
The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review


Department of Biological Sciences, Northern Arizona University, Flagstaff, AZ 86011, USA
*
Authors to whom correspondence should be addressed.
Biomimetics 2022, 7(3), 87; https://doi.org/10.3390/biomimetics7030087
Submission received: 29 April 2022
/
Revised: 23 June 2022
/
Accepted: 29 June 2022
/
Published: 1 July 2022
Abstract
:The extracellular matrix (ECM) is a 3-dimensional structure and an essential component in all human tissues. It is comprised of varying proteins, including collagens, elastin, and smaller quantities of structural proteins. Studies have demonstrated the ECM aids in cellular adherence, tissue anchoring, cellular signaling, and recruitment of cells. During times of integumentary injury or damage, either acute or chronic, the ECM is damaged. Through a series of overlapping events called the wound healing phases—hemostasis, inflammation, proliferation, and remodeling—the ECM is synthesized and ideally returned to its native state. This article synthesizes current and historical literature to demonstrate the involvement of the ECM in the varying phases of the wound healing cascade.
1. Introduction
Wound healing is a complex and coordinated cascade of events that helps to maintain homeostasis within the integument [1,2] and subsequently protect the whole organism [3]. When considering dermal wound healing, the longer a wound takes to heal, the greater the opportunity for foreign agents to enter the body and have pathological effects. To help mitigate the duration of the wound, implanting materials into the integumentary can alter the wound healing cascade of events [4,5]. Currently, translational scientists are researching and developing novel ways to create bio-inspired scaffolds able to reproduce the extracellular environment of the native tissues [6,7]. It has been estimated that the advanced wound care market will be approximately 19 billion dollars by 2027 [8]. Having these technological advancements offers patients within this growing market novel treatment regimens many of which will involve reconstruction of the native ECM. Granted, the type of wound would be the greatest indicator of the need to treat with the addition of an implanted bio-inspired scaffold.
The etiologic agent will dictate the type of wound healing that occurs. First intention wound healing is often a result of surgical incisions or a clean laceration [9]. These first intention wounds do not lead to fibrotic formation rather heal through connective tissue and epithelial regeneration [9,10,11]. Second intention wound healing is a more involved process and often occurs during soft tissue loss such as ulcerations, severe burns, and major surgeries [9,10]. Second intention healing occurs through the formation of granulation tissue which is then followed by the synthesis of extracellular matrix (ECM), much of which is due to fibroblasts [9]. These second intention wounds close through wound contraction by way of myofibroblasts and re-epithelialization from the wound margins [12,13]. Second intention (full thickness) wounds healing time is often delayed due to infection or disease states such as diabetes, which alter the normal physiological processes [9,14,15,16]. During healing by second intention, the protein elastin, responsible for the elastic recoil of the dermal matrix, is absent from the healing granulation tissue deposited by resident fibroblasts [17]. The lack of elastin creates a more rigid and inelastic ECM. Collagen, which provides tensile strength to the skin, is also secreted into the healing wound space [17]. The secreted collagen is disorganized, lacks structural integrity, and the resulting scar tissue never achieves the tensile strength of native skin [17]. Current methods to treat intractable wounds that involve second intention healing include the use of antibacterial, debridement, irrigation, vacuum-assisted closure, oxygenation, moist wound healing, hirudotherapy, and collagen-based dressings/fillers [18,19,20,21,22].
Restoration of a dermal matrix that mimics the unwounded structure and function, preserving the dermal integrity, would theoretically improve the resulting scar so that the tensile strength and elastic recoil would approximate that of unwounded, intact dermis [17]. The microarchitecture of scaffolds is relevant to tissue engineering due to the ability of the scaffolds to mimic native ECM in scale, which is thought to encourage cellular ingrowth, ECM deposition, and neotissue formation [23,24,25,26]. Table 1 contains a list of products used for wound healing which utilize ECM proteins, and a short list of medical devices created from synthetic polymers which are manufactured to mimic the ECM.
2. Involvement of Extracellular Matrix in Normal Wound Healing
Normal wound healing is a dynamic process that involves epithelial and dermal regeneration as well as scar development, which includes ECM regeneration. When normal tissue is disrupted, either cut surgically or through ulcerations, a healthy organism must be able to repair itself for longevity and survival. The normal wound healing model is characterized by four phases: clotting and coagulation (hemostasis), inflammation, proliferation, and remodeling [9,43,44,45,46]. These phases are not mutually exclusive, overlap in the organism (Figure 1), and typically occur quickly (e.g., days to weeks), while the remodeling phase of the wound healing responses can take up to months (12 months) for completion [45,47].
The ECM is an acellular, protein-rich matric that is essential for structural support and cellular attachment [48,49,50,51]. Previously published studies have supported the ECM in modulating wound healing by regulating biochemical pathways and biomechanical signaling pathways [49,51,52,53,54]. The ECM directly modulates aspects of cell behavior, including adhesion, proliferation, migration, and survival [12,54,55,56]. Indirect modulation involves cells acting on the ECM stimulates extracellular protease secretion and modulating growth factor bioavailability [54].
The two most ubiquitous proteins found in the native ECM are collagen and elastin. Collagen is the most prevalent protein found in the ECM [57,58]. It comprises approximately 50–90% of the integument [19,59]. Table 2 details the percent composition of the ECM. Collagen is predominately synthesized by fibroblasts and there are many variants of collagen. Collagen type I is the dominant form in adults, while collagen type III is the most abundant during gestational development [60]. Interestingly, in 2020 it was published that the precursor to collagen, pre-collagen, is regulated by the 24-h circadian cycle [61]. Elastin, on the other hand, comprises 0.6–7.9% of the dermal ECM depending on anatomical location, gender, and age [59,62,63,64,65,66,67]. Other proteins found in the ECM include laminin, fibronectin, vitronectin, and tenascin, as well as proteoglycans and glycosaminoglycans (GAGs) that make up the remaining 3–5% of the ECM [68,69]. For the most part these proteins will be created in the newly formed ECM, with the exception of elastin. In human skin and most organs, the bulk of elastogenesis occurs during late fetal and early neonatal periods. By maturity production it is complete and synthesis of new tropoelastin ceases [67,70,71]. Elastin essentially does not turn over in healthy tissues, therefore fiber function and tissue integrity are compromised due to this limited pattern of elastin production over the life of specific organs [70,72,73,74].
3. Clotting and Coagulation (Hemostasis)
Wound healing begins immediately following insult to the tissue when platelets contact exposed collagen from the endothelial wall and ECM initiating clotting and coagulation [75,76,77,78]. Upon adhesion to collagen, adenosine diphosphate (ADP) is released from platelet granules and erythrocytes to initiate platelet aggregation. An anti-hemophilic factor (Factor VIII) is also released from the platelet alpha granules [79,80]. Factor VIII accelerates the formation of the platelet plug and, in conjunction with the fibrin network, fills the damaged tissue space. This structure provides a scaffold for cellular migration and proliferation, essentially a provisional extracellular matrix (ECM) [12,38,81,82]. The fibrin network is rapidly degraded by plasmin and neutrophil elastase, which can release the plasma growth factors trapped in the fibrin clot [38,83]. The fibrin degradation products also stimulate ECM deposition [84], fibroblast proliferation [85], and angiogenesis [86].
The recruitment of cells to the damaged tissue area is an important component of wound healing [44]. This mechanism primarily relies on two growth factors: platelet-derived growth factor (PDGF) and transforming growth factor β (TGF-β) [68,77,87,88,89]. PDGF and TGF-β initiate chemotaxis of neutrophils, macrophages, fibroblasts, and smooth muscle cells. These processes are required for the wound healing cascade to progress to the next stage, inflammation [77,90,91].
4. Inflammation
Following platelet activation, the first inflammatory cells to invade the wound site are the neutrophils, which are recruited to the site by PDGF and TGF-β [12]. These neutrophils aid in protection against infection and removal of tissue debris. Following neutrophils, monocytes and T-lymphocytes occupy the tissue. The recruitment of monocytes and T-lymphocytes are the first overlap of steps between hemostasis and inflammation. The fragmentation of fibronectin and other ECM components act as chemoattractants for monocytes which bind ECM proteins [17,92]. This binding stimulates phagocytosis, [17,93] leading the monocytes/macrophages to further break down ECM fragments and other debris in the area [17,91,94]. Adherence of monocytes to ECM proteins also stimulates the expression of growth factors [17,95] that can then act on cells to affect the synthesis of ECM components (e.g., proteoglycan synthesis by fibroblasts) [17,96,97]. Inflammation plays a primary role in prevention of infection of the tissue [98,99].
Acute inflammation initiates within 24 h of injury and is characterized by initial vasoconstriction followed by vasodilation and increased capillary permeability [100]. Neutrophils are recruited to the wound site within an hour by factors released from the platelets in the hemostasis phase (including interleukin-1, TNF-α, TGF-β, and platelet factor-4,) and have a sustained migration for 48 h [78,101,102]. Neutrophils from the circulating blood are recruited through molecular changes on the surface of endothelial cells [103].
The primary function of infiltrating neutrophils is to remove foreign or damaged particles, bacteria, and non-functioning host cells found in the wound [104]. Neutrophils accomplish this through phagocytosis, degranulation, and the production of chromatin and protease traps [98]. Granules release several toxic enzymes that include lactoferrin (antibacterial effect), proteases, neutrophil elastase, and cathepsin [54,98]. The resulting debris either becomes part of the scab and is sloughed off or phagocytized by macrophages [100,101]. At the same time neutrophils are recruited, a circulating monocyte influx is stimulated by the fragmentation of fibronectin [102], elastin [103], collagen [104], enzymatically active thrombin [105], complement components, PDGF, TGF-β, leukotriene B4, and PF-4 [10]. Infiltrating monocytes become activated macrophages and function to phagocytize effete neutrophils and residual bacterial particles at the wound site. Activated macrophages release growth factors such as PDGF, vascular endothelial growth factor (VEGF) and cytokines that are required for the formation of granulation tissue and fibroblast proliferation [77,81,106,107]. Macrophages continue cleaning the wound site and stay in the wound site much longer [107,108,109].
The final cells recruited during inflammation are lymphocytes. These cells are attracted by several chemoattractants including IL-1, complement components and immunoglobulin G (IgG) breakdown products [98,100,107]. The lymphocytes regulate the proliferative phase of wound healing [98,107,110]. The role of T-lymphocytes is not completely understood and is a current area of research [2].
Another important cell in the inflammation process is the mast cell [111,112,113,114]. Mast cells release granules of histamine, heparin, interleukins (IL-6, IL8), VEGF, enzymes, and other active amines which together are responsible increasing vascular permeability and the characteristic signs of inflammation, calor (heat), tumor (swelling), rubor (redness), and dolor (pain), that surrounds the wound site [111,113,114]. The proteins released from these granules cause the surrounding vessels to become more permeable, which allows mononuclear cells and fluid to pass into the wound area giving way to the signs of inflammation [114,115].
Inflammatory resolution is largely controlled by anti-inflammatory interleukins (IL-10) and the genetic regulation of inflammatory genes [116]. During this down regulating event, additional cells will migrate to the injured tissue site. These cells include mesenchymal stem cells, fibroblasts that create extracellular matrix, and nearby endothelial cells that create new blood vessels (i.e., angiogenesis) [117]. At the end of the inflammatory phase, a rich granulation tissue has begun to be synthesized that contains a significant number of fibroblasts, capillaries, and inflammatory cells [117,118].
5. Proliferation
The next stage of wound healing is proliferation, which is marked by the rapid formation of new connective tissue that functions to repopulate the wound bed with a newly formed ECM. However, during adult wound healing the newly formed ECM does not resemble the native ECM prior to the injury [17]. The ECM during the adult wound healing event is much looser which allows for cellular invasion [81]. This stage is termed granulation tissue formation due to the gross granular appearance of the tissue when excised from the wound [43]. The tissue has a granular appearance due to the large amount of neovascularization (new blood vessels) that has taken place [43,117]. Figure 2 below details granulation tissue in a chronic wound.
During proliferation, granulation tissue converts from a cell-rich, highly vascular medium to relatively avascular and acellular matrix of collagen [119,120]. However, elastin is absent from the formed granulation tissue deposited by resident fibroblasts [17]. Elastin has only been demonstrated to be present after these wound stages have progressed, in some cases months after this proliferative event [53,67,121]. Instead, collagen provides tensile strength to the skin and is mostly secreted into the healing wound space in a disorganized way, lacking structural integrity [17,122].
After approximately 96 h, the proliferation phase initiates, and new stroma begin to invade the wound site [82]. Figure 3 details the phases of wound healing, including proliferation. Granulation tissue consists of a dense population of macrophages, fibroblasts, and neovasculature embedded in a loose matrix of collagen, fibronectin, and hyaluronic acid [123,124]. All these cells and proteins move into or are deposited into the wound space at about the same time [81]. One such cell type is the fibroblast which are signaled into the wound bed by following the collagen protein orientation that has been deposited [120,125]. As fibroblasts invade the fibrin clot, the cells are lysed and then deposit hyaluronan and fibronectin; this forms the early granulation tissue [117]. Hyaluronic acid (HA) is a large component of early granulation tissue [126,127]. The process of granulation initially occurs in the periphery of the clot and moves toward the center as the granulation tissues grow into the wound space [117,127]. The ECM of the healing wound undergoes rapid changes as the fibrin clot is replaced by fibronectin and hyaluronan and subsequently by collagen types I and II [43,117,128].
The newly forming vessels that arise during neovascularization sprout from existing vessels that are stimulated by the cytokine VEGF, fibroblast growth factor (FGF)-2, PDGF, and members of the TGF-β family [127,129,130]. Small capillary projections are initially formed through the accumulation of endothelial cells on the leading edge, with further extension occurring from endothelial proliferation [131]. The maturation of these vessels occurs with the emergence of peri-endothelial cells that aid in matrix formation and undergo a change to become pericytes (contractile cells that wrap around endothelial cells) [132]. This process is perpetuated by recruited macrophages, which continue to provide the cytokines necessary for fibroplasia.
The persistence of fibroblasts produces a new extracellular matrix that supports new cellular growth [117]. The accumulation of fibroblasts does not necessarily ensure that collagen will be synthesized and deposited; the fibroblasts must first be provided with a stimulus. This stimulus comes from the production of lactate synthesized by metabolically active macrophages and new blood vessels [133,134,135]. The neovasculature must be present to provide oxygen and nutrients to support cellular metabolism [81]. Along with neovascularization, collagen production, macrophage activation, and epithelialization occur in the proliferating tissue.
Epithelialization is stimulated by the presence of epidermal growth factors and TGF-α. These growth factors are produced by activated macrophages, platelets, and keratinocytes [136,137]. For the epidermal cells to differentiate and re-establish the permeability barrier, fibroblasts in the granular tissue must transform into myofibroblasts that contract the wound [43,124]. Once the new epithelial cells span the wound site, enzymes are released to allow the scab to be removed from the damaged tissue. The result of the proliferative phase is the increased presence of new vessels and fibrous tissue that will provide the basic foundation for tissue remodeling. Throughout the granulation tissue stage, matrix formation and remodeling occur simultaneously and overlap considerably.
6. Remodeling
The final phase of wound healing is tissue remodeling. Tissue remodeling allows for the restoration of the tissue architecture by macrophages and fibroblasts, which helps to restore tissue strength [77,96]. The length of this phase is highly variable, many times related to the wound size and pre-existing conditions of the patient. On average, this phase begins two to three weeks after the initial tissue injury event and can take up to a year or longer for remodeling to occur [138]. The length of remodeling is predominately due to the synthesis and degradation of collagen in the ECM of the wound bed [98]. During the remodeling period of wound healing, the fibronectin-rich matrix is replaced with a stronger collagenous matrix [139]. TGF-β1 stimulates production of collagen type I and III in fibroblasts [44,138]. This collagenous matrix is strengthened due to the crosslinking and structural modifications it undergoes throughout the remodeling phase. This crosslinking is completed by varying enzymes including transglutaminases and lysyl oxidases [140]. Wound resolution begins as the amount of extracellular collagen increases and fibroblasts decrease their collagen production [43,141]. During the course of a year, type III collagen is replaced by type I collagen through regulation by matrix metalloproteinases (MMPs) [142,143]. There are 24 distinct extracellular endopeptidases in the MMP family [144,145]. MMPs are proteases that breakdown ECM proteins which modulate the ECM environment [146]. The result of normal healing is a type I collagenous scar that is largely avascular and lacks any ordered structure. This is exemplified by the fact that healed wounds never regain full tensile strength [147,148]. On average, the wounds reach 50% tensile strength at three (3) months and the maximum tensile strength a wound can achieve is approximately 70–80% of normal tissue [16,43,77,100,133].
7. Conclusions
The extracellular matrix (ECM) is a critical structural component of any tissue, including the integument. Damage to the ECM through an acute or chronic source can lead to a series of healing events, coined the “wound healing cascade”. These overlapping events begin with the initial cessation of blood flow and clotting called hemostasis, followed by the inflammatory stage, progressing to the granulation-rich proliferative stage and finally the remodeling phase, which produces a collagen I tissue. All of this is responsible from cellular signaling events, many of which involve the ECM. This review addresses the role of the ECM throughout the wound healing cascade of events.
Many wound healing products currently on the market in the US have realized that using these structural proteins can increase the rate of wound heling in second intention wounds. However, this may not be enough. It is the opinion of the authors that more research needs to be devoted to not only utilizing the structural components of the ECM but to also mimic the physiological structure.
Author Contributions
Both authors contributed equally to this project. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Conflicts of Interest
The authors have the following relevant disclosures: Robert B. Diller is employed by Amnio Technology (Phoenix, AZ, USA) and Aaron J. Tabor is employed by Axolotl Biologix (Flagstaff, AZ, USA). Both companies offer membrane based wound care products.
References
- Childs, D.R.; Murthy, A.S. Overview of wound healing and management. Surg. Clin. 2017, 97, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.A.; DiPietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Sanford, J.A.; Gallo, R.L. Functions of the skin microbiota in health and disease. In Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2013; Volume 25, pp. 370–377. [Google Scholar]
- Robson, M.C.; Steed, D.L.; Franz, M.G. Wound healing: Biologic features and approaches to maximize healing trajectories. Curr. Probl. Surg. 2001, 38, 72–140. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M. Biological responses to materials. Annu. Rev. Mater. Res. 2001, 31, 81–110. [Google Scholar] [CrossRef]
- Stevens, M.M.; George, J.H. Exploring and engineering the cell surface interface. Science 2005, 310, 1135–1138. [Google Scholar] [CrossRef]
- Traversa, E.; Mecheri, B.; Mandoli, C.; Soliman, S.; Rinaldi, A.; Licoccia, S.; Forte, G.; Pagliari, F.; Pagliari, S.; Carotenuto, F.; et al. Tuning hierarchical architecture of 3D polymeric scaffolds for cardiac tissue engineering. J. Exp. Nanosci. 2008, 3, 97–110. [Google Scholar] [CrossRef]
- Sen, C.K. Human wound and its burden: Updated 2020 compendium of estimates. Adv. Wound Care 2021, 10, 281–292. [Google Scholar] [CrossRef]
- Enoch, S.; Leaper, D.J. Basic science of wound healing. Surgery 2008, 26, 31–37. [Google Scholar]
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef]
- Alhajj, M.; Goyal, A. Physiology, granulation tissue. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Midwood, K.S.; Williams, L.V.; Schwarzbauer, J.E. Tissue repair and the dynamics of the extracellular matrix. Int. J. Biochem. Cell Biol. 2004, 36, 1031–1037. [Google Scholar] [CrossRef]
- Kushwaha, A.; Goswami, L.; Kim, B.S. Nanomaterial-based therapy for wound healing. Nanomaterials 2022, 12, 618. [Google Scholar] [CrossRef]
- Hess, C.T.; Kirsner, R.S. Orchestrating wound healing: Assessing and preparing the wound bed. Adv. Ski. Wound Care 2003, 16, 246–257. [Google Scholar] [CrossRef]
- Gantwerker, E.A.; Hom, D.B. Skin: Histology and physiology of wound healing. Clin. Plast. Surg. 2012, 39, 85–97. [Google Scholar] [CrossRef]
- Carr, N.J. The pathology of healing and repair. Surgery 2021, 40, 13–19. [Google Scholar] [CrossRef]
- Schultz, G.S.; Wysocki, A. Interactions between extracellular matrix and growth factors in wound healing. Wound Repair Regen. 2009, 17, 153–162. [Google Scholar] [CrossRef]
- Mustoe, T. Understanding chronic wounds: A unifying hypothesis on their pathogenesis and implications for therapy. Am. J. Surg. 2004, 187, S65–S70. [Google Scholar] [CrossRef]
- Brett, D. A Review of Collagen and Collagen-based Wound Dressings. Wounds 2008, 20, 347–356. [Google Scholar]
- Mees, J.; Mardin, W.A.; Senninger, N.; Bruewer, M.; Palmes, D.; Mees, S.T. Treatment options for postoperatively infected abdominal wall wounds healing by secondary intention. Langenbeck’s Arch. Surg. 2012, 397, 1359–1366. [Google Scholar] [CrossRef]
- Yamaoka, H.; Sumiyoshi, H.; Higashi, K.; Nakao, S.; Minakawa, K.; Sumida, K.; Saito, K.; Ikoma, N.; Mabuchi, T.; Ozawa, A.; et al. A novel small compound accelerates dermal wound healing by modifying infiltration, proliferation and migration of distinct cellular components in mice. J. Dermatol. Sci. 2014, 74, 204–213. [Google Scholar] [CrossRef]
- Nair, H.K.; Ahmad, N.W.; Lee, H.L.; Ahmad, N.; Othamn, S.; Mokhtar, N.S.H.M.; Chong, S.S.Y. Hirudotherapy in wound healing. Int. J. Low. Extrem. Wounds 2020. [Google Scholar] [CrossRef]
- Barnes, C.P.; Sell, S.A.; Boland, E.D.; Simpson, D.G.; Bowlin, G.L. Nanofiber technology: Designing the next generation of tissue engineering scaffolds. Adv. Drug Deliv. Rev. 2007, 59, 1413–1433. [Google Scholar] [CrossRef] [PubMed]
- Bonvallet, P.P.; Culpepper, B.K.; Bain, J.L.; Schultz, M.J.; Thomas, S.J.; Bellis, S.L. Microporous dermal-like electrospun scaffolds promote accelerated skin regeneration. Tissue Eng. Part A 2014, 20, 2434–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norouzi, M.; Boroujeni, S.M.; Omidvarkordshouli, N.; Soleimani, M. Advances in skin regeneration: Application of electrospun scaffolds. Adv. Healthc. Mater. 2015, 4, 1114–1133. [Google Scholar] [CrossRef] [PubMed]
- Bargavi, P.; Ramya, R.; Chitra, S.; Vijayakumari, S.; Chandran, R.R.; Durgalakshmi, D.; Rajashree, P.; Balakumar, S. Bioactive, degradable and multi-functional three-dimensional membranous scaffolds of bioglass and alginate composites for tissue regenerative applications. Biomater. Sci. 2020, 8, 4003–4025. [Google Scholar] [CrossRef] [PubMed]
- Underwood, P.; Cardinal, P.; Keller, E.; Goodfellow, R.; Scalea, T.; Henry, S.; Lauerman, M.H. Extending Limb Salvage after Fourth and Fifth Transmetatarsal Amputation in Diabetic Foot Infections Using ACell® Urinary Bladder Matrix. Am. Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, R.V.; James, S.L.; James, S.E. A review of tissue-engineered skin bioconstructs available for skin reconstruction. J. R. Soc. Interface 2010, 7, 229–258. [Google Scholar] [CrossRef] [Green Version]
- Kuo, S.; Kim, H.M.; Wang, Z.; Bingham, E.L.; Miyazawa, A.; Marcelo, C.L.; Feinberg, S.E. Comparison of two decellularized dermal equivalents. J. Tissue Eng. Regen. Med. 2018, 12, 983–990. [Google Scholar] [CrossRef]
- Tierney, B.P. Comparison of 30-day Clinical Outcomes with SimpliDerm and AlloDerm RTU in Immediate Breast Reconstruction. Plast. Reconstr. Surg. Glob. Open 2021, 9, e3648. [Google Scholar] [CrossRef]
- Vo, A.; Diller, R.; Kellar, R. Characterization and Clinical Applications of Amniotic Membranes. J. Pharmacol. Clin. Res. 2017, 4. [Google Scholar] [CrossRef]
- Valentin, J.E.; Badylak, J.S.; McCabe, G.P.; Badylak, S.F. Extracellular matrix bioscaffolds for orthopaedic applications: A comparative histologic study. JBJS 2006, 88, 2673–2686. [Google Scholar] [CrossRef]
- Chocarro-Wrona, C.; López-Ruiz, E.; Perán, M.; Gálvez-Martín, P.; Marchal, J.A. Therapeutic strategies for skin regeneration based on biomedical substitutes. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 484–496. [Google Scholar] [CrossRef]
- Lisa, A.V.E.; Galtelli, L.; Vinci, V.; Veronesi, A.; Cozzaglio, L.; Cananzi, F.C.M.; Sicoli, F.; Klinger, M. Adoption of a newly introduced dermal matrix: Preliminary experience and future directions. BioMed Res. Int. 2020, 2020, 3261318. [Google Scholar] [CrossRef]
- Dai, C.; Shih, S.; Khachemoune, A. Skin substitutes for acute and chronic wound healing: An updated review. J. Dermatol. Treat. 2020, 31, 639–648. [Google Scholar] [CrossRef]
- Yeh, D.D.; Nazarian, R.M.; Demetri, L.; Mesar, T.; Dijkink, S.; Larentzakis, A.; Velmahos, G.; Sadik, K.W. Histopathological assessment of OASIS ultra on critical-sized wound healing: A pilot study. J. Cutan. Pathol. 2017, 44, 523–529. [Google Scholar] [CrossRef]
- Kellar, R.S.; Diller, R.B.; Tabor, A.J.; Dominguez, D.D.; Audet, R.G.; Bardsley, T.A.; Talbert, A.J.; Cruz, N.D.; Ingraldi, A.L.; Ensley, B.D. Improved wound closure rates and mechanical properties resembling native skin in murine diabetic wounds treated with a tropoelastin and collagen wound healing device. J. Diabetes Clin. Res. 2020, 2, 86. [Google Scholar]
- Hsu, P.W.; Salgado, C.J.; Kent, K.; Finnegan, M.; Pello, M.; Simons, R.; Atabek, U.; Kann, B. Evaluation of porcine dermal collagen (Permacol) used in abdominal wall reconstruction. J. Plast. Reconstr. Aesthetic Surg. 2009, 62, 1484–1489. [Google Scholar] [CrossRef]
- Veves, A.; Sheehan, P.; Pham, H.T. A randomized, controlled trial of Promogran (a collagen/oxidized regenerated cellulose dressing) vs. standard treatment in the management of diabetic foot ulcers. Arch. Surg. 2002, 137, 822–827. [Google Scholar] [CrossRef]
- Thakar, H.; Sebastian, S.M.; Mandal, S.; Pople, A.; Agarwal, G.; Srivastava, A. Biomolecule-conjugated macroporous hydrogels for biomedical applications. ACS Biomater. Sci. Eng. 2019, 5, 6320–6341. [Google Scholar] [CrossRef]
- Patil, P.; Russo, K.A.; McCune, J.T.; Pollins, A.C.; Cottam, M.A.; Dollinger, B.R.; Colazo, J.M.; Yu, F.; Martin, J.R.; Gupta, M.K.; et al. ROS-Degradable Polythioketal Urethane Foam Dressings to Promote Porcine Skin Wound Repair. bioRxiv 2021. [Google Scholar] [CrossRef]
- Puhl, D.L.; Mohanraj, D.; Nelson, D.W.; Gilbert, R.J. Designing electrospun fiber platforms for efficient delivery of genetic material and genome editing tools. Adv. Drug Deliv. Rev. 2022, 183, 114161. [Google Scholar] [CrossRef]
- Clark, R.A.; Ashcroft, G.S.; Spencer, M.J.; Larjava, H.; Ferguson, M.W.J. Re-epithelialization of normal human excisional wounds is associated with a switch from αvβ5 to αvβ6 integrins. Br. J. Dermatol. 1996, 135, 46–51. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of acute wound healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef]
- Ellis, S.; Lin, E.J.; Tartar, D. Immunology of wound healing. Curr. Dermatol. Rep. 2018, 7, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Becerra-Bayona, S.M.; Solarte-David, V.A.; Sossa, C.L.; Mateus, L.C.; Villamil, M.; Pereira, J.; Arango-Rodríguez, M.L. Mesenchymal Stem Cells Derivatives as a Novel and Potential Therapeutic Approach to Treat Diabetic Foot Ulcers. Endocrinol. Diabetes Metab. Case Rep. 2020, 2020, 19–0164. [Google Scholar] [CrossRef]
- Wallace, H.A.; Basehore, B.M.; Zito, P.M. Wound Healing Phases; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- Lukashev, M.E.; Werb, Z. ECM signalling: Orchestrating cell behaviour and misbehaviour. Trends Cell Biol. 1998, 8, 437–441. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Pfisterer, K.; Shaw, L.E.; Symmank, D.; Weninger, W. The extracellular matrix in skin inflammation and infection. Front. Cell Dev. Biol. 2021, 9, 682414. [Google Scholar] [CrossRef] [PubMed]
- David, V.A.S.; Güiza-Argüello, V.R.; Arango-Rodríguez, M.L.; Sossa, C.L.; Becerra-Bayona, S.M. Decellularized Tissues for Wound Healing: Towards Closing the Gap Between Scaffold Design and Effective Extracellular Matrix Remodeling. Front. Bioeng. Biotechnol. 2022, 10, 821852. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. The extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almine, J.F.; Bax, D.V.; Mithieux, S.M.; Nivison-Smith, L.; Rnjak, J.; Waterhouse, A.; Wise, S.; Weiss, A.S. Elastin-based materials. Chem. Soc. Rev. 2010, 39, 3371–3379. [Google Scholar] [CrossRef] [PubMed]
- Maquart, F.X.; Monboisse, J.C. Extracellular matrix and wound healing. Pathol. Biol. 2014, 62, 91–95. [Google Scholar] [CrossRef]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular matrix and dermal fibroblast function in the healing wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef]
- Rousselle, P.; Montmasson, M.; Garnier, C. Extracellular matrix contribution to skin wound re-epithelialization. Matrix Biol. 2019, 75, 12–26. [Google Scholar] [CrossRef]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef]
- Fidler, A.L.; Boudko, S.P.; Rokas, A.; Hudson, B.G. The triple helix of collagens–an ancient protein structure that enabled animal multicellularity and tissue evolution. J. Cell Sci. 2018, 131, jcs203950. [Google Scholar] [CrossRef] [Green Version]
- Har-Shai, Y.; Sela, E.; Rubinstien, I.; Lindenbaum, E.S.; Mitz, V.; Hirshowitz, B. Computerized morphometric quantitation of elastin and collagen in SMAS and facial skin and the possible role of fat cells in SMAS viscoelastic properties. Plast. Reconstr. Surg. 1998, 102, 2466–2470. [Google Scholar] [CrossRef]
- Uitto, J.; Olsen, D.R.; Fazio, M.J. Extracellular matrix of the skin: 50 years of progress. J. Investig. Dermatol. 1989, 92, S61–S77. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Garva, R.; Pickard, A.; Yeung, C.Y.C.; Mallikarjun, V.; Swift, J.; Holmes, D.F.; Calverley, B.; Lu, Y.; Adamson, A.; et al. Circadian control of the secretory pathway maintains collagen homeostasis. Nat. Cell Biol. 2020, 22, 74–86. [Google Scholar] [CrossRef]
- Weinstein, G.D.; Boucek, R.J. Collagen and elastin of human dermis. J. Investig. Dermatol. 1960, 35, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Uitto, J.; Paul, J.L.; Brockley, K.; Pearce, R.H.; Clark, J.G. Elastic fibers in human skin: Quantitation of elastic fibers by computerized digital image analyses and determination of elastin by radioimmunoassay of desmosine. Lab. Investig. J. Tech. Methods Pathol. 1983, 49, 499–505. [Google Scholar]
- Matsuoka, L.Y.; Wortsman, J.; Uitto, J.; Hashimoto, K.; Kupchella, C.E.; Eng, A.M.; Dietrich, J.E. Altered skin elastic fibers in hypothyroid myxedema and pretibial myxedema. Arch. Intern. Med. 1985, 145, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sephel, G.C.; Davidson, J.M. Elastin production in human skin fibroblast cultures and its decline with age. J. Investig. Dermatol. 1986, 86, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Nieves, D.; Chaikof, E.L. Collagen and Elastin Biomaterials for the Fabrication of Engineered Living Tissues. ACS Biomater. Sci. Eng. 2017, 3, 694–711. [Google Scholar] [CrossRef]
- Amadeu, T.P.; Braune, A.S.; Porto, L.C.; Desmoulière, A.; Costa, A.M.A. Fibrillin-1 and elastin are differentially expressed in hypertrophic scars and keloids. Wound Repair Regen. 2004, 12, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Anitua, E.; Andia, I.; Ardanza, B.; Nurden, P.; Nurden, A.T. Autologous platelets as a source of proteins for healing and tissue regeneration. Thromb. Haemost. 2004, 91, 4–15. [Google Scholar] [CrossRef]
- Baumann, L.; Bernstein, E.F.; Weiss, A.S.; Bates, D.; Humphrey, S.; Silberberg, M.; Daniels, R. Clinical relevance of elastin in the structure and function of skin. Aesthetic Surg. J. Open Forum 2021, 3, ojab019. [Google Scholar] [CrossRef]
- Swee, M.H.; Parks, W.C.; Pierce, R.A. Developmental Regulation of Elastin Production. Expression of Tropoelastin Pre-Mrna Persists after Down-Regulation of Steady-State mRNA Levels. J. Biol. Chem. 1995, 270, 14899–14906. [Google Scholar] [CrossRef] [Green Version]
- Mecham, R.P. Regulation of Matrix Accumulation; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Davidson, J.M. Biochemistry and turnover of lung interstitium. Eur. Respir. J. 1990, 3, 1048–1063. [Google Scholar]
- Rucker, R.B.; Dubick, M.A. Elastin metabolism and chemistry: Potential roles in lung development and structure. Environ. Health Perspect. 1984, 55, 179. [Google Scholar] [CrossRef]
- Shapiro, S.D.; Endicott, S.K.; Province, M.A.; Pierce, J.A.; Campbell, E.J. Marked longevity of human lung parenchymal elastic fibers deduced from prevalence of D-aspartate and nuclear weapons-related radiocarbon. J. Clin. Investig. 1991, 87, 1828. [Google Scholar] [CrossRef] [Green Version]
- Pignatelli, P.; Pulcinelli, F.M.; Lenti, L.; Gazzaniga, P.P.; Violi, F. Hydrogen peroxide is involved in collagen-induced platelet activation. Blood 1998, 91, 484–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, A.R.; Sands, D.; Sandberg, E.; Seitz, R.; Barrowcliffe, T.W. A multi-centre collaborative study on the potency estimation of ReFacto. Thromb. Haemost. 2003, 90, 1088–1093. [Google Scholar] [PubMed]
- Diegelmann, R.F.; Evans, M.C. The wound healing process: An overview of acute, fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef]
- Broughton, G.; Janis, J.E.; Attinger, C.E. The basic science of wound healing. Plast. Reconstr. Surg. 2006, 117 (Suppl. 7), 12S–34S. [Google Scholar] [CrossRef]
- Blair, P.; Flaumenhaft, R. Platelet α-granules: Basic biology and clinical correlates. Blood Rev. 2009, 23, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Montgomery, R.R. Platelets as delivery systems for disease treatments. Adv. Drug Deliv. Rev. 2010, 62, 1196–1203. [Google Scholar] [CrossRef] [Green Version]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Clark, R.A. Fibrin and wound healing. Ann. N. Y. Acad. Sci. 2001, 936, 355–367. [Google Scholar] [CrossRef]
- Adams, S.A.; Kelly, S.L.; Kirsch, R.E.; Robson, S.C.; Shephard, E.G. Role of neutrophil membrane proteases in fibrin degradation. Blood Coagul. Fibrinolysis 1995, 6, 693–702. [Google Scholar] [CrossRef]
- Ahmann, K.A.; Weinbaum, J.S.; Johnson, S.L.; Tranquillo, R.T. Fibrin degradation enhances vascular smooth muscle cell proliferation and matrix deposition in fibrin-based tissue constructs fabricated in vitro. Tissue Eng. Part A 2010, 16, 3261–3270. [Google Scholar] [CrossRef] [Green Version]
- Gray, A.J.; Bishop, J.E.; Reeves, J.T.; Mecham, R.P.; Laurent, G.J. Partially degraded fibrin (ogen) stimulates fibroblast proliferation in vitro. Am. J. Respir. Cell Mol. Biol. 1995, 12, 684–690. [Google Scholar] [CrossRef]
- Dvorak, H.F.; Harvey, V.S.; Estrella, P.; Brown, L.F.; McDonagh, J.; Dvorak, A.M. Fibrin containing gels induce angiogenesis. Implications for tumor stroma generation and wound healing. Lab. Investig. 1987, 57, 673–686. [Google Scholar]
- Pierce, G.F.; Mustoe, T.A.; Lingelbach, J.; Masakowski, V.R.; Griffin, G.L.; Senior, R.M.; Deuel, T.F. Platelet-derived growth factor and transforming growth factor-beta enhance tissue repair activities by unique mechanisms. J. Cell Biol. 1989, 109, 429–440. [Google Scholar] [CrossRef]
- Pierce, G.F.; Mustoe, T.A.; Altrock, B.W.; Deuel, T.F.; Thomason, A. Role of platelet-derived growth factor in wound healing. J. Cell. Biochem. 1991, 45, 319–326. [Google Scholar] [CrossRef]
- Kim, L.R.; Whelpdale, K.; Zurowski, M.; Pomeranz, B. Sympathetic denervation impairs epidermal healing in cutaneous wounds. Wound Repair Regen. 1998, 6, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.M.; Hunt, D.A.; Wakefield, L.M.; McCartney-Francis, N.; Wahl, L.M.; Roberts, A.B.; Sporn, M.B. Transforming growth factor type beta induces monocyte chemotaxis and growth factor production. Proc. Natl. Acad. Sci. USA 1987, 84, 5788–5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ather, S.; Harding, K.G.; Tate, S.J. Wound management and dressings. In Advanced Textiles for Wound Care; Woodhead Publishing: Sawston, UK, 2019; pp. 1–22. [Google Scholar]
- Clark, R.A.; Wikner, N.E.; Doherty, D.E.; Norris, D.A. Cryptic chemotactic activity of fibronectin for human monocytes resides in the 120-kDa fibroblastic cell-binding fragment. J. Biol. Chem. 1988, 263, 12115–12123. [Google Scholar] [CrossRef]
- Brown, E.J.; Goodwin, J.L. Fibronectin receptors of phagocytes. J. Exp. Med. 1988, 167, 777–793. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A. Regulation of fibroplasia in cutaneous wound repair. Am. J. Med. Sci. 1993, 306, 42–48. [Google Scholar] [CrossRef]
- Shaw, R.J.; Doherty, D.E.; Ritter, A.G.; Benedict, S.H.; Clark, R.A.F. Adherence-dependent increase in human monocyte PDGF (B) mRNA is associated with increases in c-fos, c-jun, and EGR2 mRNA. J. Cell Biol. 1990, 111, 2139–2148. [Google Scholar] [CrossRef]
- Mascharak, S.; desJardins-Park, H.E.; Davitt, M.F.; Griffin, M.; Borrelli, M.R.; Moore, A.L.; Chen, K.; Duoto, B.; Chinta, M.; Foster, D.S.; et al. Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring. Science 2021, 372, eaba2374. [Google Scholar] [CrossRef]
- Lin, F.; Ren, X.D.; Doris, G.; Clark, R.A. Three-dimensional migration of human adult dermal fibroblasts from collagen lattices into fibrin/fibronectin gels requires syndecan-4 proteoglycan. J. Investig. Dermatol. 2005, 124, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.; Young, A.; McNaught, C.E. The physiology of wound healing. Surgery 2014, 32, 445–450. [Google Scholar] [CrossRef]
- Mu, X.; Li, Y.; Fan, G.C. Tissue-resident macrophages in the control of infection and resolution of inflammation. Shock 2021, 55, 14–23. [Google Scholar] [CrossRef]
- Mast, B.A.; Diegelmann, R.F.; Krummel, T.M.; Cohen, I.K. Scarless wound healing in the mammalian fetus. Surg. Gynecol. Obstet. 1992, 174, 441–451. [Google Scholar]
- Pohlman, T.H.; Stanness, K.A.; Beatty, P.G.; Ochs, H.D.; Harlan, J.M. An endothelial cell surface factor(s) induced in vitro by lipopolysaccharide, interleukin 1, and tumor necrosis factor-alpha increases neutrophil adherence by a CDw18-dependent mechanism. J. Immunol. 1986, 136, 4548–4553. [Google Scholar]
- Young, A.; McNaught, C.E. The physiology of wound healing. Surgery 2011, 29, 475–479. [Google Scholar] [CrossRef]
- Martin, U.; Bock, D.; Arseniev, L.; Tornetta, M.A.; Ames, R.S.; Bautsch, W.; Köhl, J.; Ganser, A.; Klos, A. The human C3a receptor is expressed on neutrophils and monocytes, but not on B or T lymphocytes. J. Exp. Med. 1997, 186, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Parani, M.; Lokhande, G.; Singh, A.; Gaharwar, A.K. Engineered nanomaterials for infection control and healing acute and chronic wounds. ACS Appl. Mater. Interfaces 2016, 8, 10049–10069. [Google Scholar] [CrossRef]
- Bar-Shavit, R.; Kahn, A.; Fenton, J.W.; Wilner, G.D. Chemotactic response of monocytes to thrombin. J. Cell Biol. 1983, 96, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Janis, J.; Harrison, B. Wound healing: Part II. Clinical applications. Plast. Reconstr. Surg. 2014, 133, 383e–392e. [Google Scholar] [CrossRef]
- Viaña-Mendieta, P.; Sánchez, M.L.; Benavides, J. Rational selection of bioactive principles for wound healing applications: Growth factors and antioxidants. Int. Wound J. 2022, 19, 100–113. [Google Scholar] [CrossRef]
- Pierce, G.F.; Berg, J.V.; Rudolph, R.; Tarpley, J.; Mustoe, T.A. Platelet-derived growth factor-BB and transforming growth factor beta 1 selectively modulate glycosaminoglycans, collagen, and myofibroblasts in excisional wounds. Am. J. Pathol. 1991, 138, 629. [Google Scholar]
- Ramasastry, S.S. Acute wounds. Clin. Plast. Surg. 2005, 32, 195–208. [Google Scholar] [CrossRef]
- Szpaderska, A.M.; Zuckerman, J.D.; DiPietro, L.A. Differential injury responses in oral mucosal and cutaneous wounds. J. Dent. Res. 2003, 82, 621–626. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Baram, D.; Mekori, Y.A. Mast cells. Physiol. Rev. 1997, 77, 1033–1079. [Google Scholar] [CrossRef]
- Artuc, M.; Hermes, B.; Stckelings, U.M.; Grützkau, A.; Henz, B.M. Mast cells and their mediators in cutaneous wound healing? active participants or innocent bystanders? Exp. Dermatol. 1999, 8, 1–16. [Google Scholar] [CrossRef]
- Martin, P.; Leibovich, S.J. Inflammatory cells during wound repair: The good, the bad and the ugly. Trends Cell Biol. 2005, 15, 599–607. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A review of the contribution of mast cells in wound healing: Involved molecular and cellular mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef]
- Gethin, G. Understanding the inflammatory process in wound healing. Br. J. Community Nurs. 2012, 17. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Tonnesen, M.G.; Feng, X.; Clark, R.A. Angiogenesis in wound healing. J. Investig. Dermatol. Symp. Proc. 2000, 5, 40–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eppley, B.L.; Pietrzak, W.S.; Blanton, M. Platelet-rich plasma: A review of biology and applications in plastic surgery. Plast. Reconstr. Surg. 2006, 118, 147e–159e. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häkkinen, L.; Larjava, H.; Koivisto, L. Granulation tissue formation and remodeling. Endod. Top. 2011, 24, 94–129. [Google Scholar] [CrossRef]
- Stadelmann, W.K.; Digenis, A.G.; Tobin, G.R. Physiology and healing dynamics of chronic cutaneous wounds. Am. J. Surg. 1998, 176, 26S–38S. [Google Scholar] [CrossRef]
- Raghunath, M.; Bachi, T.; Meuli, M.; Altermatt, S.; Gobet, R.; Bruckner-Tuderman, L.; Steinmann, B. Fibrillin and elastin expression in skin regenerating from cultured keratinocyte autografts: Morphogenesis of microfibrils begins at the dermo-epidermal junction and precedes elastic fiber formation. J. Investig. Dermatol. 1996, 106, 1090–1095. [Google Scholar] [CrossRef] [Green Version]
- Sarojini, H.; Bajorek, A.; Wan, R.; Wang, J.; Zhang, Q.; Billeter, A.T.; Chien, S. Enhanced Skin Incisional Wound Healing with Intracellular ATP Delivery via Macrophage Proliferation and Direct Collagen Production. Front. Pharmacol. 2021, 12, 594586. [Google Scholar] [CrossRef]
- Lee, J.H.; Parthiban, P.; Jin, G.Z.; Knowles, J.C.; Kim, H.W. Materials roles for promoting angiogenesis in tissue regeneration. Prog. Mater. Sci. 2021, 117, 100732. [Google Scholar] [CrossRef]
- Noskovicova, N.; Hinz, B.; Pakshir, P. Implant fibrosis and the underappreciated role of myofibroblasts in the foreign body reaction. Cells 2021, 10, 1794. [Google Scholar] [CrossRef]
- Guido, S.; Tranquillo, R.T. A methodology for the systemic and quantitative study of cell contact guidance in oriented collagen gels. J. Cell Sci. 1993, 105, 317–331. [Google Scholar] [CrossRef]
- Gailit, J.; Clark, R.A. Wound repair in the context of extracellular matrix. Curr. Opin. Cell Biol. 1994, 6, 717–725. [Google Scholar] [CrossRef]
- Assunção, M.; Yiu, C.H.K.; Wan, H.Y.; Wang, D.; Tuan, R.S.; Blocki, A. Hyaluronic acid drives mesenchymal stromal cell-derived extracellular matrix assembly by promoting fibronectin fibrillogenesis. J. Mater. Chem. B 2021, 9, 7205–7215. [Google Scholar] [CrossRef]
- Bainbridge, P. Wound healing and the role of fibroblasts. J. Wound Care 2013, 22, 407–411. [Google Scholar]
- Banks, R.E.; Forbes, M.A.; Kinsey, S.E.; Stanley, A.; Ingham, E.; Walters, C.; Selby, P.J. Release of the angiogenic cytokine vascular endothelial growth factor (VEGF) from platelets: Significance for VEGF measurements and cancer biology. Br. J. Cancer 1998, 77, 956. [Google Scholar] [CrossRef] [Green Version]
- DiPietro, L.A. Angiogenesis and scar formation in healing wounds. Curr. Opin. Rheumatol. 2013, 25, 87–91. [Google Scholar] [CrossRef]
- Pettet, G.J.; Byrne, H.M.; McElwain, D.L.S.; Norbury, J. A model of wound-healing angiogenesis in soft tissue. Math. Biosci. 1996, 136, 35–63. [Google Scholar] [CrossRef]
- Kurz, H. Physiology of angiogenesis. J. Neuro-Oncol. 2000, 50, 17–35. [Google Scholar] [CrossRef]
- Hunt, T.K. The physiology of wound healing. Ann. Emerg. Med. 1988, 17, 1265–1273. [Google Scholar] [CrossRef]
- Ghani, Q.P.; Wagner, S.; Becker, H.D.; Hunt, T.K.; Hussain, M.Z. Regulatory role of lactate in wound repair. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2004; Volume 381, pp. 565–575. [Google Scholar]
- Shen, Y.; Jiang, L.; Wen, P.; Ye, Y.; Zhang, Y.; Ding, H.; Luo, J.; Xu, L.; Zen, K.; Zhou, Y.; et al. Tubule-derived lactate is required for fibroblast activation in acute kidney injury. Am. J. Physiol.-Ren. Physiol. 2020, 318, F689–F701. [Google Scholar] [CrossRef]
- Schultz, G.S.; White, M.; Mitchell, R.; Brown, G.; Lynch, J.; Twardzik, D.R.; Todaro, G.J. Epithelial wound healing enhanced by transforming growth factor-alpha and vaccinia growth factor. Science 1987, 235, 350–352. [Google Scholar] [CrossRef]
- Yates, R.A.; Nanney, L.B.; Gates, R.E.; King, L.E. Epidermal growth factor and related growth factors. Int. J. Dermatol. 1991, 30, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Kurkinen, M.; Vaheri, A.N.T.T.I.; Roberts, P.J.; Stenman, S. Sequential appearance of fibronectin and collagen in experimental granulation tissue. Lab. Investig. 1980, 43, 47–51. [Google Scholar] [PubMed]
- Coelho, N.M.; McCulloch, C.A. Contribution of collagen adhesion receptors to tissue fibrosis. Cell Tissue Res. 2016, 365, 521–538. [Google Scholar] [CrossRef]
- Lee, Y.I.; Lee, S.G.; Kim, J.; Choi, S.; Jung, I.; Lee, J.H. Proteoglycan Combined with Hyaluronic Acid and Hydrolyzed Collagen Restores the Skin Barrier in Mild Atopic Dermatitis and Dry, Eczema-Prone Skin: A Pilot Study. Int. J. Mol. Sci. 2021, 22, 10189. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Overall, C.M.; López-Otín, C. Strategies for MMP inhibition in cancer: Innovations for the post-trial era. Nat. Rev. Cancer 2002, 2, 657–672. [Google Scholar] [CrossRef]
- Chen, P.; Parks, W.C. Role of matrix metalloproteinases in epithelial migration. J. Cell. Biochem. 2009, 108, 1233–1243. [Google Scholar] [CrossRef]
- Martins, V.L.; Caley, M.; O’Toole, E.A. Matrix metalloproteinases and epidermal wound repair. Cell Tissue Res. 2013, 351, 255–268. [Google Scholar] [CrossRef]
- Welch, M.P.; Odland, G.F.; Clark, R.A. Temporal relationships of F-actin bundle formation, collagen and fibronectin matrix assembly, and fibronectin receptor expression to wound contraction. J. Cell Biol. 1990, 110, 133–145. [Google Scholar] [CrossRef]
- Kumar, I.; Staton, C.A.; Cross, S.S.; Reed, M.W.R.; Brown, N.J. Angiogenesis, vascular endothelial growth factor and its receptors in human surgical wounds. Br. J. Surg. 2009, 96, 1484–1491. [Google Scholar] [CrossRef]
- Abercrombie, M.; Flint, M.H.; James, D.W. Wound contraction in relation to collagen formation in scorbutic guinea-pigs. Development 1956, 4, 167–175. [Google Scholar] [CrossRef]
Figure 1.
Schematic of the classical wound healing cascade with important stages of cellular infiltration and protein deposition.
Figure 1.
Schematic of the classical wound healing cascade with important stages of cellular infiltration and protein deposition.

Figure 2.
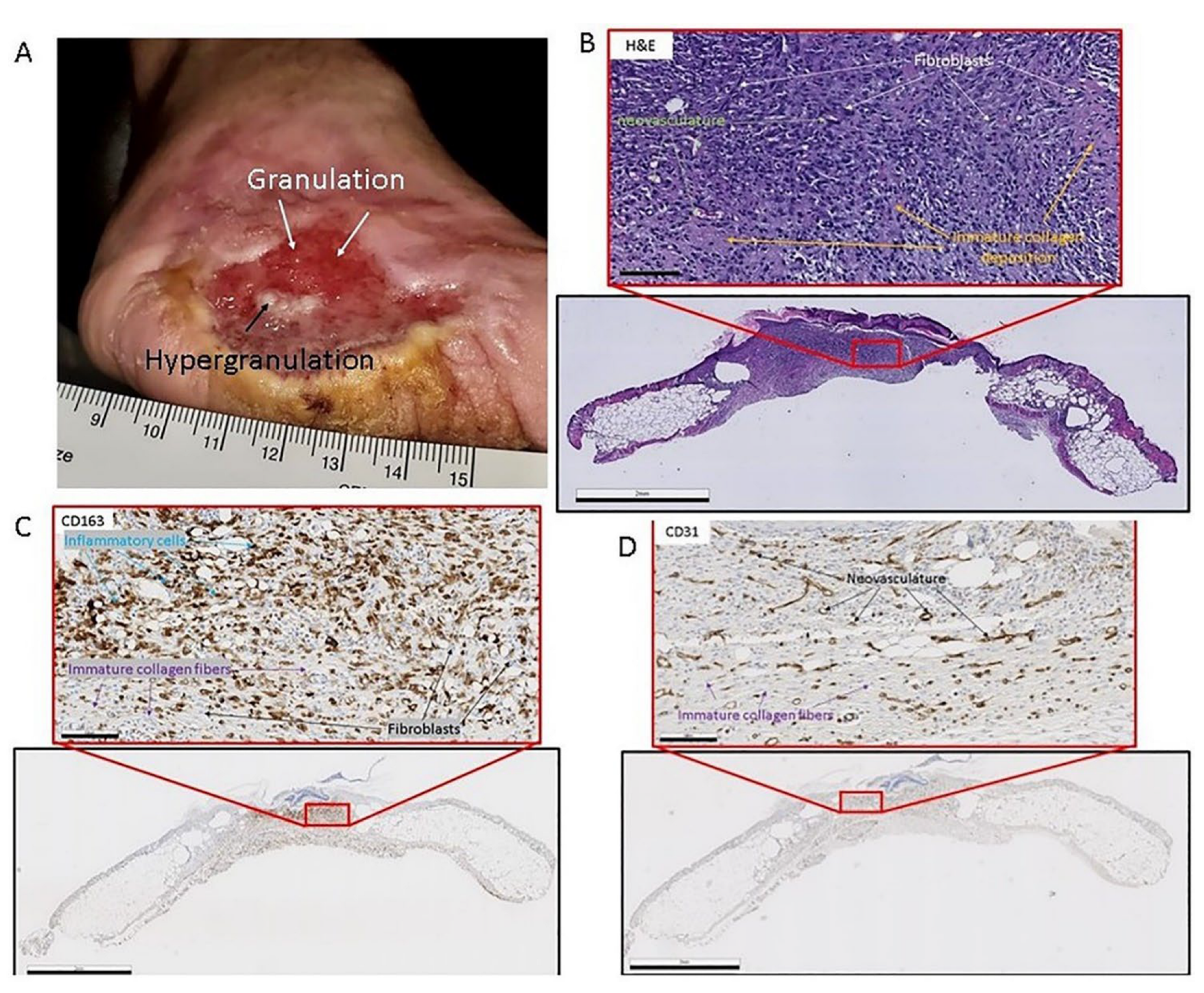
(A) Clinical image of chronic diabetic injury with granulation tissue presence. The wound is on the medial aspect of the foot. Notice the white arrow pointing to red tissue indicating cellular infiltrate and neovascularization. The black arrow is pointing to the area in the middle of the wound which is indicative of hyper-granulation. (Patient consent provided). (B) Heamatoxalin and Eosin-stained murine skin the black frame is a gross image (scale bar = 2 mm) of the skin sample, and the red framed image (scale bar = 100 µm) is a magnified image of the area in the small red box. The white arrows point to fibroblasts in the granulation tissue. The orange arrows point to immature collagen deposition. The green arrows point to neovasculature. (C) CD-163 (Inflammatory cell marker) reacted immunohistochemistry-stained murine skin the black frame is a gross image (scale bar = 2 mm) of the skin sample and the red framed image (scale bar = 100 µm) is a magnified image of the area in the small red box. The black arrows point to fibroblasts in the granulation tissue. The purple arrows point to immature collagen deposition. The blue arrows point to inflammatory cells: macrophages, and monocytes. (D) CD-31 (endothelial cell marker) reacted immunohistochemistry-stained murine skin, the black frame is a gross image (scale bar = 2 mm) of the skin sample, and the red framed image (scale bar = 100 µm) is a magnified image of the area in the small red box. The black arrows point to neovasculature in the granulation tissue. The purple arrows point to immature collagen deposition.
Figure 2.
(A) Clinical image of chronic diabetic injury with granulation tissue presence. The wound is on the medial aspect of the foot. Notice the white arrow pointing to red tissue indicating cellular infiltrate and neovascularization. The black arrow is pointing to the area in the middle of the wound which is indicative of hyper-granulation. (Patient consent provided). (B) Heamatoxalin and Eosin-stained murine skin the black frame is a gross image (scale bar = 2 mm) of the skin sample, and the red framed image (scale bar = 100 µm) is a magnified image of the area in the small red box. The white arrows point to fibroblasts in the granulation tissue. The orange arrows point to immature collagen deposition. The green arrows point to neovasculature. (C) CD-163 (Inflammatory cell marker) reacted immunohistochemistry-stained murine skin the black frame is a gross image (scale bar = 2 mm) of the skin sample and the red framed image (scale bar = 100 µm) is a magnified image of the area in the small red box. The black arrows point to fibroblasts in the granulation tissue. The purple arrows point to immature collagen deposition. The blue arrows point to inflammatory cells: macrophages, and monocytes. (D) CD-31 (endothelial cell marker) reacted immunohistochemistry-stained murine skin, the black frame is a gross image (scale bar = 2 mm) of the skin sample, and the red framed image (scale bar = 100 µm) is a magnified image of the area in the small red box. The black arrows point to neovasculature in the granulation tissue. The purple arrows point to immature collagen deposition.

Figure 3.
Phases of the wound healing response including hemostasis (coagulation), inflammation, proliferation, and tissue remodeling (Designua/Shutterstock.com).
Figure 3.
Phases of the wound healing response including hemostasis (coagulation), inflammation, proliferation, and tissue remodeling (Designua/Shutterstock.com).


{kind=link}
{kind=link}
{kind=link}
Table 1.
A list of wound healing products created using ECM proteins and a list of medical devices manufactured from synthetic polymers, intended to mimic the ECM. These lists are not meant to be all inclusive, but instead provided the reader with several examples of products which utilize ECM proteins to assist in wound healing, and products which attempt to mimic the structure of the dermis to influence wound healing.
Table 1.
A list of wound healing products created using ECM proteins and a list of medical devices manufactured from synthetic polymers, intended to mimic the ECM. These lists are not meant to be all inclusive, but instead provided the reader with several examples of products which utilize ECM proteins to assist in wound healing, and products which attempt to mimic the structure of the dermis to influence wound healing.
| Products Manufactured Utilizing ECM Proteins | ||||
|---|---|---|---|---|
| Product Name | Source | Composition | ECM Proteins | Reference(s) |
| Acell Wound Powder | Porcine Urinary Bladder | Powdered Urinary Bladder Extracellular Matrix | Collagen I, Collagen III, and Collagen IV | [27] |
| AlloDerm | Human Dermis | Human acellular, lyophilized dermal matrix | collagen, elastin, basement membrane, hyaluronic acid glycosaminoglycan (GAG) | [28,29,30] |
| AlloPatch | Human Cadaveric Dermis | aseptically processed human reticular dermal tissue | collagen, elastin, basement membrane, hyaluronic acid glycosaminoglycan (GAG) | [29] |
| Axolotl DualGraft | Human Amnion | Dehydrated amniotic membrane | Collagen I, Collagen II, Collagen IV, Collagen V, Collagen VI, Proteoglycans, Fibronectin, Laminin | [31] |
| GraftJacket | Human cadaveric skin | Cryogenicaly preserved decellularized human dermis | Collagen and elastin | [32,33] |
| MatriDerm | Bovine Ligamentum nuchae | 3D matrix of type 1 collagen fiber coated with a 3% a-elastin hydrolysate | collagen fibrils types I, III, and V and alpha-elastin | [34,35] |
| OASIS | Porcine jejunum submucosa (PSIS) | Lyophilized small intestine submucosa | Collagen I, Collagen III, Collagen IV, Collagen VI, fibronectin, elastin, hyaluronan, chondroitin sulfate, decorin | [36,37] |
| Pelnac | Porcine Achilles tendon | Collagen sponge porous matrix | Collagen | [34] |
| PalinGen Membrane | Human Amnion | Dehydrated amniotic membrane | Collagen I, Collagen II, Collagen IV, Collagen V, Collagen VI, Proteoglycans, Fibronectin, Laminin | [31] |
| Permacol | Porcine Dermis | Collagen and Elastin crosslinked by diisocynate | Collagen I, Elastin | [38] |
| Promogran | Bovine Dermis | 55% Bovine Dermal Collagen 45% Oxidized regenerated cellulose | Collagen | [39] |
| SimliDerm | Human Cadaveric Dermis | pre-hydrated human acellular dermal matrix | Collagen, elastin, basement membrane, hyaluronic acid glycosaminoglycan (GAG) | [30] |
| Xcellistem | Porcine Spleen and Lung | wound powder composed of a blend of multiple porcine-based extracellular matrix material | Collagen I, Collagen III, Collagen IV, Sulfated glycosaminoglycan, Hyaluronic acid, Lipids, Elastin, Fibronectin, and Laminin | [40] |
| Novosorb | Synthetic polyers | polyurethane foam made from ethyl lysine diisocyanate, lactic acid/ethylene glycol chain extender, and PCL1000 polyol with a removable polyurethane overlayer | None | [41] |
| Phoenix Wound Matrix | Sythetic polymers | Electrospun nonwoven fibrous three-dimensional matrix comprised of Poly(lactide-co-caprolactone) and Polyglycolic acid fibers | None | [42] |
| Restrata Wound Matrix | Synthetic polymers | electrospun mat of polyglactin 910 (PGLA) and polydiaxonone (PDO) fibers | None | [41] |
Table 2.
Listing of proteins found in ECM, percent composition, and the general functions within the ECM of the integument. Note that varying degrees of composition can occur because of the location of ECM.
Table 2.
Listing of proteins found in ECM, percent composition, and the general functions within the ECM of the integument. Note that varying degrees of composition can occur because of the location of ECM.
| Protein | Percent Composition | Function(s) |
|---|---|---|
| Collagen(s) | 50–90 | Synthesized by fibroblasts. Gives structural and tissue integrity, aids in epidermal/dermal differentiation. |
| Elastin | 0.6–7.9 | Creates an intricate network for structural support allowing for elasticity of tissue. |
| Fibronectin | <1.0 | Involved in wound healing including platelet spreading and leukocyte migration to injured tissue(s). Aids in promotion of elastin deposition and mechanical strength of ECM. |
| Laminin | <1.0 | A glycoprotein that is a part of the basal lamina, aids in cellular signaling. |
| Vitronectin | <1.0 | A glycoprotein involved in hemostasis and cellular adhesion during tissue damage. |
| Tenascin | <1.0 | Glycoprotein family that aids in cellular migration adhesions and cell proliferation. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Diller, R.B.; Tabor, A.J. The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics 2022, 7, 87. https://doi.org/10.3390/biomimetics7030087
AMA Style
Diller RB, Tabor AJ. The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics. 2022; 7(3):87. https://doi.org/10.3390/biomimetics7030087
Chicago/Turabian StyleDiller, Robert B., and Aaron J. Tabor. 2022. "The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review" Biomimetics 7, no. 3: 87. https://doi.org/10.3390/biomimetics7030087