

Review of the Current Research Progress of Seed Germination Inhibitors
1
College of Forestry, Nanjing Forestry University, Nanjing 210037, China
2
Co-Innovation Center for Sustainable Forestry in Southern China, Southern Tree Inspection Center National Forestry Administration, Nanjing 210037, China
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(4), 462; https://doi.org/10.3390/horticulturae9040462
Submission received: 25 February 2023
/
Revised: 22 March 2023
/
Accepted: 4 April 2023
/
Published: 5 April 2023
(This article belongs to the Collection Seed Dormancy and Germination of Horticultural Plants)
Abstract
:Germination inhibitors, which inhibit the germination of seeds, spores and other plant reproductive material, are abundant in the plant kingdom and include phenols, cyanides, alkaloids, essential oils, amino acids, etc. These inhibitors can be classified as germination destructors and germination retarders depending on whether they harm the morphology, structure and physiology of the seed. Germination retarders are closely related to seed dormancy, and exogenous retarders can be used to extend the “dormancy” period of non-dormant seeds or perishable seeds by applying the proper dosage. They have significant potential applications as preservatives for seed preservation following harvest or for the storage of long-term germplasm resources. Germination destructors, as a type of relatively high-efficiency, low-specificity “toxic chemicals”, are of significant benefit in the application of effective and environmentally benign herbicides. At present, the main problems related to the research methods of germination inhibitors include difficulty in determining the specific endogenous substances and the minimum inhibitory concentration to induce dormancy, as well as whether the application of exogenous inhibitors will cause physiological damage to seeds. In the future, we should strengthen the tracking of germination inhibitors, explore the mechanisms of action of specific substances and deeper molecular mechanisms and finally explore new developments and new applications of different inhibitors.
1. Introduction
Seeds are the foundation of agroforestry, the beginning of plant growth and development, and also the key link in their life cycle [1]. Seed germination is the process by which the metabolism of plant seeds is stimulated after hydration and the radicle breaks through the seed covering [2,3]. During this process, there are hormones that stimulate seed germination, such as gibberellin [4,5], and there are also hormones that inhibit seed germination, such as abscisic acid, which regulates seed dormancy [6,7,8]. In addition, certain chemicals present in different parts of the fruit and seed, including the fleshy and non-fleshy pericarp, endosperm, coat and seed embryo, also inhibit seed germination [9]. The presence of germination inhibitors is one of the main causes of seed dormancy, especially in physiologically dormant (PD) seeds [10,11].
Plant-produced germination inhibitors are closely associated with dormancy [12]. Thus, comprehensive research on germination inhibitors is crucial for crop production techniques, the conservation of germplasm resources and the protection and restoration of endangered populations. At present, although the existence of some germination inhibitors is known, their systematic and in-depth study is still in its early stages. This paper proposes a classification for germination inhibitors based on current research progress and summarizes related research problems from the qualitative, quantitative and experimental method aspects of germination inhibitors in order to serve as references for related research in the field of seed science.
2. Definition of Germination Inhibitors
Germination inhibitors are substances found in plants that can delay seeds germinating. Evenari [13] claimed that it is not possible to determine whether these substances only hinder growth without harming plants, so germination inhibitors are generally referred to as any substance that can inhibit seed germination. These germination inhibitors are not only found in angiosperms; butylated hydroxytoluene (BHT) may be present in the endosperm and embryo of Pistacia chinensis seeds [14], which can inhibit seed germination and cotyledon elongation. At the same time, it is also abundant in gymnosperm seeds, cryptophores and other reproductive material; for example, fenugreek spores do not germinate in its spore pool [13,15]. Bingöl et al. [16] found that the ethanol extract of gyelnik (Xanthoparmelia somloensis) had inhibitory effects on tomato seed germination. With the in-depth study of germination inhibitors, it has gradually been found that some non-plant substances also inhibit seed germination. For example, trimethylbutenolide (TMB), which is a smoke-derived trimethylbutenolactone [17], can significantly inhibit the germination of five weed seeds, including fleabane, hairy wild lettuce, bugweed, spilanthes and fameflower [18]. The large-scale synthesis and production of TMB has been achieved [19], extending the traditional range of seed germination inhibitors.
3. Classification of Germination Inhibitors
3.1. Classification by Function
Kockemann [20] believed that germination inhibitors could be divided into two types, one being substances found in seeds which easily fail under the action of light and other physical and chemical factors, and the other being substances found in the flesh of the fruit whose physical and chemical properties are not well understood. As the scope of research on germination inhibitors continues to expand, so does the number of chemicals, including but not limited to hydrogen cyanide, ammonia, ethylene, mustard oil, organic acids, unsaturated lactones, aldehydes, aromatic oils and alkaloids. Germination inhibitors can be divided into endogenous and exogenous inhibitors according to their mode of production and location. Evenari [13] believed that endogenous germination inhibitors are produced and used by the plants themselves, while those produced by other plants or seeds are exogenous. However, with the expansion of traditional methods of obtaining inhibitors and the development of synthetic chemistry, the scope of exogenous inhibitors was also expanded to include all inhibitors not produced by the plant itself.
Germination inhibitors can be divided into two categories according to whether they cause irreversible damage to seed germination: (1) Germination retarders, which can delay the germination of seeds, but the seeds can recover the ability to germinate under certain conditions; and (2) germination destructors, which damage the physiological activity of seeds and cause them to gradually lose their ability to germinate.
Most germination retarders are natural compounds produced by dormant seeds under natural conditions, such as ABA and 1,2,3-benzenetriol, which are produced in Cercis chinensis seeds and can regulate their dormancy [21]. After soaking in warm water, applying gibberellin or low-temperature stratification, the germination inhibitor content of these seeds was reduced or weakened and the normal germination ability could be restored [22,23]. For example, coumarin reduces reactive oxygen species (ROS) accumulation by specifically enhancing SOD and POD enzyme activities while promoting ABA synthesis to inhibit the germination of Brassica parachinensis and Oryza sativa, etc., but this germination inhibition can be recovered by adding exogenous GA4 and GA4+7 [24,25]. Germination retarders are closely related to seed dormancy, and exogenous retarders can be used to prolong the “dormancy” time of non-dormant seeds or perishable seeds. It has important application prospects as a preservative for seed preservation after harvest or long-term storage of germplasm resources.
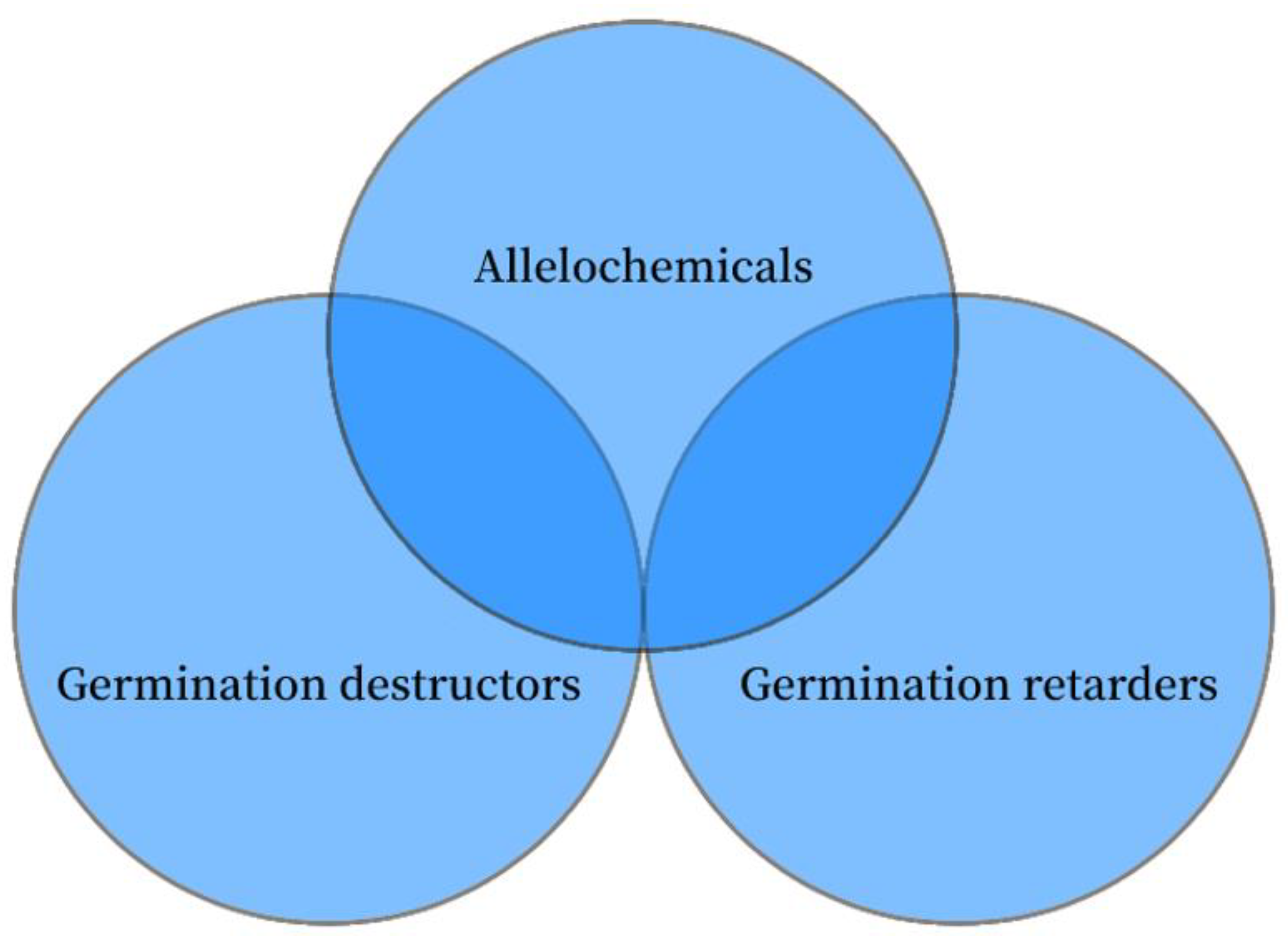
Germination destructors are mostly harmful substances that can injure seeds. For instance, after treating mustard seed with sunflower leaf extract, the seed’s H2O2 level significantly increased, ROS accumulation increased, and the cell membrane was damaged, ultimately leading to the seed’s death [26]. Unselective and selective germination destructors are additional classifications for germination destructors. Unselective germination destructors that are hazardous to seeds in all situations, such as a poisonous chemical; by inducing the production of a large number of ROS, these substances destroy the structure of cell membranes, leading to oxidative stress [27], damage macromolecules including lipids, proteins and DNA [28], and ultimately cause irreparable damage to seeds and plant morphogenesis. Selective germination destructors (Table 1) obstruct the growth and development of particular plant seeds, such as secondary metabolites produced by allelopathy, that may influence the seed activity of other plants. Lawrence et al. [29] demonstrated that leaf and stem extracts of Ailanthus inhibited seed germination and seedling growth in eight native North American plants, but had no self-toxic effects. Bauer et al. [30] found that Lonicera maackii leaf extract inhibited the germination of seeds of the herbs Bredia hirsuta, Arabidopsis thaliana, Alliaria petiolata and Impatiens capensis without harming its own seeds. Selective germination destructors may also have no germination inhibition effect on the seed germination of other plants. Water extracts from Solidago canadensis roots could damage the structure and function of Kummerowia striata seeds, but had no significant effect on seed germination of Lactuca sativa and Raphanus sativus [31,32]. Efficient germination destructors have little impact on the ecological environment due to their specific inhibitory effect and the degradability of most substances, which are of great significance in the application of new environmental protection and efficient herbicides. For example, natural secondary metabolites strigolactones (SLs) inhibited the seed germination of obligate hemiparasitic weed Striga hermonthica without any effect on the host plant [33]. However, not all allelopathic compounds inhibit the growth and development of other seeds, and other allelopathic substances just delay the germination process of seeds [34]; the effect of such allelopathic substances is similar to that of germination retarders (Figure 1). The extract from the leaves of Rhododendron maximum, Kalmia latifolia and Lonicera maackii had no significant effect on the final germination rate of Festuca arundinacea seeds, but the time to reach the maximum seed germination rate was delayed by up to 4 days, which significantly delayed the seed germination process [35,36].
3.2. Classification by Chemical Structure
Germination inhibitors can be divided into the following types according to their chemical structure (Table 2).
3.2.1. Phenols
Phenolic compounds are secondary metabolites that are ubiquitous in plant tissues [52] and are one of the most common allelopathic substances in plants [53]. Li et al. [54] found that phenolic compounds (cinnamic acid) and their related phenolic derivatives (trans-cinnamic acid, coumarin, chlorogenic acid and ferulic acid) had a significant inhibitory effect on the germination of lettuce seeds. Williams and Hoagland [37] found that phenolic compounds (caffeic acid, chlorogenic acid, coumarin, p-coumarin, ferulic acid, fumaric acid, gallic acid, hydrocinnamic acid, p-hydroxybenzoic acid, juglone and catechol) had a significant inhibitory effect on the germination of lettuce seeds, walnut and catechol) had inhibitory effects on the germination of crop (cotton, melon, maize and sorghum) and weed (hemp sesbania, sicklepod, velvetleaf, prickly sida and redroot pigweed) seeds. Lignans are a polyphenolic compound found in some plants; Cutillo [55] found that lignans identified from toxic extracts of Brassica had a significant inhibitory effect on the germination of lettuce seeds, and still had inhibitory activity at extremely low concentrations (1 nmol/L). Ng et al. [56] found that cinnamic acid and benzoic acid, together with some derivatives, had an inhibitory effect on the germination of Brassica napus seeds. Radwan et al. [57] found that phenolic compounds and flavonoids significantly reduced the germination rate, radicle length and germ length of wheat seeds. Chiji et al. [51] found that two phenolamides isolated from beet seed bulbs had a significant inhibitory effect on lettuce seed germination. It should also be noted that most phenolic substances have a combined inhibitory effect, and the degree of inhibition of seed germination is greater when used in combination than when used alone [37]. Einhellig and Rasmussen [58] found that vanillic acid and parahydroxybenzoic acid have inhibitory effects on radish and sorghum seed germination, that the two can inhibit synergistically and that they are more effective than when used alone. It has also been found that the combination of phenolic compounds and ABA also has a stronger inhibitory effect on seedling growth than monophenolic compounds or ABA alone [54], and that this effect is related to the concentration of phenolic substances [41]. The inhibitory mechanisms of different phenolic compounds on seed germination are different. Tannic acid, coumaric acid and vanillic acid affected the growth of mung bean hypocotyl and inhibited mitochondrial metabolic activities. Vanillic acid only inhibits mitochondrial Ca2+ transport and does not affect respiration or oxidative phosphorylation [59].
3.2.2. Alkaloids
Alkaloids are a class of structurally diverse secondary compounds widely distributed in plants [60]. Scopolamine and hyoscyamine in alkaloids largely inhibited the germination and early growth of flax and sunflower seeds [61,62]. Wink [50] found that quinicidine alkaloids inhibited the germination of lettuce (Lactuca sativa) seeds, and lupin alkaloids at a concentration of 6 mM inhibited lettuce seed germination by 100%. Aerts et al. [63] found that quinoline alkaloids at a concentration of 0.3 mM had a significant inhibitory effect on seed germination of Ocimum, Spermacoce, Catharanthus and Cinchona.
3.2.3. Essential Oils
Essential oils are products of plant metabolism, of which terpenoids, especially monoterpenes and sesquiterpenes, are the major constituents of essential oils and can inhibit plant growth activity [64]. Singh et al. [65] found that volatile oils in eucalyptus had a complete inhibitory effect on the seed germination of Parthenium hysterophorus. The inhibition of plant seed germination by essential oils is universal; Ramezani et al. [66] extracted essential oils from Eucalyptus nicholii, Eucalyptus nicholii, Chamaecyparis lowsoniana and Chamaecyparis lowsoniana, which showed significant inhibition on the germination of weed seeds. Benvenuti et al. [67] extracted essential oils from 20 species of Asteraceae which showed certain inhibition on the germination of Amaranthus retroflexus seeds.
3.2.4. Other Types
High concentrations of ‘unusual’ amino acids are found in some seeds, such as Fabaceae seeds. S-Hydroxy-L-tryptophan makes up 14% of the seed weight of Griffonia simplicifolia [68] and canucine (a non-protein amino acid) makes up 7–10% of the seed weight of Dioclea megacarpa [69], L3,4-dihydroxyphenylalanine makes up more than 8% of the seed weight of Mucuna mutisiana [70]; these amino acids may have some inhibitory effect on seed germination. Elmore [71] found that alkaline components (amino acids) extracted from velvetleaf seeds inhibited radicle growth in 18% of beet seeds after 48 h. A study by Wilson and Bell [72] found that non-protein amino acids were more effective than protein amino acids in inhibiting lettuce seed germination and seedling growth, with the exception of lysine. Vurro et al. [73] found that a 2 mM concentration of methionine almost completely inhibited the germination of Orobanche ramosa seeds.
Niacinamide is a precursor of vitamin B3 and has a variety of biological effects. In plants, nicotinamide has been shown to reduce H2O2-induced cell death [74] and inhibit ABA-induced stomatal closure [75], but the opposite effect was found in seeds. Zheng et al. [76] found that the application of nicotinamide inhibited the growth of mung bean embryo growth, and Hunt et al. [77] found that nicotinamide inhibited Arabidopsis seed germination and that nicotinamide metabolism is required to break seed dormancy. In addition, some compounds have also been found to inhibit germination; Oster et al. [78] found that lactone compounds extracted from Sorbus pohuashanensis fruits and seeds at concentrations ≥5 × 10−4 mnL inhibited the germination of Amaranthus caudatus and Lepidium sativum seeds. Levi-Minzi et al. [79] found that butyric acid in fatty acids has a significant inhibitory effect on wheat seed germination, and the inhibitory effect increases with increasing butyric acid concentration. Uygur [80] found that mustard oil inhibited the germination of Centaurea solstitialis seeds.
4. The Core Problem of Germination Inhibitors Research
4.1. Determination of Dormant Seed Germination Inhibitors
Germination inhibitors are present in all parts of the plant: pulp, peel, endosperm, seed coat, embryo, leaf, bulb and root [13]. Germination inhibitors can inhibit seed germination in one or more plants; however, it is difficult to determine the specific type of germination inhibitor from dormant seeds, and most studies have only been able to determine the presence of germination inhibitors in dormant seeds. Bian et al. [81] found that Taxus yunnanensis seed extract contained germination inhibitors and had a significant inhibitory effect on Brassia campestris seeds, but the specific endogenous substances were difficult to determine. At the same time, the specific substances that inhibit germination cannot be determined only by the changes in the content of endogenous substances during the process of seed dormancy removal. Some germination-inhibiting substances have a small and rapid effect, and there are also some relatively large and rapidly changing substances that cannot regulate germination. Complex physiological and biochemical processes are involved in the process of seed dormancy release; once the key factors for dormancy release have been identified, the external conditions such as temperature, water and oxygen required for seed germination must be met [2]. At this point, changes in water uptake rates, reactivation of metabolic reactions, transformation of storage materials and seedling development and growth occur, involving a large number of substance metabolisms and transformations [82]. In addition, indirect metabolic changes can also block seed germination; for example, the application of paclobutrazole can inhibit seed germination by inhibiting gibberellin synthesis [83]. Seeds are sensitive to environmental changes and can regulate their dormancy in response to a number of environmental factors [84], making it difficult to determine which substance is directly or indirectly causing the disruption or delay in germination pathways in the current study.
In related studies, after determining the existence of germination inhibitors in the seeds of a given plant, the system solvent method was used to extract the contents of the all or part of the seed tissues (such as endosperm, seed coat, etc.), and then petroleum ether phase, diethyl ether phase, methanol phase and aqueous phase were separated (Figure 2) [85], followed by the analysis of the main components of each phase by gas chromatography–mass spectrometry (GC-MS) and high-performance liquid chromatography (HPLC), the germination inhibitors were inferred according to the changes in the content of each component. However, this method also has major limitations and can only be used for speculation. The release of physiological dormancy in seeds undergoes complex physiological and biochemical changes, which is a systematic process accompanied by the metabolism and transformation of various substances [86]. Therefore, it is difficult to identify germination inhibitors based solely on the reduction in substance content. On the other hand, dormant seeds can germinate after the removal of peel or seed coat tissues, and the presence of endogenous inhibitors in the removed tissues does not necessarily mean that seed dormancy is caused by chemical inhibitors in the peel or seed coat [87]. Many seeds and fruits containing endogenous inhibitors may not be dormant [9].
4.2. Concentration of Germination Inhibitors
The problem of concentration is an unavoidable key issue in the study of chemistry and biology, and the application of certain substances at different concentrations has a major impact on the results of research. For example, cyanide has a bidirectional effect on seed germination. It can enhance seed germination in some plants [88], treatment of dormant apple seeds with HCN can improve their germination [89] and cyanide also enhances the germination of dormant rice and barley seeds [90]; this enhancement may be due to the ROS compounds produced [91]. However, the inhibitory effect of cyanide is mainly related to its compound concentration, mainly showing a low promoting and high suppressing effect [92]; high concentrations of cyanide will inhibit seed germination [93]. When studying the concentration of germination inhibitors, the most important core element is to find the lowest endogenous inhibitor concentration that causes seed dormancy. Ultraviolet spectrophotometry can be used for the quantitative determination of organic compounds such as phenols [87,94]. Different seeds have different concentration requirements for dormancy caused by germination inhibitors, which requires the detection of the minimum concentration threshold of different inhibitors that cause dormancy in different seeds. From a practical point of view, the most valuable germination inhibitors are those that do not damage the seed but can prevent or delay germination. Quantity is the key problem that cannot be avoided in the production and use of germination inhibitors.
4.3. Examination of Germination Inhibitors
The presence of germination inhibitors can be verified by diluting the extracted inhibitors and applying them to non-dormant seeds such as cabbage, wheat and mung bean [95,96]. Cutillo et al. [55] found that Brassica fruticulosa methanol aqueous extract (10 mg/L) had a germination inhibition rate of 50% on Lactuca sativa seeds, which is a common method of verifying the presence of germination inhibitors in dormant seeds. However, this method has certain disadvantages: the first is the relationship between the concentration of diluted inhibitors and non-dormant seed germination; during the dormancy process, the seed itself is equivalent to an “environmental sensor”, which will adjust its dormant state according to a number of environmental factors [84], so even non-toxic substances at high concentrations can cause seeds not to germinate, and determining the minimum concentration to cause dormancy cannot be verified. In addition, whether the application of dormant seed extracts to non-dormant seeds will cause irreversible physiological damage requires the germination test of ungerminated seeds to be regerminated after removal of the inhibitors to assess the damage of the germination inhibitor. These issues are often overlooked in existing studies, leading to erroneous or unreliable conclusions, such as the use of cabbage seeds as biological materials to test seed dormancy, as in Daphne giraldii [97] and Quercus et al. [98], which did not validate the minimum concentration of germination inhibitor in seeds that was harmful or detrimental to the germination of cabbage seeds. The harmlessness of endogenous inhibitors should be an important criterion for such tests.
5. Research Prospects of Germination Inhibitors
5.1. Determine the Relationship between Germination Inhibitors and Inhibition of Germination
At present, most of the research on germination inhibitors is still focused on the last century, and a specific theoretical system and research methods have not yet been formed; some studies show difficulty in determining types of germination inhibitors after confirming the existence of germination inhibitors and their inhibitory effects. Some germination inhibitors are specific; there may be different inhibition effects on the germination of different seeds, or even no inhibition phenomenon. For different types of dormant seeds, the general inhibition of one or more well-defined germination inhibitors needs to be verified.
5.2. Tests and Examination of Germination Inhibitors
Having identified the specific germination inhibitors, it is necessary to carry out more in-depth research into the characteristics and targets of germination inhibitors to further determine the inhibition concentration and whether exogenous substances will cause physiological damage to seeds, ultimately achieving the important purpose of controlling seed dormancy and germination.
5.3. Study on the Mechanism of Action of Germination Inhibitors
Currently, different omics methods (e.g., proteomics) have been used to study various seed germination inhibitors, such as jasmonic acid (JA), methyl jasmonate (MeJA) or 12-oxophytodienoic acid (OPDA), which promote the interaction between COI1 and JAZ1 proteins, resulting in the ubiquitin-dependent degradation of JAZ1, thereby inhibiting the transcription of JA response genes [99] and ultimately inhibiting seed germination in plants such as Solanum lycopersicum, Brassica napus and Linum usitatissimum [100,101]. In addition, the mechanism of ABA inhibition of germination has been further elaborated using different methods [102]; while the mechanism of action of most germination inhibitors is still unclear, the main mechanisms include the inhibition of the electron transport system, the inhibition of seed swelling, the inhibition of translation and transcription, etc. [103]. Different germination inhibitors have different mechanisms of action, and omics methods are still needed to explore the specific inhibitory mechanisms of different substances and the deeper molecular regulatory mechanisms.
Author Contributions
P.C. and S.Y. conceived and determined the article framework; P.C. and W.Y. wrote the article; W.Y. provided a literature supplement; S.Y. and S.F. reviewed and edited the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Jiangsu Forestry Science and Technology Innovation and Promotion Project (LYKJ [2021] 03) and the Key R&D project of Shandong Province (2021LZGC023).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data generated or analysed during this research are included in this published article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- El-Maarouf-Bouteau, H.; Sajjad, Y.; Bazin, J.; Langlade, N.; Cristescu, S.M.; Balzergue, S.; Baudouin, E.; Bailly, C. Reactive oxygen species, abscisic acid and ethylene interact to regulate sunflower seed germination. Plant Cell Environ. 2015, 38, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Black, M.; Bewley, J.D.; Black, M. Seeds: Germination, structure, and composition. Seeds Physiol. Dev. Germination 1994, 1–33. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Physiology and Biochemistry of Seeds in Relation to Germination: 1 Development, Germination, and Growth; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Jacobsen, J.; Beach, L. Control of transcription of α-amylase and rRNA genes in barley aleurone protoplasts by gibberellin and abscisic acid. Nature 1985, 316, 275–277. [Google Scholar] [CrossRef]
- Jones, R.L.; Jacobsen, J.V. Regulation of synthesis and transport of secreted proteins in cereal aleurone. Int. Rev. Cytol. 1991, 126, 49–88. [Google Scholar]
- Weyers, J.D.; Paterson, N.W. Plant hormones and the control of physiological processes. New Phytol. 2001, 152, 375–407. [Google Scholar] [CrossRef] [Green Version]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Liu, X.-D.; Xie, Q.; He, Z.-H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Wareing, P. Endogenous inhibitors in seed germination and dormancy. In Differenzierung und Entwicklung/Differentiation and Development; Springer: Berlin/Heidelberg, Germany, 1965; pp. 2556–2571. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. Some considerations for adoption of Nikolaeva’s formula system into seed dormancy classification. Seed Sci. Res. 2008, 18, 131–137. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; Rubio de Casas, R.; NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef] [Green Version]
- Evenari, M. Germination inhibitors. Bot. Rev. 1949, 15, 153–194. [Google Scholar] [CrossRef]
- Guo, H.; Liu, Y.; Wang, H.; Li, S. Study on the dormancy characteristics of Chinese pistache (Pistacia chinensis Bunge) seeds. Forests 2022, 13, 1521. [Google Scholar] [CrossRef]
- Oppenheimer, H. The absence of germination in the containers of the mother plant. Imperial Akad Wiss 1922, 131, 279–312. [Google Scholar]
- Bingöl, Ö.; Battal, A.; Aslan, A.; Emre, E. Investigation of the allelopathic effects of lyophilized ethanol extract of Xanthoparmelia somloensis (Gyelnik) Hale lichen on tomato plant. Anatol. J. Bot. 2022, 6, 39–43. [Google Scholar]
- Light, M.E.; Burger, B.V.; Staerk, D.; Kohout, L.; Van Staden, J. Butenolides from plant-derived smoke: Natural plant-growth regulators with antagonistic actions on seed germination. J. Nat. Prod. 2010, 73, 267–269. [Google Scholar] [CrossRef] [Green Version]
- Papenfus, H.B.; Kulkarni, M.G.; Pošta, M.; Finnie, J.F.; Van Staden, J. Smoke-isolated trimethylbutenolide inhibits seed germination of different weed species by reducing amylase activity. Weed Sci. 2015, 63, 312–320. [Google Scholar] [CrossRef]
- Surmont, R.; Verniest, G.; De Kimpe, N. Short synthesis of the seed germination inhibitor 3, 4, 5-trimethyl-2(5H)-furanone. J. Org. Chem. 2010, 75, 5750–5753. [Google Scholar] [CrossRef]
- Kockemann, A. About an anti-germination substance in fleshy fruits. Ber. Deut. Bot. Ges. 1934, 52, 523–526. [Google Scholar]
- Gao, Y.; Zhu, M.; Wang, H.; Li, S. Dynamic Changes to Endogenous Germination Inhibitors in Cercis chinensis Seeds during Dormancy Release. HortScience 2021, 56, 557–562. [Google Scholar] [CrossRef]
- Ke, B.Y.; Zhou, C.Y.; Li, J.Y.; Pan, J.; Zhou, P. Study on Methods of Seed Dormancy and Dormancy-release for Rare Species Acrocarpus fraxinifolius. Seed 2020, 39, 139–143. [Google Scholar]
- Song, J.; Zhang, H.; Zhang, Y.; Yue, H. Effect of Stratification after lmmersion of Gibberellin on Seed Germination of Plagiorhegma dubia Maxim. J. Northeast For. Univ. 2021, 12, 34–39. [Google Scholar]
- Chen, B.-X.; Peng, Y.-X.; Gao, J.-D.; Zhang, Q.; Liu, Q.-J.; Fu, H.; Liu, J. Coumarin-induced delay of rice seed germination is mediated by suppression of abscisic acid catabolism and reactive oxygen species production. Front. Plant Sci. 2019, 828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.-X.; Peng, Y.-X.; Yang, X.-Q.; Liu, J. Delayed germination of Brassica parachinensis seeds by coumarin involves decreased GA4 production and a consequent reduction of ROS accumulation. Seed Sci. Res. 2021, 31, 224–235. [Google Scholar] [CrossRef]
- Oracz, K.; Bailly, C.; Gniazdowska, A.; Côme, D.; Corbineau, F.; Bogatek, R. Induction of oxidative stress by sunflower phytotoxins in germinating mustard seeds. J. Chem. Ecol. 2007, 33, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuypers, A.; Keunen, E.; Bohler, S.; Jozefczak, M.; Opdenakker, K.; Gielen, H.; Vercampt, H.; Bielen, A.; Schellingen, K.; Vangronsveld, J. Cadmium and copper stress induce a cellular oxidative challenge leading to damage versus signalling. In Metal Toxicity in Plants: Perception, Signaling and Remediation; Springer: Berlin/Heidelberg, Germany, 2011; pp. 65–90. [Google Scholar]
- Huybrechts, M.; Cuypers, A.; Deckers, J.; Iven, V.; Vandionant, S.; Jozefczak, M.; Hendrix, S. Cadmium and plant development: An agony from seed to seed. Int. J. Mol. Sci. 2019, 20, 3971. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, J.G.; Colwell, A.; Sexton, O.J. The ecological impact of allelopathy in Ailanthus altissima (Simaroubaceae). Am. J. Bot. 1991, 78, 948–958. [Google Scholar] [CrossRef]
- Bauer, J.T.; Shannon, S.M.; Stoops, R.E.; Reynolds, H.L. Context dependency of the allelopathic effects of Lonicera maackii on seed germination. Plant Ecol. 2012, 213, 1907–1916. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, B.; Zhang, S.; Tang, J.; Tu, C.; Hu, S.; Yong, J.W.H.; Chen, X. Enhanced allelopathy and competitive ability of invasive plant Solidago canadensis in its introduced range. J. Plant Ecol. 2012, 6, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Butcko, V.M.; Jensen, R.J. Evidence of tissue-specific allelopathic activity in Euthamia graminifolia and Solidago canadensis (Asteraceae). Am. Midl. Nat. 2002, 148, 253–262. [Google Scholar] [CrossRef]
- Zwanenburg, B.; Blanco-Ania, D. Strigolactones: New plant hormones in the spotlight. J. Exp. Bot. 2018, 69, 2205–2218. [Google Scholar] [CrossRef]
- Dorning, M.; Cipollini, D. Leaf and root extracts of the invasive shrub, Lonicera maackii, inhibit seed germination of three herbs with no autotoxic effects. Plant Ecol. 2006, 184, 287–296. [Google Scholar] [CrossRef]
- Koocheki, A.; Lalegani, B.; Hosseini, S. Ecological consequences of allelopathy. In Allelopathy: Current Trends and Future Applications; Springer: Berlin/Heidelberg, Germany, 2013; pp. 23–38. [Google Scholar]
- McEwan, R.W.; Arthur-Paratley, L.G.; Rieske, L.K.; Arthur, M.A. A multi-assay comparison of seed germination inhibition by Lonicera maackii and co-occurring native shrubs. Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 475–483. [Google Scholar] [CrossRef]
- Williams, R.D.; Hoagland, R.E. The effects of naturally occurring phenolic compounds on seed germination. Weed Sci. 1982, 30, 206–212. [Google Scholar] [CrossRef]
- Miller, H.G.; Ikawa, M.; Peirce, L.C. Caffeic acid identified as an inhibitory compound in asparagus root filtrate. HortScience 1991, 26, 1525–1527. [Google Scholar] [CrossRef]
- Li, X.; Gruber, M.Y.; Hegedus, D.D.; Lydiate, D.J.; Gao, M.-J. Effects of a coumarin derivative, 4-methylumbelliferone, on seed germination and seedling establishment in Arabidopsis. J. Chem. Ecol. 2011, 37, 880–890. [Google Scholar] [CrossRef]
- Muzaffar, S.; Ali, B.; Wani, N.A. Effect of catechol, gallic acid and pyrogallic acid on the germination, seedling growth and the level of endogenous phenolics in cucumber (Cucumis sativus L.). Inter J. Life Sci. Biotechnol. Pharma. Res. 2012, 1, 50–55. [Google Scholar]
- Reigosa, M.; Souto, X.; Gonz´lez, L. Effect of phenolic compounds on the germination of six weeds species. Plant Growth Regul. 1999, 28, 83–88. [Google Scholar] [CrossRef]
- Shirley, B.W. Flavonoids in seeds and grains: Physiological function, agronomic importance and the genetics of biosynthesis. Seed Sci. Res. 1998, 8, 415–422. [Google Scholar] [CrossRef]
- Jia, L.; Wu, Q.; Ye, N.; Liu, R.; Shi, L.; Xu, W.; Zhi, H.; Rahman, A.R.B.; Xia, Y.; Zhang, J. Proanthocyanidins inhibit seed germination by maintaining a high level of abscisic acid in Arabidopsis thaliana F. J. Integr. Plant Biol. 2012, 54, 663–673. [Google Scholar] [CrossRef]
- Samanta, A.; Das, G.; Das, S.K. Roles of flavonoids in plants. Carbon 2011, 100, 12–35. [Google Scholar]
- Chotsaeng, N.; Laosinwattana, C.; Charoenying, P. Inhibitory Effects of a Variety of Aldehydes on Amaranthus tricolor L. and Echinochloa crus-galli (L.) Beauv. Molecules 2018, 23, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradow, J.M.; Connick, W.J. Volatile seed germination inhibitors from plant residues. J. Chem. Ecol. 1990, 16, 645–666. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, S.F.; Boydston, R.A. Volatile allelochemicals released by crucifer green manures. J. Chem. Ecol. 1997, 23, 2107–2116. [Google Scholar] [CrossRef]
- Bialy, Z.; Oleszek, W.; Lewis, J.; Fenwick, G. Allelopathic potential of glucosinolates (mustard oil glycosides) and their degradation products against wheat. Plant Soil 1990, 129, 277–281. [Google Scholar] [CrossRef]
- Azirak, S.; Karaman, S. Allelopathic effect of some essential oils and components on germination of weed species. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2008, 58, 88–92. [Google Scholar] [CrossRef]
- Wink, M. Inhibition of seed germination by quinolizidine alkaloids: Aspects of allelopathy in Lupinus albus L. Planta 1983, 158, 365–368. [Google Scholar] [CrossRef]
- Chiji, H.; Giga, T.; Izawa, M.; Kiriyama, S. Two phenolic amides in the seed balls of sugar beet (Beta vulgaris L. var saccharifera Alefeld). Agric. Biol. Chem. 1984, 48, 1653–1654. [Google Scholar] [CrossRef] [Green Version]
- Laura, A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Phenolic compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–271. [Google Scholar]
- Noperi-Mosqueda, L.C.; López-Moreno, F.J.; Navarro-León, E.; Sánchez, E.; Blasco, B.; Moreno, D.A.; Soriano, T.; Ruiz, J.M. Effects of asparagus decline on nutrients and phenolic compounds, spear quality, and allelopathy. Sci. Hortic. 2020, 261, 109029. [Google Scholar] [CrossRef]
- Li, H.-H.; Inoue, M.; Nishimura, H.; Mizutani, J.; Tsuzuki, E. Interactions of trans-cinnamic acid, its related phenolic allelochemicals, and abscisic acid in seedling growth and seed germination of lettuce. J. Chem. Ecol. 1993, 19, 1775–1787. [Google Scholar] [CrossRef]
- Cutillo, F.; D’Abrosca, B.; DellaGreca, M.; Fiorentino, A.; Zarrelli, A. Lignans and neolignans from Brassica fruticulosa: Effects on seed germination and plant growth. J. Agric. Food Chem. 2003, 51, 6165–6172. [Google Scholar] [CrossRef]
- Ng, P.; Ferrarese, M.; Huber, D.; Ravagnani, A.; Ferrarese-Filho, O. Canola (Brassica napus L.) seed germination influenced by cinnamic and benzoic acids and derivatives: Effects on peroxidase. Seed Sci. Technol. 2003, 31, 39–46. [Google Scholar] [CrossRef]
- Radwan, A.M.; Alghamdi, H.A.; Kenawy, S.K. Effect of Calotropis procera L. plant extract on seeds germination and the growth of microorganisms. Ann. Agric. Sci. 2019, 64, 183–187. [Google Scholar] [CrossRef]
- Einhellig, F.A.; Rasmussen, J.A. Synergistic inhibitory effects of vanillic and p-hydroxybenzoic acids on radish and grain sorghum. J. Chem. Ecol. 1978, 4, 425–436. [Google Scholar] [CrossRef]
- Demos, E.; Woolwine, M.; Wilson, R.; McMillan, C. The effects of ten phenolic compounds on hypocotyl growth and mitochondrial metabolism of mung bean. Am. J. Bot. 1975, 62, 97–102. [Google Scholar] [CrossRef]
- Levin, D.A.; York, B.M., Jr. The toxicity of plant alkaloids: An ecogeographic perspective. Biochem. Syst. Ecol. 1978, 6, 61–76. [Google Scholar] [CrossRef]
- Levitt, J.; Lovett, J. Activity of allelochemicals of Datura stramonium L.(thorn-apple) in contrasting soil types. Plant Soil 1984, 79, 181–189. [Google Scholar] [CrossRef]
- Levitt, J.; Lovett, J. Alkaloids, antagonisms and allelopathy. Biol. Agric. Hortic. 1985, 2, 289–301. [Google Scholar] [CrossRef]
- Aerts, R.J.; Snoeijer, W.; van der Meijden, E.; Verpoorte, R. Allelopathic inhibition of seed germination by Cinchona alkaloids? Phytochemistry 1991, 30, 2947–2951. [Google Scholar] [CrossRef]
- Weston, L.A.; Duke, S.O. Weed and crop allelopathy. Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Singh, H.; Batish, D.; Setia, N.; Kohli, R. Herbicidal activity of volatile oils from Eucalyptus citriodora against Parthenium hysterophorus. Ann. Appl. Biol. 2005, 146, 89–94. [Google Scholar] [CrossRef]
- Ramezani, S.; Saharkhiz, M.J.; Ramezani, F.; Fotokian, M.H. Use of essential oils as bioherbicides. J. Essent. Oil Bear. Plants 2008, 11, 319–327. [Google Scholar] [CrossRef]
- Benvenuti, S.; Cioni, P.L.; Flamini, G.; Pardossi, A. Weeds for weed control: Asteraceae essential oils as natural herbicides. Weed Res. 2017, 57, 342–353. [Google Scholar] [CrossRef]
- Bell, E.A. ’Uncommon’amino acids in plants. Febs Lett. 1976, 64, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, G.A.; Hughes, C.G.; Janzen, D.H. L-Canavanine, a dietary nitrogen source for the seed predator Caryedes brasiliensis (Bruchidae). Science 1982, 217, 353–355. [Google Scholar] [CrossRef]
- Bell, E.; Janzen, D.H. Medical and ecological considerations of L-dopa and 5-HTP in seeds. Nature 1971, 229, 136–137. [Google Scholar] [CrossRef]
- Elmore, C. Inhibition of turnip (Brassica rapa) seed germination by velvetleaf (Abutilon theophrasti) seed. Weed Sci. 1980, 28, 658–660. [Google Scholar] [CrossRef]
- Wilson, M.F.; Bell, E.A. The determination of the changes in the free amino acid content of the eluate from germinating seeds of Glycine wightii (L.) and its effect on the growth of lettuce fruits. J. Exp. Bot. 1978, 29, 1243–1247. [Google Scholar] [CrossRef]
- Vurro, M.; Boari, A.; Pilgeram, A.L.; Sands, D.C. Exogenous amino acids inhibit seed germination and tubercle formation by Orobanche ramosa (broomrape): Potential application for management of parasitic weeds. Biol. Control 2006, 36, 258–265. [Google Scholar] [CrossRef]
- Amor, Y.; Babiychuk, E.; Inzé, D.; Levine, A. The involvement of poly (ADP-ribose) polymerase in the oxidative stress responses in plants. Febs Lett. 1998, 440, 1–7. [Google Scholar] [CrossRef]
- Doucet-Chabeaud, G.; Godon, C.; Brutesco, C.; De Murcia, G.; Kazmaier, M. Ionising radiation induces the expression of PARP-1 and PARP-2 genes in Arabidopsis. Mol. Genet. Genom. 2001, 265, 954–963. [Google Scholar] [CrossRef]
- Zheng, X.-Q.; Hayashibe, E.; Ashihara, H. Changes in trigonelline (N-methylnicotinic acid) content and nicotinic acid metabolism during germination of mungbean (Phaseolus aureus) seeds. J. Exp. Bot. 2005, 56, 1615–1623. [Google Scholar] [CrossRef] [Green Version]
- Hunt, L.; Holdsworth, M.J.; Gray, J.E. Nicotinamidase activity is important for germination. Plant J. 2007, 51, 341–351. [Google Scholar] [CrossRef]
- Oster, U.; Bios, I.; Rüdiger, W. Natural Inhibitors of germination and growth IV compounds from fruit and seeds of mountain ash (Sorbus aucuparia). Z. Nat. C 1987, 42, 1179–1184. [Google Scholar] [CrossRef]
- Levi-Minzi, R.; Saviozzi, A.; Riffaldi, R. Organic acids as seed germination inhibitors. J. Environ. Sci. Health Part A 1994, 29, 2203–2217. [Google Scholar] [CrossRef]
- Uygur, S. Effects of mustard oil on germination and growth of yellow starthistle (Centaurea solstitialis L.). Allelopath. J. 2011, 27, 23–32. [Google Scholar]
- Bian, F.; Su, J.; Liu, W.; Li, S. Dormancy release and germination of Taxus yunnanensis seeds during wet sand storage. Sci. Rep. 2018, 8, 3205. [Google Scholar] [CrossRef] [Green Version]
- Prudente, D.d.O.; Paiva, R. Seed dormancy and germination: Physiological considerations. J. Cell Dev. Biol. 2018, 2, 2. [Google Scholar]
- Zehhar, N.; Ingouff, M.; Bouya, D.; Fer, A. Possible involvement of gibberellins and ethylene in Orobanche ramosa germination. Weed Res. 2002, 42, 464–469. [Google Scholar] [CrossRef]
- Footitt, S.; Douterelo-Soler, I.; Clay, H.; Finch-Savage, W.E. Dormancy cycling in Arabidopsis seeds is controlled by seasonally distinct hormone-signaling pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 20236–20241. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Qian, C.; Gao, Y.; Chen, L.; Zhu, M.; Pan, Y.; Li, S. Germination inhibitors detected in Sapium sebiferum seeds. J. For. Res. 2019, 30, 2305–2312. [Google Scholar] [CrossRef]
- Ali, A.S.; Elozeiri, A.A. Metabolic processes during seed germination. Adv. Seed Biol. 2017, 2017, 141–166. [Google Scholar]
- Labbafi, M.; Mehrafarin, A.; Badi, H.; Ghorbani, M.; Tavakoli, M. Improve germination of caper (Capparis spinosa L.) seeds by different induction treatments of seed dormancy breaking. Trakia J. Sci. 2018, 16, 71–74. [Google Scholar] [CrossRef]
- Hendricks, S.; Taylorson, R. Promotion of seed germination by nitrates and cyanides. Nature 1972, 237, 169–170. [Google Scholar] [CrossRef]
- Bogatek, R.; Dziewanowska, K.; Lewak, S. Hydrogen cyanide and embryonal dormancy in apple seeds. Physiol. Plant. 1991, 83, 417–421. [Google Scholar] [CrossRef]
- Roberts, E. Seed Dormancy and Oxidation Processes. Symp. Soc. Exp. Biol. 1969, 23, 161–192. [Google Scholar]
- Oracz, K.; El-Maarouf-Bouteau, H.; Kranner, I.; Bogatek, R.; Corbineau, F.; Bailly, C. The mechanisms involved in seed dormancy alleviation by hydrogen cyanide unravel the role of reactive oxygen species as key factors of cellular signaling during germination. Plant Physiol. 2009, 150, 494–505. [Google Scholar] [CrossRef] [Green Version]
- Mullick, P.; Chatterji, U. Effect of sodium cyanide on germination of two leguminous seeds. Osterr. Bot. Z. 1967, 114, 88–91. [Google Scholar] [CrossRef]
- Taylorson, R.; Hendricks, S. Promotion of seed germination by cyanide. Plant Physiol. 1973, 52, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Devlin, H.R.; Harris, I.J. Mechanism of the oxidation of aqueous phenol with dissolved oxygen. Ind. Eng. Chem. Fundam. 1984, 23, 387–392. [Google Scholar] [CrossRef]
- Yang, Z.H.; Chen, X.M.; Chen, C.H. Characteristics of Taxodinm distichum var imbricatum Seed Coat and Their Effects on Seed Dormancy. Sci. Silvae Sin. 2022, 222, 11–21. [Google Scholar]
- Li, W.; Tang, H.; He, L. Effects of Crude Extracts of Paeonia rockii Seeds on Germination and Seedling Growth of receptor Plant. J. Northeast For. Univ. 2020, 48, 8–12. [Google Scholar]
- Yan, F.; Zhang, E.; Wang, Q.; Mao, D. The Mechanisms of Seed Dormancy of Wild Daphne giraldii and Methods for Dormancy Breaking. Sci. Silvae Sin. 2016, 52, 30–37. [Google Scholar]
- Li, Q.; Liu, Y.; Liu, G.; Liu, Y.; Hou, L.; Hu, J. Study on germination inhibitors of seeds of 7 species of Quercus species. Acta Ecol. Sin. 2013, 7, 2104–2112. [Google Scholar]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef]
- Miersch, O.; Neumerkel, J.; Dippe, M.; Stenzel, I.; Wasternack, C. Hydroxylated jasmonates are commonly occurring metabolites of jasmonic acid and contribute to a partial switch-off in jasmonate signaling. New Phytol. 2008, 177, 114–127. [Google Scholar] [CrossRef]
- Oh, E.; Kang, H.; Yamaguchi, S.; Park, J.; Lee, D.; Kamiya, Y.; Choi, G. Genome-wide analysis of genes targeted by PHYTOCHROME INTERACTING FACTOR 3-LIKE5 during seed germination in Arabidopsis. Plant Cell 2009, 21, 403–419. [Google Scholar] [CrossRef] [Green Version]
- Graeber, K.; Linkies, A.; Müller, K.; Wunchova, A.; Rott, A.; Leubner-Metzger, G. Cross-species approaches to seed dormancy and germination: Conservation and biodiversity of ABA-regulated mechanisms and the Brassicaceae DOG1 genes. Plant Mol. Biol. 2010, 73, 67–87. [Google Scholar] [CrossRef]
- Pal, D.; Mukherjee, S. Tamarind (Tamarindus indica) seeds in health and nutrition. In Nuts and Seeds in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2020; pp. 171–182. [Google Scholar]
Figure 1.
Relationship between germination retarders, germination destructors and allelochemicals.

Figure 2.
Separation process of seed methanol extract.


{kind=link}
{kind=link}
Table 1.
Classification of germination inhibitors by function.
| Classification | Subdivision | Definition |
|---|---|---|
| Germination retarder | Endogenous germination retarder | Plant-produced substances that inhibit seed germination |
| Exogenous germination retarder | Non-plant substances that inhibit seed germination | |
| Germination destructor | Selective germination destructor | Substances that inflict physiological harm on the seeds of one or more plants |
| Unselective germination destructor | Substances that cause general seed damage |
Table 2.
Classification of germination inhibitors by chemical structure.
| Name of Germination Inhibitors | Locations of Existence | Form of Existence | Particular Plants | Inhibitory Effects | ||
|---|---|---|---|---|---|---|
| Phenols | Caffeic acid | Seeds, fruits, and other plant tissues | Either in a free state or conjugated with sugars as glucosides and esters | Cucumis melo [37] | retarder | Caffeic acid delayed germination rather than destroying it. |
| Asparagus [38] | destructor | Caffeic Acid Identified as an inhibitory compound in Asparagus root filtrate. | ||||
| Chlorogenic acid | Sida spinosa, Sorghum bicolor [37] | destructor | It slightly reduced germination of Sida spinosa and Sorghum bicolor seeds. | |||
| Coumarin | Amaranthus retroflexus, Sida spinosa [37] | retarder | Germination was delayed rather than destructed by chlorogenic acid. | |||
| Arabidopsis thaliana [39] | destructor | It reduces germination of Arabidopsis thaliana seeds and led to reduced primary radicle elongation. | ||||
| Oryza sativa [24] | retarder | Coumarin induced delay of Oryza sativa seed germination is mediated by suppression of ABA catabolism and reduced production of ROS. | ||||
| Brassica parachinensis [25] | retarder | It delayed germination of Brassica parachinensis seeds by decreased GA4 production and a consequent reduction of ROS accumulation. | ||||
| p-Coumarie acid | Zea mays [37] | destructor | It significantly decreased the germination of Zea mays seeds. | |||
| Ferulic acid | Gossypiumhirsutum [37] | destructor | It significantly decreased the germination of Gossypium hirsutum seeds. | |||
| Fumaric acid | Sida spinosa [37] | destructor | It reduced the germination rate of Sida spinosa seedlings slightly. | |||
| Gallic acid | Cucumis sativus [40] | destructor | It greatly reduced germination rate, radicle and hypocotyl growth, and seedling fresh and dry weight. | |||
| Hydrocinnamic acid | Amaranthus retroflexus [37] | retarder | It delayed germination rather than destroying it. | |||
| Pyrocatechol | Amaranthus retroflexus [37] | retarder | It delayed germination rather than destroying it. | |||
| p-Hydroxybenzoic acid | Chenopodium album, Plantago lanceolata, Amaranthus retroflexus, Solanum nigrum, Cirsium, Rumex crispus [41] | retarder | A significant concentration of p-hydroxybenzoic acid inhibited the germination of all of these weeds. | |||
| Juglone | Zea mays [37] | retarder | It delayed germination rather than destroying it. | |||
| Abutilon theophrasti, Sida spinosa, Amaranthus retroflexus [37] | destructor | Seed growth is destroyed by juglone. | ||||
| Flavonoids | Flavonols | Seeds | Low molecular weight polyphenolic secondary metabolic compounds | Zea mays [42] | retarder | It takes part in seeds maturation and dormancy. |
| Proanthocyanidins | Arabidopsis thaliana [43] | retarder | It contributes to the maintenance of seed dormancy by promotion of ABA. | |||
| Dihydroflavonoids | Brassica campestris and Echinochloa crusgalli [44] | destructor | It inhibits embryo growth. | |||
| Aldehydes | Crotonaldehyde | Stem and root | Natural metabolic secretions of plants | Amaranthus tricolor [45] | destructor | The substance inhibits the growth of the germ and radicle |
| (E)-2-hexenal | Trifolium alexandrinum [46] | destructor | It inhibits germination and seedling development | |||
| 3-methylbutanal | Trifolium alexandrinum [46] | destructor | It inhibits germination and seedling development | |||
| Benzaldehyde | Brassicaceae [47] | destructor | It inhibits seed germination. | |||
| Mustard oil | Mustard oil glycosides | Seeds, and other plant tissues | Phytochemical of plants | Triticumturgidumvar. durum [48] | destructor | It completely inhibited Triticum turgidum var. durum seeds germination at 500 ppm. |
| Aromatic oils | Carum carvi | Most of plant tissues | Natural metabolic secretions of plants | Amaranthus retroflexus, Centaurea salsotitialis and Raphanus raphanistrum [49] | destructor | Even at low concentrations, thymol, carvacrol, and carvone showed significant inhibition of these seeds. |
| Mentha spicata | ||||||
| Origanum onites | ||||||
| Thymbra spicata | ||||||
| Alkaloid | Quinolizidine alkaloids | Seeds | Present as ester alkaloids | Lactuca sativa [50] | destructor | The alkaloid esters resulted in the strongest inhibition: 6 mM 13-tigloyloxylupanine inhibited germination by 100%. |
| Amide | Feruloylputrescine | Seeds and leaves | Natural secondary metabolites | Beta vulgaris [51] | retarder | It hinders germination of seeds. |
| Feruloylserotonin | ||||||
| N-trans-Feruloyltyramine | ||||||
Notes: Different colors represent different plant secondary metabolites, including Phenols, Flavonoids, Aldehydes, Mustard oil, Aromatic oils, Alkaloid and Amide.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chenyin, P.; Yu, W.; Fenghou, S.; Yongbao, S. Review of the Current Research Progress of Seed Germination Inhibitors. Horticulturae 2023, 9, 462. https://doi.org/10.3390/horticulturae9040462
AMA Style
Chenyin P, Yu W, Fenghou S, Yongbao S. Review of the Current Research Progress of Seed Germination Inhibitors. Horticulturae. 2023; 9(4):462. https://doi.org/10.3390/horticulturae9040462
Chicago/Turabian StyleChenyin, Peng, Wu Yu, Shi Fenghou, and Shen Yongbao. 2023. "Review of the Current Research Progress of Seed Germination Inhibitors" Horticulturae 9, no. 4: 462. https://doi.org/10.3390/horticulturae9040462
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.