
The Effect of Biostimulants on Fruit Quality of Processing Tomato Grown under Deficit Irrigation

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Biostimulant Treatments and Experimental Design
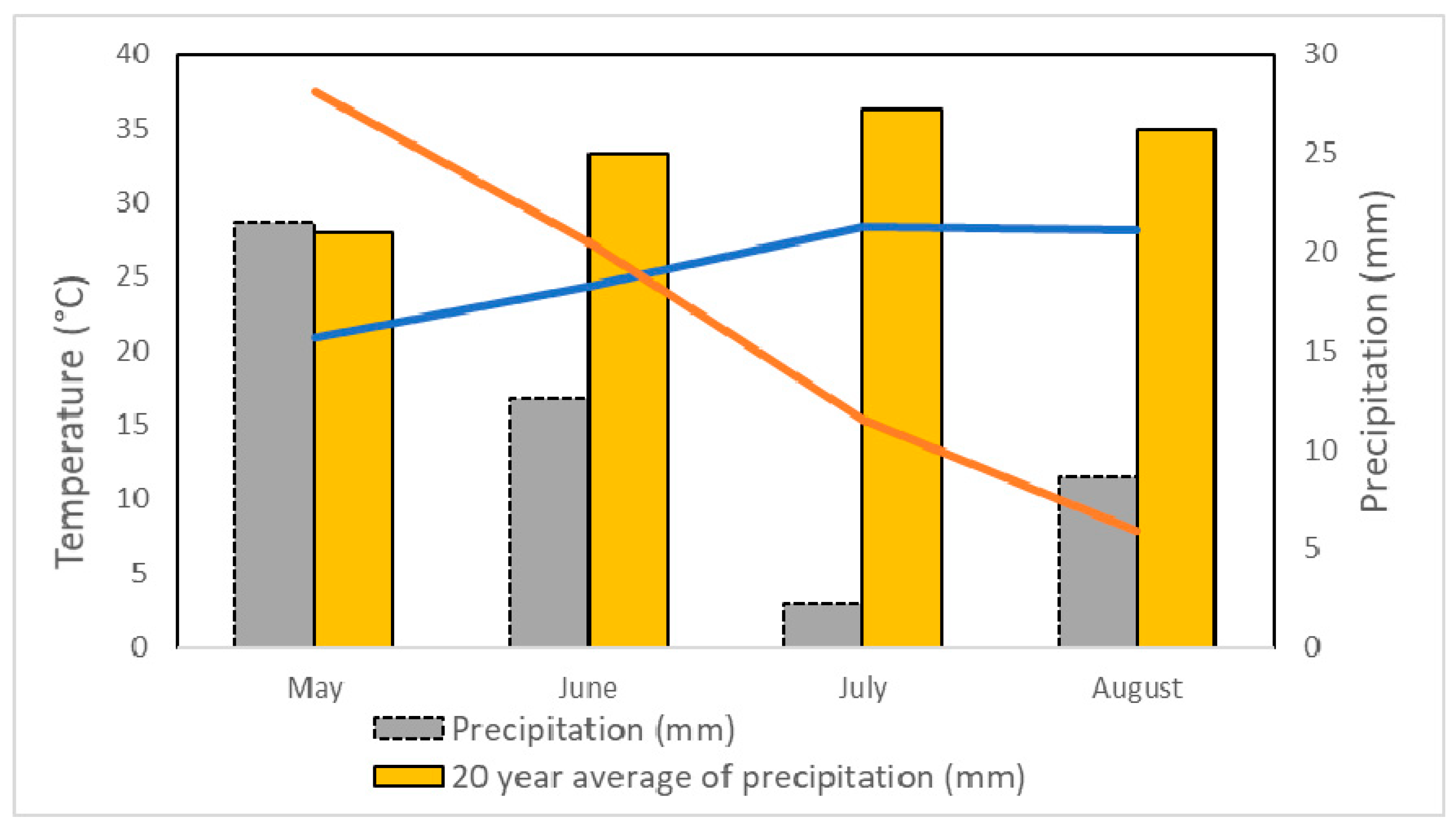
2.2. Irrigation Treatments
2.3. Chemical Composition Analysis
2.3.1. Proximate Composition and Energy
2.3.2. Free Sugars and Organic Acids
2.3.3. Fatty Acids, Tocopherols, and Carotenoids
2.4. Evaluation of Bioactive Properties
2.4.1. Preparation of Hydroethanolic Extracts
2.4.2. Total Phenolic and Flavonoids Contents
2.4.3. Thiobarbituric Acid Reactive Substances (TBARS) Assay
2.4.4. Oxidative Hemolysis Inhibition (OxHLIA) Assay
2.5. Statistical Analysis
3. Results and Discussion
3.1. Nutritional Value
3.2. Free Sugars and Organic Acids
3.3. Tocopherols and Carotenoids
3.4. Fatty Acids
3.5. Total Phenolic and Flavonoids Content and Antioxidant Activity
3.6. Principal Component Analysis (PCA)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Subramaniyan, L.; Veerasamy, R.; Prabhakaran, J.; Selvaraj, A. Biostimulation Effects of Seaweed Extract (Ascophyllum nodosum) on Phytomorpho-Physiological, Yield, and Quality Traits of Tomato (Solanum lycopersicum L.). Horticulturae 2023, 9, 348. [Google Scholar] [CrossRef]
- Perveen, R.; Ansar, H.; Suleria, R.; Anjum, F.M.; Butt, M.S.; Pasha, I.; Ahmad, S. Tomato (Solanum lycopersicum) Carotenoids and Lycopenes Chemistry; Metabolism, Absorption, Nutrition, and Allied Health Claims—A Comprehensive Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hu, T.; Feng, P.; Yao, D.; Gao, F.; Hong, X. Effect of potassium fertilization during fruit development on tomato quality, potassium uptake, water and potassium use efficiency under deficit irrigation regime. Agric. Water Manag. 2021, 250, 106831. [Google Scholar] [CrossRef]
- Fernandes, Â.; Chaski, C.; Pereira, C.; Kostić, M.; Rouphael, Y.; Soković, M.; Barros, L.; Petropoulos, S.A. Water Stress Alleviation Effects of Biostimulants on Greenhouse-Grown Tomato Fruit. Horticulturae 2022, 8, 645. [Google Scholar] [CrossRef]
- Islam, M.; Jahan, K.; Sen, A.; Urmi, T.A.; Haque, M.M. Exogenous Application of Calcium Ameliorates Salinity Stress Tolerance of Tomato (Solanum lycopersicum L.) and Enhances Fruit Quality. Antioxidants 2023, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Gorni, P.H.; de Lima, G.R.; Pereira, L.M.d.O.; Spera, K.D.; Lapaz, A.D.M.; Pacheco, A.C. Increasing plant performance, fruit production and nutritional value of tomato through foliar applied rutin. Sci. Hortic. 2022, 294, 110755. [Google Scholar] [CrossRef]
- Bantis, F.; Koukounaras, A. Ascophyllum nodosum and Silicon-Based Biostimulants Differentially Affect the Physiology and Growth of Watermelon Transplants under Abiotic Stress Factors: The Case of Drought. Horticulturae 2022, 8, 1177. [Google Scholar] [CrossRef]
- Kumari, A.; Lakshmi, G.A.; Krishna, G.K.; Patni, B.; Prakash, S.; Bhattacharyya, M.; Singh, S.K.; Verma, K.K. Climate Change and Its Impact on Crops: A Comprehensive Investigation for Sustainable Agriculture. Agronomy 2022, 12, 3008. [Google Scholar] [CrossRef]
- Gedeon, S.; Ioannou, A.; Balestrini, R.; Antoniou, C. Application of Biostimulants in Tomato Plants (Solanum lycopersicum) to Enhance Plant Growth and Salt Stress Tolerance. Plants 2022, 11, 3082. [Google Scholar] [CrossRef]
- Chaski, C.; Petropoulos, S.A. The Effects of Biostimulant Application on Growth Parameters of Lettuce Plants Grown under Deficit Irrigation Conditions. Horticulturae 2022, 8, 1089. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Petropoulos, S.A. Biostimulants application: A low input cropping management tool for sustainable farming of vegetables. Biomolecules 2021, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Atilgan, A.; Rolbiecki, R.; Saltuk, B.; Jagosz, B.; Arslan, F.; Erdal, I.; Aktas, H. Deficit Irrigation Stabilizes Fruit Yield and Alters Leaf Macro and Micronutrient Concentration in Tomato Cultivation in Greenhouses: A Case Study in Turkey. Agronomy 2022, 12, 2950. [Google Scholar] [CrossRef]
- Francaviglia, R.; Di Bene, C. Deficit Drip Irrigation in Processing Tomato Production in the Mediterranean Basin. A Data Analysis for Italy. Agriculture 2019, 9, 79. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Bertaki, M. Sustainable Water Management in Agriculture under Climate Change. Agric. Agric. Sci. Procedia 2015, 4, 88–98. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Chand, J.B.; Hewa, G.; Ph, D.; Hassanli, A.; Ph, D.; Myers, B.; Ph, D. Deficit Irrigation on Tomato Production in a Greenhouse Environment: A Review. J. Irrig. Drain. Eng. 2021, 147, 04020041. [Google Scholar] [CrossRef]
- Turhan, A.; Kuscu, H.; Asik, B.B. The Influence of Irrigation Strategies on Tomato Fruit Yield and Leaf Nutrient Contents. Gesunde Pflanz. 2022, 74, 1021–1027. [Google Scholar] [CrossRef]
- Vila e Vila, V.; Marques, P.A.A.; Rezende, R.; Wenneck, G.S.; Terassi, D.d.S.; Andrean, A.F.B.A.; Nocchi, R.C.d.F.; Matumoto-Pintro, P.T. Deficit Irrigation with Ascophyllum nodosum Extract Application as a Strategy to Increase Tomato Yield and Quality. Agronomy 2023, 13, 1853. [Google Scholar] [CrossRef]
- Jin, N.; Jin, L.; Wang, S.; Meng, X.; Ma, X.; He, X.; Zhang, G.; Luo, S.; Lyu, J.; Yu, J. A Comprehensive Evaluation of Effects on Water-Level Deficits on Tomato Polyphenol Composition, Nutritional Quality and Antioxidant Capacity. Antioxidants 2022, 11, 1585. [Google Scholar] [CrossRef]
- Wu, Y.; Yan, S.; Fan, J.; Zhang, F.; Xiang, Y. Scientia Horticulturae Responses of growth, fruit yield, quality and water productivity of greenhouse tomato to deficit drip irrigation. Sci. Hortic. 2021, 275, 109710. [Google Scholar] [CrossRef]
- Singh, M.; Singh, P.; Singh, S.; Saini, R.K.; Angadi, S.V. A global meta-analysis of yield and water productivity responses of vegetables to deficit irrigation. Sci. Rep. 2021, 11, 22095. [Google Scholar] [CrossRef] [PubMed]
- Chand, J.; Hewa, G.; Hassanli, A.; Myers, B. Evaluation of Deficit Irrigation and Water Quality on Production and Water Productivity of Tomato in Greenhouse. Agriculture 2020, 10, 297. [Google Scholar] [CrossRef]
- Coyago-cruz, E.; Corell, M.; Moriana, A.; Hernanz, D.; Stinco, C.M.; Mapelli-brahm, P.; Meléndez-Martínez, A.J. Effect of regulated deficit irrigation on commercial quality parameters, carotenoids, phenolics and sugars of the black cherry tomato (Solanum lycopersicum L.) ‘Sunchocola’. J. Food Compos. Anal. 2022, 105, 104220. [Google Scholar] [CrossRef]
- Bai, C.; Zuo, J.; Watkins, C.B.; Wang, Q.; Liang, H.; Zheng, Y.; Liu, M.; Ji, Y. Postharvest Biology and Technology Sugar accumulation and fruit quality of tomatoes under water deficit irrigation. Postharvest Biol. Technol. 2023, 195, 112112. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Agliassa, C.; Mannino, G.; Vigliante, I.; Contartese, V.; Secchi, F.; Bertea, C.M. A Biostimulant Based on Seaweed (Ascophyllum nodosum and Laminaria digitata) and Yeast Extracts Mitigates Water Stress Effects on Tomato (Solanum lycopersicum L.). Agriculture 2021, 11, 557. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]
- Rakkammal, K.; Maharajan, T.; Antony, S.; Manikandan, C. Biostimulants and their role in improving plant growth under drought and salinity. Cereal Res. Commun. 2023, 51, 61–74. [Google Scholar] [CrossRef]
- Sudiro, C.; Guglielmi, F.; Hochart, M.; Senizza, B.; Zhang, L.; Lucini, L.; Altissimo, A. A Phenomics and Metabolomics Investigation on the Modulation of Drought Stress by a Biostimulant Plant Extract in Tomato (Solanum lycopersicum). Agronomy 2022, 12, 764. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2014, 26, 619–628. [Google Scholar] [CrossRef]
- Niu, C.; Wang, G.; Sui, J.; Liu, G.; Ma, F.; Bao, Z. Scientia Horticulturae Biostimulants alleviate temperature stress in tomato seedlings. Sci. Hortic. 2022, 293, 110712. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, H.; Wu, G.; Chen, X.; Gruda, N.; Li, X.; Dong, J.; Duan, Z. Dose-Dependent Application of Straw-Derived Fulvic Acid on Yield and Quality of Tomato Plants Grown in a Greenhouse. Front. Plant Sci. 2021, 12, 7366313. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Miras Moreno, M.B.; Reynaud, H.; Canaguier, R.; Trtílek, M.; et al. A combined phenotypic and metabolomic approach for elucidating the biostimulant action of a plant-derived protein hydrolysate on tomato grown under limited water availability. Front. Plant Sci. 2019, 10, 493. [Google Scholar] [CrossRef] [PubMed]
- Domingo, G.; Marsoni, M.; Milena, Á.; Torres, M.D.; Dom, H.; Vannini, C. The Role of Protein-Rich Extracts from Chondrus crispus as Biostimulant and in Enhancing Tolerance to Drought Stress in Tomato Plants. Plants 2023, 12, 845. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zheng, W.; Lv, H.; Liang, B.; Jin, S.; Li, J. Animal-derived plant biostimulant alleviates drought stress by regulating photosynthesis, osmotic adjustment, and antioxidant systems in tomato plants. Sci. Hortic. 2022, 305, 111365. [Google Scholar] [CrossRef]
- Top, S.; Vandoorne, B.; Pauwels, E.; Perneel, M.; Christine, M.; Kathy, V.L. Plant Sensors Untangle the Water-Use and Growth Effects of Selected Seaweed-Derived Biostimulants on Drought-Stressed Tomato Plants (Solanum lycopersicum). J. Plant Growth Regul. 2023, 42, 5615–5627. [Google Scholar] [CrossRef]
- Jiménez-Arias, D.; Hernándiz, A.E.; Morales-Sierra, S.; García-García, A.L.; García-Machado, F.J.; Luis, J.C.; Borges, A.A. Applying Biostimulants to Combat Water Deficit in Crop Plants: Research and Debate. Agronomy 2022, 12, 571. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Francesca, S.; Najai, S.; Zhou, R.; Decros, G.; Cassan, C.; Delmas, F.; Ottosen, C.; Barone, A.; Manuela, M. Phenotyping to dissect the biostimulant action of a protein hydrolysate in tomato plants under combined abiotic stress. Plant Physiol. Biochem. 2022, 179, 32–43. [Google Scholar] [CrossRef]
- Alfosea-Simón, M.; Simón-Grao, S.; Zavala-Gonzalez, E.A.; Cámara-Zapata, J.M.; Simón, I.; Martínez-Nicolás, J.J.; Lidón, V.; Rodríguez-Ortega, W.M.; García-Sánchez, F. Application of biostimulants containing amino acids to tomatoes could favor sustainable cultivation: Implications for tyrosine, lysine, and methionine. Sustainability 2020, 12, 9279. [Google Scholar] [CrossRef]
- Hamedeh, H.; Antoni, S.; Cocciaglia, L.; Ciccolini, V. Molecular and Physiological Effects of Magnesium–Polyphenolic Compound as Biostimulant in Drought Stress Mitigation in Tomato. Plants 2022, 11, 586. [Google Scholar] [CrossRef]
- Cristina, M.; Lucia, D.; Baghdadi, A.; Mangione, F.; Borella, M.; Zegada-lizarazu, W.; Ravi, S.; Deb, S.; Broccanello, C.; Concheri, G.; et al. Transcriptional and Physiological Analyses to Assess the Effects of a Novel Biostimulant in Tomato. Front. Plant Sci. 2022, 12, 781993. [Google Scholar] [CrossRef]
- Petropoulos, S.A. Practical applications of plant biostimulants in greenhouse vegetable crop production. Agronomy 2020, 10, 1569. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Petropoulos, S.A.; Sun, W. Survey of the Influences of Microbial Biostimulants on Horticultural Crops: Case Studies and Successful Paradigms. Horticulturae 2023, 9, 193. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Tzortzakis, N.; Petropoulos, S.A. Sustainable Agriculture Systems in Vegetable Production Using Chitin and Chitosan as Plant Biostimulants. Biomolecules 2021, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of Association of Official Analytical Chemists; Horwitz, W., Latimer, G., Eds.; AOAC Inter.: Gaithersburg, MD, USA, 2019; ISBN 0935584773. [Google Scholar]
- Spréa, R.M.; Fernandes, Â.; Calhelha, R.C.; Pereira, C.; Pires, T.C.S.P.; Alves, M.J.; Canan, C.; Barros, L.; Amaral, J.S.; Ferreira, I.C.F.R. Chemical and bioactive characterization of the aromatic plant Levisticum officinale W.D.J. Koch: A comprehensive study. Food Funct. 2020, 11, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Keutgen, A.J.; Pawelzik, E. Quality and nutritional value of strawberry fruit under long term salt stress. Food Chem. 2008, 107, 1413–1420. [Google Scholar] [CrossRef]
- Pereira, C.G.; Prado, J.M.; Meireles, M.A.A. Economic Evaluation of Natural Product Extraction Processes. In Natural Product Extraction: Principles and Applications; RSC Green Chemistry: Cambridge, UK, 2013; pp. 442–471. [Google Scholar]
- Pinela, J.; Barreira, J.C.M.; Barros, L.; Cabo Verde, S.; Antonio, A.L.; Carvalho, A.M.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Suitability of gamma irradiation for preserving fresh-cut watercress quality during cold storage. Food Chem. 2016, 206, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon. Shokuhin Kogyo Gakkaishi 1992, 39, 925–928. [Google Scholar] [CrossRef]
- Añibarro-Ortega, M.; Pinela, J.; Ćirić, A.; Martins, V.; Rocha, F.; Soković, M.D.; Barata, A.M.; Carvalho, A.M.; Barros, L.; Ferreira, I.C.F.R. Valorisation of table tomato crop by-products: Phenolic profiles and in vitro antioxidant and antimicrobial activities. Food Bioprod. Process. 2020, 124, 307–319. [Google Scholar] [CrossRef]
- Lockowandt, L.; Pinela, J.; Roriz, C.L.; Pereira, C.; Abreu, R.M.V.; Calhelha, R.C.; Alves, M.J.; Barros, L.; Bredol, M.; Ferreira, I.C.F.R. Chemical features and bioactivities of cornflower (Centaurea cyanus L.) capitula: The blue flowers and the unexplored non-edible part. Ind. Crops Prod. 2019, 128, 496–503. [Google Scholar] [CrossRef]
- Mu, L.; Fang, L. Changes in Tomato Fruit Quality and Antioxidant Enzyme Activities under Deficit Irrigation and Fertilizer Application in a Solar Greenhouse in Northwest China. Commun. Soil Sci. Plant Anal. 2016, 47, 1329–1341. [Google Scholar] [CrossRef]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef] [PubMed]
- Stagnari, F.; Galieni, A.; Pisante, M. Drought stress effects on crop quality. In Water Stress and Crop Plants: A Sustainable Approach; Ahmad, P., Ed.; John Willey & Sons, Ltd.: Hoboken, NJ, USA, 2016; Volume 2, pp. 375–392. [Google Scholar]
- Lipan, L.; Issa-issa, H.; Moriana, A.; Zurita, M.; Galindo, A.; Martín-Palomo, M.J.; Andreu, L.; Carbonell-Barrachina, Á.A.; Hernández, F.; Corell, M. Scheduling Regulated Deficit Irrigation with Leaf Water Potential of Cherry Tomato in Greenhouse and its Effect on Fruit Quality. Agriculture 2021, 11, 669. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Bonini, P.; Colla, G.; Rouphael, Y. Yield and nutritional quality of Vesuvian piennolo tomato PDO as affected by farming system and biostimulant application. Agronomy 2019, 9, 505. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Sánchez-Hernández, C.V.; Palmeros-Suárez, P.A.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Meza-Canales, I.D.; Becerril-Espinosa, A. Seaweed Extract Improves Growth and Productivity of Tomato Plants under Salinity Stress. Agronomy 2022, 121, 2495. [Google Scholar] [CrossRef]
- Distefano, M.; Steingass, C.B.; Leonardi, C.; Giuffrida, F.; Schweiggert, R.; Mauro, R.P. Effects of a plant-derived biostimulant application on quality and functional traits of greenhouse cherry tomato cultivars. Food Res. Int. 2022, 157, 111218. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Carillo, P.; Colla, G.; Graziani, G.; Di Mola, I.; Mori, M.; Kyriacou, M.C.; Rouphael, Y.; Soteriou, G.A.; et al. Plant-Derived Biostimulants Differentially Modulate Primary and Secondary Metabolites and Improve the Yield Potential of Red and Green Lettuce Cultivars. Agronomy 2022, 12, 1361. [Google Scholar] [CrossRef]
- Ramawat, N.; Bhardwaj, V. Biostimulants: Exploring Sources and Applications; Springer: Berlin/Heidelberg, Germany, 2022; ISBN 978-981-16-7079-4. [Google Scholar]
- Agius, C.; Von Tucher, S.; Poppenberger, B.; Rozhon, W. Quantification of sugars and organic acids in tomato fruits. MethodsX 2018, 5, 537–550. [Google Scholar] [CrossRef]
- Ma, W.F.; Li, Y.B.; Nai, G.J.; Liang, G.P.; Ma, Z.H.; Chen, B.H.; Mao, J. Changes and response mechanism of sugar and organic acids in fruits under water deficit stress. PeerJ 2022, 10, e13691. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, X.; Ali, M.M.; Rizwan, H.M.; Li, B.; Li, H.; Jia, K.; Yang, X.; Ma, S.; Li, S.; et al. Changes in the Content of Organic Acids and Expression Analysis of Citric Acid Accumulation-Related Genes during Fruit Development of Yellow (Passiflora edulis f. flavicarpa) and Purple (Passiflora edulis f. edulis) Passion Fruits. Int. J. Mol. Sci. 2021, 22, 5765. [Google Scholar] [CrossRef]
- Murtic, S.; Oljaca, R.; Murtic, M.S.; Vranac, A.; Koleska, I. Effects of seaweed extract on the growth, yield and quality of cherry tomato under different growth conditions. Acta Agric. Slov. 2018, 111, 315–325. [Google Scholar] [CrossRef]
- Saldeen, K.; Saldeen, T. Importance of tocopherols beyond a -tocopherol: Evidence from animal and human studies. Nutr. Res. 2005, 25, 877–889. [Google Scholar] [CrossRef]
- Pereira, C.; Dias, M.I.; Petropoulos, S.A.; Plexida, S.; Chrysargyris, A.; Tzortzakis, N.; Calhelha, R.C.; Ivanov, M.; Stojković, D.; Soković, M.; et al. The effects of biostimulants, biofertilizers and water-stress on nutritional value and chemical composition of two spinach genotypes (Spinacia oleracea L.). Molecules 2019, 24, 4494. [Google Scholar] [CrossRef]
- Oh, M.; Trick, H.N.; Rajashekar, C.B. Secondary metabolism and antioxidants are involved in environmental adaptation and stress tolerance in lettuce. J. Plant Physiol. 2009, 166, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Mathur, P.; Ding, Z.; Saldeen, T. Tocopherols in the Prevention and Treatment of Atherosclerosis and Related Cardiovascular Disease. Clin. Cardiol. 2015, 38, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Fernandes, Â.; Plexida, S.; Chrysargyris, A.; Tzortzakis, N.; Barreira, J.C.M.; Barros, L.; Ferreira, I.C.F.R. Biostimulants application alleviates water stress effects on yield and chemical composition of greenhouse green bean (Phaseolus vulgaris L.). Agronomy 2020, 10, 181. [Google Scholar] [CrossRef]
- Zushi, K.; Matsuzoe, N. Metabolic profile of organoleptic and health-promoting qualities in two tomato cultivars subjected to salt stress and their interactions using correlation network analysis. Sci. Hortic. 2015, 184, 8–17. [Google Scholar] [CrossRef]
- Costan, A.; Stamatakis, A.; Chrysargyris, A.; Petropoulos, S.A.; Tzortzakis, N. Interactive effects of salinity and silicon application on Solanum lycopersicum growth, physiology and shelf-life of fruit produced hydroponically. J. Sci. Food Agric. 2020, 100, 732–743. [Google Scholar] [CrossRef]
- Zhang, L.; Freschi, G.; Rouphael, Y.; De Pascale, S.; Lucini, L. The differential modulation of secondary metabolism induced by a protein hydrolysate and a seaweed extract in tomato plants under salinity. Front. Plant Sci. 2023, 13, 1072782. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional Indices for Assessing Fatty Acids: A Mini-Review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An Increase in the Omega-6 / Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Kaliannan, K.; Li, X.; Wang, B.; Pan, Q.; Chen, C.; Hao, L.; Xie, S.; Kang, J.X. Multi-omic analysis in transgenic mice implicates omega-6/omega-3 fatty acid imbalance as a risk factor for chronic disease. Commun. Biol. 2019, 2, 276. [Google Scholar] [CrossRef]
- Borguini, R.G.; da Silva Torres, E.A.F. Tomatoes and tomato products as dietary sources of antioxidants. Food Rev. Int. 2009, 25, 313–325. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regulated Deficit Irrigation | Moisture (g 100 g−1 fw) | Proteins (g 100 g−1 fw) | Crude Fat (g 100 g−1 fw) | Ash (g 100 g−1 fw) | Carbohydrates (g 100 g−1 fw) | Energy (kcal 100 g−1 fw) |
|---|---|---|---|---|---|---|
| Tr1 | 92.7 ± 1.8 c | 1.04 ± 0.03 bc | 0.082 ± 0.003 b | 0.47 ± 0.02 d | 5.7 ± 0.2 b | 27.9 ± 0.1 bc |
| Tr2 | 92.1 ± 1.6 d | 1.04 ± 0.02 bc | 0.075 ± 0.005 cd | 0.61 ± 0.03 a | 6.2 ± 0.2 a | 29.7 ± 0.8 a |
| Tr3 | 92.0 ± 0.7 d | 1.25 ± 0.03 a | 0.075 ± 0.005 cd | 0.59 ± 0.02 a | 6.1 ± 0.2 a | 30.1 ± 0.8 a |
| Tr4 | 92.8 ± 1.3 c | 1.07 ± 0.06 b | 0.062 ± 0.003 f | 0.59 ± 0.03 a | 5.5 ± 0.2 b | 26.6 ± 0.8 c |
| Control | 92.5 ± 0.5 c | 1.28 ± 0.02 a | 0.077 ± 0.003 c | 0.55 ± 0.02 b | 5.6 ± 0.2 b | 28.2 ± 0.8 b |
| Regular Irrigation | Moisture (g 100 g−1 fw) | Proteins (g 100 g−1 fw) | Crude fat (g 100 g−1 fw) | Ash (g 100 g−1 fw) | Carbohydrates (g 100 g−1 fw) | Energy (kcal 100 g−1 fw) |
| Tr1 | 94.0 ± 1.3 a | 1.01 ± 0.01 c | 0.066 ± 0.001 ef | 0.50 ± 0.01 c | 4.4 ± 0.2 de | 22.2 ± 0.8 e |
| Tr2 | 93.8 ± 1.1 a | 0.86 ± 0.03 d | 0.070 ± 0.004 de | 0.49 ± 0.02 cd | 4.7 ± 0.2 cd | 23.0 ± 0.8 e |
| Tr3 | 93.9 ± 1.5 a | 0.86 ± 0.01 d | 0.077 ± 0.005 bc | 0.49 ± 0.01 cd | 4.6 ± 0.2 d | 22.7 ± 0.8 e |
| Tr4 | 94.2 ± 2.2 a | 1.05 ± 0.03 bc | 0.069 ± 0.002 e | 0.50 ± 0.02 c | 4.2 ± 0.2 e | 21.7 ± 0.8 e |
| Control | 93.3 ± 1.4 b | 1.04 ± 0.05 bc | 0.090 ± 0.005 a | 0.55 ± 0.03 b | 5.1 ± 0.2 c | 25.2 ± 0.7 d |
| Free Sugars (g 100 g−1 fw) | Organic Acids (mg 100 g−1 fw) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Regulated Deficit Irrigation | Fructose | Glucose | Sucrose | Total | Oxalic Acid | Malic Acid | Ascorbic Acid | Citric Acid | Total |
| Tr1 | 3.3 ± 0.2 a | 1.58 ± 0.08 a | 0.040 ± 0.002 b | 4.9 ± 0.2 a | 71 ± 1 c | 544 ± 14 b | 14.5 ± 0.2 c | 754 ± 21 d | 1383 ± 12 c |
| Tr2 | 2.74 ± 0.06 c | 1.36 ± 0.05 c | 0.030 ± 0.001 c | 4.13 ± 0.02 c | 48 ± 3 f | 616 ± 26 a | 17.0 ± 0.7 a | 815 ± 29 c | 1497 ± 59 b |
| Tr3 | 2.4 ± 0.2 d | 1.02 ± 0.06 d | 0.030 ± 0.002 c | 3.4 ± 0.2 d | 61 ± 2 d | 473 ± 19 c | 7.9 ± 0.2 f | 821 ± 25 c | 1362 ± 7 c |
| Tr4 | 3.05 ± 0.08 b | 1.48 ± 0.05 b | 0.040 ± 0.002 b | 4.57 ± 0.04 b | 61 ± 4 de | 602 ± 29 a | 16.3 ± 0.2 b | 724 ± 30 e | 1403 ± 63 c |
| Control | 2.37 ± 0.02 d | 1.00 ± 0.05 d | 0.060 ± 0.003 a | 3.43 ± 0.04 d | 57 ± 3 e | 588 ± 39 a | 13.3 ± 0.2 d | 941 ± 9 a | 1599 ± 12 a |
| Regular Irrigation | Fructose | Glucose | Sucrose | Total | Oxalic Acid | Malic Acid | Ascorbic Acid | Citric Acid | Total |
| Tr1 | 2.2 ± 0.1 e | 0.91 ± 0.05 e | 0.030 ± 0.001 d | 3.16 ± 0.08 ef | 57 ± 2 de | 329 ± 16 d | 3.8 ± 0.1 g | 634 ± 7 f | 1024 ± 25 e |
| Tr2 | 2.04 ± 0.09 f | 0.97 ± 0.07 de | 0.0100 ± 0.0003 e | 3.0 ± 0.2 f | 49 ± 4 f | 276 ± 9 e | 0.50 ± 0.01 j | 577 ± 18 g | 902 ± 10 g |
| Tr3 | 1.84 ± 0.04 g | 0.75 ± 0.04 f | 0.010 ± 0.001 e | 2.60 ± 0.09 g | 61 ± 2 de | 321 ± 22 d | 2.6 ± 0.1 h | 701 ± 10 e | 1085 ± 30 d |
| Tr4 | 1.88 ± 0.05 g | 0.57 ± 0.02 g | 0.0100 ± 0.0002 e | 2.46 ± 0.06 g | 102 ± 3 a | 303 ± 6 de | 1.7 ± 0.1 i | 565 ± 20 g | 972 ± 10 f |
| Control | 2.33 ± 0.02 de | 0.95 ± 0.05 de | 0.010 ± 0.001 e | 3.30 ± 0.03 de | 77 ± 6 b | 541 ± 25 b | 11.8 ± 0.5 e | 888 ± 15 b | 1517 ± 36 b |
| Tocopherols (μg 100 g−1 fw) | Carotenoids (μg 100 g−1 fw) | ||||||
|---|---|---|---|---|---|---|---|
| Regulated Deficit Irrigation | α-Tocopherol | β-Tocopherol | γ-Tocopherol | δ-Tocopherol | Total | Lycopene | β-Carotene |
| Tr1 | 520 ± 6 b | 58 ± 1 c | 143 ± 3 b | 11.5 ± 0.3 a | 732 ± 10 b | 404 ± 4 g | 346 ± 4 c |
| Tr2 | 448 ± 4 c | 73 ± 3 b | 108 ± 1 c | 9.8 ± 0.4 b | 639 ± 9 c | 728 ± 8 b | 506 ± 4 a |
| Tr3 | 555 ± 29 a | nd | 233 ± 11 a | nd | 788 ± 39 a | 761 ± 16 a | 515 ± 26 a |
| Tr4 | 406 ± 12 d | 78 ± 1 a | 110 ± 1 c | 6.9 ± 0.5 c | 601 ± 11 d | 568 ± 9 c | 398 ± 11 b |
| Control | 408 ± 4 d | 33 ± 1 h | 99 ± 1 d | nd | 540 ± 6 e | 489 ± 5 e | 391 ± 1 b |
| Regular Irrigation | Fructose | Glucose | Sucrose | Total | Oxalic Acid | Malic Acid | Ascorbic Acid |
| Tr1 | 212 ± 3 g | 52 ± 2 e | 51 ± 2 h | nd | 314 ± 7 h | 238 ± 14 j | 199 ± 4 f |
| Tr2 | 213 ± 2 g | 36 ± 2 g | 63 ± 4 f | nd | 312 ± 4 h | 358 ± 14 h | 224 ± 18 e |
| Tr3 | 215 ± 2 g | 18 ± 1 i | 42 ± 2 i | nd | 275 ± 5 i | 272 ± 5 i | 191 ± 7 f |
| Tr4 | 290 ± 3 f | 55 ± 2 d | 56 ± 2 g | nd | 401 ± 8 g | 544 ± 6 d | 337 ± 22 cd |
| Control | 319 ± 10 e | 48 ± 2 f | 79 ± 2 e | nd | 446 ± 14 f | 441 ± 5 f | 321 ± 7 d |
| Fatty Acids | Categories | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Deficit Irrigation | C14:0 | C16:0 | C18:0 | C18:1n9 | C18:2n6 | C18:3n3 | C20:0 | C23:0 | C24:0 | SFA | MUFA | PUFA | PUFA/SFA | n6/n3 |
| Tr1 | 0.62 ± 0.04 e | 24.9 ± 0.3 g | nd | 24.4 ± 0.8 a | 40.3 ± 0.5 a | 6.1 ± 0.4 b | nd | 0.26 ± 0.02 | nd | 26.3 ± 0.3 h | 27 ± 1 a | 47 ± 1 a | 1.79 ± 0.04 a | 6.5 ± 0.2 c |
| Tr2 | 0.75 ± 0.01 c | 27 ± 1 f | nd | 21.6 ± 0.6 b | 38.4 ± 0.5 b | 7.5 ± 0.2 a | nd | 0.34 ± 0.01 | 1.03 ± 0.04 b | 30 ± 1 g | 23 ± 1 b | 47.1 ± 0.3 a | 1.59 ± 0.04 b | 5.1 ± 0.1 d |
| Tr3 | 1.00 ± 0.06 b | 52 ± 1 a | 9.0 ± 0.4 c | 5.5 ± 0.4 fg | 24 ± 2 f | 3.3 ± 0.1 c | 1.09 ± 0.07 cd | 2.1 ± 0.2 | 0.65 ± 0.02 f | 67 ± 1 ab | 5.9 ± 0.3 f | 27 ± 2 c | 0.40 ± 0.02 d | 7.1 ± 0.5 c |
| Tr4 | 1.18 ± 0.06 a | 50 ± 1 b | 9.3 ± 0.1 c | 5.1 ± 0.1 g | 23.2 ± 0.9 f | 2.6 ± 0.1 d | 1.01 ± 0.03 e | 3.7 ± 0.1 | 0.97 ± 0.07 c | 68 ± 1 a | 5.4 ± 0.1 g | 27 ± 1 c | 0.39 ± 0.01 d | 9.00 ± 0.01 b |
| Control | 0.73 ± 0.05 c | 47 ± 1 c | 11.1 ± 0.6 a | 5.9 ± 0.1 ef | 27 ± 1 cd | 3.2 ± 0.1 c | 1.22 ± 0.06 b | 1.20 ± 0.03 | 0.63 ± 0.04 f | 63 ± 1 d | 6.3 ± 0.1 ef | 31 ± 1 b | 0.48 ± 0.01 c | 8.5 ± 0.1 b |
| Regular Irrigation | C14:0 | C16:0 | C18:0 | C18:1n9 | C18:2n6 | C18:3n3 | C20:0 | C23:0 | C24:0 | SFA | MUFA | PUFA | PUFA/SFA | n6/n3 |
| Tr1 | 0.72 ± 0.01 c | 45 ± 2 d | 9.0 ± 0.4 c | 15.5 ± 0.2 d | 21 ± 1 g | 2.2 ± 0.1 ef | 1.04 ± 0.07 de | 2.34 ± 0.09 | 0.73 ± 0.03 e | 61 ± 2 e | 15.7 ± 0.2 d | 24 ± 1 d | 0.39 ± 0.02 d | 9.6 ± 0.6 b |
| Tr2 | 0.70 ± 0.03 cd | 50 ± 1 b | 10.0 ± 0.3 b | 5.0 ± 0.4 g | 26 ± 1 de | 2.2 ± 0.2 ef | 1.13 ± 0.02 c | 2.89 ± 0.04 | 0.80 ± 0.05 d | 67 ± 2 ab | 5.2 ± 0.4 g | 28 ± 1 c | 0.42 ± 0.02 d | 11.8 ± 0.3 a |
| Tr3 | 0.65 ± 0.03 de | 47 ± 1 c | 11.3 ± 0.7 a | 6.3 ± 0.1 e | 25 ± 2 ef | 2.2 ± 0.1 ef | 1.24 ± 0.07 b | 2.9 ± 0.1 | 1.06 ± 0.07 b | 66 ± 2 bc | 6.6 ± 0.1 e | 27 ± 2 c | 0.41 ± 0.03 d | 12 ± 1 a |
| Tr4 | 0.66 ± 0.05 de | 50 ± 3 b | 11.5 ± 0.2 a | 5.0 ± 0.3 g | 28 ± 2 c | 2.4 ± 0.2 e | 1.65 ± 0.08 a | nd | 1.33 ± 0.05 a | 65 ± 3 cd | 5.0 ± 0.3 g | 30 ± 2 b | 0.47 ± 0.04 c | 11.8 ± 1.2 a |
| Control | 0.53 ± 0.01 f | 40 ± 1 e | 8.5 ± 0.3 d | 18.0 ± 0.4 c | 25.2 ± 0.6 e | 2.1 ± 0.1 f | 1.05 ± 0.04 de | 2.00 ± 0.03 | 0.67 ± 0.05 ef | 54 ± 1 f | 18.2 ± 0.4 c | 28 ± 1 c | 0.51 ± 0.01 c | 12.1 ± 0.5 a |
| Regulated Deficit Irrigation | Total Phenolic (mg GAE g−1 Extract) | Total Flavonoids (mg QE g−1 Extract) | TBARS (EC50, μg mL−1) | OxHLIA (IC50, μg mL−1) |
|---|---|---|---|---|
| Tr1 | 18.4 ± 0.7 g | 3.0 ± 0.1 b | 556 ± 21 b | 67 ± 5 de |
| Tr2 | 17.5 ± 0.5 h | 2.9 ± 0.1 c | 268 ± 17 g | 25 ± 2 f |
| Tr3 | 23.9 ± 0.3 a | 2.6 ± 0.1 f | 584 ± 17 a | 15.4 ± 0.6 g |
| Tr4 | 20.5 ± 0.6 c | 2.4 ± 0.1 g | 518 ± 7 c | 138 ± 8 b |
| Control | 24 ± 1 a | 2.7 ± 0.1 e | 190 ± 7 h | 67 ± 4 de |
| Regular Irrigation | Total Phenolic (mg GAE g−1 Extract) | Total Flavonoids (mg QE g−1 Extract) | TBARS (EC50, μg mL−1) | OxHLIA (IC50, μg mL−1) |
| Tr1 | 19.6 ± 0.7 d | 3.17 ± 0.04 a | 477 ± 17 d | 83 ± 3 d |
| Tr2 | 17.3 ± 0.4 h | 2.60 ± 0.03 ef | 270 ± 6 g | 55 ± 5 e |
| Tr3 | 18.9 ± 0.2 f | 3.0 ± 0.2 b | 332 ± 17 f | 62 ± 5 e |
| Tr4 | 22.5 ± 0.9 b | 2.6 ± 0.1 e | 326 ± 4 f | 106 ± 7 c |
| Control | 19.2 ± 0.7 e | 2.8 ± 0.1 d | 396 ± 7 e | 718 ± 21 a |
| Trolox | - | - | 5.4 ± 0.3 | 21.8 ± 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liava, V.; Chaski, C.; Añibarro-Ortega, M.; Pereira, A.; Pinela, J.; Barros, L.; Petropoulos, S.A. The Effect of Biostimulants on Fruit Quality of Processing Tomato Grown under Deficit Irrigation. Horticulturae 2023, 9, 1184. https://doi.org/10.3390/horticulturae9111184
Liava V, Chaski C, Añibarro-Ortega M, Pereira A, Pinela J, Barros L, Petropoulos SA. The Effect of Biostimulants on Fruit Quality of Processing Tomato Grown under Deficit Irrigation. Horticulturae. 2023; 9(11):1184. https://doi.org/10.3390/horticulturae9111184
Chicago/Turabian StyleLiava, Vasiliki, Christina Chaski, Mikel Añibarro-Ortega, Alexis Pereira, José Pinela, Lillian Barros, and Spyridon A. Petropoulos. 2023. "The Effect of Biostimulants on Fruit Quality of Processing Tomato Grown under Deficit Irrigation" Horticulturae 9, no. 11: 1184. https://doi.org/10.3390/horticulturae9111184