
A Novel C-Type Lectin and Its Potential Role in Feeding and Feed Selection in Ruditapes philippinarum
by
 and
and
Sentao Chen
1,
Peng Shi
1,
Qingkai Feng
2,
Xiaoting Qiu
2,
Jilin Xu
1,
Xiaojun Yan
1 and
Chengxu Zhou
2,* 1
School of Marine Sciences, Ningbo University, Ningbo 315000, China
2
College of Food and Pharmaceutical Sciences, Ningbo University, Ningbo 315000, China
*
Author to whom correspondence should be addressed.
Fishes 2023, 8(2), 62; https://doi.org/10.3390/fishes8020062
Submission received: 27 November 2022
/
Revised: 10 January 2023
/
Accepted: 13 January 2023
/
Published: 19 January 2023
(This article belongs to the Special Issue Feeding and Nutrition of Bivalves)
Abstract
:In recent years, the role of lectins in the feed selection of bivalve has become hot research topic. Manila clam Ruditapes philippinarum is a species of marine bivalve with important economic value. A new C-type lectin (Rpcl) from the clam was obtained and its potential role in feeding and feed selection was studied. Rpcl cDNA was 929 bp in length and had 720 bp of open reading frame. Rpcl encoded 235 amino acids, comprising a carbohydrate recognition domain (CRD) as well as an N-terminal signal peptide. Rpcl contained a conserved CRD disulfide bond including five cysteine residues (Cys125, Cys142, Cys213, Cys219, and Cys237) and the QPN motif (GLN204-PRO205-ASN206). Phylogenetic analysis revealed that the amino acid sequence of Rpcl was closely related to that of Vpclec-1 from R. philippinarum and C-type lectin from Mercenaria. The qPCR analysis indicated that Rpcl expression was observed in all examined tissues and was the highest in gills followed by in the hepatopancreas, and to a lesser extent in the mantle and lip. The in vitro agglutination experiments showed that, the purified Rpcl protein could selectively agglutinate with different microalgae. The strongest agglutinating effect with Chlorella sp. was observed, followed by Karlodinium veneficum and Chaetoceros debilis Cleve. No agglutination with Prorocentrum minimum was observed. In the feeding experiment, compared with that of the starvation group, Rpcl expression in the lip and gill of the clam fed with C. debilis and K. veneficum showed a significant upward trend with the change of time. In addition, it was found that the changes in the expression of the Rpcl gene in the gill and lip, the main feeding tissues, were consistent with the slope of the decrease in the number of algal cells in the water body. In summary, the structure of a new C-type lectin (Rpcl) was reported in this study and its correlation with the feeding and feed selection in R. philippinarum was confirmed.
1. Introduction
China ranks first in marine shellfish cultivation in the world, with a production of 14.808 million tons in 2021 [1]. Manila clam R. philippinarum is a species of marine bivalve mollusks of significant importance in China’s fishery industry. The huge clam breeding in China, 3 million tons of clams annually, accounts for 90% of the world’s total production [2]. R. philippinarum not only occupies an important economic position, but also an important ecological position among other bivalves. Shellfish act as a filter feeder, shellfish culture has a natural “green” attribute and plays a positive ecological role in purifying water quality and slowing down eutrophication [3].
Lectins are widely distributed in bacteria, viruses, fungi, protozoa, plants, and animals [4]. Carbohydrate recognition domains (CRDs) are present in animal lectins, which contain conserved amino acid motifs and structural domains. According to the different sugar recognition domains, animal lectins are classified into seven groups: C-type lectins (CTLs), S-type lectins (galectins), I-type lectins, P-type lectins, F-type lectins, n-pentameric proteins, and fibroadectin [5]. Galectins and CTLs are two extensively investigated lectins in invertebrates. CTLs are the most commonly explored animal lectins with diverse functions. CTLs are proteins binding to carbohydrates in a Ca2+-dependent way [6]. According to the different structures of CTLs, it is possible to determine their function. Their carbohydrate-binding activity has been found to be mediated by CRDs. CRDs are found in Ca2+-dependent lectins rather than in other lectin categories [7,8]. The CRD of a CTL has a tight globular structure, and this domain is referred to as the C-type lectin domain (CTLD) or C-type CRD [9,10]. As the number of CTLs discovered increases, proteins with C-type CRD do not necessarily require Ca2+ dependence for binding activity [11,12].
Lectins of bivalves have been identified for their feed selection mechanism. For example, a newly predicted CTL was reported to be obtained from the blue mussel Mytilus edulis [13]. The experimental results showed that this lectin is involved in the feeding behavior of M. edulis [13]. It was also demonstrated that both seasonal factors and food quality have effects on gene expression of this lectin and the sorting of particles in blue mussel mucus [13]. Moreover, it was found that a non-calcium-dependent CTL was involved in microalgal feed selection of Sinonovacula constricta [14]. In addition, a reverse genetics study found that CTLs in the mucosa of the oyster Crassostrea virginica, which also belong to bivalve shellfish, directly participate in food particle selection [15]. Cell surface carbohydrates played a crucial part in the mechanisms of cell recognition in a study by Espinosa et al. (2010) [16]. They evidence that selection of a particle by suspended feeding bivalves may be mediated by the interaction of the relevant carbohydrates on the surface of particles with lectins occurring in the mucus that covers the feeding organs of bivalves [16]. However, the lectin type involved in the feeding and feed of R. philippinarum remains unknown. The importance of studying the nutrigenomics of different crustacean species is well known. The use of specific nutrients can be effective in promoting growth, molting, lipid metabolism, antioxidant capacity, and reproduction. In the field of crustacean research, nutrigenomics has been rapidly developed for studying several aspects related to the effects of nutrients on crustacean development. Several researchers have studied the relationship between several functional genes and their expression profiles in crustaceans and several physiological functions. They found a strong link between the optimal feeding effect and efficient production, growth, reproductive development, and health status in several crustacean species. Furthermore, they showed that the regulation of gene expression in individual cells by different nutrients and formulated diets could improve growth and development and immune enhancement in several crustacean species [17]. In this paper, we investigated whether the feeding of representative toxic algae and bait algae to R. philippinarum caused the expression of functional genes in R. philippinarum, and interesting research phenomena emerged.
In this research, a CTL from R. philippinarum (Rpcl) was first characterized. Then the agglutination activity of Rpcl against different classes of microalgae was assayed. Finally, the expression of Rpcl in the feeding organs (lip and gill) was assayed in vitro after feeding. The results highlight an essential role of Rpcl in the feeding and feed selection of R. philippinarum.
2. Materials and Methods
2.1. Clams and Microalgae
The adult R. philippinarum weighed about 4–5 g. The mean body length was 4.0 ± 0.3 cm. They were collected from the mariculture farm in Xiangshan, Ningbo City, Zhejiang Province, China. Eight tissues including adductor muscle, lips, intestine, gill, pipe, mantle, hepatopancreas, and feet of six normal adult R. philippinarum were used to explore gene expression levels. All tissue samples were kept in a refrigerator at −80 °C for RNA extraction. For the feeding experiment, the clams were bought from the market and temporarily cultured in laboratory in seawater with a salinity of 28 ppt and a temperature of 18 °C, and fed with Chlorella vulgaris for 4 days. The microalgae applied in the agglutination assays were Chaetoceros debilis, Karlodinium veneficum, Chlorella pyrenoidosa and Prorocentrum minimum. That applied in feeding experiment were C. debilis, K. veneficum. After the feeding experiment, the lip and gill of R. philippinarum were sampled for further molecular studies.
2.2. Cloning of Rpcl cDNA
The extracted RNA was examined qualitatively by agarose gel electrophoresis and evaluated quantitatively using a spectrophotometer (Nano-Drop 2100, Thermo Scientific, Waltham, MA, USA) and then cDNA was synthesized (PCR, Thermo Scientific, Waltham, MA, USA). PrimeScript™ RT-PCR kits (Takara, Shiga, Japan) were adopted for cDNA synthesis in accordance with the instructions. A 20 μL total reaction volume of cDNA was prepared, comprising dNTP Mixture (1 μL), Oligo dT Primer (1 μL), Template RNA (4 μL), and RNase-Free dH2O (4 μL). The following amplification conditions were programmed: denaturation at 65 °C for 5 min and then at 4 °C. The reaction solution was taken out after annealing and 5× Prime Script Buffer (4 μL), RNase Inhibitor (0.5 μL), Prime Script RTase (0.5 μL), and RNase-Free dH2O (5 μL) were added. Amplification conditions were set as follows: denaturation at 42 °C for 15 min, 95 °C for 5 min, and 4 °C.
The cDNA fragment of Rpcl was obtained from NCBI transcriptome data (GenBank accession No. MH107141.1). The primers are shown in Table 1. Target PCR products were ligated to T vector pMD 19 (Takara Biotechnology (Dalian) Co., Ltd., Dalian, China), followed by transforming into DH5α E. coli receptor cells (Jiangsu Cowin Biotech Co., Ltd., Jiangsu, China) and sequencing (BGI Genomics Co., Ltd., Shenzhen, China).
2.3. Sequence Analysis of Rpcl
The corresponding amino acid sequences were derived from the full Rpcl cDNA sequence using DNAman software. BLAST algorithm (http://www.ncbi.nlm.nih.gov/blast (accessed on 20 November 2021)) was adopted for the analysis of cDNA and amino acid sequences of Rpcl. The compute pI/Mw tool (ExPASy database, https://web.expasy.org/compute_pi/ (accessed on 20 November 2021)) was utilized to analyze the isoelectric point and molecular weight of Rpcl. SignalP-3.0 (http://www.cbs.dtu.dk/services/SignalP/ (accessed on 20 November 2021)) was utilized for predicting signal peptides (SP). RosettaFold (https://robetta.bakerlab.org/ (accessed on 20 November 2021)) was utilized for 3D structure prediction. MEGA software was applied to construct the phylogenic trees.
2.4. qRT-PCR
The SYBR® Premix Ex Taq (Tli RNaseH Plus) kits (TaKaRa, Shiga, Japan) were adopted for performing qRT-PCR of β-actin and Rpcl following the kit instructions. The reactions of a total volume of 20 μL were prepared for qRT-PCR, including 2× SYBR Green Mix (10 μL), cDNA 2 (μL), forward and reverse primers (1 μL each), and water (6 μL) [18]. When the PCR reaction was completed, the amplification products were analyzed for dissociation curves to verify the amplification of only one PCR product. Software was used to automatically set baselines, to maintain consistency. The 2−ΔΔCt method was conducted for the analysis of Rpcl expression level [19]. All data were described by the mean ± SE of relative mRNA expression.
2.5. Recombinant Rpcl Protein Expression
Primers Rpcl-NdeI-F and Rpcl-BamHI-R (Table 1) containing restriction sites were designed according to the open reading frame (ORF) sequence of the Rpcl gene, and the ORF sequence was amplified using cDNA as template. The pMD-19T-Rpcl recombinant plasmid and pET-28a blank plasmid were extracted by using the plasmid extraction kit, double digested with NdeI and BamHI to recover the target fragment, and the pET-28a vector was ligated with T4DNA ligase to construct the pET-28a-Rpcl recombinant plasmid. The recombinant plasmids were transformed into DH5α, screened for positive clones, and sequenced for validation. Based on the sequencing results, the recombinant plasmids were then extracted and transformed into BL21(DE3) cells to construct the recombinant expression strain BL21/pET28a-Rpcl, which was then inoculated in LB liquid medium (supplemented with kanamycin 50 μg/mL) for expanded culture. The culture was grown at 37 °C until the OD value was between 0.4 and 0.6, and the inducer IPTG with a final concentration of 0.5 mmol/L was added and induced overnight at 28 °C. The control group was the recombinant solution without the inducer. The bacterial broth was gathered through centrifugation at 10,000r/min for 10 min at 4 °C, followed by adding it to binding buffer (20 mmol/L Tris-HCl, 0.5 mol/L NaCl) and ultrasonically crushing it. The hetero protein was washed twice with inclusion body washing solution, then the inclusion body was lysed with inclusion body lysis solution, and the recombinant protein was purified according to the instruction manual of His60NiSuperflowResin purification column (TaKaRa, Shiga, Japan). The BCA kit (Beyotime, Shanghai, China) was utilized for determination of purified protein concentration. The purified protein was detected with 12% polyacrylamide gel electrophoresis (SDS-PAGE).
2.6. Agglutination Activity of Rpcl
The experimental microalgae were grown to logarithmic growth phase, as described formerly [20]. Microalgae were collected for 15 min of centrifugation at 3500× g and rinsed two times with TBS, followed by resuspending in TBS containing CaCl2 (10 mM) using the method described previously [21]. Then, 50 μL microalgae suspension (1 × 107 cells/mL) was incubated for 2 h with 50 µL Rpcl (0.047 mg/mL) at indoor temperature. As a negative control, the microalgae suspension (50 µL) was added to CaCl2 (10 mM) in TBS. The above experimental group and control group were incubated in 96-well plates. The incubation solution (10 μL) was placed on a slide for observation. The agglutination phenomena were all set up in 3 sets of replicates, and a light microscope was utilized to view the results of agglutination.
2.7. Expression of Rpcl in the Lip and Gill of R. philippinarum after Feeding Microalgae
The two microalgae (C. debilis and K. veneficum) were grown to logarithmic period. Fifteen R. philippinarum were placed in an aerated box mixed with 500 mL of algal culture (1 × 106 cells/mL) and 500 mL of seawater as the experimental group. R. philippinarum was starved under the same conditions and was set as the control group. Lip and gill were gathered at 0 h, 0.5 h, 1 h, 2 h, and 4 h of rearing, respectively. All the groups were designed with 6 experimental replicates. The mRNA expression of Rpcl was detected.
2.8. Statistical Methods
Data analysis was conducted using SPSS 20.0 software (2011, IBM SPSS Inc., Chicago, IL, USA). Tukey’s HSD multiple comparisons test and one-way analysis of variance (ANOVA) were utilized for comparisons between the results among tissues. A t-test was applied for the analysis of intergroup differences. Statistically significant threshold was regarded as p < 0.05.
3. Results
3.1. Molecular Characterization of Rpcl
The full cDNA sequence of Rpcl comprised 930 bp (GenBank accession number MH107141.1), involving a 5′-untranslated region (UTR) of 18 bp, a 3′ UTR of 192 bp with a poly (A) tail, as well as an ORF of 720 bp, which encoded 239 amino acids. Bioinformatics analyses revealed that Rpcl contained an SP with no transmembrane region, which had an isoelectric point of 5.82 and a molecular weight of 27.94 kDa. Rpcl had a CRD containing 118 amino acid residues and five cysteine residues (Figure 1). The homology sequence alignment results show that Rpcl shares high similarity with the 1-CRD type CTL sequences of other mollusks (Figure 2). Phylogenetic analyses revealed that the amino acid sequence of Rpcl was closely related to that of Vpclec-1 from R. philippinarum and CTL from Mercenaria (Figure 3). The protein structure prediction showed that the Rpcl contains one calcium ion binding site, and have the binding sites of saccharides listed as follows: N-acetylglucosamine, galactose, mannose, and trehalose (Figure 4).
3.2. Tissue Distribution of Rpcl
To study Rpcl tissue distribution, total RNA was separated from eight tissues. In the dissociation curve analysis, only one peak was detected for Rpcl and ß-actin genes at the corresponding melting temperatures, indicating PCR specificity. The mRNA transcript of Rpcl expression was observed in all tissues detected (Figure 5), with the order of expression from highest to lowest being the following: gill > hepatopancreas > mantle > lip > adductor muscle (p < 0.05).
3.3. Heterologous Expression of Rpcl
The recombinant proteins were expressed as inclusion bodies. SDS-PAGE acquired distinct bands from the precipitated ultrasonic fragmentation solution with a molecular weight of approximately 27.94 kDa. This is the Rpcl target protein band (Figure 6). Lane 1 is the protein molecule standard, lane 2 is the induced pET-28a, lane 3 is the negative control (Rpcl), lane 4 is the induced Rpcl (supernatant), lane 5 is the induced Rpcl (inclusion bodies), and lane 6 is the purified protein.
3.4. Microalgae Agglutinating Activity of Rpcl
The result showed that Rpcl had agglutination activities for C. debilis, K. veneficum, and C. pyrenoidosa, but had no agglutination for P. minimum. It had the strongest agglutination activity for C. pyrenoidosa. However, it had similar agglutination with C. debilis and K. veneficum (Figure 7).
3.5. Expression of Rpcl in Feeding Experiments
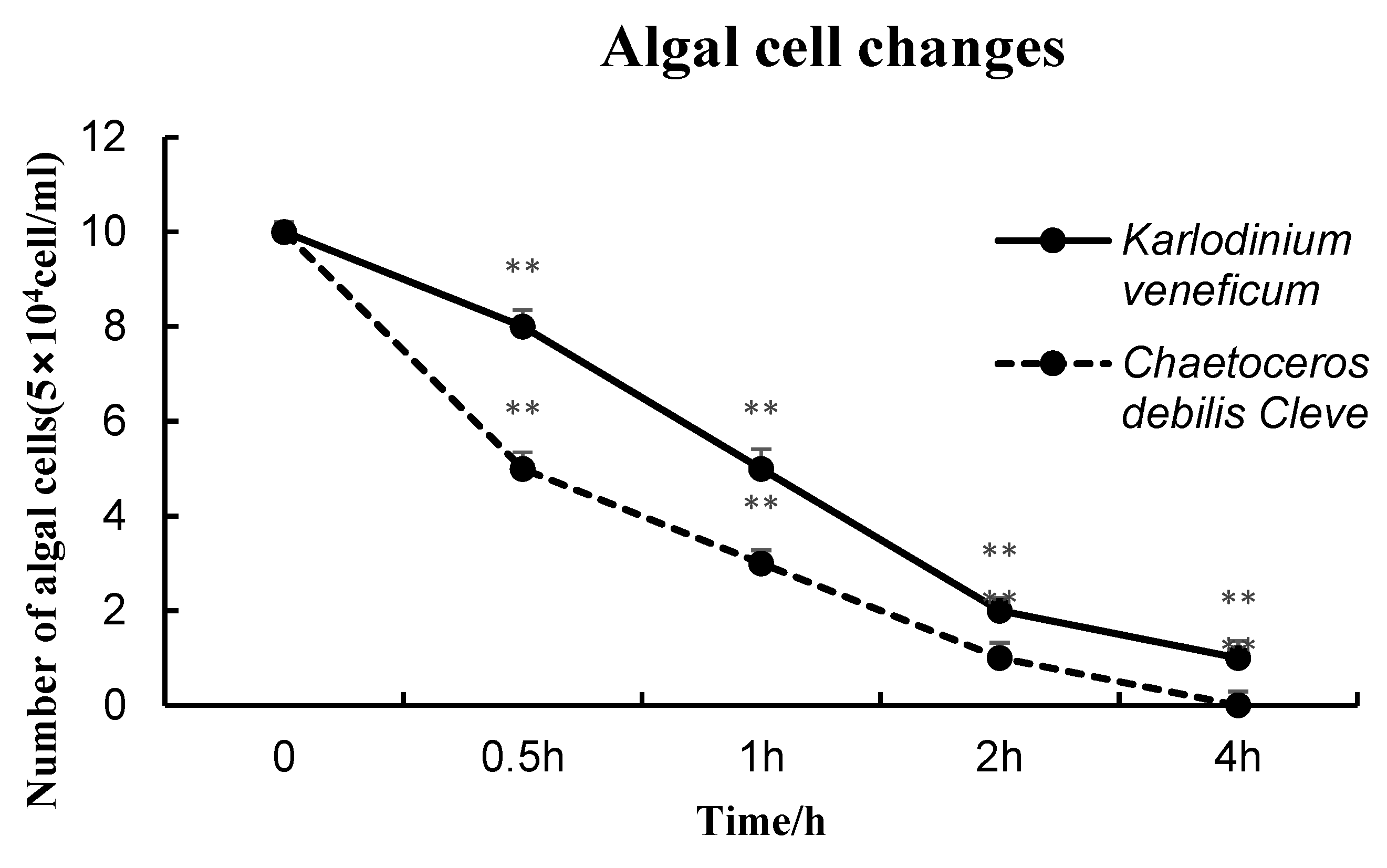
The Rpcl expression was significantly elevated at all times in the C. debilis group in contrast to the control of starving group (p < 0.05). The Rpcl expression was also significantly increased at all times in group of K. veneficum (p < 0.05, Figure 8). The gene expression in C. debilis group reached its first peak at 0.5 h, followed by a process of first decreasing and then increased back to another peak at 4 h. In K. veneficum group, the gene expression reached its peak at 1 h after feeding and declined afterward (Figure 8). In the gill tissues (Figure 9), the variation trends of the gene expressions were relatively similar as that in lips. However, compared with the two experimental groups, the expression of Rpcl in C. debilis was markedly higher compared with that in K. veneficum in most periods. In addition, it was found that the changes in the expression of the Rpcl gene in the gill and lip, the main feeding tissues, were consistent with the slope of the decrease in the number of algal cells in the water body (Figure 8, Figure 9 and Figure 10). The expression of lectins in the lip was more significant than that in the gill. Thus, it was also verified that lip lectins play a deterministic role in feeding.
4. Discussion
Invertebrates are known to contain many different molecular forms of CTLs [22]. The CTL structural domain consists of 110 to 130 amino acid residues and is greatly conserved in its structural folds. In CTLs, the lectin or lectin-like structural domain accepts various carbohydrate and non-carbohydrate ligands, including proteins and lipids [5,22]. A single lectin can bind to multiple sugar ligands and differentiate tiny structural distinctions in closely associated ligands [23]. Calcium ions form multiple ligand bonds with well-conserved amino acid residues in CTLs [24,25]. The two adjacent hydroxyl groups form hydrogen bonds with EPN or QPD motifs and coordination bonds with calcium ions in monosaccharides [26]. Motifs refer to the residues defining sugar binding. In all CTLs, coordination bonds are formed with Ca2+ via a carbonyl side chain and asparagine (Asn) and aspartate (Asp) from the WND (Trp-Asn-Asp) motif [24,27]. In addition, the EPN (Glu-Pro-Asn) motif or QPD (Gln-Pro-Asp) motif [28,29] contributed to the coordination of calcium as well as the formation of sugar-binding sites. The CTL-like structural domain will not bind calcium ions if it lacks such conserved motifs. The number of CRDs is another factor affecting the CTL binding capacity [30]. The protein may identify and bind more carbohydrate species through multiple CRD sites in invertebrates.
In this research, we characterized a CTL with a single CRD in R. philippinarum and investigated its role in feeding and feed selection. Rpcl has high sequence similarity to other mollusks such as Mercenaries mercenaria and is very similar to the previously reported CTL sequence of R. philippinarum. In addition, Rpcl aggregates phylogenetically to the identical subgroup as CTLs of other bivalve mollusks. Multiple CTL molecular features were found in Rpcl, such as cysteine and Ca2+ binding sites involved in disulfide bond formation. Structurally, Rpcl belongs to the QPN type of the calcium-dependent QPD variant, which also belongs to a domain with high affinity to galactose. The RosettaFold was used to predict the protein structural domain of Rpcl, which was found to bind N-acetylglucosamine, galactose, mannose, and trehalose.
For bivalves such as M. edulis with sympatric ctenophores (gills with monofilament type), food particles can’t be taken qualitatively in gills [31,32], and lips are the important part for them to take food particles. Their specificity was highlighted in previous reports [32,33] using endoscopy, which showed that selection is limited to the lips and that gills are only involved in transporting granules to the tentacles for further processing in M. edulis. This information was enhanced by studies demonstrating the crucial part of lip mucosal lectins in the separation of particles. Indeed, the mucosal lectin MeML is present in both lips and gills, and sorting efficiency is only associated with the mRNA transcription of MeML in the lips [16]. Combining the results of the above previous studies, we proposed that the same conclusion applies to R. philippinarum. Therefore, in this study, the lip was mainly selected in feeding trials. Combining the above findings and the expression of tissue expression profiles, the gills and lips were mainly selected for feeding experiments in this study. The conclusion that infers that lip lectins play a definitive role in feeding experiments also applies to R. philippinarum.
Tissue distribution maps revealed that Rpcl expression takes place in all tissues, and mainly in those involved in feeding and digestion. Rpcl was most significantly expressed in the gills, followed by the hepatopancreas, mantle, and lip. During protein acquisition, Rpcl protein expression was mainly found in sediment and therefore belonged to inclusion body proteins. The result that Rpcl had agglutination for C. debilis, K. veneficum, and C. pyrenoidosa, but had no agglutination for P. minimum, showed the different agglutination abilities for different microalgae. The result that agglutination activities were relatively similar between C. debilis (nontoxic) and K. veneficum (toxic), indicated that, for Rpcl protein, microalgae of toxic or nontoxic play no roles in the agglutination. However, gene expression was significantly different when the two microalgae were applied in feeding the clams.
It is demonstrated that the protein Rpcl has selective agglutination for different microalgae. This result can be explained by the concept of ligand receptor proposed by Deniaud et al. [23]. Specific interactions of carbohydrates on the microalgae cell surfaces and lectins in the mucosal secretions of bivalve feeding organs determine the process of microalgal ingestion by bivalves [34]. Therefore, studying the characteristics of microalgal cell surface polysaccharides and their relationship with bivalve selective microalgae is likewise a key research direction. Based on the experimental results, future studies are necessary to analyze whether these four algal surfaces contain the sugar substances we predicted and in what amounts.
Understanding the particle selection of bivalves is important economically and ecologically. The results of lectin expression in the feeding experiment showed that the experimental groups of both microalgae exhibited significant differences at all time points compared to the starvation-treated control group. Although the in vitro purified protein Rpcl was consistent with the agglutination effect of both algae, the gene expression level of K. veneficum, the toxic dinoflagellate [35,36,37,38,39], was significantly lower than that of C. debilis, the beneficial bait of shellfish [40], which to some extent verifies that microalgae with toxicity can cause a series of extreme effects in bivalves [41], including valve closure, set constriction, inhibition of gill hair pulsation, and reduction in gap [42,43,44].
5. Conclusions
The cDNA of Rpcl obtained in this study is 930 bp long with a 720 bp ORF and encodes 235 amino acids, containing an N-terminal SP and a CRD. Rpcl comprises a conserved CRD disulfide bond including five cysteine residues (Cys125, Cys142, Cys213, Cys219, and Cys237) and the QPN (GLN204-PRO205-ASN206) motif. The QPN motif is a variant of the QPD motif, that substitutes aspartic acid with asparagine and has galactose-binding properties. It is also predicted to have calcium-binding site for N-acetylglucosamine, galactose, mannose, and trehalose binding. Rpcl expression was commonly observed in all detected tissues. The Rpcl expression from highest to lowest was in the following order: gills > hepatopancreas > mantle > lip > adductor muscle. The results of the in vitro agglutination assay of the purified protein Rpcl with microalgae and the upregulation of the Rpcl gene in the feeding assay fully demonstrate that lectins are involved in the feeding and feed selection process of R. philippinarum. In addition, the expression of Rpcl is more significant in bait microalgae than in toxic microalgae in feeding experiments, contrary to the agglutination effect of both microalgae with in vitro purified proteins. To some extent, this verifies that microalgae with toxicity can cause considerable effects on bivalves. From this study, evidence is provided that lectin may indirectly or directly involve in feeding and particle selection.
Author Contributions
Conceptualization, S.C.; methodology, S.C. and Q.F.; software and empirical analysis, X.Q.; validation, C.Z. and X.Y.; formal analysis, X.Q. and C.Z.; investigation, P.S.; resources, C.Z.; data curation, S.C. and Q.F.; writing—original draft preparation, S.C.; writing—review and editing, S.C. and C.Z.; supervision, C.Z. and J.X.; project administration, X.Y. and C.Z.; funding acquisition, C.Z., X.Y. and J.X. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the National Key Research and Development Program of China (No. 2018YFD0900702) and the National Science and Technology Basic Resources Investigation Program of China (No. 2018FY100206).
Institutional Review Board Statement
The animal study protocol complied with the ARRIVE guidelines and was carried out in accordance with the UK Animals (Scientific Procedures) Act, 1986, and the associated guidelines.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bureau of Fisheries, Ministry of Agriculture and Rural Affairs, National Fisheries Technology Extension Center, China Society of Fisheries. China Fishery Statistical Yearbook 2021; China Agriculture Press: Beijing, China, 2021.
- Zuo, S.; Jiang, K.; Li, D.; Yan, X.; Nie, H. Transcriptomic analysis of Manila clam Ruditapes philippinarum under lipopolysaccharide challenge provides molecular insights into immune response. Fish Shellfish. Immunol. 2020, 106, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Mckindsey, C.W.; Landry, T.; O’Beirn, F.X.; Davies, I.M. Bivalve aquaculture and exotic species: A review of ecological considerations and management issues. J. Shellfish. Res. 2007, 26, 281–294. [Google Scholar] [CrossRef]
- Vasta, G.; Feng, C.; Bianchet, M.; Bachvaroff, T.; Tasumi, S. Structural, functional, and evolutionary aspects of galectins in aquatic mollusks: From a sweet tooth to the Trojan horse. Fish Shellfish. Immunol. 2015, 46, 94–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef]
- Drickamer, K. Two distinct classes of carbohydrate-recognition domeins in animal’s lectins. J. Biol. Chem. 1988, 263, 9557–9560. [Google Scholar] [CrossRef]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-type lectin superfamily in the immune system. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef]
- Kikuno, R.; Sato, A.; Mayer, W.E.; Shintani, S.; Aoki, T.; Klein, J. Clustering of C-type lectin natural killer receptor-like loci in the bony fish Oreochromis niloticus. Scand. J. Immunol. 2004, 59, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Hurtado, C.; Granja, A.G.; Bustos, M.J.; Nogal, M.L.; de Buitrago, G.G.; de Yébenes, V.G.; Salas, M.L.; Revilla, Y.; Carrascosa, A.L. The C-type lectin homologue gene (EP153R) of African swine fever virus inhibits apoptosis both in virus infection and in heterologous expression. Virology 2004, 326, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.-N.; Jin, X.-K.; Li, S.; Yu, A.-Q.; Wu, M.-H.; Tan, S.-J.; Zhu, Y.-T.; Li, W.-W.; Zhang, P.; Wang, Q. A novel C-type lectin from Eriocheir sinensis functions as a pattern recognition receptor with antibacterial activity. Fish Shellfish. Immunol. 2013, 35, 1554–1565. [Google Scholar] [CrossRef]
- Hosono, M.; Sugawara, S.; Ogawa, Y.; Kohno, T.; Takayanagi, M.; Nitta, K. Purification, characterization, cDNA cloning, and expression of asialofetuin-binding C-type lectin from eggs of shishamo smelt (Osmerus [Spirinchus] lanceolatus). Biochim. Biophys. Acta—Gen. Subj. 2005, 1725, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, E.P.; Allam, B. Food quality and season affect gene expression of the mucosal lectin MeML and particle sorting in the blue mussel Mytilus edulis. Mar. Biol. 2013, 160, 1441–1450. [Google Scholar] [CrossRef]
- Shi, Y.; Zhao, X.; Wang, Z.; Shao, Y.; Zhang, W.; Bao, Y.; Li, C. Novel Ca2+-independent C-type lectin involved in immune defense of the razor clam Sinonovacula constricta. Fish Shellfish. Immunol. 2019, 84, 502–508. [Google Scholar] [CrossRef]
- Espinosa, E.P.; Allam, B. Reverse genetics demonstrate the role of mucosal C-type lectins in food particle selection in the oyster Crassostrea virginica. J. Exp. Biol. 2018, 221, jeb174094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa, E.P.; Perrigault, M.; Allam, B. Identification and molecular characterization of a mucosal lectin (MeML) from the blue mussel Mytilus edulis and its potential role in particle capture. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Alhoshy, M.; Shehata, A.I.; Habib, Y.J.; Abdel-Latif, H.M.; Wang, Y.; Zhang, Z. Nutrigenomics in crustaceans: Current status and future prospects. Fish Shellfish. Immunol. 2022, 129, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Nie, H.; Dong, S.; Huo, Z.; Yan, X. Molecular cloning and expression analysis of C-type lectin (RpCTL) in Manila clam Ruditapes philippinarum after lipopolysaccharide challenge. Fish Shellfish. Immunol. 2019, 86, 981–993. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lou, Y.; Lai, W.; Xu, J.; Zhou, C.; Yan, X. Effect of polyamines on amplification of several marine microalgae. J. Ningbo. Univ. 2018, 31, 38–43. [Google Scholar]
- Espinosa, E.P.; Hassan, D.; Ward, J.E.; Shumway, S.E.; Allam, B. Role of Epicellular Molecules in the Selection of Particles by the Blue Mussel, Mytilus edulis. Biol. Bull. 2011, 219, 50–60. [Google Scholar] [CrossRef]
- Sancho, D.; e Sousa, C.R. Sensing of cell death by myeloid C-type lectin receptors. Curr. Opin. Immunol. 2013, 25, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Deniaud, D.; Julienne, K.; Gouin, S.G. Insights in the rational design of synthetic multivalent glycoconjugates as lectin ligands. Org. Biomol. Chem. 2011, 9, 966–979. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, M.; Zhang, H.; Song, L. The immune role of C-type lectins in molluscs. Invertebr. Surviv. J. 2011, 8, 241–246. [Google Scholar]
- Zhang, H.; Wang, H.; Wang, L.; Song, L.; Song, X.; Zhao, J.; Li, L.; Qiu, L. Cflec-4, a multidomain C-type lectin involved in immune defense of Zhikong scallop Chlamys farreri. Dev. Comp. Immunol. 2009, 33, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Zelensky, A.N.; Gready, J.E. C-type lectin-like domains in Fugu rubripes. BMC genomics 2004, 5, 51. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, E.P.; Perrigault, M.; Ward, J.E.; Shumway, S.E.; Allam, B. Lectins Associated with the Feeding Organs of the Oyster Crassostrea virginica Can Mediate Particle Selection. Biol. Bull. 2009, 217, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Zhang, H.; Jiang, S.; Wang, M.; Wang, L.; Song, L. Comparative study of two single CRD C-type lectins, CgCLec-4 and CgCLec-5, from pacific oyster Crassostrea gigas. Fish Shellfish. Immunol. 2016, 59, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Zhang, H.; Jiang, S.; Wang, L.; Liu, R.; Yi, Q.; Song, L. An EPD/WSD motifs containing C-type lectin from Argopectens irradians recognizes and binds microbes with broad spectrum. Fish Shellfish. Immunol. 2015, 43, 287–293. [Google Scholar] [CrossRef]
- Cambi, A.; Koopman, M.; Figdor, C.G. How C-type lectins detect pathogens. Cell. Microbiol. 2005, 7, 481–488. [Google Scholar] [CrossRef]
- Beninger, P.; St-Jean, S. Particle processing on the labial palps of Mytilus edulis and Placopecten magellanicus (Mollusca:Bivalvia). Mar. Ecol. Prog. Ser. 1997, 147, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.E.; Levinton, J.S.; Shumway, S.E.; Cucci, T. Particle sorting in bivalves: In vivo determination of the pallial organs of selection. Mar. Biol. 1998, 131, 283–292. [Google Scholar] [CrossRef]
- Ward, J.E.; Levinton, J.S.; Shumway, S.E.; Cucci, T. Site of particle selection in a bivalve mollusc. Nature 1997, 390, 131–132. [Google Scholar] [CrossRef]
- Espinosa, E.P.; Perrigault, M.; Ward, J.E.; Shumway, S.E.; Allam, B. Microalgal Cell Surface Carbohydrates as Recognition Sites for Particle Sorting in Suspension-Feeding Bivalves. Biol. Bull. 2010, 218, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegaret, H.; Wikfors, G.H.; Shumway, S.E. Diverse feeding responses of five species of bivalve mollusc when exposed to three species of harmful algae. J. Shellfish. Res. 2007, 26, 549–559. [Google Scholar] [CrossRef]
- Turner, A.D.; Lewis, A.M.; Bradley, K.; Maskrey, B.H. Marine invertebrate interactions with Harmful Algal Blooms—Implications for One Health. J. Invertebr. Pathol. 2021, 186, 107555. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Yin, B.; Wang, Y.; Huai, B. Significantly decreasing harmful algal blooms in China seas in the early 21st century. Mar. Pollut. Bull. 2019, 139, 270–274. [Google Scholar] [CrossRef]
- McNamee, S.E.; Medlin, L.K.; Kegel, J.; McCoy, G.R.; Raine, R.; Barra, L.; Ruggiero, M.V.; Kooistra, W.H.; Montresor, M.; Hagstrom, J.; et al. Distribution, occurrence and biotoxin composition of the main shellfish toxin producing microalgae within European waters: A comparison of methods of analysis. Harmful Algae 2016, 55, 112–120. [Google Scholar] [CrossRef]
- Karlson, B.; Andersen, P.; Arneborg, L.; Cembella, A.; Eikrem, W.; John, U.; West, J.J.; Klemm, K.; Kobos, J.; Lehtinen, S.; et al. Harmful algal blooms and their effects in coastal seas of Northern Europe. Harmful Algae 2021, 102, 101989. [Google Scholar] [CrossRef] [PubMed]
- Robbins, H.M.; Bricelj, V.M.; Ward, J.E. In vivo Effects of Brown Tide on the Feeding Function of the Gill of the Northern Quahog Mercenaria mercenaria (Bivalvia: Veneridae). Biol. Bull. 2010, 219, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, Y.; Uchida, T.; Honjo, T. Toxic effects of the dinoffagellate Heterocapsa circularisquama on clearance rate of the blue mussel Mytilus galloprovincialis. Oceanogr. Lit. Rev. 1997, 7, 731. [Google Scholar]
- Shumway, S.E. A Review of the Effects of Algal Blooms on Shellfish and Aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, Z.; Deng, Y.; Tang, Y.Z. Evidence for resting cyst production in the cosmopolitan toxic dinoflagellate Karlodinium veneficum and the cyst distribution in the China seas. Harmful Algae 2020, 93, 101788. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Deduced amino acid sequences and complete nucleotides of Rpcl. The underline is SP, the gray shading is CRD, and the C circled in red is the conserved cysteine.
Figure 1.
Deduced amino acid sequences and complete nucleotides of Rpcl. The underline is SP, the gray shading is CRD, and the C circled in red is the conserved cysteine.
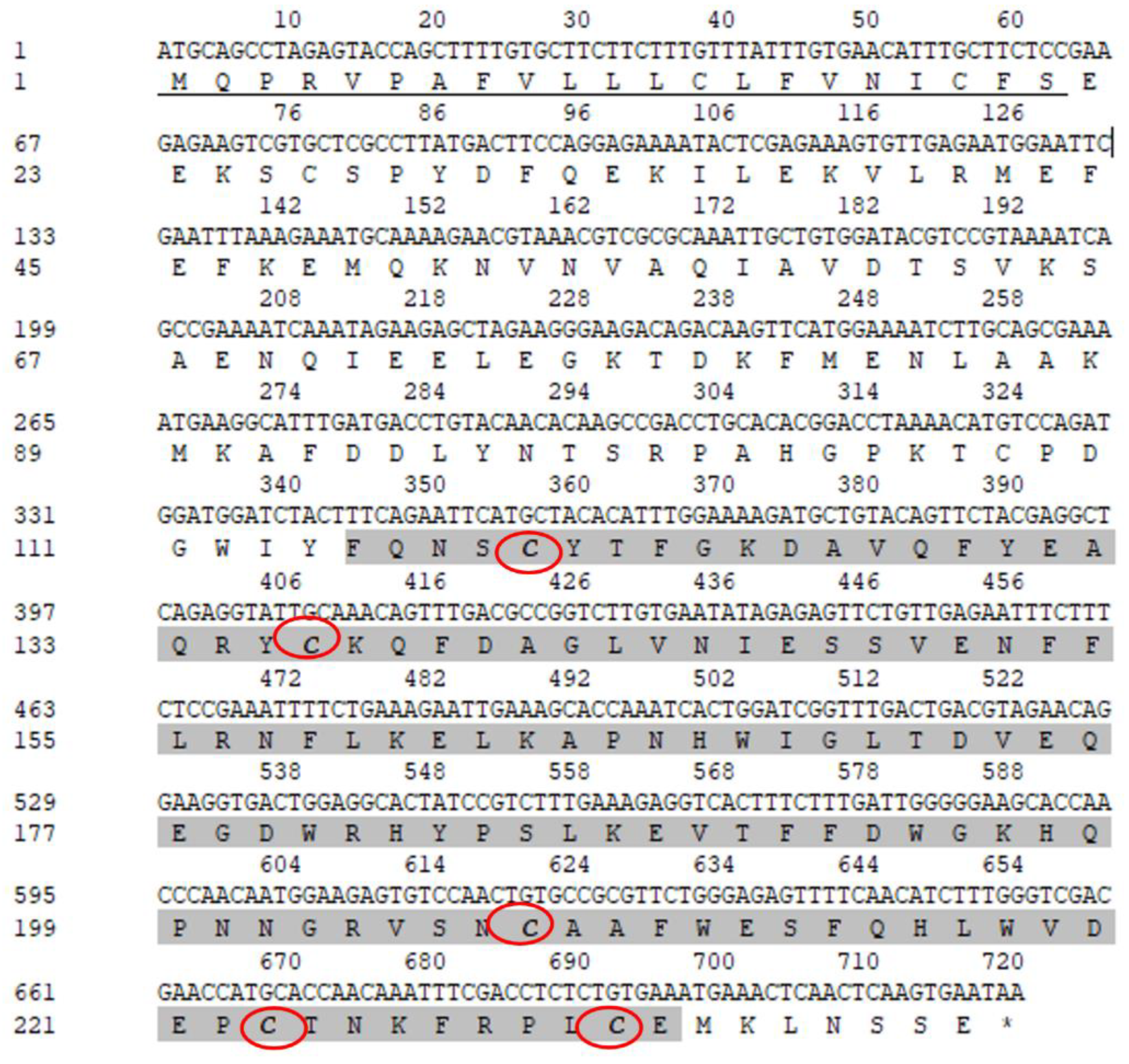
Figure 2.
Multiple alignment of amino acid sequences of CTL, Rpcl, and other mollusk lectins. Black parts are 100% similar amino acids, and the gray parts are 75% similar ones. Conserved structures (QPN) are indicated by red boxes. The GenBank accession numbers of proteins are listed: Crassostrea virginica (XP_022328732.1), Mercenaria (XP_045180913.1), Meretrix meretrix (ANS83633.1), Pinctada fucata (ACO36046.1), Rpcl (AZS54112.1), and vpclec-1 (AZS54109.1).
Figure 2.
Multiple alignment of amino acid sequences of CTL, Rpcl, and other mollusk lectins. Black parts are 100% similar amino acids, and the gray parts are 75% similar ones. Conserved structures (QPN) are indicated by red boxes. The GenBank accession numbers of proteins are listed: Crassostrea virginica (XP_022328732.1), Mercenaria (XP_045180913.1), Meretrix meretrix (ANS83633.1), Pinctada fucata (ACO36046.1), Rpcl (AZS54112.1), and vpclec-1 (AZS54109.1).

Figure 3.
Phylogenetic analysis of Rpcl and other CTLs. MEGA X was utilized to implement evolutionary analyses, and Neighbor-Joining method was applied to infer the evolutionary history. In the bootstrap test, the percentage of replicate trees where relevant taxa were clustered together (1000 replications) was indicated close to branches. We drew the tree to scale, and the units of branch length are identical to those of the evolutionary distances utilized for inferring phylogenetic trees. The Poisson correction method was performed to calculate the evolutionary distances in units of amino acid substitutions per site.
Figure 3.
Phylogenetic analysis of Rpcl and other CTLs. MEGA X was utilized to implement evolutionary analyses, and Neighbor-Joining method was applied to infer the evolutionary history. In the bootstrap test, the percentage of replicate trees where relevant taxa were clustered together (1000 replications) was indicated close to branches. We drew the tree to scale, and the units of branch length are identical to those of the evolutionary distances utilized for inferring phylogenetic trees. The Poisson correction method was performed to calculate the evolutionary distances in units of amino acid substitutions per site.

Figure 4.
The structure of Rpcl predicted by RosettaFold: the N- and C-terminal domains are indicated and differently colored. The Ca2+-binding site is enlarged and shown in the right panel.
Figure 4.
The structure of Rpcl predicted by RosettaFold: the N- and C-terminal domains are indicated and differently colored. The Ca2+-binding site is enlarged and shown in the right panel.
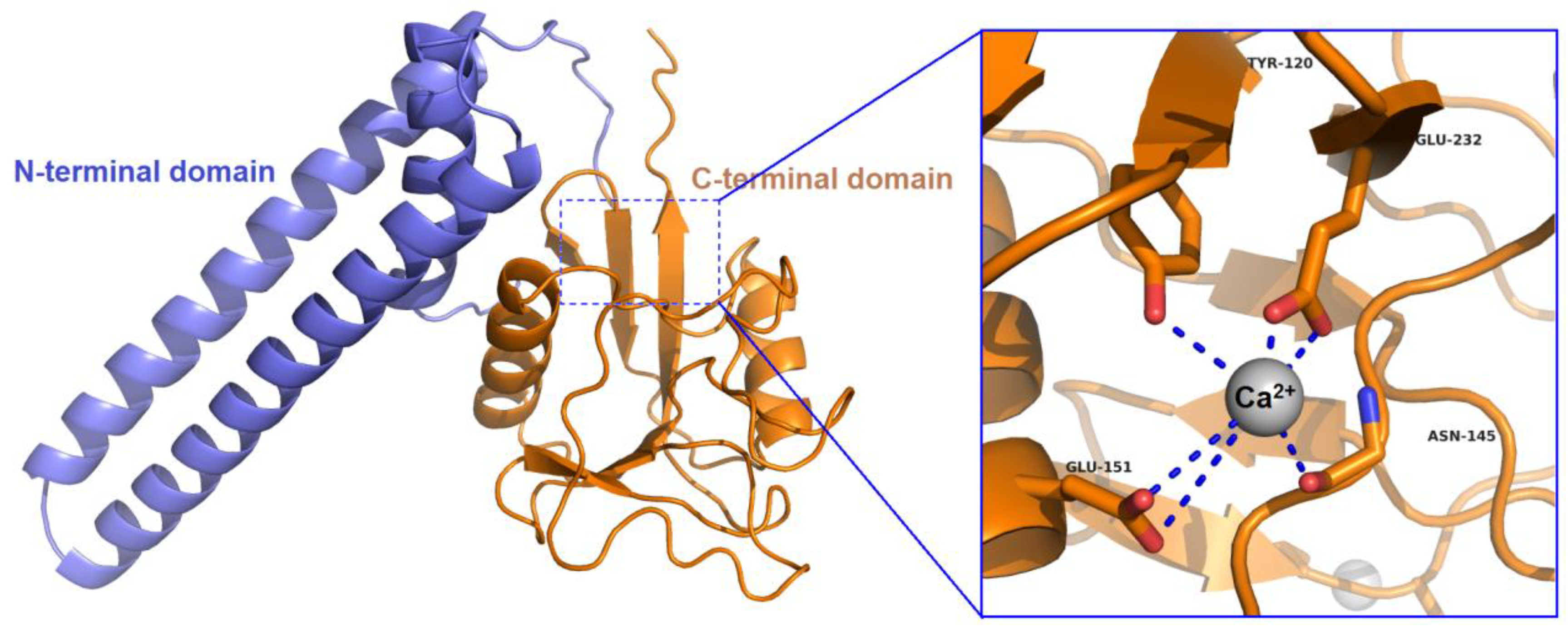
Figure 5.
The distribution of Rpcl in various tissues of R. philippinarum using qRT-PCR. Different letters represent significant differences. Three biological replicates for each tissue sample, and the data are described by mean ± SD (n = 6). Tukey HSD multiple comparisons and one-way ANOVA were carried out for analysis of statistically significant differences. Diverse letters denote significant differences, and p < 0.05.
Figure 5.
The distribution of Rpcl in various tissues of R. philippinarum using qRT-PCR. Different letters represent significant differences. Three biological replicates for each tissue sample, and the data are described by mean ± SD (n = 6). Tukey HSD multiple comparisons and one-way ANOVA were carried out for analysis of statistically significant differences. Diverse letters denote significant differences, and p < 0.05.

Figure 6.
SDS-PAGE analyses of Rpcl protein.
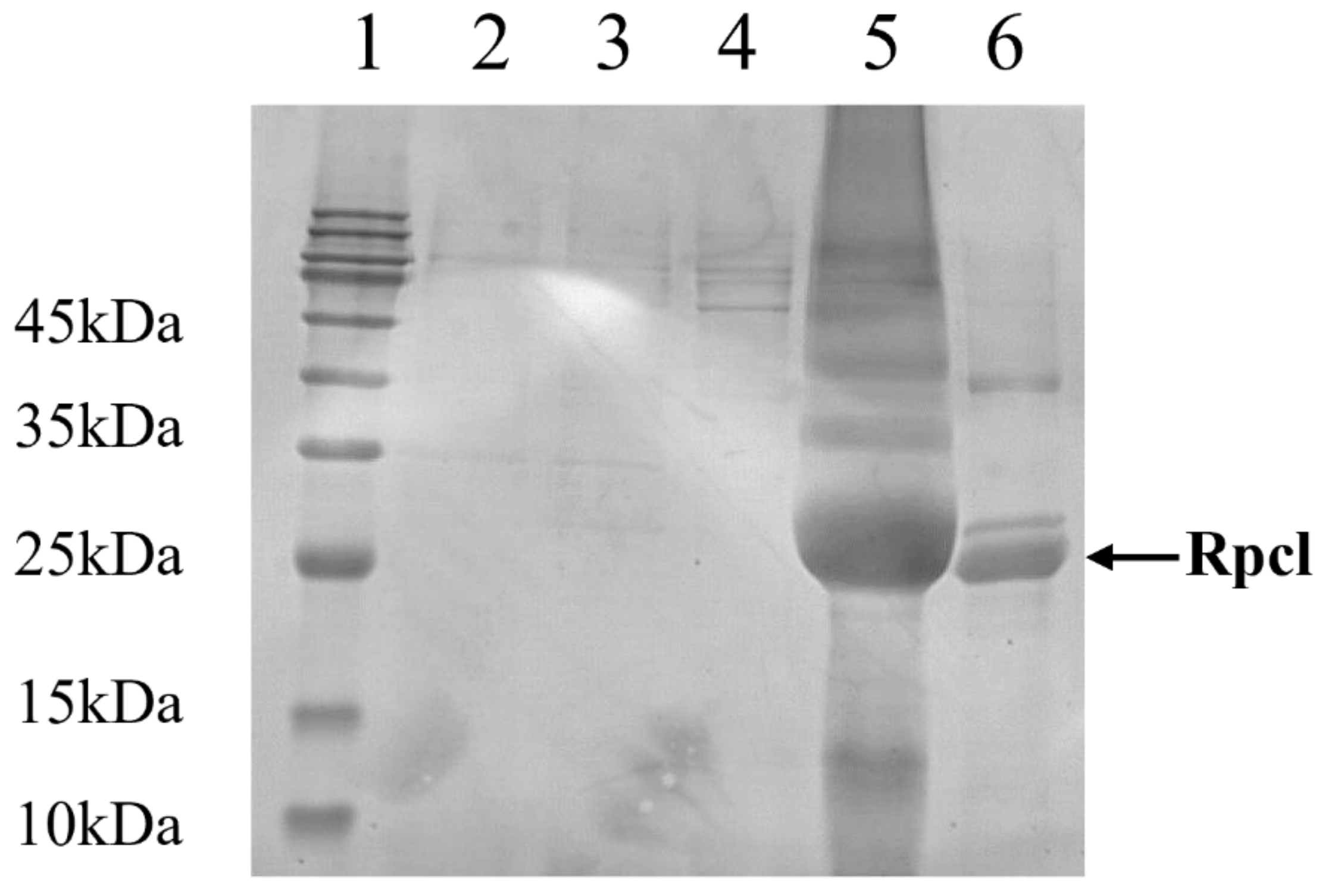
Figure 7.
Microalgae agglutination assay of Rpcl. Rpcl: microalgae were cultured in TBS containing Rpcl and CaCl2 (10 mM). Control: microalgae were cultured in TBS containing CaCl2 (10 mM) (the red box shows the protein agglutination in the microalgae).
Figure 7.
Microalgae agglutination assay of Rpcl. Rpcl: microalgae were cultured in TBS containing Rpcl and CaCl2 (10 mM). Control: microalgae were cultured in TBS containing CaCl2 (10 mM) (the red box shows the protein agglutination in the microalgae).
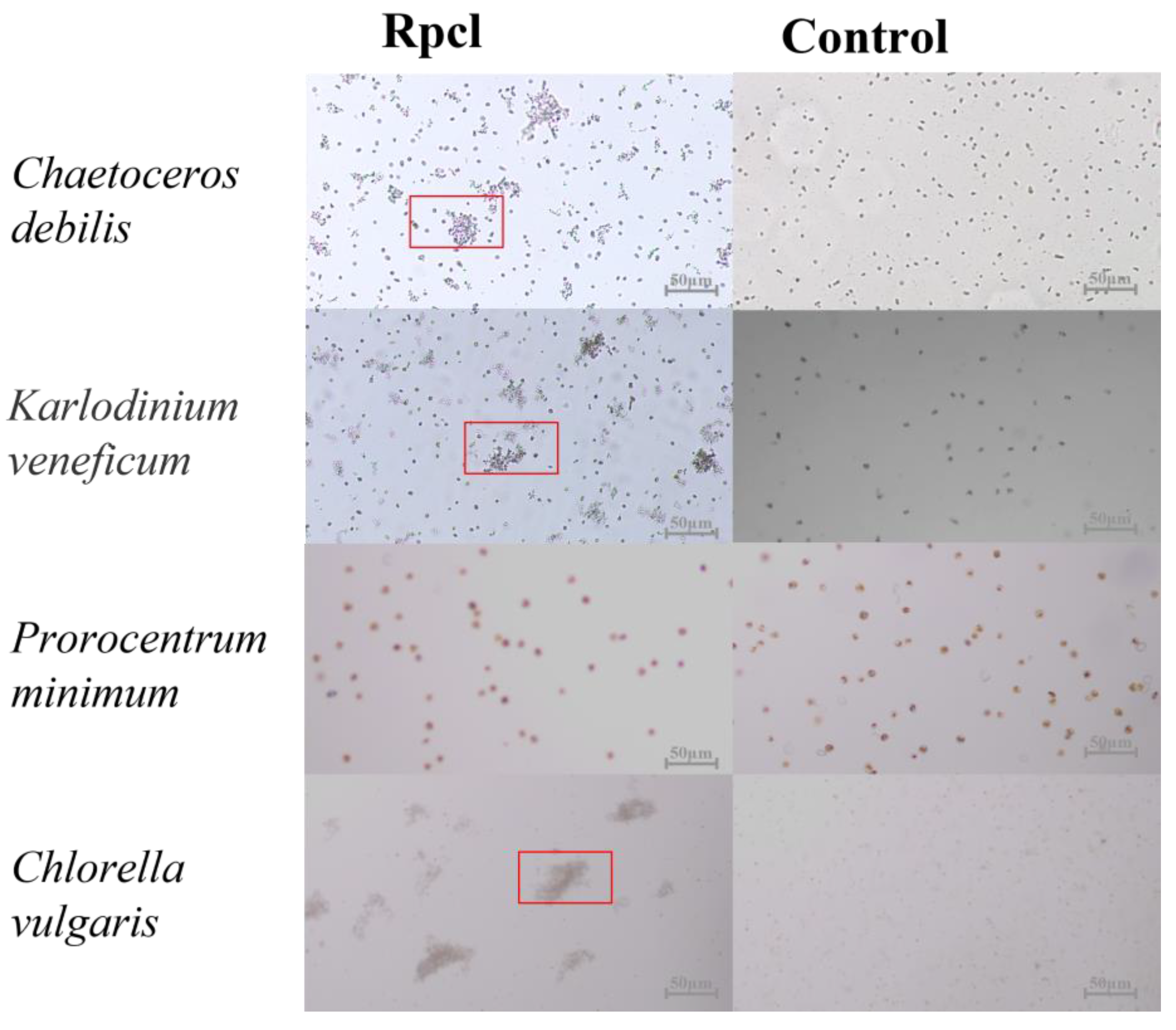
Figure 8.
Expression levels of Rpcl in the lip of R. philippinarum after feeding with Chaetoceros debilis and Karlodinium veneficum: * p < 0.05, ** p < 0.01.
Figure 8.
Expression levels of Rpcl in the lip of R. philippinarum after feeding with Chaetoceros debilis and Karlodinium veneficum: * p < 0.05, ** p < 0.01.
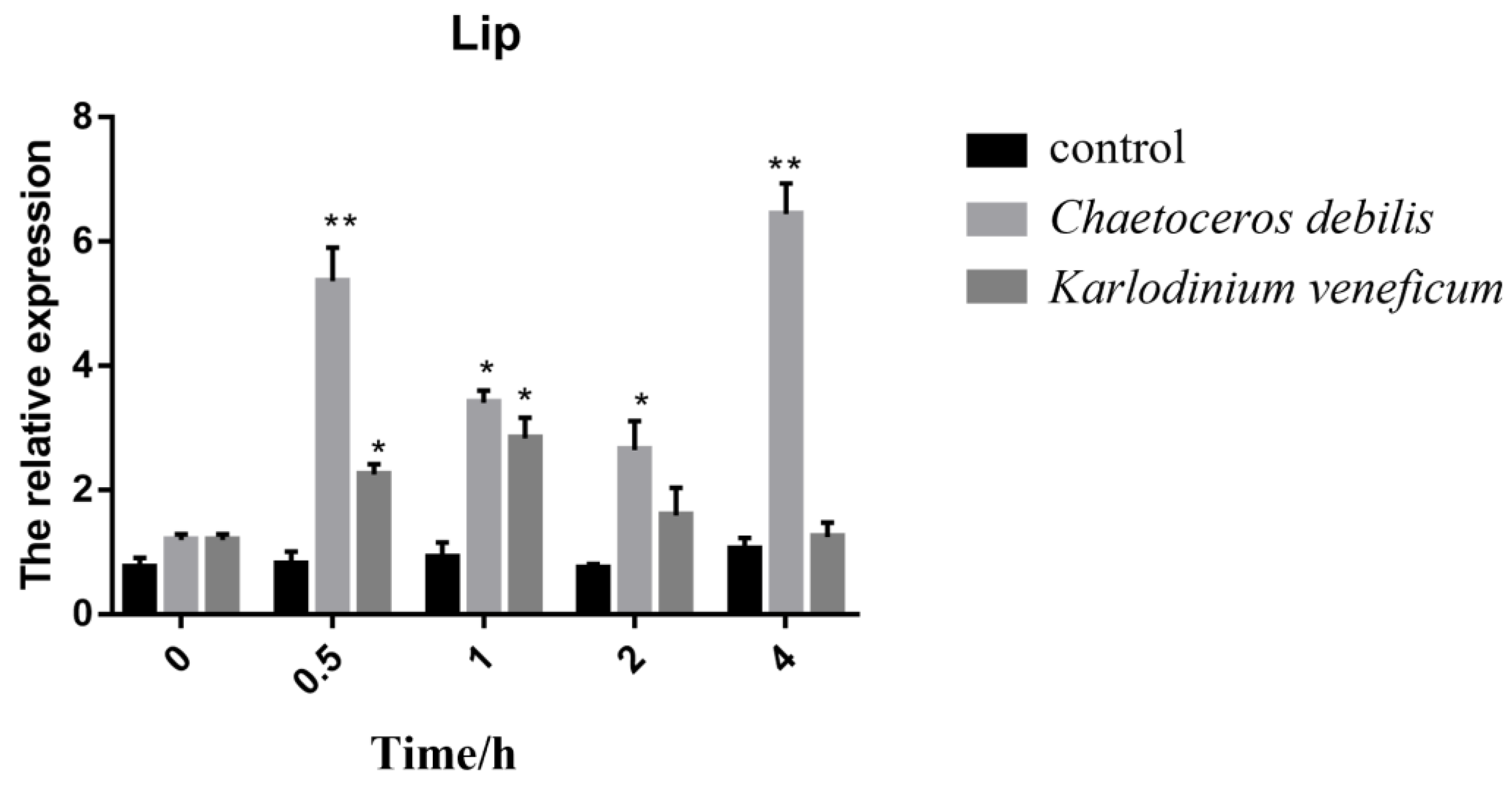
Figure 9.
Expression levels of Rpcl in the gill of R. philippinarum after feeding with Chaetoceros debilis and Karlodinium veneficum: * p < 0.05, ** p< 0.01.
Figure 9.
Expression levels of Rpcl in the gill of R. philippinarum after feeding with Chaetoceros debilis and Karlodinium veneficum: * p < 0.05, ** p< 0.01.
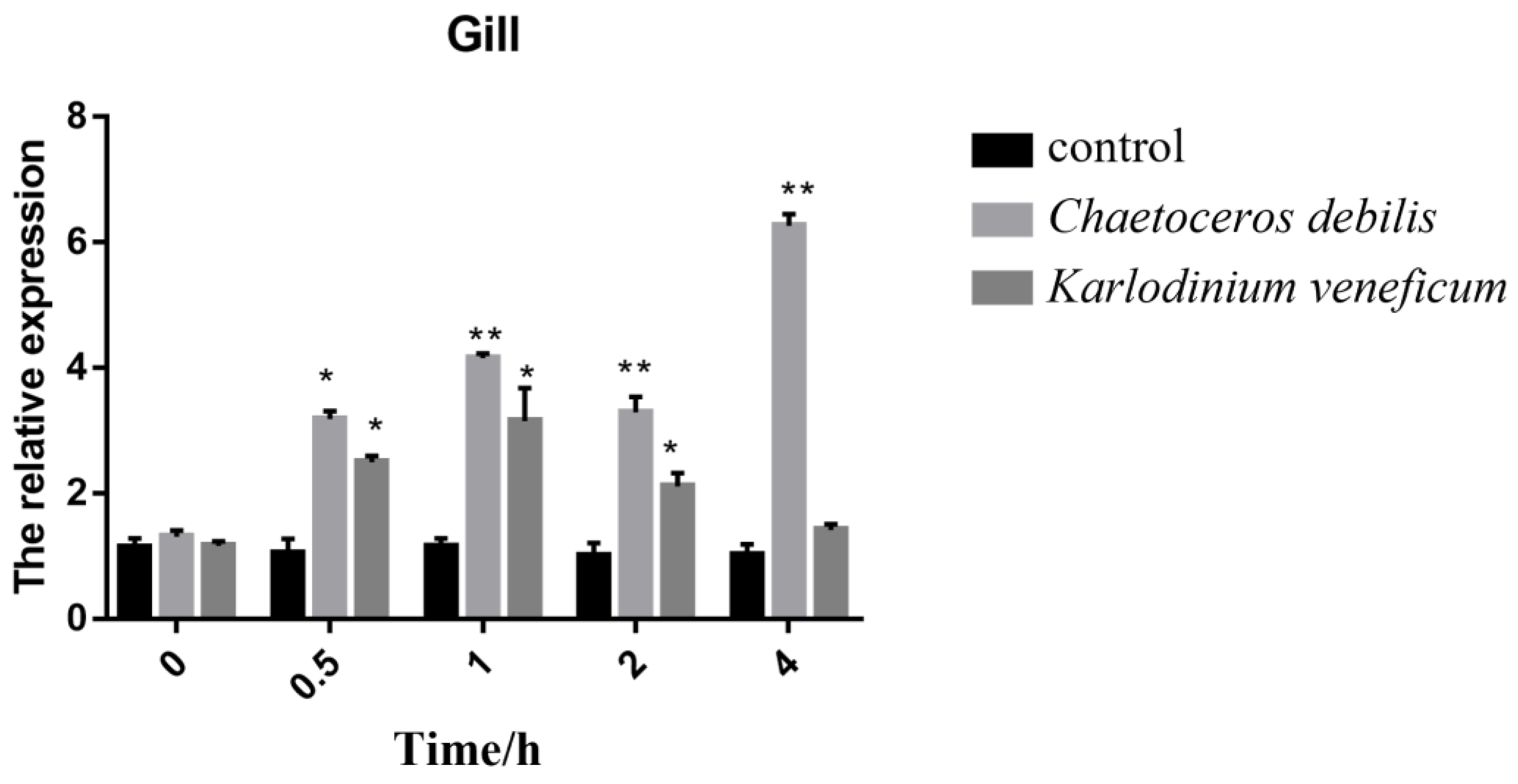
Figure 10.
Trend of algae content in feeding trials: ** p < 0.01.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers applied in this research.
| Gene | Primer | Primer Sequences (5′–3′) | Aim |
|---|---|---|---|
| Rpcl | Forward primer | GTGACTGGAGGCACTATCCG | qRT-PCR |
| ß-actin | Reverse primer | TTGTTGGTGCATGGTTCGTC | |
| Forward primer | CTCCCTTGAGAAGAGCTACGA | ||
| Rpcl-NdeI | Reverse primer | GATACCAGCAGATTCCATACCC | ORF amplification |
| Forward primer | CGCCATATGGAAGAGAAGTCGTGCTCGCCT | ||
| Rpcl-BamHI | Reverse primer | CGCGGATCCTTATTCACTTGAGTTGAGTTTCATTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, S.; Shi, P.; Feng, Q.; Qiu, X.; Xu, J.; Yan, X.; Zhou, C. A Novel C-Type Lectin and Its Potential Role in Feeding and Feed Selection in Ruditapes philippinarum. Fishes 2023, 8, 62. https://doi.org/10.3390/fishes8020062
AMA Style
Chen S, Shi P, Feng Q, Qiu X, Xu J, Yan X, Zhou C. A Novel C-Type Lectin and Its Potential Role in Feeding and Feed Selection in Ruditapes philippinarum. Fishes. 2023; 8(2):62. https://doi.org/10.3390/fishes8020062
Chicago/Turabian StyleChen, Sentao, Peng Shi, Qingkai Feng, Xiaoting Qiu, Jilin Xu, Xiaojun Yan, and Chengxu Zhou. 2023. "A Novel C-Type Lectin and Its Potential Role in Feeding and Feed Selection in Ruditapes philippinarum" Fishes 8, no. 2: 62. https://doi.org/10.3390/fishes8020062