
Wound-Induced Changes in Antioxidant Enzyme Activities in Skin Mucus and in Gene Expression in the Skin of Gilthead Seabream (Sparus aurata L.)


Immunobiology for Aquaculture Group, Department of Cell Biology and Histology, Faculty of Biology, University of Murcia, 30100 Murcia, Spain
*
Author to whom correspondence should be addressed.
Fishes 2021, 6(2), 15; https://doi.org/10.3390/fishes6020015
Submission received: 22 March 2021
/
Revised: 13 April 2021
/
Accepted: 15 April 2021
/
Published: 18 April 2021
(This article belongs to the Special Issue New Insights on Fish Immunology)
Abstract
:This study investigated the antioxidant enzyme activities in the skin mucus of gilthead seabream (Sparus aurata L.) at 3 and 7 days post-wounding (dpw). The expression levels of the genes that encode stress proteins (grp170, grp94, grp75, sod and hsp70) and skin regeneration-related proteins (tf, igf1, tgfb1, der1, apo1 and erdj3) in the skin also were determined. Mucus and skin samples were obtained from the left and right flanks of non-wounded and wounded fish. In both flanks of the wounded fish, catalase and glutathione reductase activities in the skin mucus increased (p < 0.05) at 3 and 7 dpw (100 ± 31% and 111 ± 25%, respectively), whereas superoxide dismutase activity increased (p < 0.05) only at 7 dpw (135 ± 15%). The expression levels of stress proteins in the skin of the wounded flank of the wounded fish mainly increased at 7 dpw (grp170 increased to 288 ± 85%, grp94 to 502 ± 143%, grp75 to 274 ± 69%, sod to 569 ± 99%, and hsp70 increased to 537 ± 14%) (p < 0.05). However, the expression levels of the tissue regeneration-related genes varied depending on the flank investigated, on the experimental time, and on the gene studied. To the best of our knowledge, this is the first work to determine the effect of a wound in different skin parts of the same fish.
1. Introduction
Together with its associated structures, the skin constitutes the first line of defense of the immune system of vertebrates. Histologically, the skin consists of three layers: the epidermis, dermis, and hypodermis [1]. In marine organisms, apart from its role in defense, the skin plays a role in many other important functions, such as osmoregulation or as a mineral reservoir [2]. The condition of the skin is generally a key factor influencing the welfare and health of fish, which in turn are important parameters in aquaculture. In teleosts, skin wounds are quickly repaired, as a wound surface becomes rapidly covered with skin mucus and within a few hours, re-epithelization along the margin is initiated and proceeds towards the central part of the wound, as was reported for Cyprinus carpio, Hemicrhomis bimaculatus, and Sparus aurata [2,3,4]. The other important steps in the skin healing process are the following: within 1–2 days scale differentiation is initiated; at 3–5 days, the external layer of the extracellular matrix is produced; at 6–14 days, the skin produces the basal plate matrix; and after 14–28 days, mineralization of the basal plate occurs [2].
Gilthead seabream (Sparus aurata L.) was selected for this study due to its economic importance as a farmed fish species in marine aquaculture. In the last 10 years, numerous studies have focused on the skin of this important fish species. Specifically, many studies have focused on the skin mucus (the external layer covering the entire fish surface), which confers mechanical and disease protection and participates in many other important functions, such as respiration, communication, microbiota environment, and particle trapping [5]. The skin mucus is mainly secreted by goblet cells present in the epidermis, and it forms a continuous layer of adherent gel that covers the apical portions of epithelial cells [6]. Given that most pathogen attacks begin in mucosal surfaces, gaining a thorough understanding of this barrier is of great interest [7]. The skin mucus of gilthead seabream has been studied to evaluate and compare its humoral defense mechanisms with those of the serum [8] or to compare it with the skin mucus of other fish species [9,10]. Furthermore, skin mucus has been used to evaluate fish stress [11,12] or to determine its proteomic profile [5,13]. More recently, stable isotope analysis has made it possible to study skin mucus exudation and renewal [14,15].
In aquaculture production, the importance of having a ‘normal and continuous’ skin and skin mucus is well-established, as aquatic animals are in a tight relationship with their environment where microorganisms, such as parasites, bacteria, viruses, and fungi, may be present at harmful concentrations [16]. Our previous studies revealed important differences between the skin located above and below the lateral line, indicating the structural and physiological differences of the various parts of gilthead seabream [1]. However, no significant differences in terms of the cell cycle of isolated cells or in gene expression were observed in the skin obtained from these two locations. However, the skin below the lateral line has a thicker epidermis, smaller cells, and a smaller area of microridges in the apical part of the epidermal cells compared with the skin obtained above the lateral line. Regarding functional characterization, the rate of wound healing was higher in the skin obtained below the lateral line than in its counterpart [1]. More recently, we performed a radiological characterization of gilthead seabream skin using X-ray computed tomography. The density values of the skin (ranging from −400 to −50 Hounsfield units) were used as references for the in situ analysis of the entire gilthead seabream specimen, and the skin and subcutaneous fat segmentation accounted for 31% of the total fish volume [17]. Furthermore, different dietary supplements (e.g., 1% arginine, 10 mg kg−1 of heat-killed (30 min, 60 °C) Saccharomyces cerevisiae, or 109 colony factor units Shewanella putrefaciens per gram of diet) improved the healing process in the skin of gilthead seabream [18,19,20,21].
On the basis of the above findings, this study mainly aimed to describe the response of gilthead seabream to experimental wounds by evaluating the antioxidant parameters in skin mucus and the expression of genes related to stress and skin regeneration in the skin samples obtained from both flanks of the wounded and non-wounded fish. A thorough understanding of the role of skin mucus in the regeneration process of a wound, as well as the behavior of the immune-related proteins present in the skin, will help in gaining a deeper understanding of the skin healing process in this important marine fish species.
2. Results
2.1. Antioxidant Enzyme Activity in Skin Mucus
The catalase (CAT), superoxide dismutase (SOD), and glutathione reductase (GR) activities were measured in the skin mucus of both flanks of the non-wounded and wounded gilthead seabream (Figure 1).
In the non-wounded fish, no significant differences (p > 0.05) in terms of the studied enzymes were detected between the flanks at 3 and 7 dpw. However, significant increments (p < 0.05) in CAT and GR (at 3 dpw) and in SOD (at 7 dpw) were observed in the skin mucus in both flanks of the wounded fish (the activities were always higher in the wounded flank than in the non-wounded flank). By contrast, the CAT activities in the left flank of the wounded fish at 7 dpw significantly decreased with respect to those in the non-wounded fish. Furthermore, the GR activity was significantly increased (p < 0.05) in the left flank of wounded fish at 7 dpw. However, the GR activity in the right flank of the wounded fish at 7 dpw was increased, albeit not significantly, with respect to those in the non-wounded fish, and the increments were significantly lower than those in the left flank of the same fish (Figure 1).
2.2. Gene Expression in the Skin
The expression levels of important genes that encode stress proteins (glucose-regulated protein 170 (grp170), glucose-regulated protein 94 (grp94), glucose-regulated protein 75 (grp75), superoxide dismutase (sod), and 70 kilodalton heat shock protein (hsp70)) and proteins involved in skin regeneration (transferrin (tf), insulin-like growth factor-1 (igf1), transforming growth factor beta 1 (tgfb1), derlin 1 (der1), apoptosis antigen 1 (apo1), and stress-inducible endoplasmic reticulum DnaJ homologue (erdj3)) were determined in the skin of both flanks of the wounded and non-wounded seabream specimens. Generally, the gene expression levels were highly affected by the presence of a wound (Figure 2).
No significant variations in the expression levels of the studied genes (grp170, grp94, grp75, and sod) were detected between the left and right flanks or between the non-wounded and wounded fish at 3 dpw (Figure 2A). However, the expression levels of hsp70 in both flanks of the wounded fish significantly decreased with respect to those in the skin of the non-wounded fish at 3 dpw (Figure 2A).
While no significant variations were detected in the expression levels of the studied genes in the non-wounded fish at 7 dpw, notable variations were observed in the wounded fish (Figure 2B). Specifically, the expression levels of all the genes in the skin of the wounded fish were significantly increased in the left flank relative to those in the right flank. Furthermore, significant differences in the expression levels of all the studied genes were detected between the wounded and non-wounded fish (Figure 2B).
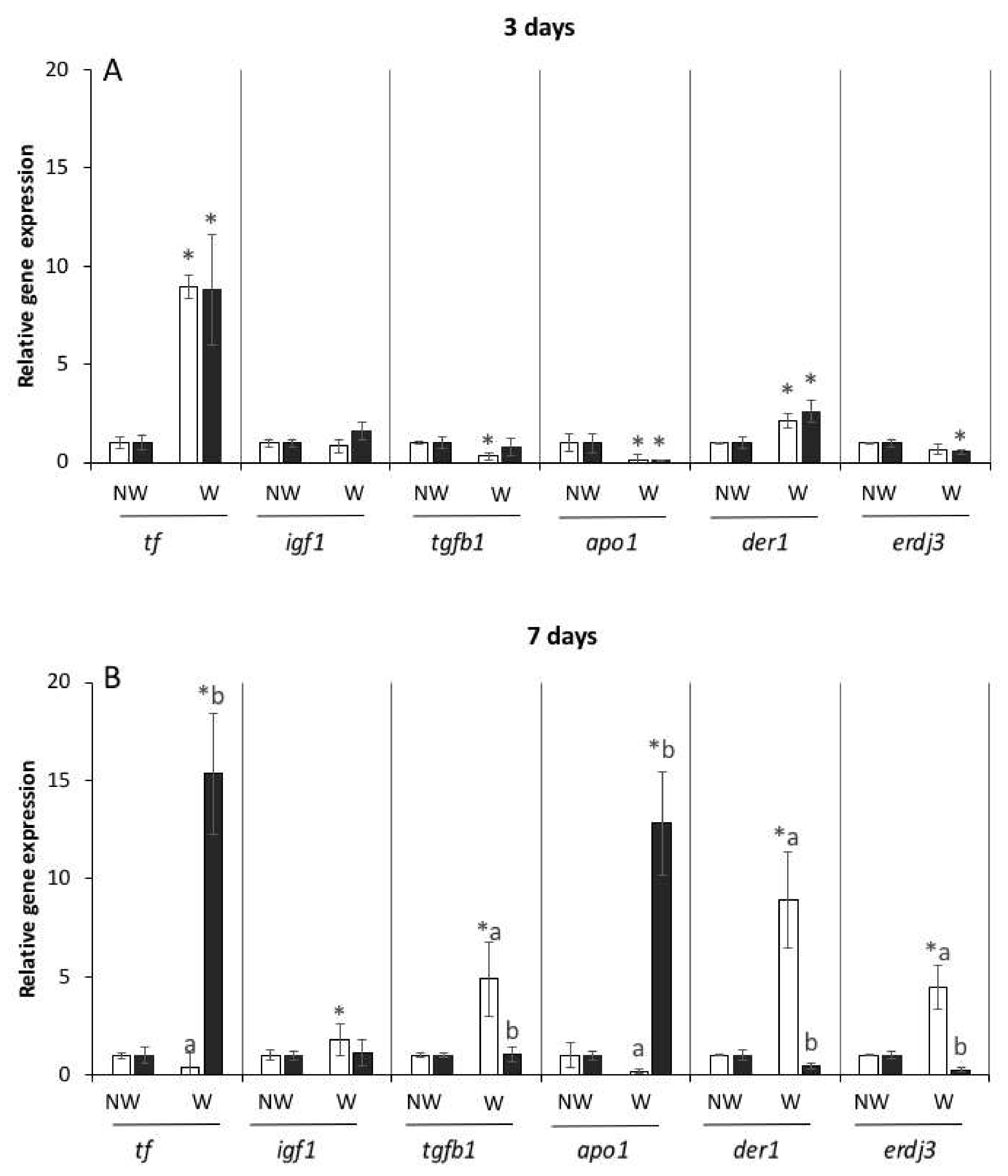
The expression levels of the genes that encode skin regeneration-related proteins (tf, igf1, tgfb1, der1, apo1, and erdj3) were similar in the skin of both flanks in the non-wounded fish. By contrast, notable differences were detected in the gene expression levels in the wounded fish, especially at 7 dpw. While the expression levels of tf and der1 were significantly increased (p < 0.05), those of apo1 were significantly decreased in both flanks of the wounded fish at 3 dpw (Figure 3A). However, the expression levels of tgfb1 in the left flank and of erdj3 in the right flank were significantly decreased with respect to those in the opposite flanks.
Additionally, in the wounded fish, the expression levels of tgfb1, der1, and erdj3 in the skin at 7 dpw were significantly decreased (p < 0.05) in the left flank, whereas those of tf and apo1 were significantly increased in the right flank with respect to those in the opposite flanks (Figure 3B). Furthermore, the expression levels of igf1, tgfb1, der1, and erdj3 in the skin of the wounded fish were significantly increased in the left flank with respect to those in the same flank of the non-wounded fish. By contrast, the expression levels of tf and apo1 in the skin of the right flank of the wounded fish were significantly increased (p < 0.05) (Figure 3B).
3. Discussion
The immune system plays a central role in maintaining physiological homeostasis by sensing changes in the normal conditions and responding to injuries, infections, and stress [20]. In all cases, depending on the induced changes to normality and their magnitude, the extent of the immune response required to eliminate dangers and restore normalcy potentially pose exhausting effects on the limited resources of an organism [20]. In this study, half of the fish were experimentally wounded in their left flank. Following the injury, a proper balance between an effective immune response and tolerance must be achieved, followed by the repair of the damage at the molecular, cellular, and tissue levels. As previously reported, in gilthead seabream, the rate of wound healing below the lateral line was higher than that above the lateral line. Furthermore, the wounds altered the mucus composition, as demonstrated by the changes in several immune parameters relative to those in the non-wounded fish. Significant variations were detected depending on the site of the wound and on the studied parameter [21]. These previous results led us to conceptualize this study, in which we focused on the possible changes caused by the experimental open wounds in terms of the activities of three important enzymes involved in redox status and present in the skin mucus, as well as on the changes in the expression levels of important genes related to stress and skin regeneration. In this study, we aimed to determine whether a wound (which was created in the same spot in the left flank of the studied fish) could cause differences between the left and right flanks of the same fish in terms of antioxidant enzyme activity in the skin mucus and in terms of gene expression in the skin.
The present results indicate that in the non-wounded fish, the antioxidant enzymes were homogeneously distributed in the skin mucus; however, after wounding, significant differences were observed in the mucus of the wounded and non-wounded flanks. In fact, notable changes in mucus composition were detected only in one flank of the fish (one flank was wounded and the other was non-wounded). The mucosal immune response plays a crucial role in the quality of tissue response to an injury [22]. The physiological redox status is determined by oxygen concentration, reactive oxygen species (ROS) levels, and antioxidant concentration (e.g., vitamins C and E or glutathione). ROS and nitrogen species production is an essential innate immune response with important functions mainly in the elimination of previously phagocytized pathogens. Although excessive ROS production is a defense mechanism, it can cause damage to an organism. Thus, the studied enzymes which convert ROS into less-harmful oxygen species are considered to be crucial in redox status maintenance. The protease, anti-protease, and peroxidase activities in the skin mucus of gilthead seabream at 7 dpw did not significantly differ to those obtained in the non-wounded animals [17]. However, the changes observed in the studied enzymes are important because decreased antioxidant defenses could severely compromise the inflamed mucosa, rendering it more susceptible to oxidative tissue damage [23]. The present results agree with a previous finding [21], in which increments in the activity of other related enzymes, such as peroxidase or protease, were observed in the skin mucus of gilthead seabream at 2–3 dpw. Similarly, the CAT expression in the muscle of zebrafish increased at 24 h post wounding, whereas it decreased at 5 dpw [24]. The rise in antioxidant enzymes could be produced by the intake of some antioxidants and is considered a sign of animal health [22]. In the present study, the increments in the antioxidant enzymes in the skin mucus of the wounded fish may have been produced to counteract the oxidative stress related to the inflammatory process developed during the course of skin healing. Furthermore, no significant differences in the CAT and GR activities were found between the left and right flanks, which seems to suggest that they were increased in the skin mucus as a general or systemic response more than a local one. This general response could be mediated by stress factors such as cortisol, growth factors, or Nrf2 [25]. At 7 dpw, the CAT activity in the wounded flank was reduced, whereas the GR activity was significantly increased (p < 0.05). These observations indicated that the normal antioxidant activity in the skin mucus of gilthead seabream was not totally recovered at 7 dpw. Moreover, significant differences in the mucus were detected between the left and right flanks. The increased GR activity in the wounded flank may have been caused by the inflammatory process triggered by the wound, whereas the decrements in the CAT activity suggested a notable oxidative process. The increments in the SOD activity could also be explained as a systemic response, as was already indicated for the CAT and GR activities [25], although with 4 days of delay. In permit (Trachinotus falcatus), it was observed that the antioxidant enzymes CAT and SOD seemed to be differently regulated in the mucus layer, with the latter remaining stable and the former being modulated by external stimuli [26]. Finally, as has been recently demonstrated by the evaluations of mucus exudation dynamics, the turnover of soluble mucus components was increased [27]. In other words, it can be interpreted that the studied enzymes, as part of the soluble fraction of the skin mucus of gilthead seabream, are continuously produced and become incorporated into the skin mucus. The notable differences detected in the activities of the two enzymes at 3 and 7 dpw seem to indicate their active participation in skin inflammation and regeneration induced by the experimental wounds.
The expression levels of several genes involved in the stress response or in tissue regeneration were also studied in the skin of the wounded and non-wounded gilthead seabream specimens. In general, the expression levels of most genes were highly affected by the presence of wound. Expectedly, no significant variations in the expression levels of any of the studied genes were detected in the skin of both flanks of the non-wounded fish. These results agree with a previous finding in which no significant differences in the expression levels of several immune-related genes were detected in the skin above and below the lateral line [1]. These results seem to support the assumption that the gene expression levels in the skin are similar in all parts of a gilthead seabream specimen in the absence of injuries or insults to the skin. Meanwhile, no significant variations were detected in the gene expression levels of grp170, grp94, grp75, and sod at 3 dpw. However, the expression levels of hsp70 in the skin of both flanks of the wounded fish were significantly reduced with respect to those in the non-wounded fish at 3 dpw, which could indicate that this protein is helping to protect cells from the adverse effects caused by the wound. Furthermore, the expression levels of grp170, grp94, grp75, sod, hsp70, igf1, tgfb1, der1, and erdj3 at 7 dpw were upregulated in the left flank (of wounded fish) with respect to those in the control group (non-wounded). Furthermore, the expression of such genes was also upregulated in the skin of the non-wounded flank of wounded fish (p < 0.05). In brief, the induction of GRP proteins (including GRP170, GRP94, and GRP75) is caused by glucose starvation and other stressors, such as hypoxia, ischemia, calcium homeostasis disorder, proteasome inhibitors, and non-steroidal anti-inflammatory drugs [28,29,30,31,32,33,34,35,36,37]. Concomitantly, most of these genes encode proteins that are secreted under stress, such as SOD, hsp70, or derline1 [38,39,40], or during the wound healing process, such as insulin-like growth factor (IGF) [41], the TGF-β superfamily (also involved in different processes and in wound healing) [42], and ERdj3 (a chaperone involved in the coordination of endoplasmic reticulum (ER) and extracellular proteostasis) [43]. The upregulation of some of these genes in the skin of the non-wounded flank of the wounded fish could be explained as a response to different mediators involved in a systemic response, as presented in the discussion on redox enzyme activities. This response could be triggered via different pathways (e.g., nrf2, cortisol, and growth factors) [1,14,23,44]. The expression levels of these genes in the wounded flank did not differ from those in the non-wounded fish (control group). In explaining such results, it could be hypothesized that local factors (e.g., interleukins, inflammation factors, and oxidative stress) could also mediate or interfere with the expression levels of these genes, although future studies should be conducted to assess the validity of this hypothesis and the possible immune and metabolic routes involved. It could also be interpreted that the observed decreases at 7 dpw were a result of a boomerang effect.
Of note, transferrin has been described to be mobilized in flesh wounds [45], although future studies should be conducted to elucidate the implications not only of this protein but also of iron in the skin regeneration process. Meanwhile, heat shock proteins (HSPs) are important factors in protein homeostasis, and they possess chaperone properties. Proteins in the Hsp70 family are the most universal molecular chaperones, and they interact with the greatest number of protein substrates. Hsp70 levels are increased under stress [46,47], and some proteins in the Hsp70 family can be released into the extracellular space, where they play an important role in intercellular communications and act as alarmins or ‘danger signals’ that modulate the immune response [48]. In the present study, the high hsp70 expression levels in the skin of the wounded flank at 7 dpw could also suggest a release of these proteins into the skin mucus. The correlation between the HSPs in the skin mucus and the modulation of this mucosal immunity in fish should be extensively investigated. Furthermore, APO1 is central to the control of lymphoid cell growth [49]; in this work, APO1 is considerably important in the inflammation process induced by wounds, whereas derlin 1 is an ER membrane protein that has been correlated with ER stress, wherein it promotes apoptosis under prolonged ER stress [38]. In general, the expression levels of the markers tf, hsp70, apo1, and der1 varied in the left and right flanks, suggesting that their expression levels were affected by a systemic response, as previously described. As for erdj3, its expression was significantly decreased in the wounded (right) flank relative to that in the control group and to that in the left side of the same fish (p < 0.05). ERdj3 is also a chaperone, but it plays a role in diverse functions related to the coordination of ER and extracellular proteostasis [43]. Under ER stress, ERdj3 is secreted into the extracellular space, working as an ATP-independent extracellular chaperone that prevents the misfolding and/or aggregation of secreted proteins [50]. Our results suggest that the increase in erdj3 expression was mediated by a local response induced by the wounding process.
4. Materials and Methods
4.1. Animals
Forty protandrous hermaphroditic seawater teleost gilthead seabream (mean weight: 42 ± 8 g) were randomly divided into eight running seawater aquaria (five animals per aquaria; 250 L, 900 L h−1 flow rate, 28‰ salinity, 25 °C, and 12 h light:12 h dark artificial photoperiod) at the Marine Fish Facilities of the University of Murcia. The animals were fed with a commercial pellet diet (Skretting, Madrid, Spain) at a rate of 2% body weight per day. The fish were allowed to acclimate for 15 days prior to the experiments. All experimental protocols were approved by the Ethical Committee of the University of Murcia.
4.2. Experimental Design and Sampling
The animals were fed with a commercial diet for a month. Afterwards, the fish from four aquaria were sedated with benzocaine (4% in acetone) (Sigma-Aldrich, Madrid, Spain) and then wounded [21] (Figure 4). All wounds, which had a diameter of 7 mm and a depth of 2 mm and were located at the middle of the left flank below the lateral line, were made by the same person using a metallic circular biopsy punch (Stiekel). Once the fish recovered from the anesthesia, they were placed back into their respective tanks. At three and seven dpw, ten animals from each trial group (n = 5, two replicates) were anesthetized, weighed and sampled. Samples of skin mucus and skin were obtained from each specimen.
Skin mucus samples were collected by gently scraping the surface of the seabream specimens using a plastic cell scraper while avoiding contamination of the sample with blood and urinogenital or intestinal excretions [8]. The novelty of this work is that two samples of skin mucus were obtained from each fish (one sample was obtained from the left or wounded flank and another from the right or non-wounded flank). Then, the mucus was centrifuged (12,000× g, 4 °C, 10 min), and the supernatant was immediately stored at −80 °C until further analysis. Skin fragments from the wounded and non-wounded flanks were also obtained for different determinations, as described below.
4.3. Antioxidant Enzyme Determination in Skin Mucus
The protein concentrations in the skin samples were determined by a routine method (using Bradford reagent), and all samples were adjusted to 500 µg protein mL−1 (diluted in 50 mM potassium phosphate buffer; pH 7.0). The mucus samples were used to determine the glutathione reductase (GR), superoxide dismutase (SOD), and catalase (CAT) activities, and all assays were performed in triplicate. GR was measured as previously described [51]. The reaction was initiated by adding 100 µL of 0.1 mM NADPH to a cuvette containing 50 µL of skin mucus and 600 µL of 50 mM potassium phosphate buffer (pH 7.0) with 2 mM EDTA and 200 µL of 0.5 mM GSSG (Sigma-Aldrich, Madrid, Spain). The change in absorbance was monitored at 340 nm for 3 min by a UV–Vis Thermo Scientific Model Evolution 300 dual beam spectrophotometer (Thermo Fisher Scientific; Berlin, Germany). One unit of GR activity was defined as the amount of enzyme that catalyzed the reduction of 1 µmol of NADPH per minute (ε340 nm for NADPH 6.22 mM−1 cm−1). GR activity was expressed in units per milligram of protein.
The SOD activity determination was based on the inhibition of the reduction of cytochrome C in the presence of SOD at 550 nm [52], with a slight modification in the methodology [51]. The SOD-like activity of the mucus samples was studied at 25 °C by an indirect method using cytochrome C as superoxide oxidant (which acted as an indicator). A mucus sample (50 µL) was added to 950 µL of 50 mM potassium phosphate buffer (pH 7.0) containing EDTA (2 mM), xanthine (100 µM), and cytochrome C (12 µM) (Sigma-Aldrich, Madrid, Spain). The formation of the reduced form of cytochrome C was detected spectrophotometrically at 550 nm using a Thermo Scientific Evolution 300 dual beam spectrophotometer. The SOD activity was expressed in units per milligram of protein.
CAT activity was measured by monitoring the consumption of hydrogen peroxide (H2O2) at 240 nm [51]. A mucus sample (50 µL) was pipetted into a cuvette containing 950 µL of H2O2 solution (10 mM) in a 50 mM phosphate buffer (pH 7). The reduction of H2O2 was monitored at 240 nm for 4 min against a blank containing 50 mM phosphate buffer. This method was based on the principle that absorbance decreases due to the decomposition of H2O2 by catalase, and the activity was expressed in U × mg protein–1.
4.4. Quantitative Real-Time PCR
Relative gene expression was analyzed in the skin samples obtained from five fish from each experimental group (3 and 7 dpw) using real-time PCR and the 2−ΔΔCT method [53]. Total RNA was extracted from seabream skin and liver fragments using TRIzol reagent (Invitrogen) following the manufacturer’s instructions. The RNA was subsequently treated with DNase I (Invitrogen) to remove genomic DNA contamination. Complementary DNA (cDNA) was synthesized from 1 mg of total RNA using SuperScript IV reverse transcriptase (Invitrogen) with an oligo-dT18 primer. The expression levels of 14 genes were analyzed by real-time PCR performed with an ABI PRISM 7500 instrument (Applied Biosystems) using SYBR Green PCR Core Reagents (Applied Biosystems), as previously described [54].
These genes encode stress proteins (grp170, grp94, grp75, sod, and hsp70) and skin regeneration-related proteins (tf, igf1, tgfb1, der1, apo1, and erdj3). For each mRNA, gene expression was corrected by the median of the elongation factor 1-alpha (ef1a), β-actin, and 18-S expression content in each sample. The primers used are shown in Table 1. Sequences were obtained from [55,56,57]. In all cases, each PCR was performed in triplicate.
4.5. Statistical Analyses
The results are expressed as mean ± SEM. The normality of the variables was confirmed by the Shapiro–Wilk test, and homogeneity of variance was confirmed by Levene’s test. Statistical differences among the experimental groups were assessed with one-way ANOVA, followed by the Tukey or Games Howell test, depending on the homogeneity of the variables. The significance level was 95% in all cases (p < 0.05). Analyses were performed using SPSS for Windows (version 15.0, SPSS Inc., Chicago, IL, USA).
5. Conclusions
Our study demonstrated that a wound in a specific spot in fish skin could induce a global response that affected the entire skin surface. In particular, the antioxidant defense in the skin mucus could locally respond to wound stress, with the antioxidant response being altered even at 7 dpw. Finally, the expression levels of several important genes related to skin stress and regeneration were similar in the earlier phases of skin regeneration, but their pattern of expression in the right and left flanks varied at 7 dpw. The analyzed markers remained upregulated in the skin of the non-wounded flank, whereas most of them were expressed at constitutive levels or even downregulated in the wounded flank. In this sense, we conclude that a single superficial wound can trigger important changes in the skin mucus composition as well as in the skin, as demonstrated in the changes in gene expression levels, which are important in cases where fishes experience stress or become infected with a pathogen.
Author Contributions
Conceptualization, M.Á.E.; methodology, C.E.-R.; validation, C.E.-R. and M.Á.E.; investigation, C.E.-R.; resources, M.Á.E.; data curation, C.E.-R.; writing—original draft preparation, C.E.-R.; writing—review and editing, M.Á.E.; project administration, M.Á.E.; funding acquisition, M.Á.E. Both authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Ministry of Economy and Competitiveness (MINECO), and co-funded with the European Regional Development Funds (ERDF/FEDER) (Grant no. AGL-2017-83370-C3-1-R) and Fundación Séneca de la Región de Murcia (Grupo de Excelencia 19883/GERM/15).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of the University of Murcia (protocol code A13150104 and date of approval of 10th November 2017).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cordero, H.; Ceballos-Francisco, D.; Cuesta, A.; Esteban, M.Á. Dorso-ventral skin characterization of the farmed fish gilthead seabream (Sparus aurata). PLoS ONE 2017, 12, e0180438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, F.A.; Gregório, S.F.; Ferraresso, S.; Thorne, M.A.; Costa, R.; Milan, M.; Bargelloni, L.; Clark, M.S.; Canario, A.V.; Power, D.M. Skin healing and scale regeneration in fed and unfed sea bream, Sparus auratus. BMC Genom. 2011, 12, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iger, Y.; Abraham, M. The process of skin healing in experimentally wounded carp. J. Fish Biol. 1990, 36, 421–437. [Google Scholar] [CrossRef]
- Quilhac, A.; Sire, J.-Y.Y. Spreading, proliferation, and differentiation of the epidermis after wounding a cichlid fish, Hemichromis bimaculatus. Anat. Rec. 1999, 254, 435–451. [Google Scholar] [CrossRef]
- Jurado, J.; Fuentes-Almagro, C.A.; Guardiola, F.A.; Cuesta, A.; Esteban, M.Á.; Prieto-Álamo, M.J. Proteomic profile of the skin mucus of farmed gilthead seabream (Sparus aurata). J. Proteom. 2015, 120, 21–34. [Google Scholar] [CrossRef]
- Esteban, M.Á.; Cerezuela, R. Fish mucosal immunity: Skin. In Mucosal Health in Aquaculture; Academic Press: Cambridge, MA, USA, 2015; pp. 67–92. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A.; Meseguer, J.; Esteban, M.Á. Increases in immune parameters by inulin and Bacillus subtilis dietary administration to gilthead seabream (Sparus aurata L.) did not correlate with disease resistance to Photobacterium damselae. Fish Shellfish Immunol. 2012, 32, 1032–1040. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Cuesta, A.; Arizcun, M.; Meseguer, J.; Esteban, M.A. Comparative skin mucus and serum humoral defence mechanisms in the teleost gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2014, 36, 545–551. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Cuesta, A.; Abellán, E.; Meseguer, J.; Esteban, M.A. Comparative analysis of the humoral immunity of skin mucus from several marine teleost fish. Fish Shellfish Immunol. 2014, 40, 24–31. [Google Scholar] [CrossRef]
- Sanahuja, I.; Fernández-Alacid, L.; Sánchez-Nuño, S.; Ordóñez-Grande, B.; Ibarz, A. Chronic cold stress alters the skin mucus interactome in a temperate fish model. Front. Physiol. 2019, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Guardiola, F.A.; Cuesta, A.; Esteban, M.Á. Using skin mucus to evaluate stress in gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2016, 59, 323–330. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Logothetis, P.; Meseguer, J.; Esteban, M.A. Evaluation of silver nanospheres on viability and innate cellular parameters of gilthead seabream (Sparus aurata L.) head-kidney leucocytes. Fish Shellfish Immunol. 2017, 69, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, M.C.; Calduch-Giner, J.A.; Fouz, B.; Estensoro, I.; Simó-Mirabet, P.; Puyalto, M.; Karalazos, V.; Palenzuela, O.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Under control: How a dietary additive can restore the gut microbiome and proteomic profile, and improve disease resilience in a marine teleostean fish fed vegetable diets. Microbiome 2017, 5, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Ibarz, A.; Pinto, P.I.S.; Power, D.M. Proteomic Approach to Skin Regeneration in a Marine Teleost: Modulation by Oestradiol-17β. Mar. Biotechnol. 2013, 15, 629–646. [Google Scholar] [CrossRef]
- Ordóñez-Grande, B.; Fernández-Alacid, L.; Sanahuja, I.; Sánchez-Nuño, S.; Fernández-Borràs, J.; Blasco, J.; Ibarz, A. Evaluating mucus exudation dynamics through isotopic enrichment and turnover of skin mucus fractions in a marine fish model. Conserv. Physiol. 2020, 8, 629–646. [Google Scholar] [CrossRef]
- Lekang, O.-I. Aquaculture Engineering, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; Available online: https://www.wiley.com/en-us/Aquaculture+Engineering%2C+2nd+Edition-p-9781118496091 (accessed on 18 March 2021).
- Ceballos-Francisco, D.; Carrillo, N.G.; Pardo-Fernández, F.J.; Cuesta, A.; Esteban, M.Á. Radiological characterization of gilthead seabream (Sparus aurata) by X-ray computed tomography. J. Fish Biol. 2020, 97, 1440–1447. [Google Scholar] [CrossRef]
- Chen, Z.; Ceballos-Francisco, D.; Guardiola, F.A.; Huang, D.; Esteban, M.Á. The alleviation of skin wound-induced intestinal barrier dysfunction via modulation of TLR signalling using arginine in gilthead seabream (Sparus aurata L). Fish Shellfish Immunol. 2020, 107, 519–528. [Google Scholar] [CrossRef]
- Chen, Z.; Ceballos-Francisco, D.; Guardiola, F.A.; Esteban, M.Á. Dietary administration of the probiotic Shewanella putrefaciens to experimentally wounded gilthead seabream (Sparus aurata L.) facilitates the skin wound healing. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, C.; Esteban, M.Á. Effect of dietary supplementation with yeast Saccharomyces cerevisiae on skin, serum and liver of gilthead seabream (Sparus aurata L). J. Fish Biol. 2020, 97, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ceballos-Francisco, D.; Guardiola, F.A.; Esteban, M.Á. Influence of skin wounds on the intestinal inflammatory response and barrier function: Protective role of dietary Shewanella putrefaciens SpPdp11 administration to gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2020, 99, 414–423. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M.; Forman, H.J. Redox homeostasis: The Golden Mean of healthy living. Redox Biol. 2016, 8, 205–215. [Google Scholar] [CrossRef]
- Ceballos-Francisco, D.; Cordero, H.; Guardiola, F.A.; Cuesta, A.; Esteban, M.Á. Healing and mucosal immunity in the skin of experimentally wounded gilthead seabream (Sparus aurata L). Fish Shellfish Immunol. 2017, 71, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.Á. An Overview of the Immunological Defenses in Fish Skin. ISRN Immunol. 2012, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Buffinton, G.D.; Doe, W.F. Depleted mucosal antioxidant defences in inflammatory bowel disease. Free Radic. Biol. Med. 1995, 19, 911–918. [Google Scholar] [CrossRef]
- Seo, S.B.; Dananjaya, S.H.S.; Nikapitiya, C.; Park, B.K.; Gooneratne, R.; Kim, T.-Y.; Lee, J.; Kim, M.C.-H.; Zoysa, D. Silver nanoparticles enhance wound healing in zebrafish (Danio rerio). Fish Shellfish Immunol. 2017, 68, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Nogales, A.; Nederlof, M.; Benedito-Palos, L.; Ballester-Lozano, G.F.; Folkedal, O.; Olsen, R.E.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Metabolic and transcriptional responses of gilthead sea bream (Sparus aurata L.) to environmental stress: New insights in fish mitochondrial phenotyping. Gen. Comp. Endocrinol. 2014, 205, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazado, C.C.; Lund, I.; Pedersen, P.B.; Nguyen, H.Q. Humoral and mucosal defense molecules rhythmically oscillate during a light-dark cycle in permit, Trachinotus falcatus. Fish Shellfish Immunol. 2015, 47, 902–912. [Google Scholar] [CrossRef]
- Sciandra, J.J.; Subjeck, J.R. The effects of glucose on protein synthesis and thermosensitivity in Chinese hamster ovary cells. J. Biol. Chem. 1983, 258, 12091–12093. Available online: http://www.ncbi.nlm.nih.gov/pubmed/6630181 (accessed on 13 May 2019). [CrossRef]
- Gao, Y.Y.; Liu, B.Q.; Du, Z.X.; Zhang, H.Y.; Niu, X.F.; Wang, H.Q. Implication of Oxygen-Regulated Protein 150 (ORP150) in Apoptosis Induced by Proteasome Inhibitors in Human Thyroid Cancer Cells. J. Clin. Endocrinol. Metab. 2010, 95, E319–E326. [Google Scholar] [CrossRef] [Green Version]
- Namba, T.; Hoshino, T.; Tanaka, K.-I.; Tsutsumi, S.; Ishihara, T.; Mima, S.; Suzuki, K.; Ogawa, S.; Mizushima, T. Up-Regulation of 150-kDa Oxygen-Regulated Protein by Celecoxib in Human Gastric Carcinoma Cells. Mol. Pharmacol. 2006, 71, 860–870. [Google Scholar] [CrossRef]
- Sciandra, J.J.; Subjeck, J.R.; Hughes, C.S. Induction of glucose-regulated proteins during anaerobic exposure and of heat-shock proteins after reoxygenation. Proc. Natl. Acad. Sci. USA 1984, 81, 4843–4847. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.-W.; Henderson, B.W.; Shen, J.-W.; Subjeck, J.R. Induction of glucose regulated proteins during growth of a murine tumor. J. Cell. Physiol. 1993, 154, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Whelan, S.A.; Hightower, L.E. Differential induction of glucose-regulated and heat shock proteins: Effects of pH and sulfhydryl-reducing agents on chicken embryo cells. J. Cell. Physiol. 1985, 125, 251–258. [Google Scholar] [CrossRef]
- Wang, H.; Pezeshki, A.M.; Yu, X.; Guo, C.; Subjeck, J.R.; Wang, X.-Y. The Endoplasmic Reticulum Chaperone GRP170: From Immunobiology to Cancer Therapeutics. Front. Oncol. 2015, 4, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzec, M.; Eletto, D.; Argon, Y. GRP94: An HSP90-like protein specialized for protein folding and quality control in the endoplasmic reticulum. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 774–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Niu, X.; Zhang, Q.; Chen, H.; Gao, A.; Qi, S.; Xiang, R.; Belting, M.; Zhang, S. Mitochondria chaperone GRP75 moonlighting as a cell cycle controller to derail endocytosis provides an opportunity for nanomicrosphere intracellular delivery. Oncotarget 2017, 8, 58536–58552. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.J.; Chang, Y.C.; Chang, H.C.; Wang, C.K.; Hung, Y.C.; Lin, Y.E.; Chan, C.C.; Chen, H.Y.; Chang, C.H.; Sang, T.K. Derlin-1 Regulates Mutant VCP-Linked Pathogenesis and Endoplasmic Reticulum Stress-Induced Apoptosis. PLoS Genet. 2014, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kalmar, B.; Greensmith, L. Activation of the heat shock response in a primary cellular model of motoneuron neurodegeneration-evidence for neuroprotective and neurotoxic effects. Cell. Mol. Biol. Lett. 2009, 14, 319–335. [Google Scholar] [CrossRef]
- Espinosa, C.; López-Jiménez, J.A.; Pérez-Llamas, F.; Guardiola, F.A.; Esteban, M.A.; Arnao, M.B.; Zamora, S. Long-term intake of white tea prevents oxidative damage caused by adriamycin in kidney of rats. J. Sci. Food Agric. 2015, 96, 3079–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chablais, F.; Jazwinska, A.; Jones, B.; Petkovich, M.; Tada, M.; Ekker, M.; Akimenko, M.A. IGF signaling between blastema and wound epidermis is required for fin regeneration. Development 2010, 137, 871–879. [Google Scholar] [CrossRef] [Green Version]
- McCartney-Francis, N.L.; Frazier-Jessen, M.; Wahl, S.M. TGF-β: A Balancing Act. Int. Rev. Immunol. 1998, 16, 553–580. [Google Scholar] [CrossRef]
- Chen, K.; Qu, S.; Chowdhury, S.; Noxon, I.C.; Schonhoft, J.D.; Plate, L.; Powers, E.T.; Kelly, J.W.; Lander, G.C.; Wiseman, R.L. The endoplasmic reticulum HSP40 co-chaperone ERdj3/DNAJB11 assembles and functions as a tetramer. EMBO J. 2017, 36, 2296–2309. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Chowdhry, S.; Dinkova-Kostova, A.T.; Sutherland, C. Dual regulation of transcription factor Nrf2 by Keap1 and by the combined actions of β-TrCP and GSK-3. Biochem. Soc. Trans. 2015, 43, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helson, L.; Rosenspire, K.; Kapellaris, A.; Bigler, R.; Richards, P.; Srivastava, S.; Meinken, G.; Chaglassian, T. Uptake of ruthenium-labeled transferrin in healing wounds. Int. J. Nucl. Med. Biol. 1983, 10, 237–239. [Google Scholar] [CrossRef]
- He, J.; Wang, J.; Xu, M.; Wu, C.; Liu, H. The cooperative expression of Heat Shock Protein 70 KD and 90 KD gene in juvenile Larimichthys crocea under Vibrio alginolyticus stress. Fish Shellfish Immunol. 2016, 58, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Hangzo, H.; Banerjee, B.; Saha, S.; Saha, N. Ammonia stress under high environmental ammonia induces Hsp70 and Hsp90 in the mud eel, Monopterus cuchia. Fish Physiol. Biochem. 2016, 95, 203–212. [Google Scholar] [CrossRef]
- Garbuz, D.G.; Zatsepina, O.G.; Evgen’ev, M.B. The Major Human Stress Protein Hsp70 as a Factor of Protein Homeostasis and a Cytokine-Like Regulator. Mol. Biol. 2019, 53, 200–217. [Google Scholar] [CrossRef]
- Debatin, K.; Goldman, C.; Waldmann, T.; Krammer, P. APO-1-induced apoptosis of leukemia cells from patients with adult T-cell leukemia. Blood 1993, 81, 2972–2977. Available online: http://www.bloodjournal.org/content/81/11/2972.long?sso-checked=true (accessed on 19 July 2019). [CrossRef] [Green Version]
- Genereux, J.C.; Wiseman, R.L. Regulating extracellular proteostasis capacity through the unfolded protein response. Prion 2015, 9, 10–21. [Google Scholar] [CrossRef]
- Espinosa, C.; Pérez-Llamas, F.; Guardiola, F.A.; Esteban, M.A.; Arnao, M.B.; Zamora, S.; López-Jiménez, J.A. Molecular mechanisms by which white tea prevents oxidative stress. J. Physiol. Biochem. 2014, 70, 891–900. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide dismutase: An enzymic Funcion for Erytrhocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cerezuela, R.; Meseguer, J.; Esteban, M.Á. Effects of dietary inulin, Bacillus subtilis and microalgae on intestinal gene expression in gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2013, 34, 843–848. [Google Scholar] [CrossRef]
- Leung, L.Y.; Kwong, A.K.Y.; Man, A.K.Y.; Woo, N.Y.S. Direct actions of cortisol, thyroxine and growth hormone on IGF-I mRNA expression in sea bream hepatocytes. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2008, 151, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Hernández, M.; Chaves-Pozo, E.; Cabas, I.; Mulero, V.; García-Ayala, A.; García-Alcázar, A. Testosterone implants modify the steroid hormone balance and the gonadal physiology of gilthead seabream (Sparus aurata L.) males. J. Steroid. Biochem. Mol. Biol. 2013, 138, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Genereux, J.C.; Qu, S.; Zhou, M.; Ryno, L.M.; Wang, S.; Shoulders, M.D.; Kaufman, R.J.; Lasmézas, C.I.; Kelly, J.W.; Wiseman, R.L. Unfolded protein response-induced ERdj3 secretion links ER stress to extracellular proteostasis. EMBO J. 2014, 34, 4–19. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Catalase, superoxide dismutase, and glutathione reductase activities (expressed in units per milligram of proteins) in the wounded and non-wounded flanks at 3 and 7 days post wounding in the control (non-wounded) and wounded fish. Values are presented as mean ± SEM (n = 5). Differences between the flanks are indicated by different letters; differences between groups (wounded and non-wounded fish) are indicated by asterisks (p < 0.05).
Figure 1.
Catalase, superoxide dismutase, and glutathione reductase activities (expressed in units per milligram of proteins) in the wounded and non-wounded flanks at 3 and 7 days post wounding in the control (non-wounded) and wounded fish. Values are presented as mean ± SEM (n = 5). Differences between the flanks are indicated by different letters; differences between groups (wounded and non-wounded fish) are indicated by asterisks (p < 0.05).

Figure 2.
Expression levels of stress-related genes in the skin of both flanks in the non-wounded (NW, control) and wounded (W) fish (left side (with wound in wounded fish) or right side (without wound)). Values are presented as mean ± SEM (n = 5). Differences between flanks are indicated by different letters; differences between groups (wounded or non-wounded fish) are indicated by asterisks (p < 0.05).
Figure 2.
Expression levels of stress-related genes in the skin of both flanks in the non-wounded (NW, control) and wounded (W) fish (left side (with wound in wounded fish) or right side (without wound)). Values are presented as mean ± SEM (n = 5). Differences between flanks are indicated by different letters; differences between groups (wounded or non-wounded fish) are indicated by asterisks (p < 0.05).

Figure 3.
Expression levels of the genes related to skin regeneration and stress in the skin obtained 3 days (A) and 7 days (B) post-wounding in the non-wounded (NW, control) and wounded (W) fish. Left side (with wound in wounded fish, white columns); right side (without wound, black columns). Values are presented as mean ± SEM (n = 5). Differences between flanks are indicated by different letters; differences between the same flank (left or right) between NW and W fish are indicated by asterisks (p < 0.05).
Figure 3.
Expression levels of the genes related to skin regeneration and stress in the skin obtained 3 days (A) and 7 days (B) post-wounding in the non-wounded (NW, control) and wounded (W) fish. Left side (with wound in wounded fish, white columns); right side (without wound, black columns). Values are presented as mean ± SEM (n = 5). Differences between flanks are indicated by different letters; differences between the same flank (left or right) between NW and W fish are indicated by asterisks (p < 0.05).

Figure 4.
Representative photographs of the experimental wounds placed below the lateral line on the left flank of gilthead seabream specimens. The photographs were taken at 3 (A,C) and 7 (B,D) days post wounding.
Figure 4.
Representative photographs of the experimental wounds placed below the lateral line on the left flank of gilthead seabream specimens. The photographs were taken at 3 (A,C) and 7 (B,D) days post wounding.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used for qPCR performance.
| Gen | Accession Number | Primer Sequence 5′-3′ | References |
|---|---|---|---|
| ef1α | AF184170 | F: CTTCAACGCTCAGGTCATCAT | [20] |
| R: GCACAGCGAAACGACCAAGGGGA | |||
| 18S | AM490061 | F: CGAAAGCATTTGCCAAGAAT | [20] |
| R: AGTTGGCACCGTTTATGGTC | |||
| b-actin | X89920 | F: TCCTGCGGAATCCATGAGA | [20] |
| R: GACGTCGCACTTCATGATGCT | |||
| grp-170 | JQ3088211 | F: CAGAGGAGGCAGACAGCAAGAC | [20] |
| R: TTCTCAGACTCAGCATTTCCAGATTTC | |||
| grp-94 | JQ3088201 | F: AAGGCACAGGCTTACCAGACAG | [20] |
| R: CTTCAGCATCATCGCCGACTTTC | |||
| grp-75 | DQ524993 | F: TCCGGTGTGGATCTGACCAAAGAC | [20] |
| R: TGTTTAGGCCCAGAAGCATCCATG | |||
| sod | AJ937872 | F: CCATGGTAAGAATCATGGCGG | [20] |
| R: CGTGGATCACCATGGTTCTG | |||
| tf | JF309047 | F: CAGGACCAGCAGACCAAGTT | [20] |
| R: TGGTGGAGTCCTTGAAGAGG | |||
| hsp70 | EU805481 | F: AATGTTCTGCGCATCATCAA | [20] |
| R: GCCTCCACCAAGATCAAAGA | |||
| igf-I | AY608674 | F: TCTCCTGTAGCCACACCCTCTC | [55] |
| R: GAAGCAGCACTCGTCCACAATG | |||
| tgfb1 | AF424703 | F: AGAGACGGGCAGTAAAGAA | [56] |
| R: GCCTGAGGAGACTCTGTTGG | |||
| der | JQ3088251 | F: ACTGCCTCGGTTGCCTTTCC | [20] |
| R: TGGCTGTCACAAGTCTCCAGATATG | |||
| apo1 | F: CTCTTCAGGGTTCCCTTTCC | [20] | |
| R: TTCAGGGCCTCAAGATCAAC | |||
| erdj3 | JQ3088271 | F: AACCGACAGCAGCAGGACAG | [57] |
| R: ACTTCTTCAAGCGTGACCTCCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Espinosa-Ruíz, C.; Esteban, M.Á. Wound-Induced Changes in Antioxidant Enzyme Activities in Skin Mucus and in Gene Expression in the Skin of Gilthead Seabream (Sparus aurata L.). Fishes 2021, 6, 15. https://doi.org/10.3390/fishes6020015
AMA Style
Espinosa-Ruíz C, Esteban MÁ. Wound-Induced Changes in Antioxidant Enzyme Activities in Skin Mucus and in Gene Expression in the Skin of Gilthead Seabream (Sparus aurata L.). Fishes. 2021; 6(2):15. https://doi.org/10.3390/fishes6020015
Chicago/Turabian StyleEspinosa-Ruíz, Cristóbal, and María Ángeles Esteban. 2021. "Wound-Induced Changes in Antioxidant Enzyme Activities in Skin Mucus and in Gene Expression in the Skin of Gilthead Seabream (Sparus aurata L.)" Fishes 6, no. 2: 15. https://doi.org/10.3390/fishes6020015