
Stepwise Optimization of the RT-qPCR Protocol and the Evaluation of Housekeeping Genes in Pears (Pyrus bretschneideri) under Various Hormone Treatments and Stresses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. RNA Isolation and cDNA Synthesis
2.3. Verification of Candidate Reference Genes
2.4. Optimization of qPCR Conditions
2.5. RT-qPCR and Data Analyses
2.6. Validation of Housekeeping Genes
2.7. Statics Analysis
3. Results
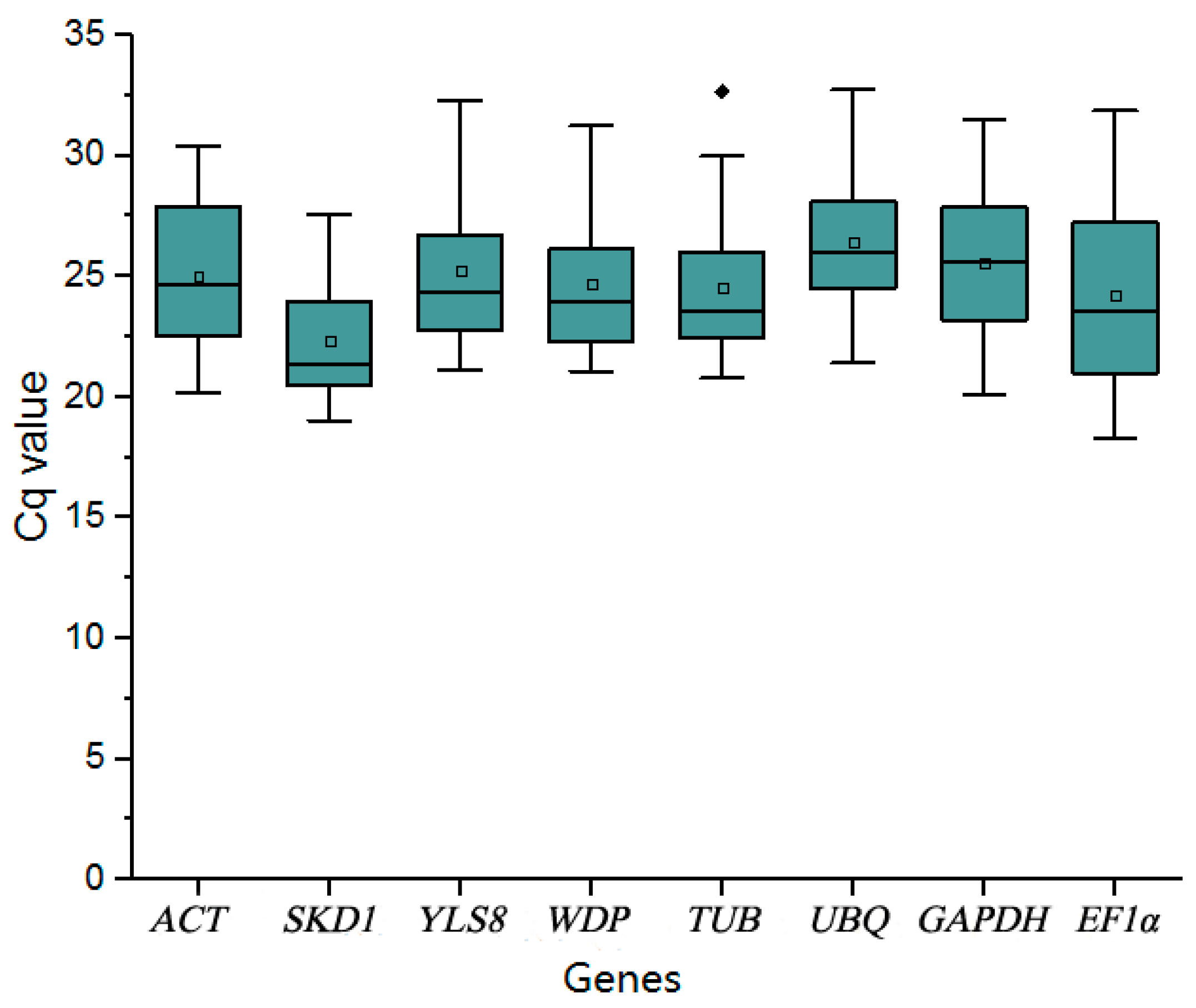
3.1. Assessment of Primer Specificity and PCR Amplification Efficiency
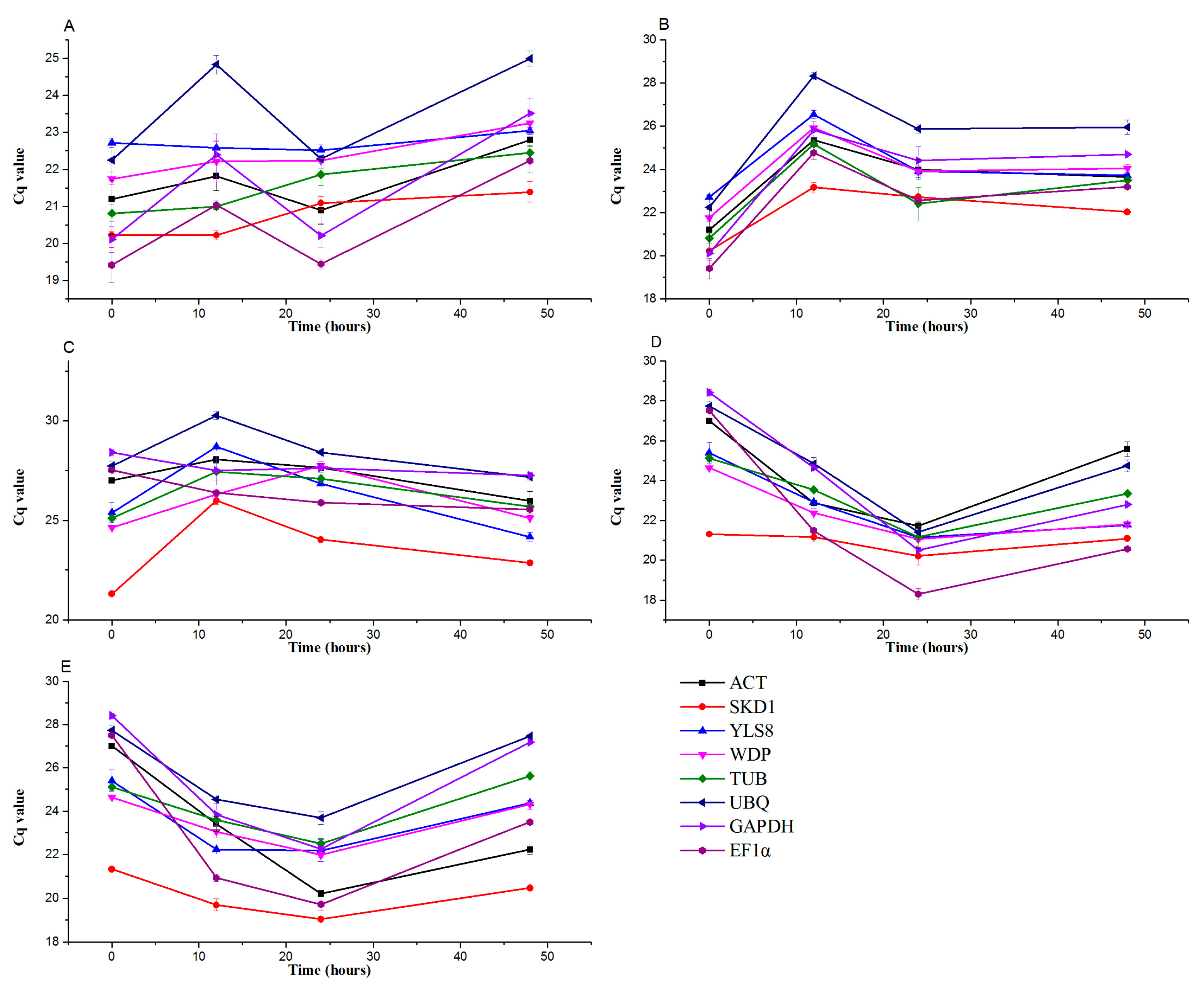
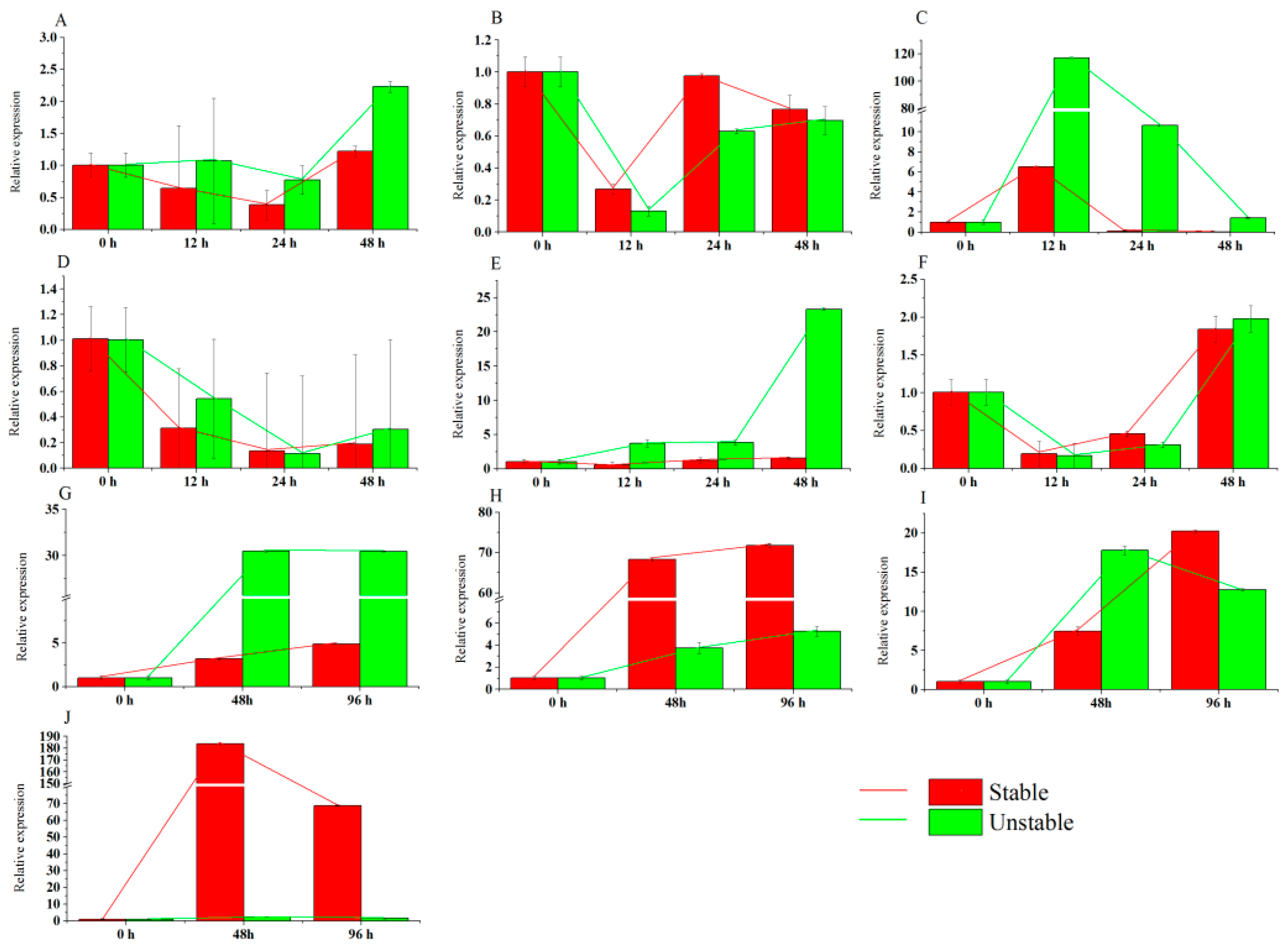
3.2. Transcript Abundance of the Eight Candidate Reference Genes in Pears under Hormone Treatments
3.3. Analysis of the Expression Stability of Reference Genes under Hormone Treatments
3.3.1. BestKeeper Analysis
3.3.2. NormFinder Analysis
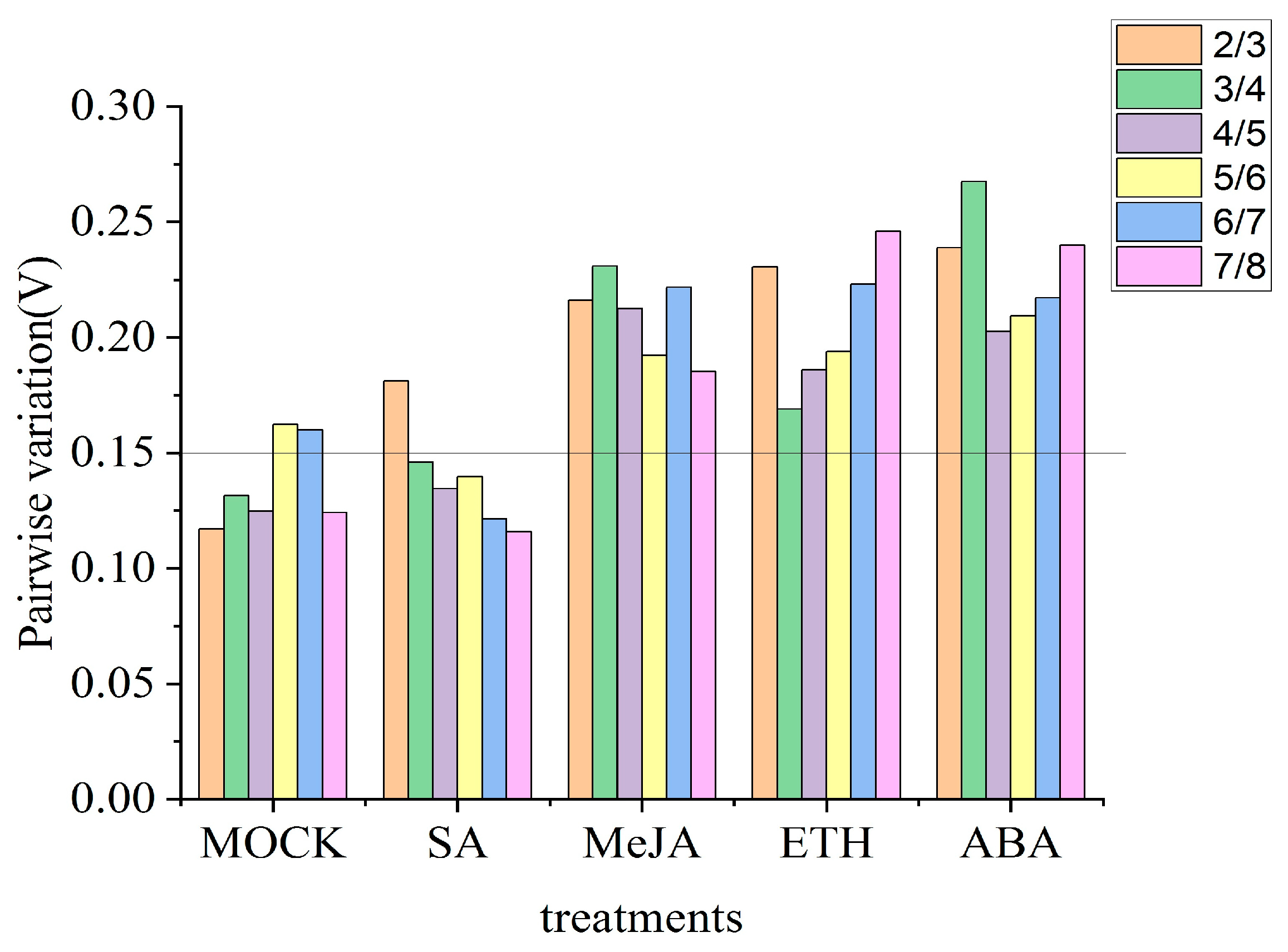
3.3.3. GeNorm Analysis
3.3.4. RefFinder Analysis
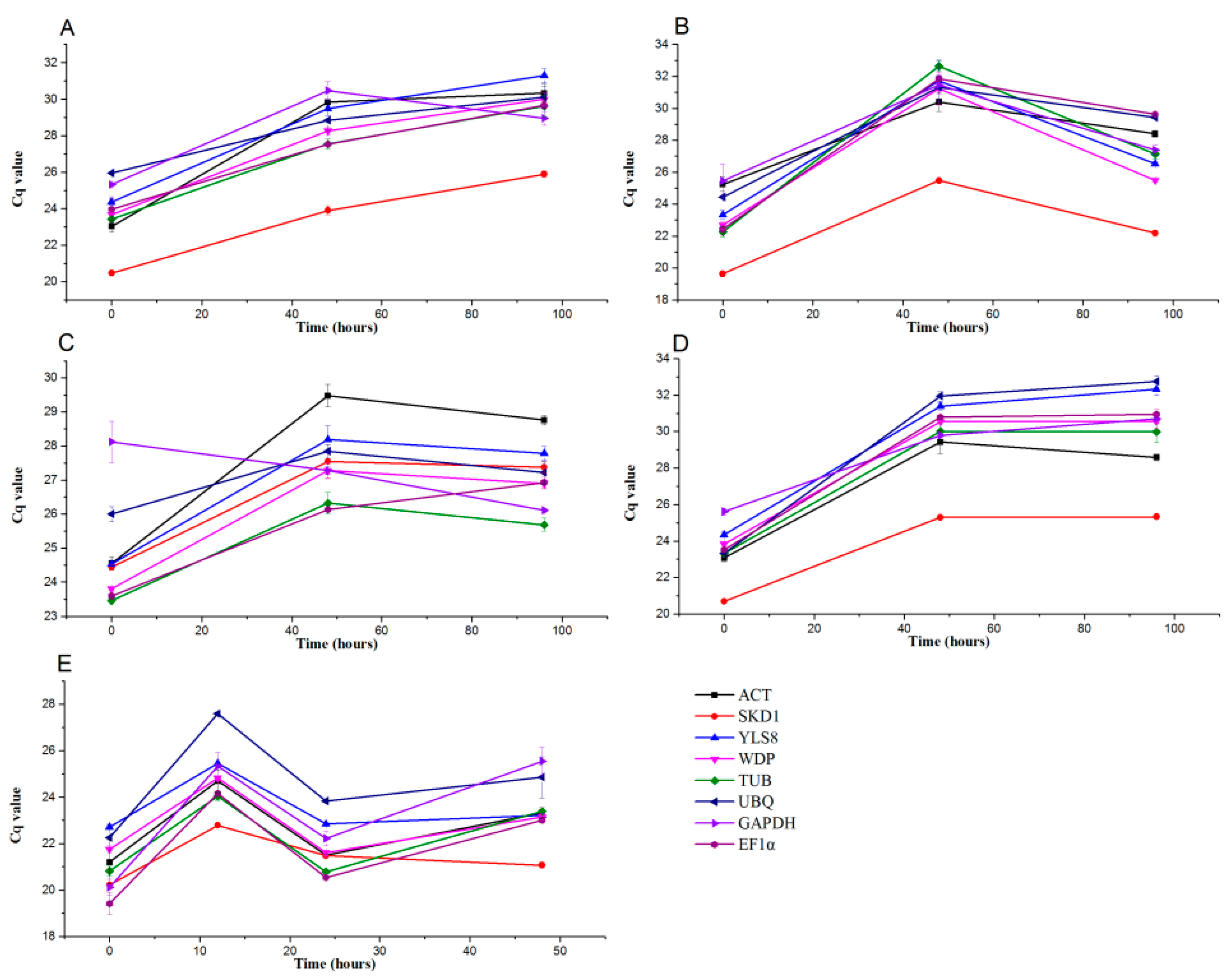
3.4. Transcript Abundance of the Eight Candidate Reference Genes in Pears under Abiotic and Biotic Stress Treatments
3.5. Analysis of the Expression Stability of Reference Genes under Abiotic and Biotic Stress Treatment
3.5.1. BestKeeper Analysis
3.5.2. NormFinder Analysis
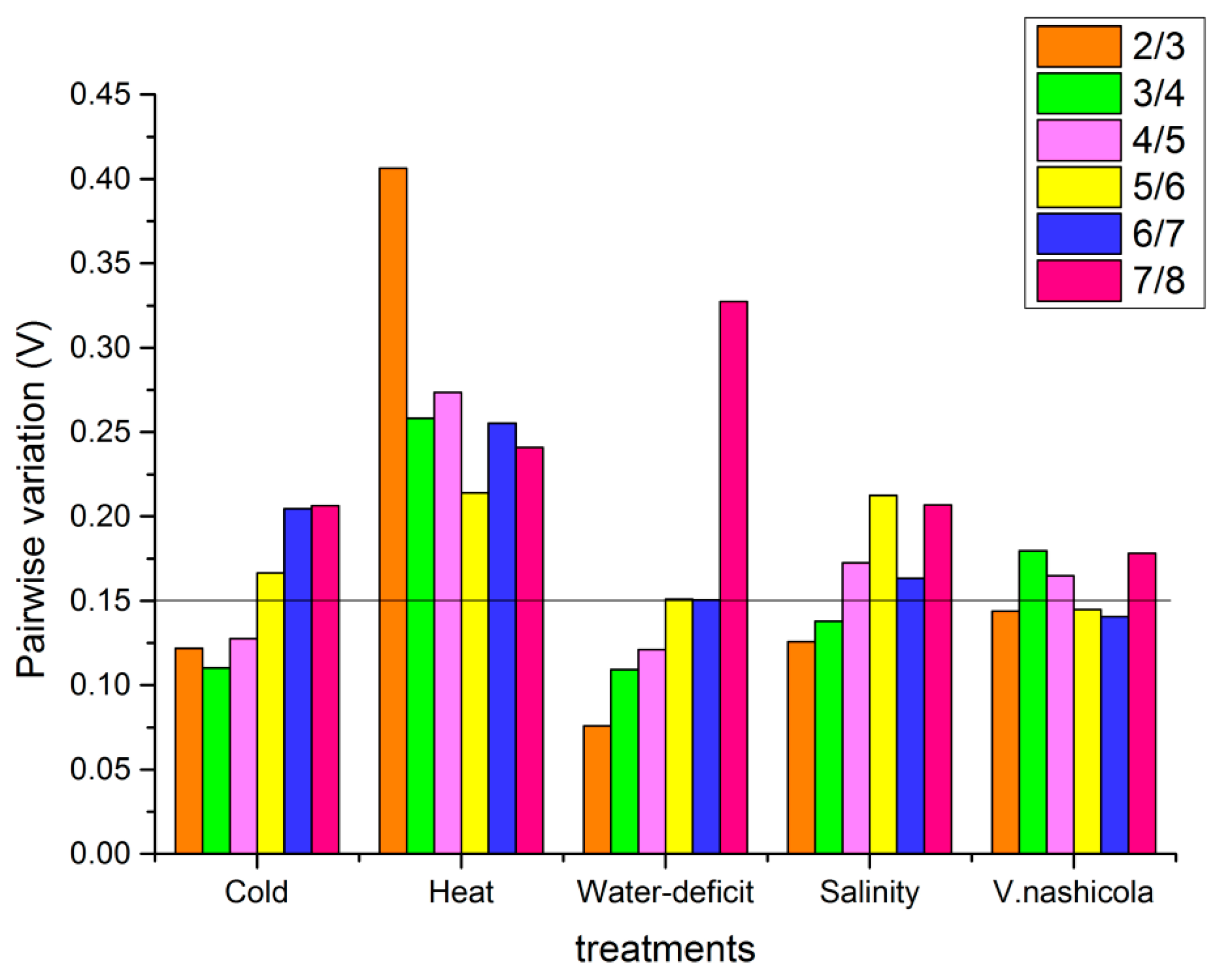
3.5.3. GeNorm Analysis
3.5.4. RefFinder Analysis
3.5.5. Validation of Selected Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wu, J.; Wang, Z.W.; Shi, Z.B.; Zhang, S.; Ming, R.; Zhu, S.L.; Khan, M.A.; Tao, S.T.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.T.; Li, Y.; Sun, Y.H. The breeding and application of the middle season ripening, scab resistant new pear variety-Huangguan pear. J. Hebei Agric. Sci. 1998, 2, 40–42. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Maren, N.A.; Kosentka, P.Z.; Kosentka, P.Z.; Liao, Y.Y.; Lu, H.Y.; Duduit, J.R.; Huang, D.B.; Ashrafi, H.; Zhao, T.J.; et al. An optimized protocol for stepwise optimization of real-time RT-PCR analysis. Hortic. Res. 2021, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, T.Y.; Lin, Z.K.; Gu, B.J.; Xing, C.H.; Zhao, L.Y.; Dong, H.Z.; Gao, J.Z.; Xie, Z.H.; Zhang, S.L.; et al. A WRKY transcription factor PbrWRKY53 from Pyrus betulaefolia is involved in drought tolerance and AsA accumulation. Plant Biotechnol. J. 2019, 17, 1770–1787. [Google Scholar] [CrossRef] [Green Version]
- Xing, C.H.; Liu, Y.; Zhao, L.Y.; Zhang, S.L.; Huang, X.S. A novel MYB transcription factor regulates ascorbic acid synthesis and affects cold tolerance. Plant Cell Environ. 2019, 42, 832–845. [Google Scholar] [CrossRef]
- Dong, H.Z.; Wang, C.M.; Xing, C.H.; Yang, T.Y.; Yan, J.X.; Gao, J.Z.; Li, D.L.; Wang, R.; Blumwald, E.; Zhang, S.L.; et al. Overexpression of PbrNHX2 gene, a Na+/H+ antiporter gene isolated from Pyrus betulaefolia, confers enhanced tolerance to salt stress via modulating ROS levels. Plant Sci. 2019, 285, 14–25. [Google Scholar] [CrossRef]
- Mizuno, A.; Tsukamoto, T.; Shimizu, Y.; Ooya, H.; Matsuura, T.; Saito, N.; Sato, S.; Kikuchi, S.; Uzuki, T.; Azegami, K. Occurrence of bacterial black shoot disease of European pear in Yamagata prefecture. J. Gen. Plant Pathol. 2010, 76, 43–51. [Google Scholar] [CrossRef]
- Rou-Zi, A.; Wang, Q. Bionomics of grapholitha molesta busck and its control measures. Biol. Disaster Sci. 2018, 41, 160–162. [Google Scholar] [CrossRef]
- Ishii, H.; Yanase, H. Venturia Nashicola, the scab fungus of Japanese and Chinese pears: A species distinct from V. pirina. Mycol. Res. 2000, 104, 755–759. [Google Scholar] [CrossRef]
- Ma, M.Z. Analysis of harmful symptoms and epidemic causes of pear scab. China Agric. Inf. 2014, 7, 100. [Google Scholar] [CrossRef]
- Gray, W.M. Hormonal regulation of plant growth and development. PLoS Biol. 2004, 2, E311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, Y.; Yamamoto, H.; Oritani, T. Biosynthesis of abscisic acid in the fungus Cercospora cruenta stimulation of biosynthesis by water stress and isolation of a transgenic mutant with reduced biosynthetic capacity. Plant Cell Physiol. 1995, 36, 557–564. [Google Scholar] [CrossRef]
- Pré, M.; Atallah, M.; Champion, A.; De Vos, M.; Pieterse, C.M.J.; Memelink, J. The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol. 2008, 147, 1347–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weingart, H.; Ullrich, H.; Geider, K.; Völksch, B. The role of ethylene production in virulence of Pseudomonas syringae pvs. glycinea and phaseolicola. Phytopathology 2001, 91, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef]
- Stintzi, A.; Weber, H.; Reymond, P.; Browse, J.; Farmer, E.E. Plant defense in the absence of jasmonic acid: The role of cyclopentenones. Proc. Natl. Acad. Sci. USA 2001, 98, 12837–12842. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, G.W.; Delaney, S.K. Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during development and abiotic stress. Mol. Genet. Genomics. 2010, 283, 233–241. [Google Scholar] [CrossRef]
- Kundu, A.; Patel, A.; Pal, A. Defining reference genes for qPCR normalization to study biotic and abiotic stress responses in Vigna mungo. Plant Cell Rep. 2013, 32, 1647–1658. [Google Scholar] [CrossRef]
- Wang, Y.J.; Dong, L.; Zhang, C.; Wang, X.Q. Reference gene selection for real-time quantitative PCR normalization in tree peony(Paeonia suffruticosa Andr). J. Agric. Biotechnol. 2012, 20, 521–528. [Google Scholar] [CrossRef]
- Yan, H.B.; Ge, W.Y.; Cheng, Y.D.; He, J.G.; Guan, J.F. Cloning and characters analysis of GAPDH gene family in Yali pear. J. Northwest AF Univ. (Nat. Sci. Ed.). 2012, 40, 181–186. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, X.; Gu, C.; Yin, H.; Zhang, S. Selection of reference genes in qRT-PCR of pear “Dangshansuli”. China Fruits 2018, 1, 16–22, 35. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, N.; Si, H.J.; Calderón-Urrea, A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods 2017, 13, 85. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qin, Z.L.; Wu, T.; Li, X.M.; Tu, J.F.; Yang, F.C.; Zhu, H.Y. Fluorescence Quantitative Reference Genes and Their Primers and Applications in Leaf Tissues of Two Tree Pears at Different Developmental Stages. CN107988407A. 2018. Available online: https://www.zhangqiaokeyan.com/patent-detail/061204415708.html (accessed on 6 November 2021).
- Abbas, A.; Yu, H.Y.; Li, X.J.; Cui, H.L.; Chen, J.C.; Huang, P. Selection and validation of reference genes for RT-qPCR analysis in Aegilops tauschii (Coss.) under different abiotic stresses. Int. J. Mol. Sci. 2021, 22, 11017. [Google Scholar] [CrossRef]
- Chen, J.Q.; Li, X.Y.; Wang, D.Q.; Li, L.T.; Zhou, H.S.; Liu, Z.; Wu, J.; Wang, P.; Jiang, X.T.; Fabrice, M.R.; et al. Identification and testing of reference genes for gene expression analysis in pollen of Pyrus bretschneideri. Sci. Hortic. 2015, 190, 43–56. [Google Scholar] [CrossRef]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.-R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Tian, C.; Song, Y. In vitro culture and rapid propagation of pear Xuehuali. Shanxi Agric. Univ. (Nat. Sci. Ed.). 2014, 34, 464–467. [Google Scholar] [CrossRef]
- Jaakola, L.; Pirttilä, A.M.; Halonen, M.; Hohtola, A. Isolation of high quality RNA from Bilberry (Vaccinium myrtillus L.) Fruit. Mol. Biotechnol. 2001, 19, 201–204. [Google Scholar] [CrossRef]
- Xie, F.L.; Xiao, P.; Chen, D.L.; Xu, L.; Zhang, B.H. miRDeepFinder:a miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. J. Genome Biol. 2002, 3, 34.1–34.11. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, J.T.; Yao, N. The role and regulation of programmed cell death in plant-pathogen interactions. Cell Microbiol. 2004, 6, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Sagi, M.; Fluhr, R. Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol. 2006, 141, 336–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepauskas, D.; Miliute, I.; Staniene, G.; Gelvonauskiene, D.; Stanys, V.; Jesaitis, A.J.; Baniulis, D. Characterization of apple NADPH oxidase genes and their expression associated with oxidative stress in shoot culture in vitro. Plant Cell Tissue Organ. Cult. 2015, 124, 621–633. [Google Scholar] [CrossRef]
- Cheng, X.; Li, G.H.; Manzoor, M.A.; Wang, H.; Abdullah, M.; Su, X.Q.; Zhang, J.Y.; Jiang, T.S.; Jin, Q.; Cai, Y.P.; et al. In silico genome-wide analysis of respiratory burst oxidase homolog (RBOH) family genes in five fruit-producing trees, and potential functional analysis on lignification of stone cells in Chinese White Pear. Cells 2019, 8, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.C.; Nadeau, K.; Abbasi, M.; Lachance, C.; Nguyen, M.; Fenrich, J. The ultimate qPCR experiment: Producing publication quality, reproducible data the first time. Trends Biotechnol. 2019, 37, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Mallona, I.; Lischewski, S.; Weiss, J.; Hause, B.; Egea-Cortines, M. Validation of Reference Genes for Quantitative Real-Time PCR during Leaf and Flower Development in Petunia Hybrida. BMC Plant Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Rapacz, M.; Stępień, A.; Skorupa, K. Internal standards for quantitative RT-PCR studies of gene expression under drought treatment in barley (Hordeum vulgare L.): The effects of developmental stage and leaf age. Acta Physiol. Plant. 2012, 34, 1723–1733. [Google Scholar] [CrossRef] [Green Version]
- Mafra, V.; Kubo, K.S.; Alves-Ferreira, M.; Ribeiro-Alves, M.; Stuart, R.M.; Boava, L.P.; Rodrigues, C.M.; Machado, M.A. Reference genes for accurate transcript normalization in Citrus genotypes under different experimental conditions. PLoS ONE 2012, 7, e31263. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Li, H.; Li, X.G.; Lin, J.; Wang, Z.H.; Yang, Q.S.; Chang, Y.H. Systematic selection and validation of appropriate reference genes for gene expression studies by quantitative real-time PCR in pear. Acta Physiol. Plant. 2015, 37, 40. [Google Scholar] [CrossRef]
- Liu, Z.; Ge, X.X.; Wu, X.M.; Kou, S.J.; Chai, L.J.; Guo, W.W. Selection and validation of suitable reference genes for mRNA qRT-PCR analysis using somatic embryogenic cultures, floral and vegetative tissues in Citrus. Plant Cell Tiss. Organ Cult. 2013, 113, 469–481. [Google Scholar] [CrossRef]
- Tong, Z.G.; Gao, Z.H.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariot, R.F.; de Oliveira, L.A.; Voorhuijzen, M.M.; Staats, M.; Hutten, R.C.B.; Van Dijk, J.P.; Kok, E.; Frazzon, J. Selection of Reference Genes for Transcriptional Analysis of Edible Tubers of Potato (Solanum tuberosum L.). PLoS ONE 2015, 10, e0120854. [Google Scholar] [CrossRef]
- Deng, Y.; Li, Y.D.; Sun, H.Y. Selection of Reference Genes for RT-qPCR Normalization in Blueberry (Vaccinium corymbosum × Angustifolium) under Various Abiotic Stresses. FEBS Open Bio 2020, 10, 1418–1435. [Google Scholar] [CrossRef]
- Pleskot, R.; Potocký, M.; Pejchar, P.; Linek, J.; Bezvoda, R.; Martinec, J.; Valentová, O.; Novotná, Z.; Žárský, V. Mutual Regulation of Plant Phospholipase D and the Actin Cytoskeleton. Plant J. 2010, 62, 494–507. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Dai, M.S.; Cai, D.Y.; Shi, Z.B. Expression stability analysis of common internal reference genes in pear fruit based on high-throughput sequencing. Mol. Plant Breed. 2019, 17, 746–753. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence ID | Amplicon Length (bp) | Annealing Tm (°C) | R2 | Amplification Efficiency (%) |
|---|---|---|---|---|---|
| ACT | GU830958 | 139 | 60 | 0.9999 | 104.39 |
| SKD1 | XM009366023 | 127 | 60 | 0.9992 | 106.66 |
| YLS8 | XM009353468 | 119 | 60 | 0.9993 | 111.02 |
| WDP | Pbr038544 | 85 | 60 | 0.9995 | 111.29 |
| TUB | Pbr042345 | 114 | 60 | 0.9949 | 111.62 |
| UBQ | Pbr008614 | 88 | 60 | 0.9977 | 108.20 |
| GAPDH | Pbr036263 | 118 | 60 | 0.9983 | 111.71 |
| EF1α | XM_009376302 | 91 | 60 | 0.9993 | 108.39 |
| Mock | SA | MeJA | ETH | ABA | |||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Value | Gene | Value | Gene | Value | Gene | Value | Gene | Value |
| Calculated in BestKeeper | |||||||||
| YLS8 | 0.17 | SKD1 | 0.91 | GAPDH | 0.36 | SKD1 | 0.37 | SKD1 | 0.76 |
| WDP | 0.45 | WDP | 1.08 | EF1α | 0.62 | WDP | 0.99 | TUB | 1.07 |
| SKD1 | 0.51 | YLS8 | 1.16 | ACT | 0.68 | TUB | 1.17 | WDP | 1.09 |
| TUB | 0.63 | ACT | 1.18 | TUB | 0.94 | YLS8 | 1.35 | YLS8 | 1.36 |
| ACT | 0.63 | TUB | 1.37 | UBQ | 0.95 | UBQ | 1.75 | UBQ | 1.63 |
| EF1α | 1.11 | EF1α | 1.54 | WDP | 1.08 | ACT | 2.00 | ACT | 2.00 |
| UBQ | 1.33 | UBQ | 1.68 | SKD1 | 1.47 | GAPDH | 2.39 | GAPDH | 2.44 |
| GAPDH | 1.40 | GAPDH | 1.82 | YLS8 | 1.50 | EF1α | 2.60 | EF1α | 2.77 |
| Calculated in NormFinder | |||||||||
| ACT | 0.22 | WDP | 0.09 | TUB | 0.33 | UBQ | 0.28 | ACT | 0.48 |
| WDP | 0.27 | ACT | 0.16 | ACT | 0.35 | YLS8 | 0.30 | YLS8 | 0.52 |
| EF1α | 0.56 | EF1α | 0.43 | UBQ | 0.44 | ACT | 0.43 | UBQ | 0.53 |
| TUB | 0.70 | TUB | 0.51 | WDP | 1.09 | WDP | 0.71 | WDP | 0.82 |
| YLS8 | 0.74 | UBQ | 0.68 | YLS8 | 1.09 | TUB | 0.77 | TUB | 0.87 |
| SKD1 | 0.77 | SKD1 | 0.84 | GAPDH | 1.27 | GAPDH | 1.05 | GAPDH | 1.18 |
| UBQ | 0.94 | YLS8 | 0.88 | SKD1 | 1.37 | SKD1 | 1.72 | SKD1 | 1.65 |
| GAPDH | 0.96 | GAPDH | 0.90 | EF1α | 1.44 | EF1α | 1.94 | EF1α | 1.89 |
| Calculated in GeNorm | |||||||||
| SKD1|TUB | 0.19 | ACT|WDP | 0.31 | ACT|UBQ | 0.69 | WDP|TUB | 0.40 | YLS8|WDP | 0.38 |
| WDP | 0.31 | TUB | 0.47 | TUB | 0.72 | YLS8 | 0.61 | TUB | 0.62 |
| YLS8 | 0.43 | EF1α | 0.56 | YLS8 | 0.86 | UBQ | 0.69 | UBQ | 0.87 |
| ACT | 0.52 | UBQ | 0.63 | SKD1 | 0.98 | ACT | 0.81 | ACT | 0.98 |
| EF1α | 0.68 | YLS8 | 0.72 | WDP | 1.08 | GAPDH | 0.94 | SKD1 | 1.09 |
| UBQ | 0.82 | SKD1 | 0.78 | GAPDH | 1.25 | SKD1 | 1.13 | GAPDH | 1.23 |
| GAPDH | 0.88 | GAPDH | 0.84 | EF1α | 1.34 | EF1α | 1.35 | EF1α | 1.42 |
| Mock | SA | MeJA | ETH | ABA | |||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Value | Gene | Value | Gene | Value | Gene | Value | Gene | Value |
| WDP | 1.86 | WDP | 1.19 | ACT | 1.57 | UBQ | 2.11 | YLS8 | 1.68 |
| ACT | 2.66 | ACT | 2.00 | TUB | 2.21 | WDP | 2.38 | WDP | 2.63 |
| TUB | 2.83 | EF1α | 3.83 | UBQ | 2.59 | YLS8 | 2.63 | ACT | 2.78 |
| SKD1 | 3.08 | TUB | 3.94 | GAPDH | 3.98 | TUB | 2.94 | TUB | 3.50 |
| YLS8 | 3.31 | SKD1 | 3.98 | YLS8 | 5.03 | ACT | 4.05 | UBQ | 3.66 |
| EF1α | 4.24 | UBQ | 5.44 | WDP | 5.18 | SKD1 | 4.30 | SKD1 | 4.14 |
| UBQ | 7.00 | YLS8 | 5.45 | EF1α | 5.66 | GAPDH | 6.24 | GAPDH | 6.48 |
| GAPDH | 8.00 | GAPDH | 8.00 | SKD1 | 6.44 | EF1α | 8.00 | EF1α | 8.00 |
| Cold | Heat | Water Deficit | Salinity | V. nashicola | |||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Value | Gene | Value | Gene | Value | Gene | Value | Gene | Value |
| Calculated using BestKeeper | |||||||||
| UBQ | 1.56 | UBQ | 1.56 | UBQ | 0.68 | SKD1 | 2.06 | SKD1 | 0.74 |
| GAPDH | 1.95 | GAPDH | 1.95 | GAPDH | 0.71 | GAPDH | 2.06 | YLS8 | 0.95 |
| SKD1 | 1.97 | SKD1 | 1.97 | TUB | 1.13 | ACT | 2.63 | WDP | 1.16 |
| EF1α | 2.06 | EF1α | 2.06 | EF1α | 1.30 | TUB | 2.97 | ACT | 1.33 |
| TUB | 2.30 | TUB | 2.30 | SKD1 | 1.34 | WDP | 2.99 | TUB | 1.46 |
| WDP | 2.43 | WDP | 2.43 | WDP | 1.46 | EF1α | 3.26 | UBQ | 1.59 |
| YLS8 | 2.68 | YLS8 | 2.68 | YLS8 | 1.54 | YLS8 | 3.33 | EF1α | 1.80 |
| ACT | 3.14 | ACT | 3.14 | ACT | 2.03 | UBQ | 4.01 | GAPDH | 2.14 |
| Calculated using NormFinder | |||||||||
| WDP | 0.11 | YLS8 | 0.67 | SKD1 | 0.09 | TUB | 0.01 | ACT | 0.15 |
| TUB | 0.32 | SKD1 | 0.92 | TUB | 0.18 | WDP | 0.01 | WDP | 0.42 |
| EF1α | 0.47 | UBQ | 0.96 | WDP | 0.29 | EF1α | 0.38 | TUB | 0.44 |
| SKD1 | 0.49 | WDP | 1.04 | YLS8 | 0.47 | YLS8 | 0.64 | EF1α | 0.55 |
| YLS8 | 0.60 | GAPDH | 1.05 | UBQ | 0.48 | ACT | 0.66 | UBQ | 0.72 |
| UBQ | 0.98 | ACT | 1.36 | EF1α | 0.62 | SKD1 | 1.24 | YLS8 | 0.85 |
| ACT | 1.34 | TUB | 1.56 | ACT | 1.33 | GAPDH | 1.28 | SKD1 | 1.06 |
| GAPDH | 1.63 | EF1α | 1.88 | GAPDH | 2.61 | UBQ | 1.63 | GAPDH | 1.41 |
| Calculated using GeNorm | |||||||||
| SKD1|EF1α | 0.14 | YLS8|WDP | 0.29 | YLS8|WDP | 0.10 | WDP|TUB | 0.03 | ACT|WDP | 0.30 |
| TUB | 0.29 | GAPDH | 0.91 | SKD1 | 0.19 | EF1α | 0.26 | TUB | 0.40 |
| WDP | 0.37 | SKD1 | 1.00 | TUB | 0.32 | YLS8 | 0.41 | EF1α | 0.57 |
| YLS8 | 0.48 | ACT | 1.17 | EF1α | 0.44 | ACT | 0.60 | UBQ | 0.70 |
| UBQ | 0.66 | UBQ | 1.27 | UBQ | 0.61 | SKD1 | 0.83 | YLS8 | 0.78 |
| ACT | 0.89 | TUB | 1.45 | ACT | 0.74 | GAPDH | 0.95 | SKD1 | 0.87 |
| GAPDH | 1.11 | EF1α | 1.63 | GAPDH | 1.22 | UBQ | 1.13 | GAPDH | 1.02 |
| Cold | Heat | Water Deficit | Salinity | V. nashicola | |||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Value | Gene | Value | Gene | Value | Gene | Value | Gene | Value |
| WDP | 2.21 | YLS8 | 1.50 | SKD1 | 1.97 | TUB | 1.68 | ACT | 1.41 |
| EF1α | 2.45 | SKD1 | 2.38 | WDP | 2.45 | WDP | 1.78 | WDP | 1.86 |
| SKD1 | 2.63 | WDP | 3.13 | TUB | 2.91 | EF1α | 3.57 | TUB | 3.41 |
| TUB | 2.78 | GAPDH | 3.41 | YLS8 | 3.25 | SKD1 | 3.83 | SKD1 | 4.30 |
| UBQ | 3.83 | ACT | 3.66 | UBQ | 3.66 | ACT | 4.40 | YLS8 | 4.36 |
| YLS8 | 5.44 | UBQ | 4.36 | EF1α | 4.95 | YLS8 | 4.60 | EF1α | 4.60 |
| GAPDH | 5.66 | TUB | 7.00 | GAPDH | 5.66 | GAPDH | 5.12 | UBQ | 5.48 |
| ACT | 7.24 | EF1α | 8.00 | ACT | 7.24 | UBQ | 8.00 | GAPDH | 8.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, P.; Huang, L.; Wang, Y.; Li, X.; Feng, X.; Li, L. Stepwise Optimization of the RT-qPCR Protocol and the Evaluation of Housekeeping Genes in Pears (Pyrus bretschneideri) under Various Hormone Treatments and Stresses. Horticulturae 2023, 9, 275. https://doi.org/10.3390/horticulturae9020275
Zhou P, Huang L, Wang Y, Li X, Feng X, Li L. Stepwise Optimization of the RT-qPCR Protocol and the Evaluation of Housekeeping Genes in Pears (Pyrus bretschneideri) under Various Hormone Treatments and Stresses. Horticulturae. 2023; 9(2):275. https://doi.org/10.3390/horticulturae9020275
Chicago/Turabian StyleZhou, Peng, Linlin Huang, Yingtao Wang, Xiao Li, Xinxin Feng, and Liulin Li. 2023. "Stepwise Optimization of the RT-qPCR Protocol and the Evaluation of Housekeeping Genes in Pears (Pyrus bretschneideri) under Various Hormone Treatments and Stresses" Horticulturae 9, no. 2: 275. https://doi.org/10.3390/horticulturae9020275