X.; Tang, S.; Zhang, Z.; Huang, Y.; Sun, Y.; Gao, Z.; Ma, Z. Effects of Broccoli Rotation on Soil Microbial Community Structure and Physicochemical Properties in Continuous Melon Cropping. Agronomy 2023, 13, 2066. [Google Scholar] [CrossRef]
- Li, N.; Gao, D.; Zhou, X.; Chen, S.; Li, C.; Wu, F. Intercropping with Potato-Onion Enhanced the Soil Microbial Diversity of Tomato. Microorganisms 2020, 8, 834. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Shi, Y.; Wu, F.; Pan, K.; Zhou, X. Intercropping of wheat changed cucumber rhizosphere bacterial community composition and inhibited cucumber Fusarium wilt disease. Sci. Agric. 2020, 77, e20190005. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Dobies, T.; Cesarz, S.; Hobbie, S.E.; Meyer, R.J.; Worm, K.; Reich, P.B. Plant diversity effects on soil food webs are stronger than those of elevated CO2 and N deposition in a long-term grassland experiment. Proc. Natl. Acad. Sci. USA 2013, 110, 6889–6894. [Google Scholar] [CrossRef] [PubMed]
- Steinauer, K.; Chatzinotas, A.; Eisenhauer, N. Root exudate cocktails: The link between plant diversity and soil microorganisms? Ecol. Evol. 2016, 6, 7387–7396. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Martiny JB, H.; Eisen, J.A.; Penn, K.; Allison, S.D.; Horner-Devine, M.C. Drivers of bacterial β-diversity depend on spatial scale. Proc. Natl. Acad. Sci. USA 2011, 108, 7850–7854. [Google Scholar] [CrossRef] [PubMed]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Xu, Y.; Zhang, J.; Hao, X.; Lu, Y. Core Microbiota in Agricultural Soils and Their Potential Associations with Nutrient Cycling. Msystems 2019, 4, 10–1128. [Google Scholar] [CrossRef]
- Gao, H.; Tian, G.; Rahman, M.K.U.; Wu, F. Cover Crop Species Composition Alters the Soil Bacterial Community in a Continuous Pepper Cropping System. Front. Microbiol. 2022, 12, 789034. [Google Scholar] [CrossRef]
- Hu, H.Y.; Li, H.; Hao, M.M.; Ren, Y.N.; Zhang, M.K.; Liu, R.Y.; Zhang, Y.; Li, G.; Chen, J.S.; Ning, T.Y.; et al. Nitrogen fixation and crop productivity enhancements co-driven by intercrop root exudates and key rhizosphere bacteria. J. Appl. Ecol. 2021, 58, 2243–2255. [Google Scholar] [CrossRef]
- Wongkiew, S.; Polprasert, C.; Noophan, P.; Koottatep, T.; Kanokkantapong, V.; Surendra, K.C.; Khanal, S.K. Effects of vermicompost leachate on nitrogen, phosphorus, and microbiome in a food waste bioponic system. J. Environ. Manag. 2023, 339, 117860. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; Dewaak, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 2010, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.G.; Zhang, J.Y.; Rahman, M.K.U.; Gao, D.M.; Wei, Z.; Wu, F.Z. p-Coumaric can alter the composition of cucumber rhizosphere microbial communities and induce negative plant-microbial interactions. Biol. Fertil. Soils 2018, 54, 363–372. [Google Scholar] [CrossRef]
- Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Hartman, K.; van der Heijden, M.G.A.; Wittwer, R.A.; Banerjee, S.; Walser, J.C.; Schlaeppi, K. Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 2018, 6, 1–14. [Google Scholar]
- Tilman, D. Benefits of intensive agricultural intercropping. Nat. Plants 2020, 6, 604–605. [Google Scholar] [CrossRef]
- Yang, X.; Xiong, J.; Du, T.; Ju, X.; Gan, Y.; Li, S.; Xia, L.; Shen, Y.; Pacenka, S.; Steenhuis, T.S.; et al. Diversifying crop rotation increases food production, reduces net greenhouse gas emissions and improves soil health. Nat. Commun. 2024, 15, 198. [Google Scholar] [CrossRef] [PubMed]
- Furey, G.N.; Tilman, D. Plant biodiversity and the regeneration of soil fertility. Proc. Natl. Acad. Sci. USA 2021, 118, e2111321118. [Google Scholar] [CrossRef]
- Abidemi, A.A.; Ewulo, B.S.; Aiyelari, O.P.; Hu, J. Effects of NPK Fertilizer and Vine Care on Soil Chemical Properties and Cucumber (Cucumis sativus L.) Growth and Yield Parameters. Int. J. Plant Soil Sci. 2021, 33, 136–151. [Google Scholar]
- Cappelli, S.L.; Domeignoz-Horta, L.A.; Loaiza, V.; Laine, A.L. Plant biodiversity promotes sustainable agriculture directly and via belowground effects. Trends Plant Sci. 2022, 27, 674–687. [Google Scholar] [CrossRef]
- Zhang, X.; Mauzerall, D.L.; Davidson, E.A.; Kanter, D.R.; Cai, R. The Economic and Environmental Consequences of Implementing Nitrogen-Efficient Technologies and Management Practices in Agriculture. J. Environ. Qual. 2015, 44, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Kermah, M.; Franke, A.C.; Adjei-Nsiah, S.; Ahiabor, B.D.K.; Abaidoo, R.C.; Giller, K.E. Maize-grain legume intercropping for enhanced resource use efficiency and crop productivity in the Guinea savanna of northern Ghana. Field Crops Res. 2017, 213, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, S.; Huang, Z.; Huang, T.; Tang, X.; He, L.; Li, Z.; Xiong, J.; Zhong, R.; Jiang, J.; et al. Effects of Intercropping and Nitrogen Application on Soil Fertility and Microbial Communities in Peanut Rhizosphere Soil. Agronomy 2024, 14, 635. [Google Scholar] [CrossRef]
- Jiao, N.; Wang, J.; Ma, C.; Zhang, C.; Guo, D.; Zhang, F.; Jensen, E.S. The importance of aboveground and belowground interspecific interactions in determining crop growth and advantages of peanut/maize intercropping. Crop J. 2021, 9, 1460–1469. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant-soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Nannipieri, P.; Hannula, S.E.; Pietramellara, G.; Schloter, M.; Sizmur, T.; Pathan, S.I. Legacy effects of rhizodeposits on soil microbiomes: A perspective. Soil Biol. Biochem. 2023, 184, 109107. [Google Scholar] [CrossRef]
- Yan, X.; Niu, J.; Li, Y.; Sun, S.; Li, X.; Jin, K. Species-specific soil legacy effects on ecosystem multifunctionality via regulating plant overyielding. Land Degrad. Dev. 2023, 34, 1235–1245. [Google Scholar] [CrossRef]
- Li, L.; Tilman, D.; Lambers, H.; Zhang, F.S. Plant diversity and overyielding: Insights from belowground facilitation of intercropping in agriculture. New Phytol. 2014, 203, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, Z.; Liu, J.; Li, X.; Wang, X.; Dai, C.; Zhang, T.; Carrion, V.J.; Wei, Z.; Cao, F.; et al. Crop rotation and native microbiome inoculation restore soil capacity to suppress a root disease. Nat. Commun. 2023, 14, 8126. [Google Scholar] [PubMed]
- Rice, A.V.; Currah, R.S. Two new species of Pseudogymnoascus with Geomyces anamorphs and their phylogenetic relationship with Gymnostellatospora. Mycologia 2006, 98, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Si, H.; He, J.; Fan, L.; Li, L. The shifts of maize soil microbial community and networks are related to soil properties under different organic fertilizers. Rhizosphere 2021, 19, 100388. [Google Scholar] [CrossRef]
- Qin, J.; Bian, C.; Duan, S.; Wang, W.; Li, G.; Jin, L. Effects of different rotation cropping systems on potato yield, rhizosphere microbial community and soil biochemical properties. Front. Plant Sci. 2022, 13, 999730. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, L. Effects of additives on the co-composting of forest residues with cattle manure. Bioresour. Technol. 2023, 368, 128384. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Shen, S.; Wan, C.; Wang, S.; Yang, F.; Zhang, K.; Gao, W. Organic fertilizer substitution over six years improves the productivity of garlic, bacterial diversity, and microbial communities network complexity. Appl. Soil Ecol. 2023, 182, 104718. [Google Scholar]
- Yang, Y.; Xu, N.; Zhang, Z.; Lei, C.; Chen, B.; Qin, G.; Qiu, D.; Lu, T.; Qian, H. Deciphering Microbial Community and Nitrogen Fixation in the Legume Rhizosphere. J. Agric. Food Chem. 2024, 72, 5659–5670. [Google Scholar] [CrossRef]
- Tang, C.; Sun, B.; Zeeshan, M.; Li, J.; Zhang, X. Funneliformis mosseae-induced changes of rhizosphere microbial community structure enhance Capsicum annuum L. plant growth and fruit yield. Soil Sci. Soc. Am. J. 2023, 87, 843–855. [Google Scholar] [CrossRef]
- Novello, G.; Gamalero, E.; Bona, E.; Boatti, L.; Mignone, F.; Massa, N.; Cesaro, P.; Lingua, G.; Berta, G. The Rhizosphere Bacterial Microbiota of Vitis vinifera cv. Pinot Noir in an Integrated Pest Management Vineyard. Front. Microbiol. 2017, 8, 1528. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Yamanaka, H.; Xu, Z.X.; Shiratori, Y.; Aono, T.; Amachi, S.; Senoo, K.; Itoh, H. Diazotrophic Anaeromyxobacter Isolates from Soils. Appl. Environ. Microbiol. 2020, 86, e00956-20. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Liu, Y.; Yao, D.; Wang, N.; Ye, X.; Cui, Z.; Wang, H. Phylogenetic diversity of stochasticity-dominated predatory myxobacterial community drives multi-nutrient cycling in typical farmland soils. Sci. Total Environ. 2023, 871, 161680. [Google Scholar] [CrossRef] [PubMed]
- Samac, D.A.; Kinkel, L.L. Suppression of the root-lesion nematode (Pratylenchus penetrans) in alfalfa (Medicago sativa) by Streptomyces spp. Plant Soil 2001, 235, 35–44. [Google Scholar] [CrossRef]
- Law, J.W.F.; Ser, H.L.; Khan, T.M.; Chuah, L.H.; Pusparajah, P.; Chan, K.G.; Goh, B.H.; Lee, L.H. The Potential of Streptomyces as Biocontrol Agents against the Rice Blast Fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 215552. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Bender, S.F.; Van Der Heijden, M.G.A. Soil biota enhance agricultural sustainability by improving crop yield, nutrient uptake and reducing nitrogen leaching losses. J. Appl. Ecol. 2015, 52, 228–239. [Google Scholar] [CrossRef]
- Chen, J.; Qiao, M.; Yang, Y.; Gao, Z.; Yang, Z.; Lin, W. Exogenous Streptomyces spp. benefit naked oat growth under dry farming conditions by modifying rhizosphere bacterial communities. Appl. Soil Ecol. 2023, 189, 104946. [Google Scholar] [CrossRef]
- Rathi, M.; Nandabalan, Y.K. Copper-tolerant rhizosphere bacteria-characterization and assessment of plant growth promoting factors. Environ. Sci. Pollut. Res. 2017, 24, 9723–9733. [Google Scholar] [CrossRef]
- Xiao, C.Q.; Chi, R.A.; He, H.; Zhang, W.X. Characterization of tricalcium phosphate solubilization by Stenotrophomonas maltophilia YC isolated from phosphate mines. J. Cent. South Univ. Technol. 2009, 16, 581–587. [Google Scholar] [CrossRef]
- Oteino, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wang, Z.; Wu, F.; Li, X.; Zhou, X. Litter Mixing Alters Microbial Decomposer Community to Accelerate Tomato Root Litter Decomposition. Microbiol. Spectr. 2022, 10, e00186-22. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Meng, S.; Wang, L.; Zhang, Z. Optimum Fermentation Condition of Soybean Curd Residue and Rice Bran by Preussia aemulans using Solid-State Fermentation Method. Int. J. Biol. 2015, 7, 66–74. [Google Scholar] [CrossRef]
- Ma, A.; Zhuang, X.; Wu, J.; Cui, M.; Lv, D.; Liu, C.; Zhuang, G. Ascomycota Members Dominate Fungal Communities during Straw Residue Decomposition in Arable Soil. PLoS ONE 2013, 8, e66146. [Google Scholar] [CrossRef]
Figure 1.
Diagram of 1:1 and 2:2 rotational strip beans and celery intercropping.

Figure 2.
Effects of rotational strip bean (a) and celery (b) intercropping on yield. ControlB and ControlC, bean monoculture and celery monoculture; Bean1:1 and Celery1:1, rotational strip intercropping of bean and celery in 1:1 ratio; Bean2:2 and Celery2:2, rotational strip intercropping of bean and celery in 2:2 ratio. Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).
Figure 2.
Effects of rotational strip bean (a) and celery (b) intercropping on yield. ControlB and ControlC, bean monoculture and celery monoculture; Bean1:1 and Celery1:1, rotational strip intercropping of bean and celery in 1:1 ratio; Bean2:2 and Celery2:2, rotational strip intercropping of bean and celery in 2:2 ratio. Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).

Figure 3.
Effects of rotational strip bean (a) and celery (b) intercropping on the soil nutrients. Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).
Figure 3.
Effects of rotational strip bean (a) and celery (b) intercropping on the soil nutrients. Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).

Figure 4.
Abundances and diversity of bacterial and fungal communities detected for rotational strip bean (a) and celery (b) intercropping. Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).
Figure 4.
Abundances and diversity of bacterial and fungal communities detected for rotational strip bean (a) and celery (b) intercropping. Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).

Figure 5.
Different bacterial and fungi phyla with average relative abundances > 1% altered by rotational strip intercropping of bean (a) and celery (b). Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).
Figure 5.
Different bacterial and fungi phyla with average relative abundances > 1% altered by rotational strip intercropping of bean (a) and celery (b). Values (mean ± standard error) in the same figure followed by different letters are significantly different (p < 0.05, Tukey’s HSD test).

Figure 6.
Fungal taxa-sensitive OTUs to rotational strip bean (a) and celery (b) intercropping. Solid circles represent sensitive OTU enrichment in the treatment.
Figure 6.
Fungal taxa-sensitive OTUs to rotational strip bean (a) and celery (b) intercropping. Solid circles represent sensitive OTU enrichment in the treatment.

Figure 7.
Spearman correlation heat map of fungal-sensitive OTUs in bean (a) and celery (b) soil samples under rotational strip intercropping. Sensitive OTUs not correlated with soil nutrients or yield were not shown. ***, ** and * denote significance at 0.001, 0.01 and 0.05 probability levels, respectively.
Figure 7.
Spearman correlation heat map of fungal-sensitive OTUs in bean (a) and celery (b) soil samples under rotational strip intercropping. Sensitive OTUs not correlated with soil nutrients or yield were not shown. ***, ** and * denote significance at 0.001, 0.01 and 0.05 probability levels, respectively.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, S.; Yuan, T.; Ibrahim, M.; Wu, F. Rotational Strip Bean and Celery Intercropping Alters the Microbial Community to Improve Crop Yield and Soil Nutrients. Horticulturae 2024, 10, 432. https://doi.org/10.3390/horticulturae10050432
AMA Style
Li S, Yuan T, Ibrahim M, Wu F. Rotational Strip Bean and Celery Intercropping Alters the Microbial Community to Improve Crop Yield and Soil Nutrients. Horticulturae. 2024; 10(5):432. https://doi.org/10.3390/horticulturae10050432
Chicago/Turabian StyleLi, Shuang, Tao Yuan, Musawar Ibrahim, and Fengzhi Wu. 2024. "Rotational Strip Bean and Celery Intercropping Alters the Microbial Community to Improve Crop Yield and Soil Nutrients" Horticulturae 10, no. 5: 432. https://doi.org/10.3390/horticulturae10050432
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.