1. Introduction
The 5-years DIVERSIFY project [
1] identified a number of new/emerging finfish species with a great potential for the expansion of the aquaculture industry. Six species were selected based both on their biological and economical potential: Meager (
Argyrosomus regius), greater amberjack (
Seriola dumerili), wreckfish (
Polyprion americanus), Atlantic halibut (
Hippoglossus hippoglossus), grey mullet (
Mugil cephalus), and pikeperch (
Sanders lucioperca). This new/emerging species are fast growing and/or a large species marketed at a large size and can be processed into a range of products to provide to the consumer.
Wreckfish (
Polyprion americanus) is one of the largest serranid species, reaching a size of 100 kg. It is a deep-water fish found almost throughout the world and is characterized by an extended pelagic juvenile phase [
2,
3,
4].It is one interesting new species for aquaculture, due to its fast growth [
5,
6], late reproductive maturation [
2], high market price and limited fisheries landings—quotas have been reduced by 90% in 2012 in the U.S.A. [
7] and 80% in 2017 in Galicia, Spain [
8]—and easy manipulation in captivity [
6,
9]. Its large size lends itself to processing and development of the value-added products.
Wreckfish acclimatizes easily to captivity and, despite its large size, no mortalities have been reported due to handling. It accepts commercial feed easily, being a very voracious carnivore. In a study of wild-caught individuals it was shown that the fish grew from 1 kg to 5 kg in a period of 10 months [
6]. The slow reproductive maturation, which occurs at an age of 5–10 years in captivity, may be a problem for broodstock development and management. On the contrary, its long juvenile stage is a great advantage from the aquaculture viewpoint, allowing for commercialization before sexual maturity, and thus avoiding problems linked to maturation, such as reduction in growth, or loss of flesh quality and organoleptic properties. It has been demonstrated that growth is strongly influenced by sex and that wreckfish females are significantly heavier than males, as observed in many other marine fish species [
10].
There are three genetically different subpopulations [
2]: North Atlantic and Mediterranean Sea, Brazil, and South Pacific. Wreckfish is a deep-water fish found almost throughout the world. The first part of their life phase (from hatching to a body length about 60 cm) is pelagic and juveniles live associate with floating debris near the coast. Demersal wreckfish individuals inhabit rocky and muddy bottoms at depths of 40–200 m, however, individuals are frequently found in waters deeper than 300 m with a maximum recorded depth of 1000 m [
11]. It is a gonochoristic species with no sexual dimorphism and spawns at the continental slope at depths of 300–500 m, with the formation of spawning aggregations [
12].
In Galicia, Spain, the cities of Vigo and A Coruña are the two main ports for wreckfish sales and most of the catches came from the Azores fishery. The price varied in last ten years between 13–22 €/Kg (
Figure 1).
The establishment of methods for the control of spawning and the production of good quality eggs are essential for the culture of any animal species. The description of the reproductive cycle, along with allowing for the identification of the spawning period and spawning preferences of each species (temperature and photoperiod), enables the recognition of possible reproductive dysfunctions and leads to the development of protocols for spawning induction and production of viable eggs [
13].
The lack of reproductive control and larval rearing protocols have been the major bottlenecks preventing wreckfish aquaculture thus far. Limited egg collection has been achieved from captive spawners using hormonal induction [
9] or stripping [
14]. Embryonic development and the early life stages have been described [
9,
13], indicating that the large egg size of this fish (~2 mm in diameter) may offer significant advantages for its larval rearing. Reproduction and larval rearing of a very close relative, the hapuku (
Polyprion oxygeneios) has been achieved recently in New Zealand [
15,
16,
17,
18]. The scarcity of broodstock is a disadvantage for this fish, but the clear biological and economical potential of this species justified the invested effort to overcome its documented bottlenecks (i.e., reproduction and larval rearing) in order to produce appropriate numbers of juveniles to launch commercial production.
In the present work, we provide a global information of the studies carried out during the 5-years of the DIVERSIFY project, related to the acquisition of new broodstock, control of reproduction, nutrition, and larval rearing. This information is targeted towards commercial and research organizations interested in the potential of wreckfish for aquaculture.
2. Reproduction
2.1. Acquisition of Wild Fish and Establishment of Captive Broodstocks
There are two main methods for catching wreckfish: By a net (similar to a purse seine) that surrounds a floating object with fish underneath for juveniles (weight <3 kg) directly, and by a long-line for adults (weight >3kg). The fishing season of wreckfish takes place between April and July.
The decline in wreckfish catches in Galicia it difficult to obtain individuals to increase the wreckfish broodstocks. A small number of young fish (<3kg) were captured on the surface with a net in the Atlantic coast (NW of Spain) by a coastal boat and kept in quarantine. For the adults (>3 kg), the lines were lifted very slowly from the bottom of the sea to avoid the over expansion and rupture of the swimming bladder that would produce hemorrhage, collapse of internal organs, and death of the animals in the following hours. Once the fish were on the surface, they were placed inside a tank with anesthetic (clove oil) at a concentration of 0.03 mL L
−1 [
19] and the hooks were removed. Finally, they were transferred to the facilities in a truck and kept in quarantine.
2.2. Description of the Reproductive Cycle in Captivity
From March 2015 to October 2016 the broodstocks of all the facilities were monitored and samples of oocytes, sperm, and blood were taken to describe the reproductive cycle of wreckfish in captivity. The results, published in reference [
20], showed that vitellogenesis began several months before the spawning period in wreckfish. The analysis of the ovarian biopsies in females, the sperm samples in males, and the blood samples in both sexes showed a correlation between estradiol and testosterone levels in females as vitellogenesis progressed, as in the cases of testosterone and 11- ketotestosterone that were higher in males with high rates of spermiation.
2.3. Spontaneous and Induced Spawning
The four wreckfish broodstocks at the Hellenic Center for Marine Research (HCMR), the Instituto Español de Oceanografía (IEO), the Aquarium Finisterrae (MC2) and Consellería do Mar (CMRM) facilities were monitored from January 2014 to July 2018 and the best conditions to obtain spontaneous and induction spawns for the species were evaluated. Low feeding rates were recorded during the spawning season from March 2015 to July 2015, while high feeding rates were observed during autumn in the two tanks at IEO facilities (
Figure 2). During this period, they were fed ad libitum twice a week with two different diets: The wet feed (semi-moist diet) was a mixture of white fish (15%), fish oil (15%), mussels (18%), squid (18%), and fishmeal (34%), with 64.7% and 17.3% of total proteins and lipids respectively; and a specific dry feed formulated for wreckfish broodstock (Sparos Lda, Portugal) composed by 68.2% of proteins and 12.5% of lipids.
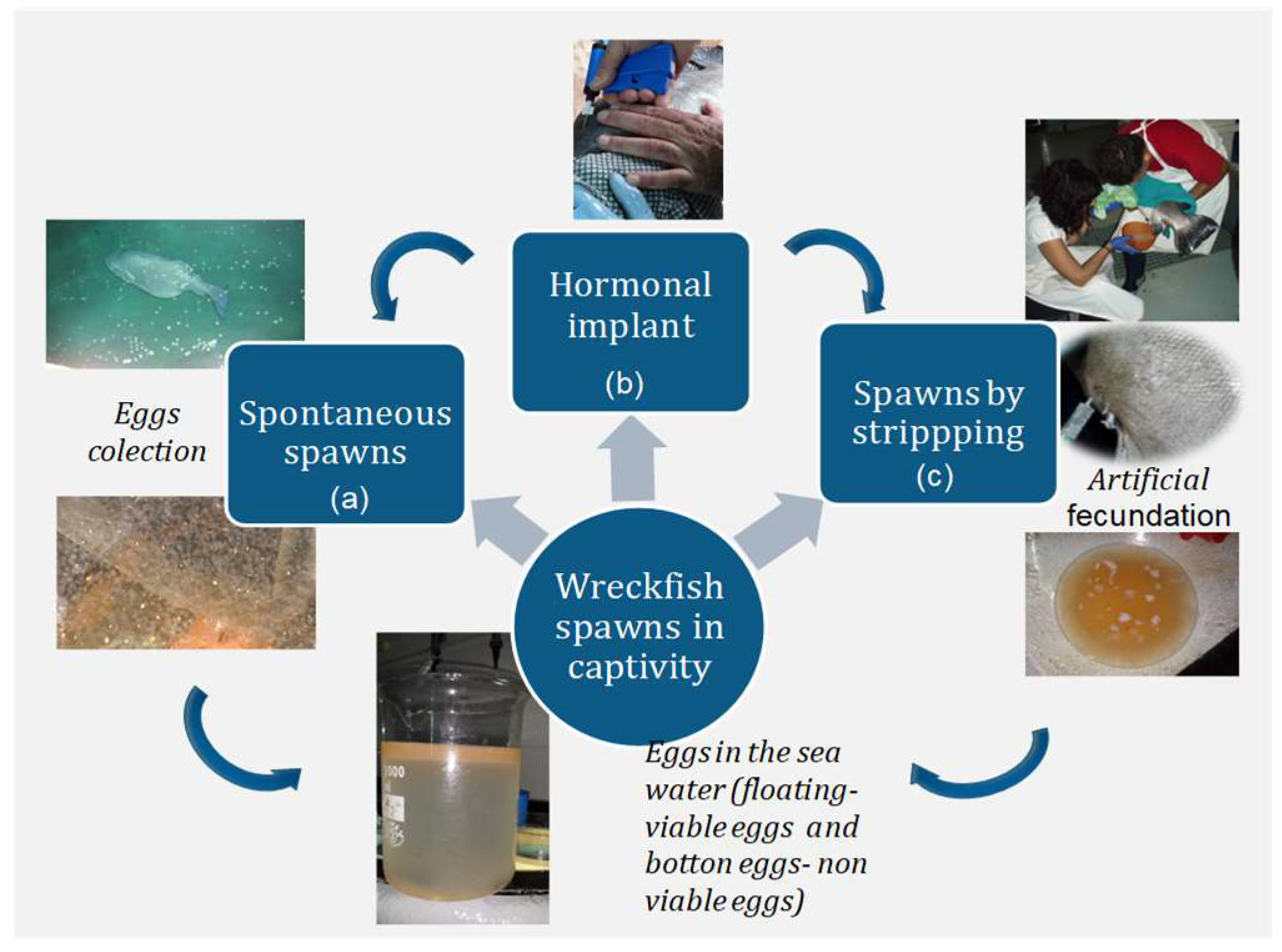
A variety of environmental conditions like photothermal regime and tank size and location were used: Indoor (IEO) and outdoor (CMRM) tanks with natural photothermal conditions, indoor tanks with simulated natural photothermal conditions (MC2) and constant temperature (HCMR). Three different strategies to obtain wreckfish spawns in captivity were tested (
Figure 3).
Natural and spontaneous spawns in large tanks (>40 m3), collecting the eggs as they exit the outflow of the tank (a).
Spawning induction with exogenous gonadotropin-releasing hormone agonist (GnRHa) (b) carried out in large tanks (>40 m3) under controlled photothermal conditions, allowing the fish to spawn spontaneously.
Spawning induction followed by in vitro fertilization by stripping of the mature females and males, maintained in smaller tanks (<40 m3) (c).
Natural vitellogenesis and oocyte maturation took place in females in captivity and the number of spontaneous spawns increased while the number of induced spawns was reduced during the last year (2018), mainly in the three Spanish stocks (
Figure 4).
The reason was probably a better adaptation of the females to the captive conditions and the promotion of a natural maturation cycle, resulting in not only vitellogenesis but also, spontaneously, oocyte maturation. The natural spawning behavior was characterized by males chasing females followed by the release of the gametes. Spawning took place in the night or very early in the morning. In 2017 and 2018, spontaneous spawns at IEO, MC2, and CMRM facilities produced a large number of fertilized eggs (
Figure 5).
Sexual maturation in males covered the same period as females, with high values of sperm concentrations between April and June and maximums of 25–35 × 109 spermatozoa mL−1.
For hormonal trials, 11 females in the three Spanish stocks were inducted with GnRHa implants between June 2015 and 2018 with doses between 25 and 100 µg kg−1 of female. The results of these treatments varied from no response in 2015, spontaneous or stripping spawns with non-viable eggs in 2016 to good fertilization results but not hatching in 2017. There was only one spawn that was successfully reared until 25 days post hatching (dph) in 2016. In 2018, after 6 days from the hormonal induction, an implanted female from MC2 facilities spawned periodically every 5 days with egg fertility higher than 50%. Similar results in spawning frequency were achieved with the GnRHa injection test a year before at the HCMR facilities. Ovarian biopsies were performed before the implants to determine the size of the oocytes. Results showed that females with oocyte size <1200 μm did not respond to the implants doses used. In addition, it was found that GnRHa injections were as effective as GnRHa implants and, as observed with the implants, a response time of approximately six days after the injection was observed.
On the other hand, the effect of recombinant sea bass gonadotropins hormones follicle-stimulating hormone (FSH), and luteinizing hormone (LH) produced by Rara Avis Biotec, Spain, has been evaluated in some wreckfish specimens (
n = 4) presenting arrest of the oogenesis in captivity. The results showed that the use of FSH and LH stimulated gonadal development [
21], but more trials with a greater number of individuals and different hormonal doses are required to verify the effects in wreckfish.
It could be possible to apply the method of artificial spawning by stripping in wreckfish only with mature females that exhibit problems of spontaneous spawns after GnRHa induction but not for females that undergo oocyte maturation naturally, because the adult fishes are large and heavy and the stress caused by handling resulted in problems with egg quality and fertilization success.
2.4. Sperm Characteristics and Cryopreservation
In DIVERSIFY, we established a Computer Assisted Sperm Analysis (CASA) for the evaluation of wreckfish sperm. This method is available as a movie describing the procedure of sperm activation and CASA analysis on the website of the project [
1]. The best adapted CASA parameters for wreckfish sperm analyses were determined and reported to optimize their abilities to check fertility potential of the semen in the course of their future spawning induction experiments. The analyses demonstrated that sperm of captive wreckfish share a common pattern of motility with both marine and freshwater fish, based on a general activation of all the sperm at the same time of ejaculation in the activating environment, then a decrease with time down to zero in a rapid lapse of time from 30 s to more than 20 min due to exhaustion of energy stores, that are not replenished due to poor respiration.
The mean concentration of wreckfish sperm was 2.41 × 10
10 (standard deviation: 0.4 × 10
10,
n=9) spermatozoa mL
−1 in Galicia in January, while it remained around 1 × 10
10 from April to September, with no significant variation between sampling dates in Crete, Greece. In 2015, the concentration reached higher values of up to 2 × 10
10 spermatozoa mL
−1. These concentrations levels did not differ from earlier data (from 1.5 to 2.71 × 10
10 spermatozoa mL
−1) from the HCMR broodstock. Finally, the spermatozoa concentration in wreckfish stripped semen was of the same order of magnitude as that of pelagic fish such as European sea bass (
Dicentrarchus labrax), gilthead sea bream (
Sparus aurata) or meagre (
Argyrosomus regius) [
22,
23,
24] and it was higher than that of sole (
Solea solea) and turbot
(Scophthalmus maximus) [
25,
26].
3. Nutrition
Nutrition studies performed in the DIVERSIFY project were focused mainly on the development of adequate live prey enrichments for wreckfish larvae and broodstock feeds for enhancing fecundity and spawn quality. These are very important for the improvement of wreckfish nutrition and culture development of this polyprionidae species.
The quality of the first feeding regimes play an important role in the success of larval culture with dietary lipids being recognized as one of the most important nutritional factors that affect larval growth and survival [
27], and an adequate broodstock nutrition is essential to get success in fish intensive culture. In marine fish, dietary lipids and, in particular, polyunsaturated fatty acids (PUFAs) play a critical role in the successful production of high-quality gametes and eggs of marine fish [
28,
29].
Marine fish have restricted ability or are unable to synthesize n-3 and n-6 LC-PUFAs from their precursors, alpha linolenic acid (18:3n-3) and linoleic acid (18:2n-6) respectively [
30,
31,
32]. Therefore, 22:6n-3 (docosahexaenoic acid, DHA), 20:5n-3 (eicosapentaenoic acid, EPA) and 20:4n-6 (arachidonic acid, ARA) are considered essential fatty acids for marine fish and must be included in their diets.
Due to the scarce information about wreckfish nutrition and with the objective of knowing the nutritional requirements, a study of the composition of different tissues (muscle, liver, and gonad) from wreckfish wild fish was done to get some basic information of this species. Furthermore, studies carried out in DIVERSIFY about the composition of wild wreckfish mature gonads and eggs from reared females were very useful as the basis to formulate the enrichment products for live prey used as feeding of wreckfish larvae and the specific dry feed for wreckfish broodstock.
The preliminary study on the biochemical composition of some tissues of wild wreckfish shows that wild wreckfish have a big amount of proteins in muscle (84.4%of dry weight, DW) and a low level of lipids (6.9%). Docosahexaenoic acid (26%), was the predominant fatty acid in the fatty acid composition of wild wreckfish muscle, similar to the one obtained by the authors in reference [
33] in the Mediterranean wreckfish with values among 24.2–25.7%. Others were oleic acid (18:1), palmitic acid (16:0), EPA (20:5n-3) and stearic acid, representing the major fatty acids in both Atlantic and Mediterranean wreckfish. A high variability among individuals was found in liver and immature gonads composition. Samples of wild males and female mature gonads were taken out showing high values of proteins (60% in females and 44% in males) and the lipid content represented 21% in females and 13% in males’ gonads, n-3 PUFA values varied between 35% and 40%, a high level of ARA was found (7–10% of total fatty acids, TFA) and the EPA/ARA ratio was nearly 1.
3.1. Effectiveness of Live Prey and the Influence of Enrichments
The development of enrichment products of live prey is very important for the success in the larval culture. The understanding of the PUFA requirements of marine fish larvae requires the definition of optimal dietary ratio of DHA, EPA, and ARA [
29].
Based on data of biochemical analyses of gonads from wild wreckfish females, eggs, and larvae obtained from reared fish, some live feed enrichment products were developed for larval wreckfish. Total lipids reached 15–19% of DW in wreckfish eggs and PUFA content reached values of 43–45% of TFA. Similar values were found in 1 to 10 dph larvae (no feeding). Three experimental enrichment products were formulated during 2017 and 2018 to meet the EPA, DHA, and ARA levels obtained from tissues of wild-cached wreckfish. For experimental enrichment preparation, a combination of different products based on microalgae was used.
The experimental enrichments were formulated using two different levels of ARA for rotifer (
Brachionus plicatilis) and one level for Artemia. The effect of the enrichment of these new products on the biochemical composition of rotifers and Artemia was evaluated and the results showed an efficient enrichment in both preys although the enrichment was less effective in Artemia than in rotifers specially in DHA as it was said recently by the authors in reference [
34] who reported that, in general, rotifers assimilated PUFA in a much higher ratio than Artemia.
Some nutritional experiments with wreckfish larvae were performed in DIVERSIFY showing that larvae exhibit, in general, a good acceptance of the enriched live prey tested and no differences in fatty acid composition of wreckfish larvae fed with the prey enriched with the enrichment products tested were found at different days of live. The fatty acid profile of wreckfish larvae along the larval development shows big amounts of PUFA, specially DHA, EPA, and ARA.
3.2. Influence of Broodstock Feeds on Fecundity and Spawning Quality
Regarding wreckfish broodstock feeding regimes, results obtained showed that most of commercial dry feeds have too much fat for wreckfish. Results obtained with dry feed specifically formulated for wreckfish broodstock (25% fish meal, 34% squid meal, 7.5% krill meal) in DIVERSIFY demonstrated that the diet must contain a big amount of proteins (60%), low level of lipids (12–13%), a high amount of n-3 PUFA (30%TFA), and the EPA/ARA ratio must be similar to that obtained in wild females.
A clear relationship between fatty acid profile of broodstock diets (semi-moisture, dry feed, and a mixture of hake and squid) and fatty acid profile of oocytes and eggs from females fed with different diets were found. Furthermore, first data on the fatty acid profile of sperm from wreckfish males of different broodstock were obtained.
A relationship was found between broodstock diets and fecundity, number of spawnings of the females, etc. Relative fecundity (n° of eggs/Kg of female) and number of spawns per female have been increasing in females fed with dry feed over the years, from 2015 to 2018.
4. Larval Husbandry
4.1. Development of the Digestive System in Wreckfish
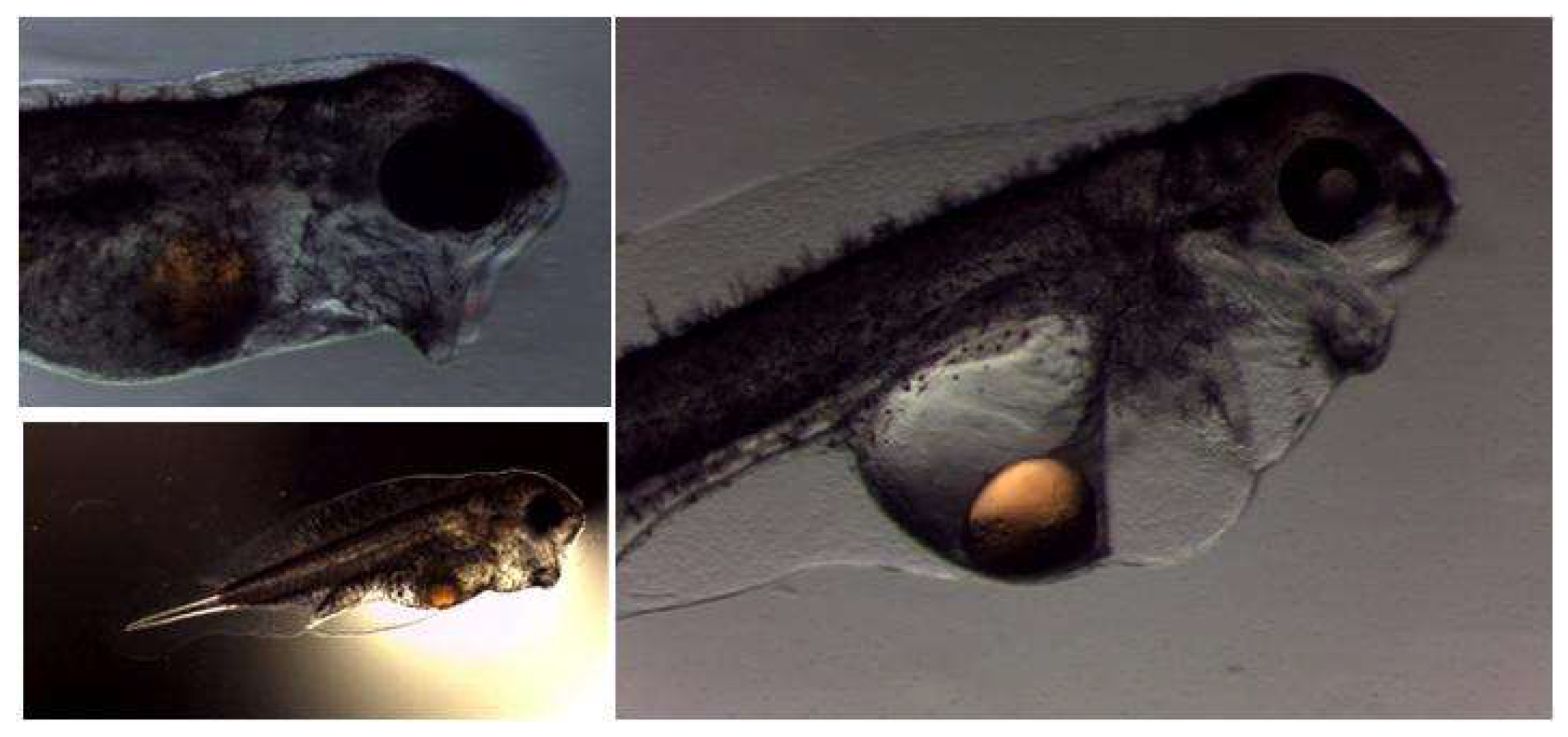
Larval rearing of wreckfish is currently the major bottleneck for the successful culture of this species, due to the low survival rates observed during this period. One of the main scientific goals for wreckfish larval rearing is the development of protocols according to the specific requirements of the larvae during the early developmental stages. The study of the development of the organs related with larval feeding behavior offers part of the necessary information for the optimization of the larval rearing protocols. During larval stages, the systems that were closely related with the feeding behavior were the vision system, by which the fish perceived the different feed items in the rearing environment, and the digestive system, which enables fish larvae to capture, ingest, digest, and absorb nutrients from the feed. These two systems and the structures of which they are composed are related with the larval rearing feeding protocols. The vision system (i.e., the eye) determines the ability of the larvae to identify the prey under the light conditions that exist in the rearing environment, whereas the digestive system was also determined by the qualitative and quantitative composition of the feeding protocol that was used during rearing.
In wreckfish, the ontogenesis of the digestive system was considered a slow procedure in comparison with other species. The development of the digestive system was controlled by endogenous factors and generally it was genetically programmed, but the time of appearance of the digestive system structures could be influenced by a number of exogenous factors, with temperature being one of the most important [
35]. The ontogenesis of the organs related to the digestive and the vision system was not completed until 23 dph (
Figure 6).
Most of the organs (except for the maxillary teeth at the upper jaw that became visible at 19 dph) appeared at 8 dph.
The ontogeny of the retina of the wreckfish was found to be similar to the general pattern shown in most fish species. At hatching, the retina was an undifferentiated and non-functional tissue, as occurs in most marine fishes with pelagic early life stages [
36,
37,
38,
39,
40,
41]. Cone cells were the first photoreceptors that appeared (6 dph). This fact indicates that at this developmental stage wreckfish larvae were able to see different items in the rearing environment only during daylight hours. Thus, it is necessary to provide light in the rearing tanks of wreckfish after 5 dph to facilitate that the fish are able to see feed items like rotifers and Artemia nauplii.
4.2. Optimum Conditions for Larval Rearing
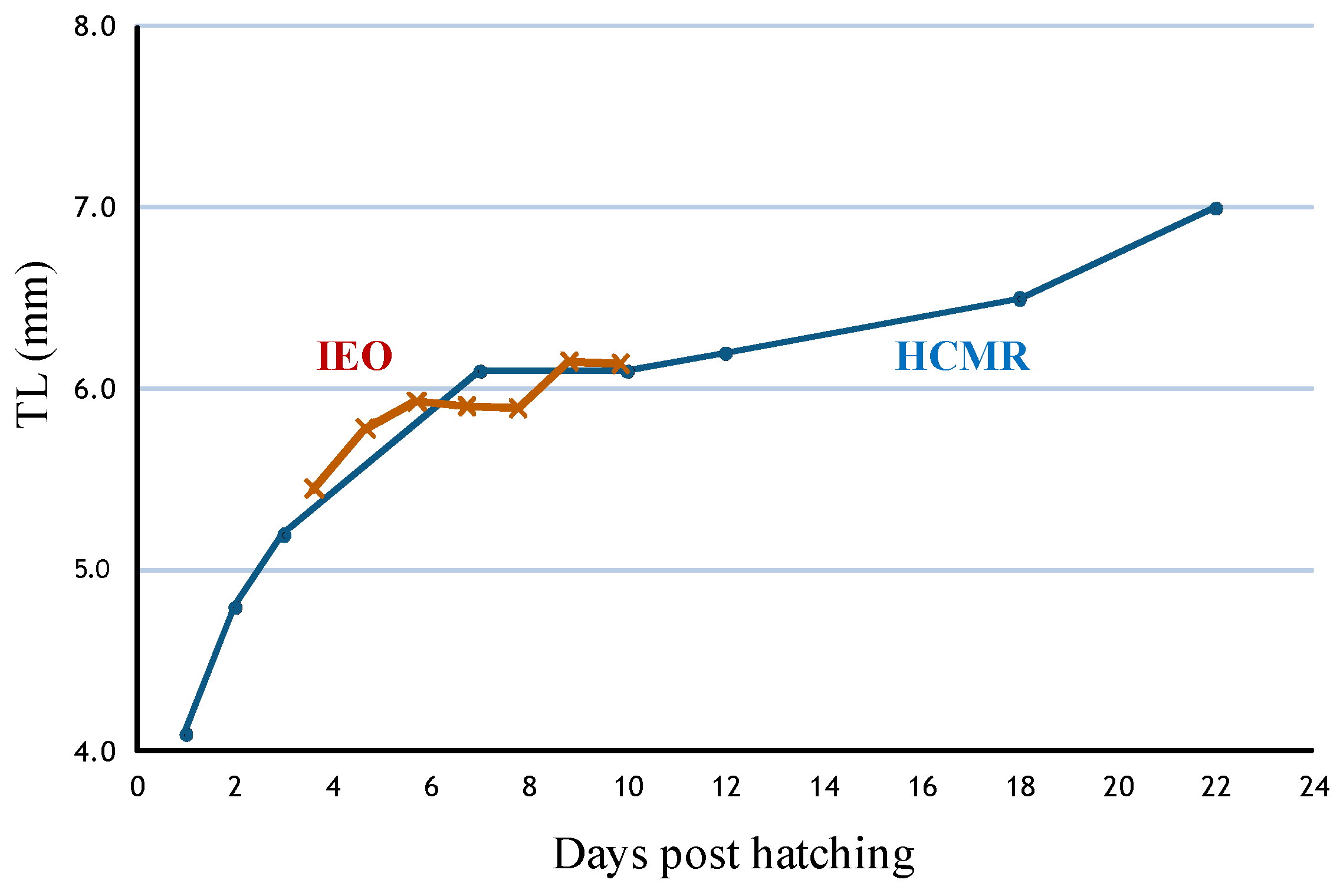
The main objectives were to develop a culture protocol and study the influence of different sea water temperatures on larval survival and growth. After incubation and during the autotrophic stage, temperature was maintained at 16.0 °C, and gradually increased afterwards to 17.5 °C. A common feeding protocol with rotifers and Artemia was applied and larvae survived until 20 dph. Growth performance until 22 dph was obtained (
Figure 7), with similar results for the Mediterranean and Atlantic stocks (MC2, CMRM and IEO) (
Figure 8). Larval length was 4.70 ± 0.27 mm at 1 dph, yolk sac consumption occurred at 11 dph at 14–17 °C and 8 dph at 17–20 °C, and the mouth opened at 7 dph and 4 dph at 14–17 °C and 17–20 °C, respectively [
42]. The large yolk sac and the large oil droplet indicated the presence of a long autotrophic larval stage.
No statistical differences (
p > 0.05) in size or weight were observed in the larval growth at different temperatures (
Figure 9).
The average length increased from 4.56 ± 0.08 mm (0 dph) to 6.02 ± 0.27 mm (26 dph). This development occurred mainly during the first 10 days of life, staying relatively constant afterwards. Growth showed a different pattern with an irregular distribution with the larvae age. Newly hatched larvae weighed 0.34 ± 0.07 mg (dry weight) and maximum values of 0.47 mg were registered at 10 dph and 26 dph.
Some malformed individuals were observed in the rearing trials (
Figure 10). The problem was identified as similar to a syndrome related to swollen yolk sac (SYSS) described in Murray cod (freshwater fish in Australia), that it is related to inadequate nutrition of the broodstock [
43]. Furthermore, similar appearance has been also described in the Blue Sac Disease that is common in trout [
44]. Toxicity from nitrogen compounds such as ammonia or oxidative stress have been suggested to play an important role in the appearance of this type of deformity. A situation of SYSS could be happening in wreckfish larvae but further studies are required to confirm this conclusion.
During the last year (2018), experimental larval rearing trials were carried out at IEO, MC2, and Instituto Galego de Formación en Acuicultura (IGAFA) facilities focused on the use of different culture systems such as flow-through, mesocosm and aquaculture recirculating system (RAS). The best results were achieved with the RAS conditions in two different batches of larvae fed with rotifers, Artemia nauplii and enriched metanauplii at the IGAFA facilities, where both reached the juvenile stage.
The study was approved by the Committee of Research Ethics of the University of Oviedo (Project identification code ES360570189801/15/FUN.01/FIS.02). The animals used in all the experiments described in this manuscript have been treated with the strictest respect and in accordance with the ethical norms approved at both the State and European level in the following legal regulations: “Royal Decree 53/2013, of February 1, which establishes the basic rules applicable for the protection of animals used in experimentation and other scientific purposes, including teaching” and “Directive 2010/63/EU of the European parliament and the council of 22 September 2010 on the protection of animals used for scientific purposes”.
5. Conclusions
Reproduction in captivity:
The spawning season covers the months of January to June, and occurs sequentially in batches, with spawned eggs having a diameter of 2 mm, as also described in other studies [
17].
The males produce large amounts of sperm with a maximum in April and June, which overlaps with the period of maturity of the females. Despite the easy handling of this species in captivity, its large size requires large volumes of sea water for its welfare in captivity and avoids stress that would affect gametogenesis and, therefore, the achievement of maturation in captivity. The stripping method seems unfavorable for wreckfish because it entails a regular handling of the breeding specimens at the time of spawning, and especially when it has been verified that the spontaneous spawnings in large-volumes tanks occurred at the scheduled time.
The positive response to the induction trials only took place in females with oocyte sizes >1200 μm.
This study complements the previous observations on the biology of this species described for wild populations [
8], and brings new knowledge about their reproduction in captivity.
Sperm characteristics:
Wreckfish males produced a high volume of easily expressible milt with a concentration considered as medium range for marine fish, and much higher than flatfish. On the top of those general features, the setup of a CASA protocol adapted to wreckfish sperm demonstrated that wreckfish sperm exhibits a high percentage of motile cells at activation, and one of the highest initial speeds recorded for fish sperm. This high speed was associated with a long swimming duration compared to other marine fish. The long duration exhibited a double trajectory shape. The first trajectory was straight (associated with the search of target eggs) and then the trajectory began bending, which was interpreted as a phase of searching for the micropyle on the egg surface.
Nutrition:
Results obtained from tissues composition of wild wreckfish were very useful to advance the knowledge of nutritional requirements of this species showing a big amount of proteins and low levels of lipids in muscle. Gonads from females of wild wreckfish have a high level of proteins, n-3 PUFA and ARA. The EPA/ARA ratio was nearly 1.
Enrichment products for live prey (rotifer and Artemia) were developed and wreckfish larvae exhibit, in general, a good acceptance of the enriched live prey. The first results of larval culture are promising, but it is necessary to continue with the research on nutritional requirements and their impact on the growth, survival, and larval quality.
A specific dry feed was formulated for wreckfish broodstock with a big amount of proteins, low level of lipids, and a high amount of n-3 PUFA. The EPA/ARA ratio must be similar to the one obtained in wild females’ gonads and a relation was found between fatty acid profiles of diet and those of the oocytes and eggs. Relative fecundity and number of spawns per female have been increasing in females fed with dry feed over the years, from 2015 to 2018.
Development of digestive and vision systems:
Wreckfish larvae were also characterized by the large size of the yolk sac and oil droplet. During the autotrophic stage the digestive system and the vision system of wreckfish larvae were developed to such an extent that larvae were able to identify, capture, and assimilate zooplanktonic organisms and this should be included to the feeding rearing protocol. As the main organs such as the gastric glands did not appear until the length of 5.5 mm, a combination of easily captured and more digestible preys as rotifers or different types of copepods in different developmental stages, have to be included in the larval rearing feeding protocol of wreckfish. The above, in combination with the optimization of the rearing conditions, such as the tank hydrodynamics, the temperature protocol during the rearing procedure, and the photic conditions in the rearing tank, is considered necessary for the development of the wreckfish larval rearing protocol. Summarizing the results of this study, it appears that after the first 23 days of rearing, the digestive system and the eye of the larvae were developed to such a degree that by that time fish were, in principle, able to detect, capture, and utilize the different types of zooplanktonic organisms.
Larval husbandry:
Results thus far suggest that the optimal water temperature for artificial incubation of wreckfish eggs is in a range of 16.5–19.5 °C. Lower temperatures (14.0 ± 0.5 °C) promote higher number of deformed larvae and lower hatching rates, with more egg mortalities during the first three days of incubation.
There are 25 juveniles that reached 150 dph that were cultured at approximately 18 °C at the IGAFA facilities and it represents a significant step forward in wreckfish larval culture and provides a basis for further studies. This was the first time in the project that we succeeded in producing juveniles weaned to feed, and it signifies a milestone in the efforts to produce wreckfish under aquaculture conditions. This trial achieved important data on growth and increased our knowledge about the feeding protocol and the specific behavior and metamorphosis of wreckfish larvae.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









