
Pomegranate Cultivars with Diverse Origins Exhibit Strong Resistance to Anthracnose Fruit Rot Caused by Colletotrichum gloeosporioides, a Major Disease in Southeast United States
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Pomegranate Cultivars
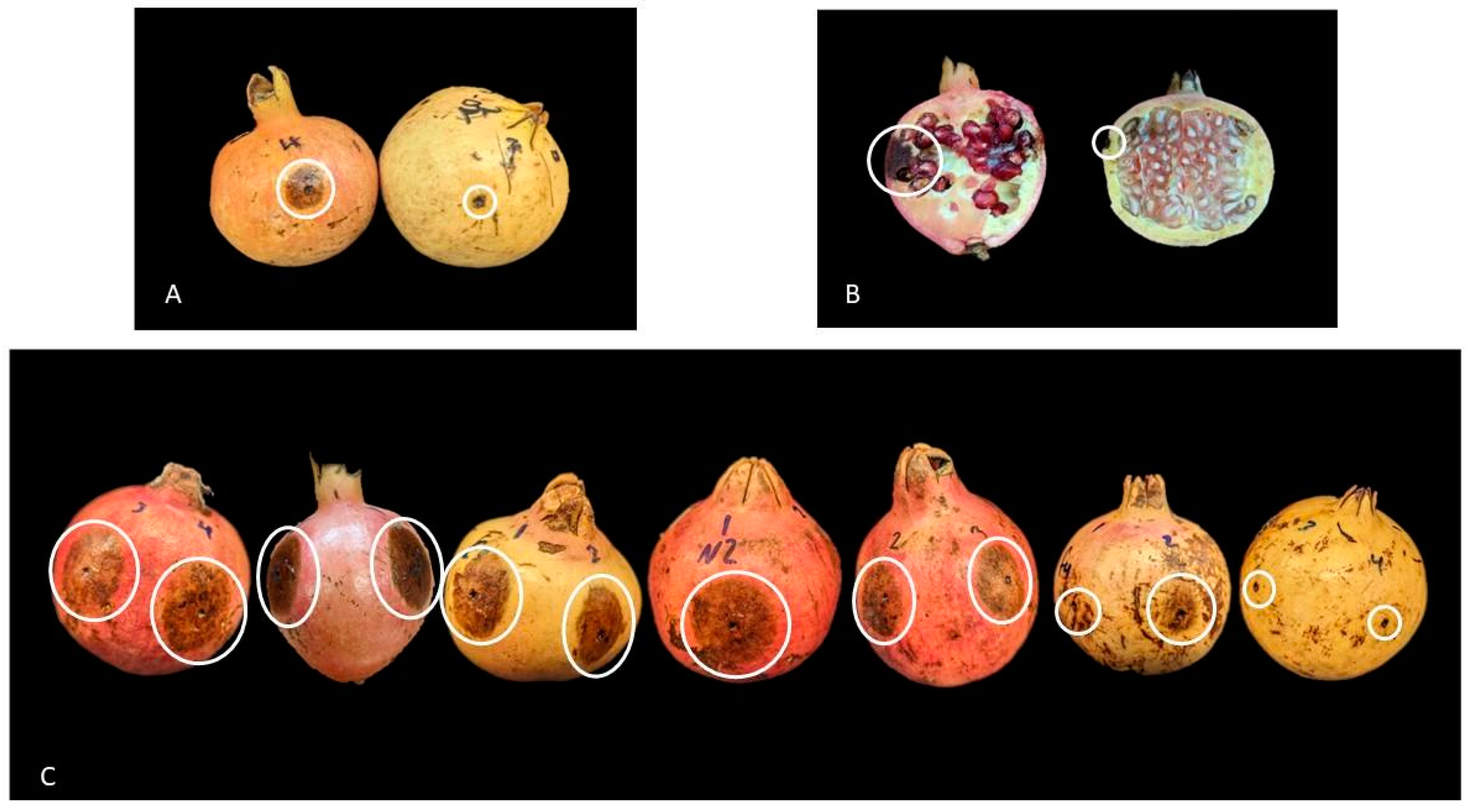
2.2. Evaluation of Fruit Rot Severity under Natural Disease Pressure
2.3. Preparation of Fungal Spore Suspension for Artificial Inoculation
2.4. Inoculation of Fungal Spores to Open Flowers and Young Fruit on Plants in the Field
2.5. Measuring Lesion Size and Disease (Fruit Rot) Development after Artificial Inoculation of In-Planta Fruit the Field
2.6. Ranking of Fruit Rot Resistance Levels
2.7. Re-Isolation of Fungal Pathogen from Inoculated Fruit
2.8. Evaluation of Detached Mature Fruit for Resistance to Fruit Rot
2.9. Statistical Analysis
3. Results
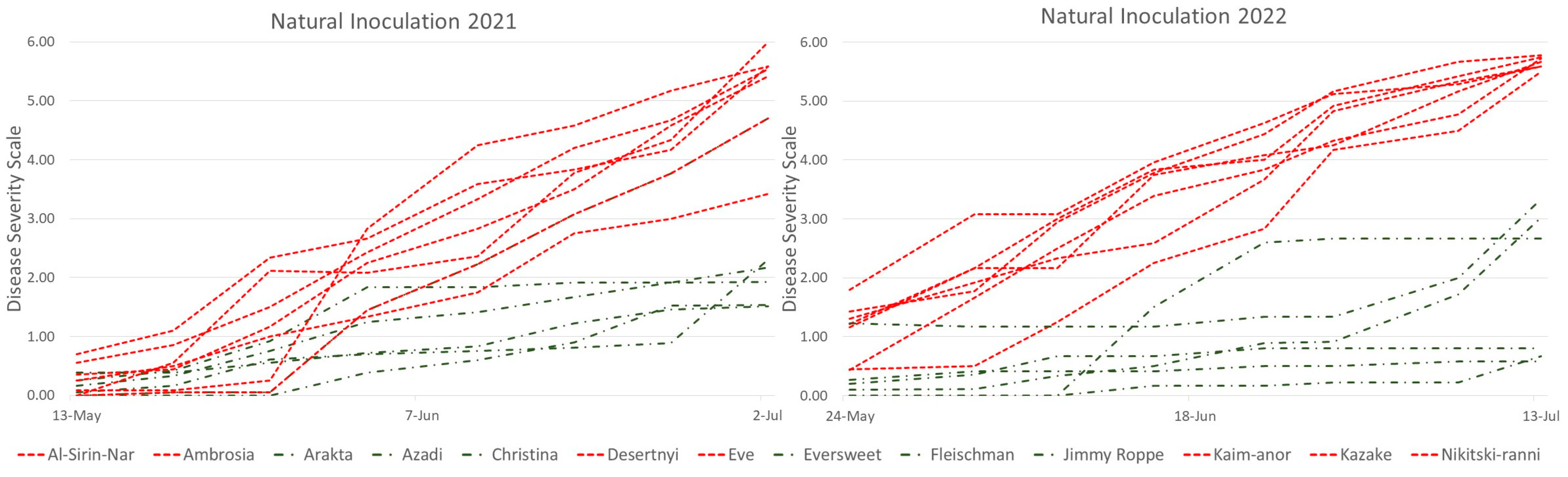
3.1. Fruit Rot under Natural Disease Pressure
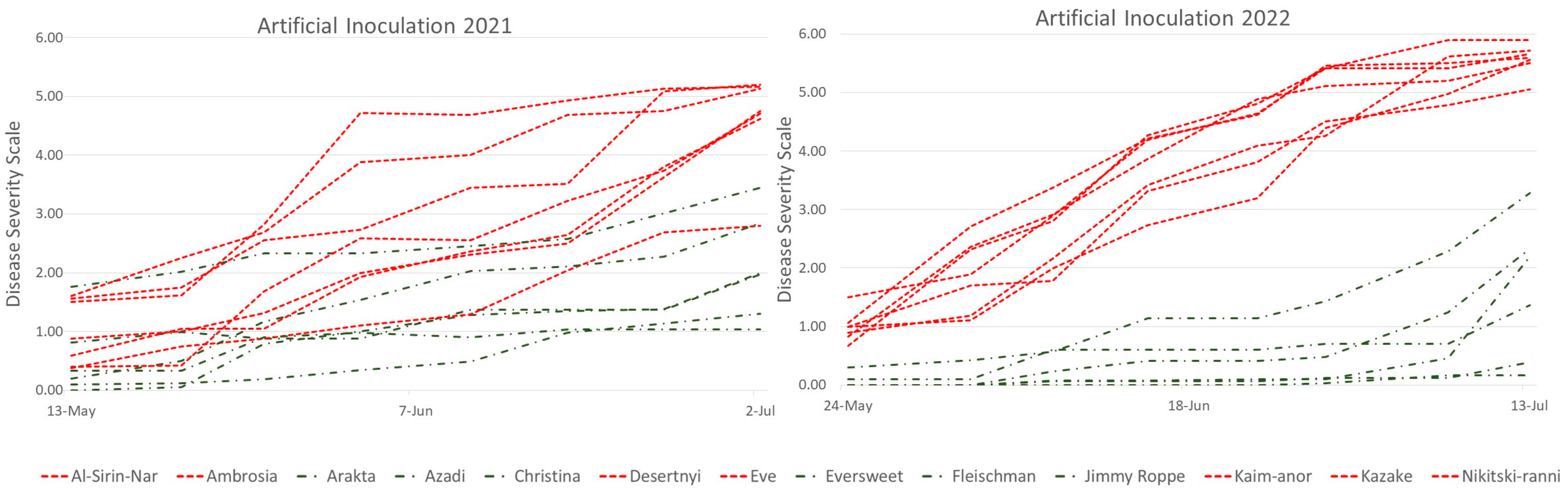
3.2. Fruit Rot after Artificial Inoculation
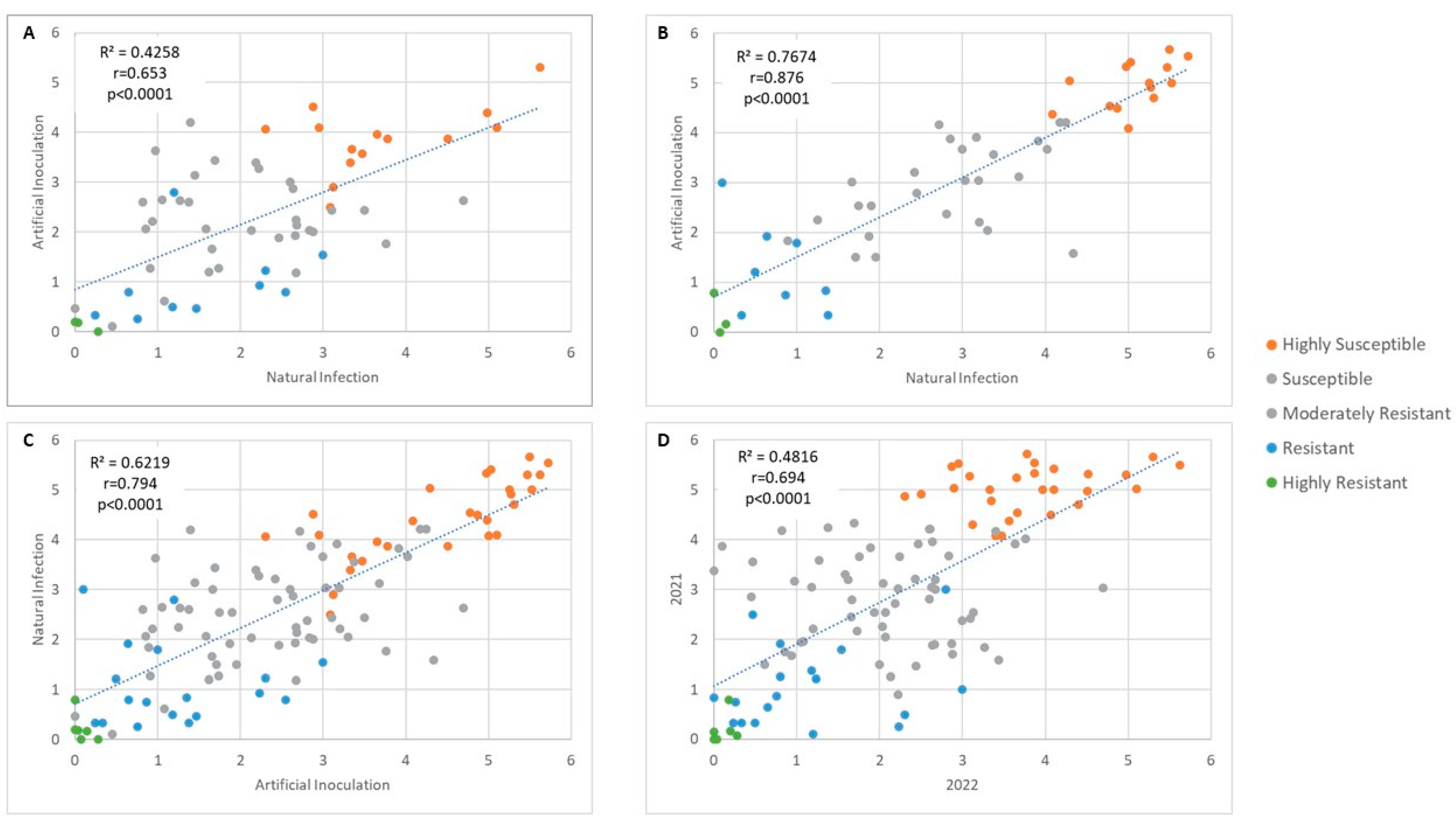
3.3. Ranking of Resistance Level and Correlation of Inoculation Methods and Years
3.4. Detached Fruit Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spragnoli, F.; Shirazi, R.; Shirazi, Z.; Saeidi Ghavi Andam, S.; Djamali, M. Archaeology, history and symbolism. In The Pomegranate: Botany, Production and Uses; Sarkhosh, A., Alimohammad, M.Y., Zamani, Z., Eds.; CABI: Oxfordshire, UK, 2021; pp. 1–14. [Google Scholar]
- Melgarejo-Sánchez, P.; Núñez-Gómez, D.; Martínez-Nicolás, J.J.; Hernández, F.; Legua, P.; Melgarejo, P. Pomegranate variety and pomegranate plant part, relevance from bioactive point of view: A review. Bioresour. Bioprocess 2021, 8, 2. [Google Scholar] [CrossRef]
- Kahramanoglu, I. Trends in pomegranate sector: Production, postharvest handling and marketing. Int. J. Agric. For. Life Sci. 2019, 3, 239–246. [Google Scholar]
- Bartual, J.; Zamudio, M.; Gómez, M. Situation of the production, research and economics of the pomegranate industry in Spain. Acta Hortic. 2015, 1089, 345–349. [Google Scholar] [CrossRef]
- Marty, A.; Castillo, A.; Zoppolo, R. Pomegranate: A growing alternative for fruit production in Uruguay. Acta Hortic. 2015, 1089, 351–355. [Google Scholar] [CrossRef]
- Saroj, P.L.; Kumar, R. Recent advances in pomegranate production in India—A review. Ann. Hortic. 2019, 12, 1–10. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Ávila-Gálvez, M.Á.; Espín, J.C.; González-Sarrías, A. Evidence for health properties of pomegranate juices and extracts beyond nutrition: A critical systematic review of human studies. Trends Food Sci. Technol. 2021, 114, 410–423. [Google Scholar] [CrossRef]
- Asgary, S.; Azadeh, Z.; Dodman, S.; Soleymani, A.; Mirjalili, M.H.; Farzaei, M.H.; Hoseinzadeh-chahkandak, F.; Hozeifi, S.; Jasemi, E.; Keshvari, M.; et al. Pomegranate bioactive compounds and health. In The Pomegranate: Botany, Production and Uses; Sarkhosh, A., Alimohammad, M.Y., Zamani, Z., Eds.; CABI: Oxfordshire, UK, 2021; pp. 504–547. [Google Scholar]
- Howell, A.; D’Souza, D. The pomegranate: Effects on bacteria and viruses that influence human health. Evid. Based Complement Alternat. Med. 2013, 2013, 606212. [Google Scholar] [CrossRef]
- LaRue, J.H. Growing Pomegranates in California; Leaflet 2459. 1980. Available online: http://ucanr.edu/sites/Pomegranates/files/122804.pdf (accessed on 20 March 2023).
- Stover, E.; Mercure, E.W. The pomegranate: A new look at the fruit of paradise. HortScience 2007, 42, 1088–1092. [Google Scholar] [CrossRef]
- USDA National Agricultural Statistics Service, 2017 Census of Agriculture. Available online: www.nass.usda.gov/AgCensus (accessed on 14 March 2023).
- Chater, J.M.; Merhaut, D.J.; Jia, Z.; Mauk, P.A.; Preece, J.E. Fruit quality traits of ten California-grown pomegranate cultivars harvested over three months. Sci. Hortic. 2018, 237, 11–19. [Google Scholar] [CrossRef]
- Hooks, T.; Niu, G.; Masabni, J.; Sun, Y.; Ganjegunte, G. Performance and phytochemical content of 22 pomegranate (Punica granatum) varieties. HortScience 2021, 56, 217–225. [Google Scholar] [CrossRef]
- Castle, W.S.; Baldwin, J.C.; Singh, M. Pomegranate in Florida for commercial enterprises and homeowners. Proc. Fla. State Hortic. Soc. 2011, 124, 33–40. [Google Scholar]
- Deng, Z.; Castle, W.; Vallad, G.E.; Agehara, S.; Thetford, M.; Dıáz-Pérez, J.C. Pomegranate: An emerging fruit crop in southeast United States? Acta Hortic. 2019, 1254, 149–156. [Google Scholar] [CrossRef]
- Xavier, K.; Kc, A.N.; Peres, N.A.; Deng, Z.; Castle, W.; Lovett, W.; Vallad, G.E. Characterization of Colletotrichum species causing anthracnose of pomegranate in the southeastern United States. Plant Dis. 2019, 103, 2771–2780. [Google Scholar] [CrossRef]
- Kc, A.N.; Vallad, G. Monitoring pomegranate pathogens towards developing effective disease management program. Phytopathology 2016, 106, 94. [Google Scholar]
- Xavier, K.; Kc, A.N.; Vallad, G. Diseases of pomegranate (Punica granatum) in Florida: PP349, 10/2019. EDIS 2019, 5. [Google Scholar] [CrossRef]
- Rahimlou, S.; Babaeizad, V.; Sayari, M. First report of fruit spot of pomegranate caused by Colletotrichum gloeosporioides in Iran. J. Plant Pathol. 2014, 96, 605. [Google Scholar]
- Thomidis, T. Fruit rots of pomegranate (cv. Wonderful) in Greece. Australas. Plant Pathol. 2014, 43, 583–588. [Google Scholar] [CrossRef]
- Munhuweyi, K.; Lennox, C.L.; Meitz-Hopkins, J.; Caleb, O.J.; Opara, U.L. Major diseases of pomegranate (Punica granatum L.), their causes and management—A review. Sci. Hortic. 2016, 211, 126–139. [Google Scholar] [CrossRef]
- Uysal, A.; Kurt, Ş. Colletotrichum gloeosporiodes causing anthracnose on pomegranate in Turkey. Australas. Plant Dis. Notes 2018, 13, 19. [Google Scholar] [CrossRef]
- Silva-Cabral, J.R.A.; Batista, L.R.L.; Costa, J.F.d.O.; Ferro, M.M.d.M.; Silva, S.J.C.; Lima, G.S.d.A.; Assunção, I.P. First report of Colletotrichum tropicale causing anthracnose on pomegranate in Brazil. Plant Dis. 2018, 103, 583. [Google Scholar] [CrossRef]
- Patel, D.S. Chemical management of fruit spot of pomegranate caused by Colletotrichum gloeosporioides Penz. and Sacc. Indian Phytopathol. 2009, 62, 252–253. [Google Scholar]
- Dev, D.; Narendrappa, T. In vitro evaluation of fungicides against Colletotrichum gloeosporioides (Penz.) Penz and Sacc. causing anthracnose of pomegranate (Punica granatum L.). J. Appl. Nat. Sci. 2016, 8, 2268–2272. [Google Scholar] [CrossRef]
- Golakiya, B.; Akbari, L.F.; Marakna, N.M. In vitro evaluation of different fungicides against pomegranate anthracnose caused by Colletotrichum gloeosporioides. Int. J. Chem. Stud. 2020, 8, 3669–3674. [Google Scholar] [CrossRef]
- Xavier, K.; Vallad, G.E. Efficacy of biological and conventional fungicide programs for foliar disease management on pomegranate (Punica granatum) in Florida. Plant Health Prog. 2020, 21, 199–204. [Google Scholar] [CrossRef]
- Xavier, K.; Kc, A.N.; Vallad, G.E. Fungicide application timing essential for the management of leaf spot and fruit rot on pomegranate (Punica granatum L.) in Florida. Plant Dis. 2020, 104, 1629–1637. [Google Scholar] [CrossRef]
- Sarkhosh, A.; Zamani, Z.; Fatahi, R.; Ranjbar, H. Evaluation of genetic diversity among Iranian soft-seed pomegranate accessions by fruit characteristics and RAPD markers. Sci. Hortic. 2009, 121, 313–319. [Google Scholar] [CrossRef]
- Chandra, R.; Vilas Jadhav, T.; Sharma, J. Global scenario of pomegranate (Punica granatum L.) culture with special reference to India. Fruit Veg. Cereal Sci. Biotechnol. 2010, 4, 7–18. [Google Scholar]
- Joshi, M.S.; Sawant, D.M.; Gaikwad, A.P. Isolate variations in Colletotrichum gloeosporioides infecting pomegranate. J. Plant Prot. Sci. 2014, 6, 21–26. [Google Scholar]
- Jayalakshmi, K.; Nargund, V.B.; Raju, J.; Benagi, V.I.; Raghu, S.; Giri, M.S.; Basamma, R.B.; Priti, S.; Rajput, R.B. Pomegranate anthracnose caused by Colletotrichum gloeosporioides: A menace in quality fruit production. J. Pure Appl. Microbiol. 2015, 9, 3093–3097. [Google Scholar]
- Yu, X.; Xavier, K.; Vallad, G.E.; Deng, Z. Diseases resistance in pomegranates: Importance, sources, breeding approaches, and progress. Proc. Fla. State Hortic. Soc. 2018, 131, 1–5. Available online: https://journals.flvc.org/fshs/article/view/114705/110032 (accessed on 20 May 2023).
- Castle, W. Pomegranates for Now-Accessions. 2022. Available online: https://crec.ifas.ufl.edu/extension/pomegranates/accessions/ (accessed on 20 May 2023).
- Chater, J.M.; Yavari, A.; Sarkhos, A.; Jia, Z.; Merhaut, D.J.; Preece, J.E.; Cossio, F.; Qin, G.; Liu, C.; Li, J.; et al. World pomegranate cultivars. In The Pomegranate: Botany, Production and Uses; Sarkhosh, A., Alimohammad, M.Y., Zamani, Z., Eds.; CABI: Oxfordshire, UK, 2021; pp. 157–199. [Google Scholar]
- GRIN-Global. U.S. National Plant Germplasm System. Available online: https://npgsweb.ars-grin.gov/gringlobal/search.aspx (accessed on 20 May 2023).
- de Mendiburu, F.; de Mendiburu, M.F. Package‘Agricolae’; R Package Version; 2021; pp. 3–5. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 14 April 2023).
- Phillips, D.A.; Harmon, P.F.; Olmstead, J.W.; Peres, N.A.; Munoz, P.R. Screening for susceptibility to anthracnose stem lesions in southern highbush blueberry. HortScience 2018, 53, 920–924. [Google Scholar] [CrossRef]
- Mangandi, J.; Peres, N.A.; Whitaker, V.M. Identifying resistance to crown rot caused by Colletotrichum gloeosporioides in strawberry. Plant Dis. 2015, 99, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Grice, K.R.E.; Bally, I.S.E.; Wright, C.L.; Maddox, C.; Ali, A.; Dillon, N.L. Mango germplasm screening for the identification of sources of tolerance to anthracnose. Australas. Plant Pathol. 2023, 52, 27–41. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, C.; Pervaiz, T.; Zhao, P.; Liu, Z.; Wang, B.; Wang, C.; Zhang, L.; Fang, J.; Qian, J. Jasmonic acid involves in grapefruit ripening and resistant against Botrytis cinerea. Funct. Integr. Genom. 2016, 16, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Sivankalyani, V.; Feygenberg, O.; Diskin, S.; Wright, B.; Alkan, N. Increased anthocyanin and flavonoids in mango fruit peel are associated with cold and pathogen resistance. Postharvest Biol. Technol. 2016, 111, 132–139. [Google Scholar] [CrossRef]
- Gat, T.; Liarzi, O.; Skovorodnikova, Y.; Ezra, D. Characterization of Alternaria alternata causing black spot disease of pomegranate in Israel using a molecular marker. Plant Dis. 2012, 96, 1513–1518. [Google Scholar] [CrossRef]
- Sharma, K.; Sharma, J.; Jadhav, V. Recent developments in bacterial blight of pomegranate and its management. In Recent Advances in the Diagnosis and Management of Plant Diseases; Awasthi, L.P., Ed.; Springer: New Delhi, India, 2015; pp. 119–126. ISBN 978-81-322-2571-3. [Google Scholar]
- Mirabolfathy, M.; Groenewald, J.Z.; Crous, P.W. First report of Pilidiella granati causing dieback and fruit rot of pomegranate (Punica granatum) in Iran. Plant Dis. 2011, 96, 461. [Google Scholar] [CrossRef]
- Peduto Hand, F.; Choudhury, R.A.; Gubler, W.D. First report of Cytospora punicae causing wood canker and branch dieback of pomegranate (Punica granatum) in the United States. Plant Dis. 2014, 98, 853. [Google Scholar] [CrossRef]
- Palavouzis, S.C.; Tzamos, S.; Paplomatas, E.; Thomidis, T. First report of Neofusicoccum parvum causing shoot blight of pomegranate in northern Greece. New Dis. Rep. 2015, 32, 10. [Google Scholar] [CrossRef]
- Kumari, N.; Ram, V. Evaluation of pomegranate germplasm for resistance against leaf spot and dry fruit rot (Coniella granati). Int. J. Farm Sci. 2015, 5, 97–104. [Google Scholar]
- Jabnoun-Khiareddine, H.; Aydi Ben Abdallah, R.; Daami-Remadi, M.; Mars, M. Response of Tunisian pomegranate (Punica granatum L.) cultivars and several plant hosts to Coniella granati (Saccardo). J. Hortic. 2018, 5, 1000245. [Google Scholar] [CrossRef]
- Mincuzzi, A.; Ippolito, A.; Brighenti, V.; Marchetti, L.; Benvenuti, S.; Ligorio, A.; Pellati, F.; Sanzani, S.M. The effect of polyphenols on pomegranate fruit susceptibility to Pilidiella granati provides insights into disease tolerance mechanisms. Molecules 2020, 25, 515. [Google Scholar] [CrossRef] [PubMed]
- Priya, B.T.; Murthy, B.N.S.; Gopalakrishnan, C.; Artal, R.B.; Jagannath, S. Identification of new resistant sources for bacterial blight in pomegranate. Eur. J. Plant Pathol. 2016, 146, 609–624. [Google Scholar] [CrossRef]
- Kumar, P.; Dashyal, M.S.; Doddaraju, P.; Meti, B.S.; Girigowda, M. Differential gene responses in different varieties of pomegranate during the pathogenesis of Xanthomonas axonopodis pv. punicae. 3 Biotech. 2021, 11, 180. [Google Scholar] [CrossRef]
- Singh, N.V.; Abburi, V.L.; Ramajayam, D.; Kumar, R.; Chandra, R.; Sharma, K.K.; Sharma, J.; Babu, K.D.; Pal, R.K.; Mundewadikar, D.M.; et al. Genetic diversity and association mapping of bacterial blight and other horticulturally important traits with microsatellite markers in pomegranate from India. Mol. Genet. Genom. 2015, 290, 1393–1402. [Google Scholar] [CrossRef]
- Usha, T.; Middha, S.K.; Babu, D.; Goyal, A.K.; Das, A.J.; Saini, D.; Sarangi, A.; Krishnamurthy, V.; Prasannakumar, M.K.; Saini, D.K.; et al. Hybrid assembly and annotation of the genome of the Indian Punica granatum, a superfood. Front. Genet. 2022, 13, 786825. [Google Scholar] [CrossRef]
- Roopa Sowjanya, P.; Shilpa, P.; Patil, G.P.; Babu, D.K.; Sharma, J.; Sangnure, V.R.; Mundewadikar, D.M.; Natarajan, P.; Marathe, A.R.; Reddy, U.K.; et al. Reference quality genome sequence of Indian pomegranate cv. ‘Bhagawa’ (Punica granatum L.). Front. Plant Sci. 2022, 13, 947164. [Google Scholar] [CrossRef]
- Luo, X.; Li, H.; Wu, Z.; Yao, W.; Zhao, P.; Cao, D.; Yu, H.; Li, K.; Poudel, K.; Zhao, D.; et al. The pomegranate (Punica granatum L.) draft genome dissects genetic divergence between soft- and hard-seeded cultivars. Plant Biotechnol. J. 2020, 18, 955–968. [Google Scholar] [CrossRef]
- Yuan, Z.; Fang, Y.; Zhang, T.; Fei, Z.; Han, F.; Liu, C.; Liu, M.; Xiao, W.; Zhang, W.; Wu, S.; et al. The pomegranate (Punica granatum L.) genome provides insights into fruit quality and ovule developmental biology. Plant Biotechnol. J. 2018, 16, 1363–1374. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Artificial Inoculation 2021 | Natural Infection 2021 | Artificial Inoculation 2022 | Natural Infection 2022 | ||||
|---|---|---|---|---|---|---|---|---|
| Afganski | 2.1 ± 0.71 | bc | 2.3 ± 0.37 | a–i | 4.0 ± 0.38 | abcd | 3.7 ± 0.37 | a–f |
| Al-Sirin-Nar | 3.0 ± 0.52 | abc | 4.0 ± 0.29 | abcd | 5.3 ± 0.24 | ab | 5.0 ± 0.20 | ab |
| Ambrosia | 4.0 ± 0.47 | ab | 3.7 ± 0.28 | a–e | 4.5 ± 0.27 | abc | 4.9 ± 0.18 | abc |
| Angel Red | 1.3 ± 0.56 | bc | 1.8 ± 0.48 | c–i | 3.3 ± 0.52 | a–f | ||
| Apseronski Krasnyj | 3.6 ± 0.18 | ab | 3.9 ± 0.33 | abcd | ||||
| Arakta | 1.4 ± 0.98 | bc | 0.4 ± 0.56 | hi | 0.8 ± 0.68 | gh | 0.8 ± 0.14 | gh |
| Azadi | 0.1 ± 0.12 | c | 0.1 ± 0.08 | i | 0.1 ± 0.07 | h | 0.3 ± 0.29 | h |
| Bala Miursal | 2.1 ± 0.63 | bc | 2.8 ± 0.46 | a–g | 1.4 ± 0.41 | efgh | 2.3 ± 0.29 | defg |
| Christina | 1.6 ± 0.94 | bc | 1.4 ± 0.45 | d–i | 0.2 ± 0.20 | h | 2.8 ± 0.95 | c–g |
| Cranberry | 1.2 ± 0.68 | bc | 3.8 ± 0.30 | abcd | 4.3 ± 1.09 | abcd | 1.6 ± 0.46 | fgh |
| Desertnyi | 3.3 ± 0.29 | ab | 4.4 ± 0.17 | ab | 4.9 ± 0.18 | ab | 4.3 ± 0.26 | abcd |
| Don Somner South | 1.1 ± 0.71 | bc | 1.9 ± 0.62 | c–i | 3.1 ± 0.33 | cde | 3.9 ± 0.43 | a–e |
| Eve | 2.9 ± 0.64 | abc | 4.3 ± 0.38 | abc | 5.5 ± 0.11 | a | 5.2 ± 0.24 | ab |
| Eversweet | 2.0 ± 1.39 | bc | 1.2 ± 0.54 | e–i | 0.7 ± 0.45 | h | 1.6 ± 0.51 | fgh |
| Fleischman | 1.0 ± 0.52 | bc | 0.4 ± 0.20 | ghi | 1.1 ± 0.60 | fgh | 0.5 ± 0.18 | gh |
| Gainey Sweet | 1.2 ± 0.64 | bc | 0.7 ± 0.38 | ghi | ||||
| Girkanets | 1.7 ± 0.65 | bc | 2.2 ± 0.49 | b–i | 2.3 ± 0.41 | defg | 2.6 ± 0.35 | defg |
| Gissarskii Rozovyi | 1.3 ± 0.58 | bc | 1.3 ± 0.39 | d–i | ||||
| Grenada | 1.6 ± 0.68 | bc | 2.8 ± 0.45 | a–g | 3.1 ± 0.58 | cde | 3.9 ± 0.27 | a–e |
| Jimmy Roppe | 1.4 ± 0.59 | bc | 0.6 ± 0.21 | ghi | 2.0 ± 0.60 | d–h | 2.0 ± 0.66 | d–h |
| Kaim-anor | 3.2 ± 0.64 | ab | 3.3 ± 0.51 | a–f | 4.5 ± 0.32 | abc | 4.8 ± 0.26 | abc |
| Kazake | 5.2 ± 0.38 | a | 4.6 ± 0.00 | a | 5.3 ± 0.16 | ab | 5.3 ± 0.12 | a |
| Larkin | 3.8 ± 0.88 | ab | 2.5 ± 0.15 | a–h | 2.7 ± 0.45 | def | 2.6 ± 0.45 | defg |
| Medovyi Vahsha | 3.1 ± 0.47 | abc | 1.8 ± 0.59 | c–i | ||||
| Molla Nepes | 4.4 ± 0.00 | ab | 1.2 ± 0.39 | e–i | ||||
| Nikitski Ranni | 3.4 ± 0.52 | ab | 3.3 ± 0.44 | a–e | 5.2 ± 0.17 | ab | 4.7 ± 0.15 | abc |
| Parfianka | 3.2 ± 0.64 | ab | 1.9 ± 0.40 | c–i | 3.7 ± 0.24 | bcd | 3.4 ± 0.28 | a–f |
| Rose | 3.2 ± 1.02 | abc | 1.9 ± 0.76 | c–i | ||||
| Sakerdze | 1.5 ± 0.75 | bc | 2.0 ± 0.41 | c–i | 2.5 ± 0.39 | def | 2.0 ± 0.68 | d–h |
| Salavatski | 2.6 ± 0.87 | bc | 2.3 ± 0.38 | a–i | 1.6 ± 0.41 | efgh | 1.9 ± 0.27 | efgh |
| Sin Pepe | 1.9 ± 0.66 | bc | 1.5 ± 0.45 | d–i | ||||
| Sirenevyi | 2.2 ± 1.27 | bc | 0.6 ± 0.47 | ghi | ||||
| Surh-Anor | 1.0 ± 0.54 | bc | 0.9 ± 0.48 | fghi | 3.0 ± 0.81 | cde | 3.2 ± 0.30 | b–f |
| Sweet | 1.6 ± 1.02 | bc | 2.3 ± 0.59 | a–i | 4.1 ± 0.42 | abcd | 4.0 ± 0.31 | abcd |
| Vkusnyi | 1.6 ± 0.60 | bc | 1.7 ± 0.34 | d–i | 1.8 ± 0.37 | efgh | 2.3 ± 0.44 | d–h |
| Categories of Fruit Rot Resistance Level | Cultivars | % Fruit Rot Mean | Disease Rating Mean (0–6 Scale) |
|---|---|---|---|
| Highly Resistant | Azadi | 2 | 0.2 |
| Resistant | Arakta, Christina, Eversweet, Fleishman, Jimmy Roppe, | 25 ± 1 | 1.2 ± 0.15 |
| Moderately Resistant | Angel Red, Bala Miursal, Cranberry, Don Somner South, Sakerdze, Salavatski, Surh-Anor, Vkusnyi | 39 ± 2 | 2.2 ± 0.10 |
| Susceptible | Afganski, Girkanets, Grenada, Larkin, Parfianka, Sweet | 61 ± 2 | 2.8 ± 0.12 |
| Highly Susceptible | Al-Sirin-Nar, Ambrosia, Desertnyi, Eve, Kaim-anor, Kazake, Nikitski Ranni | 87 ± 2 | 4.4 ± 0.13 |
| Experiment | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Cultivar | 1 | 2 | 3 | 4 | Average | ||||
| Afganski | 22.1 ± 1.9 | a | 20.6 ± 3.8 | abc | 20.3 ± 1.6 | a | 12.8 ± 2.8 | abc | 19.6 |
| Al-Sirin-Nar | 17.8 ± 1.1 | a | 21.2 ± 2.8 | ab | 9.4 ± 2.0 | bc | 8.3 ± 1.9 | bcd | 14.9 |
| Azadi | 7.6 ± 1.7 | b | 8.9 ± 1.9 | c | 4.7 ± 0.9 | c | 3.0 ± 0.0 | d | 6.0 |
| Eversweet | 21.7 ± 4.5 | a | 27.5 ± 4.0 | a | 8.3 ± 1.5 | bc | 4.0 ± 0.5 | cd | 15.5 |
| Fleishman | 13.8 ± 2.2 | ab | 18.8 ± 3.7 | abc | 5.1 ± 1.2 | bc | 12.7 | ||
| Kazake | 22.0 ± 1.9 | a | 12.0 ± 1.7 | bc | 20.8 ± 1.4 | a | 18.5 ± 1.5 | a | 18.4 |
| Nikitski Ranni | 15.3 ± 2.1 | ab | 23.8 ± 3.0 | a | 11.7 ± 2.1 | b | 12.9 ± 1.9 | ab | 15.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaller, A.; Chater, J.M.; Vallad, G.E.; Moersfelder, J.; Heinitz, C.; Deng, Z. Pomegranate Cultivars with Diverse Origins Exhibit Strong Resistance to Anthracnose Fruit Rot Caused by Colletotrichum gloeosporioides, a Major Disease in Southeast United States. Horticulturae 2023, 9, 1097. https://doi.org/10.3390/horticulturae9101097
Schaller A, Chater JM, Vallad GE, Moersfelder J, Heinitz C, Deng Z. Pomegranate Cultivars with Diverse Origins Exhibit Strong Resistance to Anthracnose Fruit Rot Caused by Colletotrichum gloeosporioides, a Major Disease in Southeast United States. Horticulturae. 2023; 9(10):1097. https://doi.org/10.3390/horticulturae9101097
Chicago/Turabian StyleSchaller, Alexander, John M. Chater, Gary E. Vallad, Jeff Moersfelder, Claire Heinitz, and Zhanao Deng. 2023. "Pomegranate Cultivars with Diverse Origins Exhibit Strong Resistance to Anthracnose Fruit Rot Caused by Colletotrichum gloeosporioides, a Major Disease in Southeast United States" Horticulturae 9, no. 10: 1097. https://doi.org/10.3390/horticulturae9101097