
Main Components of Fish Immunity: An Overview of the Fish Immune System
by
 , , , and
, , , and
Doaa M. Mokhtar
1,2,
Giacomo Zaccone
3 ,
,
Alessio Alesci
4,
Michal Kuciel
5,*,
Manal T. Hussein
1 and
Ramy K. A. Sayed
6
1
Department of Cell and Tissues, Faculty of Veterinary Medicine, Assuit University, Assiut 71526, Egypt
2
Department of Histology and Anatomy, School of Veterinary Medicine, Badr University in Assiut, Assiut 71526, Egypt
3
Department of Veterinary Sciences, Polo Universitario dell’Annunziata, University of Messina, 98168 Messina, Italy
4
Department of Chemical, Biological, Pharmaceutical, and Environmental Sciences, University of Messina, 98166 Messina, Italy
5
Laboratory of Toxicological Information and Laboratory Analysis, Department of Toxicology and Environmental Disease, Faculty of Medicine, Jagiellonian University, 31-008 Kraków, Poland
6
Department of Anatomy and Embryology, Faculty of Veterinary Medicine, Sohag University, Sohag 82524, Egypt
*
Author to whom correspondence should be addressed.
Fishes 2023, 8(2), 93; https://doi.org/10.3390/fishes8020093
Submission received: 17 January 2023
/
Revised: 28 January 2023
/
Accepted: 30 January 2023
/
Published: 5 February 2023
{kind=link}
Abstract
:Cellular immune responses consist of innate and adaptive cell-mediated immune mechanisms, where all leukocyte subpopulations are included. Among these are vital processes such as cell-mediated cytotoxicity and phagocytosis. The main cellular constituents of the fish immune system are macrophages, granulocytes, dendritic cells, NK cells, and cytotoxic T cells. This review provides the latest information on cellular defense mechanisms of fish and provides an overview of the function of the mucosal immune system in maintaining the general health of fish. Here, we discuss the fundamental ideas that underpin mucosal immune responses in teleosts, as well as the innate and adaptive immune cells and the molecules that play a role in these immune responses. Moreover, cytokine molecules and pathways in teleosts have been reported to focus on several kinds of associated immunity. Importantly, we also review antigen processing and presentation. The knowledge reported here will enable better understanding, determination, and modulation of the pathways responsible for protective immunity, thus consequently improving the health of the fish in aquaculture.
1. Introduction
The innate immune system and the adaptive immune system are the two components of the vertebrate immune system. Initial infections and diseases are first addressed by the innate immune system. Innate immune system components include physical barriers, cellular processes, and humoral components [1]. With the persistence of the pathogen, the adaptive immune system entices the pathogen with specificity and memory. The adaptive immune system is highly specific to a specific antigen and can provide long-standing immunity [2]. Several ingredients of the adaptive system, including T cell receptors (TCR), immunoglobulins (Igs), and major histocompatibility complex (MHC), are presumed to have arisen in the first jawed vertebrates [3].
Physiologically, the immune systems of fish and higher vertebrates are similar. In contrast to higher vertebrates, fish are mostly dependent on non-specific immune systems during their survival in the environment from the early embryonic stage. The innate immunity acts by identifying pathogen-associated molecular patterns of pathogenic microorganisms, while the acquired immunity acts through antigen-specific receptors [4]. In the teleost immune system, antigens are handled and presented by a wide variety of accessory or antigen-presenting cells (APC), including macrophages or monocytes, to specific lymphocytes in a seemingly alloantigen-restricted fashion [5].
Jawed vertebrate species are distinguished by their adaptive immune system, which is based on B and T cells, along with high degrees of diversity and specificity of their antigen receptors, the immunoglobulins (IG) or antibodies and the T cell receptors (TCRs), respectively [6]. Teleost antibodies are found in the skin, intestine, bile, gill mucus, and systemically in the plasma [7,8,9,10]. Furthermore, specific antibodies can be generated in these regions without generating a systemic response [9,11,12].
Previous studies and reviews have reported the existence of all major cytokine families in fish [13,14,15,16]. IL-1β and TNF-α are cytokines implicated in inflammatory response induction in fish. Moreover, IL-6 has been reported to be involved in the cascade that induces an inflammatory response [17]. Furthermore, cytokines involved in leukocyte differentiation have been identified in fish [18,19]. This review provides a background to the immune system of fish, describes the main parameters of the innate and adaptive immune systems, and highlights recent knowledge on the immune components in teleost fish.
2. Innate Immune System
Physiologically, the fish and higher vertebrate immune systems are similar, with certain differences, as fish are free-living organisms that depend on the innate immune system for their survival from the early embryonic stages of life [20]. The immune system of the fish is constituted by two immune mechanisms: innate (non-specific) and adaptive (specific). The components of the innate immunity respond to pathogens via identifying pathogen-associated molecular patterns (PAMPs) that demonstrate no expression in host cells. Among these PAMPs recognized by the innate immune receptors are lipopolysaccharides (LPS), lipoteichoic acid (LTA), phospholipomannan, beta-glucan, chitin, and hemagglutinin [21,22,23]. Following pathogen recognition, the pathogens are destructed by phagocytosis, one of the effector mechanisms of innate immunity. However, adaptive immunity recognizes pathogens through highly specific receptors, and consequently induces specific B and T cell clone proliferation and differentiation [24].
In fish, the innate immunity is the primary defense line that shows reactions to pathogens within a very short time and does not provide long-standing protection [25]. In contrast to mammals, the fish innate immunity is an essential component in the prohibition of pathogen entry because of the inefficiency of the adaptive immune response [26]. Most of the primary and secondary lymphoid organs described in mammals are detected in fish, with the exception of the bone marrow and lymph nodes, which lack the organs’ complexity in fish compared to mammals. This results in a potential limitation and delays the generation of fully functional adaptive immune responses [27].
The immune system of teleost fish is constituted by two receptor categories: the antigen-specific receptors and germline-encoded pattern recognition receptors (PRRs). Moreover, numerous other receptors or molecules can participate in the innate immunity. The innate mechanisms can be categorized into constitutive and inducible. The constitutive mechanism confers a quick response and rapid ongoing ligand binding to receptors, whereas the inducible mechanism acts slower, with a higher magnitude [28].
The innate immune system components in fish are generally categorized into three compartments: physical or surface, humoral, and cellular factors [29,30]. The physical or surface factors include scales, the mucus layer, and epithelial cells lining the gills, skin, and alimentary tract, providing a fundamental role in resisting infection [31,32,33]. Moreover, the epithelial cells of the epidermis prevent the entry of foreign materials, and are also populated with macrophages, eosinophilic granular cells, and lymphocytes [33,34]. The integrity of these cells plays a pivotal role in preserving the osmotic balance and eliminating microorganisms [35].
Humoral immune parameters are an array of soluble substances, which have pre-emptive functions through the inhibition of the microorganism’s growth and neutralization of the enzymes on which the pathogen depends. Teleost fish contain many non-specific defense substances, including transferrin, lectins, lysozyme, antimicrobial peptides, cytokines, natural antibodies, and complement components, which can destroy microorganisms or suppress their growth [29,36]. Transferrin is an essential element for the growth of microorganisms and is an acute phase protein invoked during the inflammatory response process for removing iron from deteriorated tissue [37]. It is also an activator of fish macrophages [38].
A wide variety of lytic enzymes may cause the lysis of pathogenic cells. These enzymes are either single or in combination. In fish, lysins include complement, lysozyme, and antimicrobial peptides. Lysozyme works on the peptidoglycan layer of bacterial cellular walls, causing bacterial lysis [39]. Lysozymes are synthesized in the liver and extra hepatic sites, and are found in mucus, plasma, and lymphoid tissue. They are involved in a comprehensive defense mechanism, such as bacteriolysis, in addition to restricted antiviral and antineoplastic activities [40]. Antimicrobial peptides play an essential role in host defense against viruses and bacteria. They are found in the mucus, liver tissue, and gills of teleosts [41,42]. The complement system plays a major role in the connection between both innate and adaptive immune responses, which permits an incorporated host defense to pathogenic challenges [43]. This system serves diverse functions including mediation of inflammatory vasodilation, lysis of bacterial and infected cells, phagocytosis of foreign particles, and clearance of apoptotic cells. In many fish, the bactericidal activity of the complement system has been reported [44].
The cellular components of the fish’s innate immune system include a wide diversity of cells. These immune cells are activated when they recognize a pathogen via its PAMP, and, consequently, these cells can share in diverse responses, relying on their cell subtype [27].
3. Antigen Processing and Presentation
Antigen processing and presentation (APP) is defined as the process through which an antigenic protein is processed into peptides. These peptides are loaded and transferred to the cell surface on major histocompatibility complex (MHC) proteins [45]. Histocompatibility molecules are glycoprotein receptors that are expressed in all cells of an organism. They are encoded by a gene complex. MHC plays a vital role in exogenous and endogenous antigens [46]. The adaptive immunity emergence is connected to the extremely polymorphic MHC genes encoding MHC, which are linked and scattered throughout a chromosomal genomic region in most jawed vertebrates, with varying levels of recombination. In teleost fish, the MHC class I gene family exists on several chromosomes; however, the MHC class II genes are missing in some fish species [47,48]. There is adequate evidence to confirm the similar functions of MHC class I and II molecules in fish and mammals [49]. The APP occurrence in teleost fish has been demonstrated through studies with the channel catfish (Ictalurus punctatus), with evidence revealing that MHC-like proteins govern the similar restricted APP to that observed in mammals [50], and with exogenous antigens being endocytosed by antigen-presenting cells (APCs) and inducing the proliferation of lymphocytes [51].
All cells that express MHC class I and can implement APP are described as APC. The spleen and kidney are hematopoietic organs capable of prompting and exerting innate immune responses [30,52]. In teleost, the spleen serves as a major secondary immune organ, and has a prime role in the antigen presentation and initiation of the adaptive immune response [26]. Furthermore, the anterior kidney is the main immune organ accountable for antigen processing, phagocytosis, IgM, and immune memory formation through melanomacrophage centers [53,54,55].
MHC class I molecule expression has been reported in teleost fish [56], and there is evidence indicating that the activation of CD8 + T lymphocyte-mediated cytotoxicity is MHC class I restricted [57], where MHC I molecules can reveal a cross-presentation process, through which these molecules can recognize and display exogenous antigens to cytotoxic T cells [58]. MHC class II is expressed only in the professional antigen-presenting cells (pAPCs), which mainly populate lymphoid organs including the head kidney, spleen, or thymus, and are capable of priming T lymphocytes to become cytotoxic CD8 + T lymphocytes or CD4 + T helper lymphocytes. Furthermore, the pAPCs express MHC class II molecules that normally engulf pathogens, and display peptides processed from exogenous antigens via lysosomal pathways [59]. In some fish species, the pAPCs are B cells, dendritic cells (DCs), monocytes/macrophages, granulocytes, and erythrocytes [60,61,62].
4. Cellular Immune Components
The immune cell populations present in fish are not very different from those found in mammals. Immune cells in teleosts include natural killer cells, non-specific cytotoxic cells, macrophages, granular leucocytes, thrombocytes, monocytes, dendritic cells, lymphocytes, mast cells, and eosinophilic granule cells. In addition, fish also possess rodlet cells and melanomacrophage centers.
4.1. Natural Killer (NK) Cells
In teleost fish, there are two different types of NK cell homologs: non-specific cytotoxic cells (NCCs) and NK-like cells [63]. In contrast to NK cells, which have a large and granular morphology, NCCs are tiny, agranular cells that resemble catfish monocytes. [64]. NCCs in seabream are highly variable in morphology [65]. The discovery of NK-like cells in teleosts suggests that NCCs are not the same as NK cells in fish. Accordingly, NK-like cells destroy allogeneic and virus-infected cells, according to a few investigations conducted on rainbow trout and catfish [63]. From teleost NK-like cells, the gene that acts as a cell marker, NK cell enhancement factor (NKEF), has been discovered. After viral and bacterial infection, NKEF gene expression is elevated in tissues such as the skin, gills, and other organs. [66]. Interestingly, a recombinant NKEF protein enhances the cytotoxicity of NCC from the kidney in Nile tilapia [66].
The NCCs were the first recognized and are the most thoroughly investigated killer cell population in teleosts. The NCCs serve functions like those of the higher vertebrates, acting on various target cells, including virus-infected cells, tumor cells, and protozoan parasites. NCCs may also be involved in antibacterial immunity by triggering the production and secretion of cytokines [67]. The NCCs of tilapia and catfish express components of the granule exocytosis pathway of cell-mediated cytotoxicity (CMC) analogous to cytotoxic lymphocytes of mammals [68]. Non-specific CMC reactions in mammals are mainly performed by NK cells. Non-specific cytotoxic cells (NCCs) and NK-like cells, two classes of NK cell homologues, are responsible for non-specific CMC mechanisms in fish. Fish NK-like cells have been isolated from blood leukocytes and have been shown to kill virus-infected, allogeneic, and xenogeneic target cells on their own [34]. NCCs, on the other hand, tend to target a variety of cells, including some protozoa and tumor cells, and are particularly active in the spleen and head kidney. NCCs are capable of spontaneously killing the affected cells through necrotic and apoptotic mechanisms [27].
These cells show variable morphological features, varying from tiny agranular monocyte-like cells in catfish to a mixture of acidophilic granulocytes, monocyte-macrophages, and lymphocytes in seabream [69]. Moreover, a population of circulating lymphocytes that resemble mammalian natural killer (NK) cells in terms of morphology and functionality has also been reported [70]. Additionally, NCCs express the NCC receptor protein 1 (NCCRP-1) on their cell surface and present a vimentin-like surface molecule [71]. It has been shown that catfish NCCs recognize and kill various human cell lines [64]. Further research reported that granulysin, perforin, and serglycin, gene-encoding molecules with lytic capacity, are expressed by NCCs [72]. Finally, the fish possess different NCC subsets in several of their immune compartments [73].
4.2. Macrophages
Macrophages have a pivotal role in specific immune responses because of their function in lymphocyte activation and phagocytosis. Macrophages possess specific receptors capable of recognizing β-glucan, so that the immunostimulants augment leukocytes’ respiratory burst, which produces reactive oxygen species with bactericidal activity [74]. Another bactericidal mechanism is represented by the production of nitric oxide (NO), which is catalyzed by an NO synthase. Using enzyme histochemical techniques, Schoor and Plumb [75] demonstrated inducible NO production from the head kidney of channel catfish (Ictalurus punctatus) infected with Edwardsiella ictaluri. In addition, Stafford et al. [76] distinguished the molecules found in crude leukocyte supernatants that stimulate NO production in macrophages of goldfish, indicating that transferrin is an essential mediator for the activation of both fish granulocytes and macrophages. In fish, neutrophils and macrophages play an essential role in controlling the spread of infectious agents and are accountable for the destruction of phagocytosed pathogens [77].
Macrophages act as antigen-presenting cells in the distal intestine, enabling antigens to interact with the adaptive immune system for identification. Other granulocytes can also be identified in the intestinal segments for innate clearance, in addition to the resident macrophages [78]. The lamina propria and epithelial linings of the intestine contain these innate populations, allowing a close vicinity to the digested pathogens.
Macrophages express several receptors on their cell surface, including TLRs, PRRs, and CLRs, in addition to complement and scavenger receptors. Macrophages are also an essential source of chemokines and cytokines, linking innate and adaptive immunity, which mediates an efficient immune response [79]. Moreover, macrophages are important for antigen presentation to T cells [80].
Circulating monocytes are typically CD14+, and express chemokine receptors, TLRs, adhesion molecules, and surface molecules that are involved in the pathogen-associated molecular pattern recognition at sites of inflammation and/or infection [81].
4.3. Neutrophils
In the blood, peritoneal cavity, and lymphoid organs, neutrophils are polymorphonuclear cells that have the ability to phagocytose cells or foreign particles and manufacture superoxide anions, a bactericidal component [82]. Neutrophils play an essential role in the inflammatory immune response against various viral, bacterial, fungal, and protozoan pathogens [79]. Neutrophils are the first granulocytes to be detected at the injury site, followed by macrophages, both of which are directed by chemotactic factors produced by wounded tissue. At the site of injury, neutrophils use antimicrobial peptides, proteolytic enzymes, and reactive oxygen species (ROS) to phagocytose microorganisms and destroy them [83]. Additionally, fish neutrophils have the ability to release extracellular fibers known as neutrophil extracellular traps (NETs) that include DNA, histones, and proteins that can bind, kill, and inactivate viruses, bacteria, fungi, and parasites [84]. Furthermore, the granular leukocyte-derived myeloperoxidase enzyme can interact with hydrogen peroxide to create hypochlorite, which in turn can be used to produce cloramins, oxidative substances that can attack microorganism membranes [85].
4.4. Eosinophils, Basophils, Thrombocytes, and Monocytes
Eosinophils are widely distributed cells in connective tissue, particularly in the gastrointestinal tract, bloodstream, ovaries, and gills, and offer degranulation when parasite infections are present [86].
Basophils are large polymorphonuclear granular leucocytes that are rarely observed in teleost species [87]. Their cytoplasmic granules have an inflammatory mediator, histamine. These cells are involved in allergy and anaphylaxis [88].
Thrombocytes are oval-shaped, nucleated, and agranular cells. They possess both a coagulation function and phagocytic ability. They show acid phosphatase activity that causes cells to gather at the inflammatory site [89].
The monocytes show phagocytosis and non-specific cytotoxic activities. These cells are thought to be transient blood cells because they migrate to the connective tissue during the inflammatory phase and change into macrophages [89]. Professional phagocytes that can be deployed to the site of inflammation include monocytes, macrophages, granulocytes, and dendritic cells [90].
4.5. Dendritic Cells (DCs)
The DCs are among the antigen presenting cells found within several tissues and can initiate both innate and acquired immune responses [91]. DCs are characterized by their small cell bodies with many cytoplasmic processes (dendrites) [92] and have been found in gills, gut, skin, and ovaries. Cell populations with DC-like functions and morphology have been demonstrated in many fish species [93]. With the assistance of MHC class 2 receptors, these cells deliver the processed antigens to T lymphocytes, triggering the cell-mediated acquired response. These cells appear to activate T cells in an antigen-dependent manner in zebrafish by expressing genes associated with antigen presentation, such as MHC class II invariant chain iclp1, csf1r, and il12 [94]. The trout gut has been shown to include putative CD8 + DC-like cells that appear to have phagocytic activity and the capacity to excite naive T cells [95].
Recently, Alesci, et al. [96] identified the intestinal DCs in several fish species including Scyliorhinus canicula (Chondrichthyes), Eptatretus cirrhatus (myxines), Polypterus senegalus (Osteichthyes, Brachiopterygii), Clarias batrachus (Osteichthyes, Teleostei), and Lepisosteus oculatus (Osteichthyes, Holostei). These findings showed that intestinal DCs and DC-like cells were positive to TLR-2, Langerin/CD207, and MHC II.
4.6. Lymphocytes
B lymphocytes, which are professional antigen-presenting cells (APCs) produce antibodies against antigens and mediate humoral immunity. They also develop immunological memory by differentiating into memory B cells [27]. A key location for B cell production and development into mature naive B cells is the fish head kidney. Immunoglobulins are secreted into the plasma by B cells to form an antigen-binding complex [96]. These secretory cells, also known as antibody secreting cells (ASCs) or plasmablasts, resemble mammalian plasma cells [97].
When B cells come into contact with antigens, activation occurs, which can be either T cell-independent (for polysaccharides- and unmethylated CpG DNA-derived antigens) or T cell-dependent (for protein-derived antigens) [98]. Once an antigen is bound by the BCR on an antigen-presenting B cell, the antigen is processed and delivered to T helper cells by MHC class II on the surface of B cells. T helper cells are then activated and differentiated as a result, and they subsequently express the surface protein cluster of differentiation 40 ligand (CD40L). When CD40L interacts with the B cell surface protein CD40, it causes naive B cells to activate and differentiate [99,100]. Furthermore, T cell activation is not necessary for T cell-independent B cell activation, which is triggered by the direct binding of an antigen to the BCR [99].
Activated B cells can either differentiate into memory B cells, which circulate throughout the body and are in charge of secondary antibody response, or into plasmablasts, which are proliferating, short-lived, and low affinity antibody-producing cells that later differentiate into plasma cells, which are not-proliferating, long-lived, or high-affinity antibody-producing cells [98]. When memory B cells come into contact with the same antigen, which activates their parent B cells since these two share the same surface BCR, a secondary response is generated [100].
In the cell-mediated adaptive immune system, T cells play an inherent role in directing immune activity after the early non-specific defenses and offer an immunological response after antigen identification by APCs. The T cell may then direct the clearance of the foreign material by phagocytosis or cytokine stimulation after pattern recognition through the TCR. As a result, specific antigen–receptor interactions and the MHC pairing stimulate the cellular responses of the two main groups of T cells, T helper cells and cytotoxic T cells [101]. T cell differentiation and control are influenced in distinct ways by surrounding immunological molecules. While cytotoxic T cells are controlled by cytokine concentrations and surrounding helper T cells. T helper cells are affected by antigen binding affinity, antigen concentrations, and signaling from APCs [102]. Moreover, the release of IL-2 acts as a growth promoter for the overall development of T cells [103]. After differentiation, helper T cells produce lymphotoxin (LT) to interact with intracellular pathogens and release IFN-γ to help in resisting viral infections [103]. The helper T cells can be further classified into Th1 and Th2 according to their functions and target pathogens. The Th1 subset offers cell-mediated removal of microbes. On the other hand, the Th2 cell tends to target particular pathogens, such as parasites [104].
In teleosts, T cells are correlated to the mammalian subset and possess complementarity-determining region 3 (CD3) binding domains within the TCR [105]. The head kidney and thymus nurse the developing T cells and act as a base for plasma circulating T cells. Salmonid CD3 was proven by Koppang et al.to be an effective T cell marker to assess the anatomical distributions of T cells and expression regions [105]. Numerous T cells have been reported to populate the thymus before migrating to the kidney and spleen after development [101]. The antigen presentation of both MHC class I and II has demonstrated the directional activation of a T cell response, in addition to T cell characterization. MHC class I interacts with CD8 + cytotoxic T cells, whereas MHC class II helps with complex formation in T helper cells [101].
4.7. Mast Cells
In most teleost species, mast cells are a constituent of innate immunity and are located near the blood vessels in the ovaries, skin, intestine, and gills [106]. The mast cell granules express S100 protein, desmin, and CD117. Moreover, their granules are stained positively with bromophenol blue and Safranin O. Using electron microscopy, these cells are characterized by the presence of numerous cytoplasmic electron-dense granules [86].
Several stress conditions, such as exposure to herbicide and heavy metals, chronic inflammation, and parasitic infections, cause an increased number of mast cells in the organs and tissues of teleosts [107]. Teleost fish have also demonstrated mast cell degranulation, cytokine release, and the ensuing inflammatory response after bacterial infection [108].
Piscidins, host-defense peptides, were initially discovered in hybrid striped bass. It is interesting to note that piscidins are found in mast cells, highly prevalent granulocytes that are present in all vertebrate classes [109,110]. The recent localization of ACh in mast cells and the pavement cells, as well as the interaction of the mast cells with eosinophils and the innervation of the latter by nicotinic acetylcholine receptor (nAChR) alpha 2 subunits, were correlated with the modulation of the immune function in fish gill [111]. Both muscarinic and nicotinic cholinergic receptors have been identified on mast cells [112].
4.8. Eosinophilic Granule Cells (EGCs)
Salmonids and cyprinids are among many species that frequently possess EGCs. EGCs are typically present in the skin’s epidermis, gills, connective tissue surrounding the spinal cord, and intestinal mucosa [113]. In the connective tissue stroma surrounding the bile duct, EGCs can also be identified [114]. They have spherical granules in their cytoplasm that are brightly stained red by hematoxylin and eosin (HE) because they contain basic proteins. Additionally, their granules are stained deep magenta with PAS and alcian blue, due to their content of neutral and sulfated glycosaminoglycans. They exhibit a positive reaction to bromophenol blue, which shows that they contain protein, and they show a metachromatic affinity to toluidine blue.
Many functions are reported for these cells, including the release of oxygen radicals and toxic proteins onto the surface of multicellular parasites in inflammatory areas. Additionally, acute tissue damage results in EGC degranulation and release of inflammatory mediators, but chronically inflamed tissues frequently exhibit an increase in the number of these cells [115].
4.9. Rodlet Cells
Fish only have rodlet cells, which are distinguished by their thicker capsule-like cell borders and rodlet cytoplasmic inclusions [116]. The rodlet cells of the grass carp head kidney expresses α- SMA [117]. Rodlet cells possess an actin cytoskeleton, which may contribute to the contraction that ejects their contents. During the expulsion phase, they go through a one-way contraction that eventually destroys the cell; the rodlets and all the contents of the cell are evacuated [118]. The rodlet cells are frequently distributed in the heart, thymus, kidney, spleen, gills, skin, gall bladder, liver, blood vessel endothelium, and pancreas [106].
Several functions have been reported for these cells, including a secretory function [119], which is a defensive role, especially against parasitic helminths [79]. They may play a role in water or electrolyte transport or lubrication, pH control, and antibiotic reactions. They may be modified goblet cells or may play a regulatory role in osmoregulation and transportation of ions. They can also be viewed as non-specific immune cells that are implicated in immunity, as they frequently occur in parasitic infestation. Rodlet cells, on the other hand, may be triggered because of tissue damage, simulating leukocyte reactions to various chemotactic stimuli [120]. Moreover, rodlet cells can be regarded as a type of eosinophilic granule cell or mast cell. The nature of their contents must be determined to throw additional light on the functions of RCs. The molecular structure of RCs has been investigated using lectin histochemistry and immunohistochemistry to clearly distinguish rodlet cells from mucous cells, macrophages, and granulocytes [121]. Complex carbohydrates were detected in the RCs of fish, primarily glucosidic residues of N-acetyl-galactosamine and, less commonly, N-acetyl-glucosamine and sialic acid. Sialic acid residues in the carbohydrate backbone help the mucosae to defend themselves against infections [122].
5. Mucosal Barriers
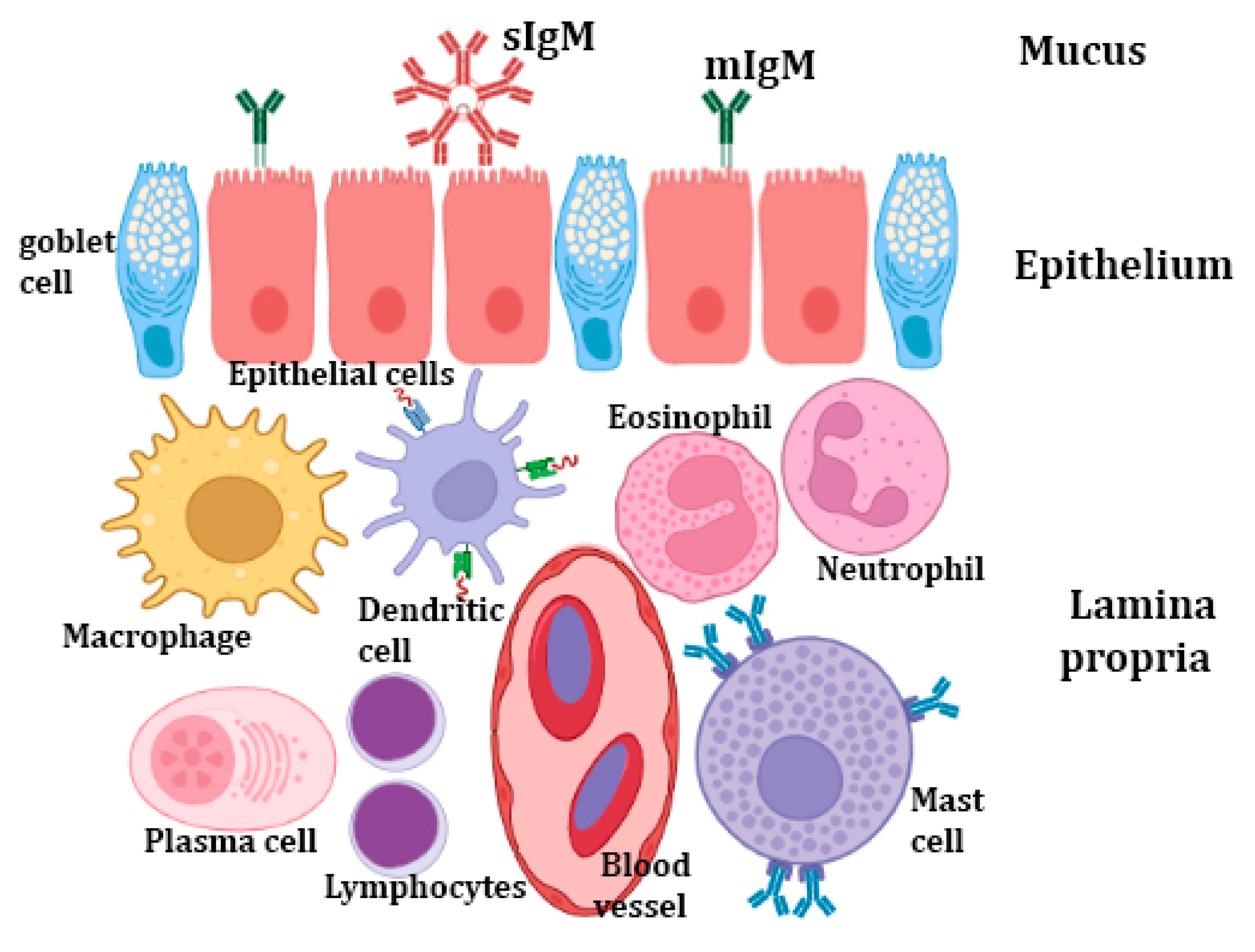
Fish can interact with their surroundings through mucosal barriers while still maintaining homeostasis. Mucosal barriers (Figure 1) are extremely complex tissues with special adaptations that enhance a variety of physiological functions, including immunity, gas exchange, and nutrition. The principal mucosa-associated lymphoid tissues (MALTs) of teleosts are the gut-associated lymphoid tissue (GALT), gill-associated lymphoid tissue (GIALT), skin-associated lymphoid tissue (SALT),) and nasopharynx-associated lymphoid tissue (NALT). These component mucosal barriers serve as the host’s local immune system and protect it from pathogens and other threats. The fish MALT possesses the major populations of innate immune cells, as well as the lymphocyte populations of CD4 and CD8 T cells, and B cells [3]. However, only three immunoglobulins’ (Igs) isotypes have been discovered in teleost to date (IgM, IgD, and IgT/Z), each of which consists of two identical heavy and two identical light polypeptide chains [123]. IgT/Z is thought to be specialized in the mucosal immunity. In mammalian mucosal surfaces, the main immunoglobulin is IgA, which is mostly produced by plasma cells present in the gut lamina propria. In a similar way, the teleost IgA homologue, IgT/IgZ, has a dominant role in gut mucosal immunity [124]. The transport of IgM and IgT through mucosal barriers is carried out by the polymeric immunoglobulin receptor (pIgR), which is also expressed in the gut and skin of teleosts [125]. In zebrafish and rainbow trout, dendritic-like cells have recently been identified [94]. It is intriguing that DCs only make up around 15% of the mononuclear phagocytes in the skin of adult zebrafish but are sparse in the gut, spleen, and kidney [126]. It has been hypothesized that zebrafish skin has DCs comparable to mammalian Langerhans cells [127]. Additionally, innate immune genes such as TNF have been found in the mucosal tissues, indicating that innate immunity has a role in the antiviral process [128].
5.1. Fish Mucus
Fish mucus that covers the mucosal surface is produced by many types of secretory cells, mainly the goblet cells (GCs), sacciform cells, and club cells. Goblet cells, which produce mucus granules and contain glycoproteins, are prevalent on all external surfaces, as well as the surface of the gills [129]. The initial line of defense against pathogen invasion is fish mucus [130]. Mucins are the major molecules found in mucus and are responsible for the mucus viscosity, pathogen entrapment, physical protection of the skin’s surface, and signaling at the cell surface. Mucins are a heterogeneous family of high molecular weight glycoproteins with single or many protein domains that have numerous locations where O-glycan attachment can be formed. Mucins contribute to the innate immune system in two ways that are significant. First, they prevent the adhesion of pathogens, sustained colonization of potentially infectious bacteria, and invasion of parasites by being continuously generated and routinely shed [131]. Second, they contain a variety of innate immunity-related proteins and enzymes, including lysozyme, esterases, complement proteins, lectins, C-reactive protein (CRP), transferring, alkaline phosphatise (ALP), proteases, immunoglobulins, and antimicrobial peptides (AMPs), and various other antibacterial proteins and peptides, which are frequently responsible for deteriorating, inactivating, and controlling infections [132,133]. In addition, the fish skin mucus has been reported to have few metabolites with antibacterial properties such as N-acetylneuraminic acid, azelaic acid, and hydroxyisocaproic acid [134].
According to reports, fish skin mucus contains a variety of saturated fatty acids (SFAs), monounsaturated fatty acids (MUFAs), and polyunsaturated fatty acids (PUFAs). Palmitic acid and stearic acid are SFAs that are present in mucus. Oleic acid is an example of a monosaturated fatty acid. Moroctic and alpha-linolenic acids are polyunsaturated fatty acids. These fatty acids are thought to perform a significant defensive function against pathogens [135]. Skin transcriptome analysis of S. trutta, M. anguillicaudatus, and C. striatus demonstrated that several genes are implicated in immunology and epidermal mucus secretion. Mucins are divided into two structurally separate families: membrane-bound and large secreted gel forming (SGFM) forms, and, to date, more than 20 mucin genes have been identified in higher vertebrates [136]. Cystein-rich (C8) and trypsin inhibitor-like cysteine-rich (TIL) domains are two of the domain structures that distinguish the secreted gel-forming mucins from other types. These domains aid in the disulfide bond formation that leads to the oligomerization of the mucin proteins, which gives mucus its ability to gel. Additionally, long repetitive sequences rich in proline, threonine, and serine residues, known as the PTS-domain, are present in the released gel-forming mucins [137,138]. MUC2, MUC5AC, MUC5B, MUC6, and MUC19 are examples of SGFM. However, the membrane-bound mucins include MUC1, MUC3, MUC4, MUC12, MUC13, MUC14, MUC15, MUC16, MUC17, and MUC18 [139]. The mucin genes of a few teleost fish species, such as zebrafish (Danio rerio) [140], gilthead sea bream (Sparus aurata) [141], common carp (Cyprinus carpio) [142], and sea trout (Salmo trutta), have been discovered [143]. Seven secreted gel-forming mucin genes were discovered in the Atlantic salmon based on many levels of evidence, including annotation, transcription, phylogeny, and domain structure. Five genes were labelled as muc5 and two as muc2. The muc5 genes were largely expressed on the epidermis and gills, while the muc2 genes were primarily expressed in the intestinal region [144]. The mucus cells and the composition of the mucus are influenced by several exogenous (e.g., hyperosmolarity, stress, infections, and pH) and endogenous (e.g., developmental stage, sex) factors [31].
The properties of the mucus vary among fish species, and they are subject to change in response to disease and environmental factors. Amoebic gill disease (AGD) is one of the main disorders affecting the mariculture of Atlantic salmon (Salmo salar L.). AGD is characterized by increased gill mucus production and hyperplasia of the lamellar epithelium. The regulation of two mucins (muc5, muc18) changed in response to illness. The secreted muc5 was significantly upregulated in AGD, but the membrane-bound muc18 showed the reverse pattern [145]. Surprisingly, the properties of the skin mucus of carp appear to alter in response to increases in the total amount of bacteria in the water. Increases in acidic glycoconjugates and overall glycosylation can be noticed as changes in the terminal presence of certain sugars [146]. Similar to this, myxozoan parasite infection alters the characteristics of seabream (Sparus aurata) gut mucus, resulting in higher levels of glycosylation and terminal glycosylation of mucus proteins in the posterior gut [147].
The fish mucus has a wide range of functions, including protection, respiration, disease resistance, as well as osmotic regulation, excretion, reproduction, communication, nest building, and feeding. Fish mucus extract is increasingly known to possess a wide range of biological qualities, for instance, antiviral, antibacterial, antiparasitic, and antifungal qualities, which may be employed in both human medicine and fish farming [148]. Sacciform cells and acidophilic granular cells (orserous goblet cells) have also been demonstrated in fish skin whose secretions mix with the secretions of goblet cells to form mucus. Sacciform cells may be analogous to granular glands of amphibians, which produce crinotoxic and repellent substances, but secretions of these cells also have a protective and regulatory role [149].
On the other hand, the adaptive immune components include Igs, which are expressed on the B cells’ surface as B cell receptors or in a soluble form in body fluids [150]. Moreover, these mucosal Igs (secretory Igs “sIgs”) are widespread in the mucosal barriers of teleosts, including the gut [124], olfactory organ [151], gills [152], skin [153], and buccal and pharyngeal cavities [154]. Recently, a bursa of Fabricius-like structure in Atlantic salmon (Salmo salar) cloaca was identified where IgT+ and IgM+ B cells could be distinguished, which indicates that sIgs may be secreted into the mucus around this structure [155].
The expressions of constituents involved in the cholinergic system, including acetylcholinesterase (ChAT) and vesicular ACh transporter (VAChT), have been confirmed in various immune cells including macrophages and epithelial cells [156]. Several studies have demonstrated that most epithelial cells do not concentrate ACh into vesicles; instead, they release it directly from the cytoplasm [157].
5.2. The Intestinal Mucosa
The intestinal mucosa of teleosts is a saturated substrate for leukocytes, and they are housed within the lamina propria [78]. These strata contain proliferating granulocytes, lysozyme, and NK cells, which together promote the production of lysozyme and superoxides [158,159]. The epithelial cells also play the role of a secondary barrier, securing the passage while permitting proper nutrient flows, signaling permeability, and benefiting gut microflora to improve digestion [160].
The intestinal regions possess an essential population of immunoglobulins and isolated populations of both T and B cells, called intra-epithelial lymphocytes (IELs) [161]. Dezfuli et al. [162] have found that mast-cell analogues have a role in the indirect propagation of immunoglobulins following viral infection. Inami et al. [163] found a larger number of IgM-positive cells in the rectum of Atlantic cod (Gadus morhua), compared with the stomach regions and foregut. This result agrees with the familiar adaptive immunocompetency of the posterior intestine and demonstrates the capability of Igs to aggregate in specific regions away from common GALT. As fish lack IgA, serum or secretory immunoglobulin, IgM is rapidly degraded within the harsh environment [164].
6. Immunoglobulins
In bony fish and other jawed vertebrates, the adaptive immune system relies on B and T cells and on the tremendous variety and specificity of their antigen receptors, the immunoglobulins (IG) or antibodies, and the T cell receptors, respectively [165,166]. Immunoglobulins are the main constituents of the immune response against pathogens [30]. Among the mechanisms implying humoral immunity mediated by IGs are pathogen elimination via phagocytosis, toxin and virus neutralization, and complement cascade activation [167,168].
In bony fish, three isotypes of IGs are produced by B cells: IgD, IgM, and IgT/Z. Therefore, three major B cell lineages have been characterized in teleosts [169]. The predominant and most ancient immunoglobulin in fish is the IgM class tetramer, which contains eight antigenic combining sites [170]. Although IgM can be expressed at the surface of B cells, secreted tetrameric IgM is considered the most predominant IG in fish serum [166,171]. Many studies have reported that mucosal and serum noncovalently accompanies IgM monomers in diverse fish species [172,173].
In rainbow trout, a new teleost fish Ig was detected and was named IgT. This new IG was identified as IgZ in zebrafish [174]. This IgT has been distinguished in most studied teleost fish, except for medaka, channel catfish, and turquoise killifish [175,176,177]. In rainbow trout, at least three subclasses of IgT are expressed. Mucosal and systemic lymphoid tissues have been reported to express the IgT1 subclass, while IgT2 has been documented to be mainly expressed in systemic lymphoid organs. Furthermore, IgT3 protein is reported in rainbow trout serum [178].
IgD is the second immunoglobulin isotype characterized in fish, specifically channel catfish. As a result of its sequence similarity with mammalian IgD, its immediate location under the IgM gene, and its expression on B cells, IgD has been suggested to be as old as IgM [168,179,180]. The secreted form of IgD has been interestingly identified in trout and catfish, with a different structure [175,181].
7. Cytokines
Cytokines play an essential role in the immune system through their specific receptors binding to the cellular membrane, providing a cascade-enhancing induction, stimulation, or suppression of nuclear cytokine-regulated genes. Several critical cytokines have been recognized in teleost fish, including tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), transforming growth factor-β (TGF-β), interferon (IFN), and many chemokines [182,183,184,185]. Recently, numerous other cytokines have been identified in teleosts [186,187,188,189].
The interferons (IFNs) are the key cytokines present in all jawed vertebrates and are involved in antiviral immunity [190]. In vertebrate host cells, INFs play a critical role in the defense mechanism against viral infection through INFα/β secretion upon the recognition of viral nucleic acid [191]. These INFs keep other cells away from viral infection via binding to different receptors, resulting in the induction of multiple genes, where some of these genes encode antiviral proteins [192,193]. The IFNs have been categorized into three broad groups, where type I and III are originally embroiled in antiviral defense, while type II IFN has a broader function and shares in cell-mediated immunity in response to a wide variety of pathogens during adaptive immune responses [190].
IL-1β and TNFα are considered the proximal cytokines that are first released and boost response through enhancing the secretion of distal cytokines such as IL-6 and IL-8. All these cytokines have been detected in fish, where IL-1β, IL-6, and TNFα, with their family members, have been reported in several fish studies investigating inflammatory responses against infection [194].
IL-1β was one of the first characterized cytokines in teleosts [183,195,196,197]. As reported in zebrafish, IL-1β is produced as a precursor molecule that is cleaved to release a bioactive mature protein, which is linked to the activation of the inflammasome complex that is accountable for the secretion of IL-1β [198]. IL-1β is produced by various cell types following the stimulation of host pattern recognition receptors through PAMPs or DAMPs [199,200,201]. IL-1β improves vaccine efficacy as it boosts antibody production when administered with bacterial vaccines [202,203]. In primary leucocytes and macrophages, IL-1β action has been extensively analyzed, where induced expression of pro-inflammatory genes such as TNF-α, IL-1β, and IL-6 is evident [204,205,206]. Furthermore, IL-1β activates the expression of genes that are considered to be immune response suppressors [207]. IL-1β genes have revealed wide variations among fish species. In carp, two IL-1β genes were found: IL-β1 and IL-1β2 [208]. Three functional IL-1β genes (IL-1β1–3), in addition to pseudogene (IL-1β4), have been identified in salmonids [204], while only a single type II IL-1β was detected in gadoids [209].
IL-18 is an essential cytokine responsible for the induction of IFN-γ production and promotion of TH1 immunity in vertebrates. It exerts an essential role in regulating inflammation within mucosal tissues [210]. IL-18 has been described in a few fish species including elephant shark, pufferfish, rainbow trout, and seabream [211,212,213,214]. IL-18 is fundamentally expressed in fish immune or non-immune tissues, unlike IL-1β, which is induced by inflammatory stimuli [214]. A recent study has suggested that IL-18 is likely developed in an evolutionary-independent event to IL-1β [215].
TNFα is a member of the TNF superfamily, which primarily comprises type II transmembrane proteins, which can be cleaved to release a soluble cytokine. TNF-α presence has been reported in teleosts [216,217,218]. In carp, many TNF-α copies have been detected [219]. In catfish cell lines, TNF-α expression has been observed in T cells and macrophages; however, there are no expressions in B cells or fibroblasts [220]. In salmonids, four copies of TNF-α have been reported [205,221]. Several fish studies have suggested that TNF-α and -β are essential macrophage activators. Previous studies in some fish species, including goldfish, rainbow trout, sea bream, turbot, and catfish, have revealed that TNF induces macrophage activation, resulting in increased respiratory activity, phagocytosis, and nitric oxide production [222,223].
IL-6 was first discovered in teleost fish [187,224]. It is a member of the IL-6 family, and is considered a pivotal acute phase response cytokine. Moreover, it is implicated in the differentiation of B cells into antibody-secreting cells [225]. It is highly expressed in the kidney and spleen, and during caudal fin regeneration following amputation [226]. IL-6 has been used as a vaccine adjuvant, to achieve higher serum antibody levels for protection [227]. In salmonids, two IL-6 genes have been detected (IL-6A and IL-6B) [227,228]. IL-11 is one of the other members of the IL-6 cytokine family related to inflammatory responses. It was first detected in teleosts in the bacterial-challenged fish [229]. Two IL-11 types (IL-11a, IL-11b) have been reported in teleosts, where IL-11a is highly expressed in the intestine and gills of trout [229,230].
TGF-β is one of the TGF-β protein family. It plays a fundamental role in various functions, including cell proliferation, differentiation, and regulation. In fish, three TGF-β proteins have been detected (TGF-β1, TGF-β2, TGF-β3) [231]. TGF-β1 is the main immune active isoform, and two TGF-β1 paralogues (TGF-β1a and TGF-β1b) have been found in zebrafish [232]. In carp, TGF-β1 is highly expressed in the spleen [233], and it can induce expressions of pro inflammatory cytokine and IL-10 [234]. In rainbow trout, three paralogues of TGF-β1 have been reported [235], with expression highest in lymphoid tissues. The highest levels of TGF-β1 expression have been found in macrophages and the gills of seabream [236].
8. Immune Cells—Nerve Interaction
To monitor infections and inflammation and to take appropriate action, the neurological and immune systems work in concert as an integrated physiological system. The nervous system, including the brain and peripheral divisions, can stimulate or inhibit a variety of immune system functions, including those of the innate and adaptive immune systems. It has been established that the anti-bacterial peptides (AMPs), which are signaling molecules involved in the antimicrobial immunity, are synthesized by the enzymatic processing of neuropeptide precursors such as proenkephalin A (PEA) and chromogranin B [237,238]. It has been shown that some peptides involved in neuronal or neuroendocrine signaling also have potent antibacterial activities. Additionally, they are widely distributed across immunological, endocrine, neuroendocrine, and neuronal cells [239]. The microbiota–gut–brain axis (MGB axis) is based on the reciprocal interactions between the CNS and gut microbiota that involve endocrine, immunological, and neurological pathways [240]. In the MGB axis, lymphocytes stimulate the gut lumen and internally release cytokines, which are then activated by gut peptides released by enteroendocrine cells from sensory nerve terminals, such as those of the vagus nerve. To create the link between the immune system and the central nervous system (CNS), which results in tissue homeostasis, communication pathways based on neurotransmitters, neuropeptides, cytokines, hormones, and growth factors are required [241]. The innervation of lymphoid tissue has been investigated in the Coho salmon (Oncorhynchus kisutch) spleen, where nerve fibers are accompanied by the vascular supply and MMCs. Additionally, adrenergic receptors are among the neurotransmitter receptors that are expressed by immune cells. Adrenergic receptors have so far been sequenced in the leukocytes of zebrafish, trout, catfish, and goldfish. [242]. The activation of the sympathetic nervous system can modulate the production and circulation of leucocytes, through activation of hematopoietic stem cells and NK cells, upregulation of myelopoiesis, and production of monocytes and neutrophils by the spleen [243]. Gut neuropeptides, such as gut antimicrobial neuropeptides, constitute a complex network between the neurological and immunological systems, where they play a crucial modulatory role [244]. In response to various stimuli, both enteroendocrine cells and enteric neurons release gut neuropeptides. Most neuropeptides created by neurons during an immune response have neuroendocrine properties that can affect both the stomach and the brain [245]. The role of the mast cells, mucous cells, and neuroepithelial endocrine cells (NECs) of teleosts’ gills in innate and adaptative immunity has been examined in several studies. These substances, such as immunoglobulins (Igs), antibody-secreting cells, and piscidins, are produced by these molecules or cells through immune-related pathways [246,247]. Piscidin 1, an antimicrobial peptide, and enkephalin, were found to be co-localized in fish gill NECs and the neurological system of the gill filaments, suggesting a potential role in the bidirectional relationship between endocrine, immune cells, and their interaction with the filament nervous system [111,248]. Investigations have been conducted into the possibility of tight communication between immune cells and afferent nerve terminals in the gill. The eosinophils in the gill lamellae of Boleophthalmus are immunostained with antibodies against Pis 1 and 5HT (serotonin), and they come into contact with neurons that are also immunolabeled with antibodies against acetylated tubulin [111]. Many neuroactive substances and AMPs, including 5-HT, piscidin 1, nitric oxide (NO), tyrosine hydroxylase (TH), and the nicotinic acetylcholine receptor, are found in the terminals of afferent nerves. These substances are released by mast cells and eosinophils. Additionally, it has been discovered that eosinophils and NECs include neuropeptide receptors, such as the GABA B R1 receptor, which suggests that the immune and neurological systems interact closely [249]. During lung infections, neuro-immune interactions also take place at the surface of the lung air barrier. Bacterial pathogens are killed by the neuropeptide calcitonin-gene related peptide (CGRP). In addition to sensory nerves, the pulmonary neuroepithelial endocrine cells (PNECs) also express CGRP. PNECs have been identified to be co-localized with innate lymphoid cells (ILC2) [250].
Cytokines or other mediator molecules stimulated by cytokines would be accessible to the fish brain. However, the preoptic region of the carp brain contains the cytokine receptor IL-1RI mRNA [251]. Prostaglandins, which are smaller, neuroactive, and lipophilic, can be the mediators [252]. There is evidence that immune mechanisms that signal to the brain in fish are similar to those in mammals. It has been suggested that the vagus nerve could act as a channel for cytokines to reach the brain [252]. However, because stresses can increase peripheral cytokines by stimulating pathways such as the noradrenergic pathway, these, in turn, may cause a rise in brain IL-1 via one of the immune to brain pathways. The role of cytokines in the activation of the HPI and sympatho-chromaffin (SC) axis and the central regulation of the stress reaction in fish is not well understood because there have been few studies that have demonstrated the signaling pathways from peripheral blood molecules to the brain. Adrenaline has been demonstrated to modulate cytokines in fish [253], which can cause the signaling pathways to the brain to activate. Pro-inflammatory cytokines in the brain decrease the activity of neurotransmitters such as noradrenaline, serotonin, and dopamine, which will have a substantial impact on how behavioral patterns are modulated [254]. To further integrate peripheral pro-inflammatory and antigenic signals with physiological and behavioral response, cytokines also activate a number of other signaling pathways in the brain. Under stressful conditions, all these processes will cooperate to combine immune cells’ necessary protective response with the physiological and behavioral response.
Finally, other forms of indirect control, such as those connected to the relationship between the gut and the brain, can also exist in fish. It has been demonstrated that information is transmitted between the two organs via chemical signals in both fish and mammals [255]. As a result, when the neuro-immuno-endocrine relationship is effectively initiated, other processes such as reproduction or growth will also be impacted. The Sockeye salmon (Oncorhynchus nerka) provides a clear case study, as degeneration of various glands and organs has been linked to extremely high steroid levels and the loss of immunocompetence, particularly innate responses, during the migratory period [256]. In order to gather energy that ensures the fish can spawn, cortisol and other reproductive steroids redirect energy from numerous biological processes, including immunological functions. Sex steroids have also been found to modulate the cortisol production in the internal tissues of salmonids [257].
9. Conclusions
The immune system of the fish comprises innate and adaptive immune responses. Among these, the innate immune response is essential for combating pathogens and for providing resistance to diseases because of delayed adaptive immune response. Three major parameters are responsible for the innate immune response: physical, humoral, and cellular. Furthermore, there is a complicated cytokine network in fish for the regulation and activation of their immune system, and for producing appropriate protective responses to various pathogens. The data described here will facilitate better understanding, determination, and modulation of the protective immunity pathways for improving fish health. Furthermore, this knowledge on fish immunology will facilitate the development of vaccines designed to target immune mechanisms responsible for disease protection.
Author Contributions
Conceptualization and supervision, D.M.M., G.Z. and R.K.A.S.; literature search and screening of articles, D.M.M., G.Z., A.A., M.K., M.T.H. and R.K.A.S.; validation and investigation, D.M.M., G.Z. and R.K.A.S.; data curation and formal analysis, D.M.M., A.A. and R.K.A.S.; writing—original draft preparation, D.M.M., M.T.H. and R.K.A.S.; writing—review and editing, G.Z. and M.K.; project administration, D.M.M. and G.Z.; funding acquisition, M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Riera Romo, M.; Pérez-Martínez, D.; Castillo Ferrer, C. Innate immunity in vertebrates: An overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Innate immunity. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A comparison of the innate and adaptive immune systems in cartilaginous fish, ray-finned fish, and lobe-finned fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.; Banu, H.; Prakash, A.; Tripathi, G. Immune system of fish: An evolutionary perspective. In Antimicrobial Immune Response; del Mar Ortega-Villaizan, M., Chico, V., Eds.; IntechOpen: London, UK, 2021; p. 1. Available online: https://www.intechopen.com/chapters/78026 (accessed on 17 December 2022).
- Vallejo, A.N.; Miller, N.W.; Clem, L.W. Antigen processing and presentation in teleost immune responses. Annu. Rev. Fish Dis. 1992, 2, 73–89. [Google Scholar] [CrossRef]
- Mendoza, M.; Magadán, S. Immunoglobulins in Teleost. In Principles of Fish Immunology; Springer: Berlin, Germany, 2022; pp. 229–251. [Google Scholar]
- Hatten, F.; Fredriksen, Å.; Hordvik, I.; Endresen, C. Presence of IgM in cutaneous mucus, but not in gut mucus of Atlantic salmon, Salmo salar. Serum IgM is rapidly degraded when added to gut mucus. Fish Shellfish. Immunol. 2001, 11, 257–268. [Google Scholar] [CrossRef]
- Rombout, J.W.; Blok, L.J.; Lamers, C.H.; Egberts, E. Immunization of carp (Cyprinus carpio) with a Vibrio anguillarum bacterin: Indications for a common mucosal immune system. Dev. Comp. Immunol. 1986, 10, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, J.; Ostland, V.; Byrne, P.; Ferguson, H. Detection of a distinct gill-surface antibody response following horizontal infection and bath challenge of brook trout Salvelinus fontinalis with Flavobacterium branchiophilum, the causative agent of bacterial gill disease. Dis. Aquat. Org. 1993, 16, 21–27. [Google Scholar] [CrossRef]
- Jenkins, P.; Wrathmell, A.; Harris, J.; Pulsford, A. Systemic and mucosal immune responses to enterically delivered antigen in Oreochromis mossambicus. Fish Shellfish. Immunol. 1994, 4, 255–271. [Google Scholar] [CrossRef]
- Cain, K.D.; Jones, D.R.; Raison, R.L. Characterisation of mucosal and systemic immune responses in rainbow trout (Oncorhynchus mykiss) using surface plasmon resonance. Fish Shellfish. Immunol. 2000, 10, 651–666. [Google Scholar] [CrossRef]
- Jones, D.R.; Hannan, C.M.; Russell-Jones, G.J.; Raison, R.L. Selective B cell non-responsiveness in the gut of the rainbow trout (Oncorhynchus mykiss). Aquaculture 1999, 172, 29–39. [Google Scholar] [CrossRef]
- Secombes, C.; Zou, J.; Bird, S. Cytokines of cartilaginous fish. In Immunobiology of the Shark; Smith, S., Sim, R., Flajnik, M., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 123–142. [Google Scholar]
- Secombes, C.J.; Wang, T.; Bird, S. The interleukins of fish. Dev. Comp. Immunol. 2011, 35, 1336–1345. [Google Scholar] [CrossRef]
- Secombes, C.J.; Wang, T.; Bird, S. Vertebrate cytokines and their evolution. In The Evolution of the Immune System: Conservation and Diversification; Malagoli, D., Ed.; Academic Press Inc.: Cambridge, MA, USA, 2016; pp. 87–150. [Google Scholar]
- Wang, T.; Secombes, C.J. The cytokine networks of adaptive immunity in fish. Fish Shellfish. Immunol. 2013, 35, 1703–1718. [Google Scholar] [CrossRef] [PubMed]
- Savan, R.; Sakai, M. Genomics of fish cytokines. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Hanington, P.C.; Wang, T.; Secombes, C.J.; Belosevic, M. Growth factors of lower vertebrates: Characterization of goldfish (Carassius auratus L.) macrophage colony-stimulating factor-1. J. Biol. Chem. 2007, 282, 31865–31872. [Google Scholar] [CrossRef]
- Kono, T.; Bird, S.; Sonoda, K.; Savan, R.; Secombes, C.J.; Sakai, M. Characterization and expression analysis of an interleukin-7 homologue in the Japanese pufferfish, Takifugu rubripes. FEBS J. 2008, 275, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Rombout, J.; Huttenhuis, H.; Picchietti, S.; Scapigliati, G. Phylogeny and ontogeny of fish leucocytes. Fish Shellfish. Immunol. 2005, 19, 441–455. [Google Scholar] [CrossRef]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef]
- Silva-Gomesa, S.; Decouta, A.; Nigoua, J. Pathogen-associated molecular patterns (PAMPs). In Encyclopedia of Inflammatory Diseases; Parnham, M.E., Ed.; Springer: Basel, Switzerland, 2015; pp. 1–16. [Google Scholar]
- Taghavi, M.; Khosravi, A.; Mortaz, E.; Nikaein, D.; Athari, S.S. Role of pathogen-associated molecular patterns (PAMPS) in immune responses to fungal infections. Eur. J. Pharmacol. 2017, 808, 8–13. [Google Scholar] [CrossRef]
- Bonilla, F.A.; Oettgen, H.C. Adaptive immunity. J. Allergy Clin. Immunol. 2010, 125, S33–S40. [Google Scholar] [CrossRef]
- Turvey, S.E.; Broide, D.H. Innate immunity. J. Allergy Clin. Immunol. 2010, 125, S24–S32. [Google Scholar] [CrossRef]
- Whyte, S.K. The innate immune response of finfish—A review of current knowledge. Fish Shellfish. Immunol. 2007, 23, 1127–1151. [Google Scholar] [CrossRef]
- Firdaus-Nawi, M.; Zamri-Saad, M. Major Components of Fish Immunity: A Review. Pertanika J. Trop. Agric. Sci. 2016, 39, 393–420. [Google Scholar]
- Paludan, S.R.; Pradeu, T.; Masters, S.L.; Mogensen, T.H. Constitutive immune mechanisms: Mediators of host defence and immune regulation. Nat. Rev. Immunol. 2021, 21, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B. Innate immunity of fish (overview). Fish Shellfish. Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Uribe, C.; Folch, H.; Enríquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486. [Google Scholar] [CrossRef]
- Ellis, A. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Magnadottir, B. Immunological control of fish diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Ángeles Esteban, M. An overview of the immunological defenses in fish skin. Int. Sch. Res. Not. 2012, 2012, 29. [Google Scholar] [CrossRef]
- Fischer, U.; Utke, K.; Somamoto, T.; Köllner, B.; Ototake, M.; Nakanishi, T. Cytotoxic activities of fish leucocytes. Fish Shellfish. Immunol. 2006, 20, 209–226. [Google Scholar] [CrossRef]
- Sveen, L.; Karlsen, C.; Ytteborg, E. Mechanical induced wounds in fish—A review on models and healing mechanisms. Rev. Aquac. 2020, 12, 2446–2465. [Google Scholar] [CrossRef]
- García-Fernández, C.; Sánchez, J.A.; Blanco, G. Characterization of the gilthead seabream (Sparus aurata L.) transferrin gene: Genomic structure, constitutive expression and SNP variation. Fish Shellfish. Immunol. 2011, 31, 548–556. [Google Scholar] [CrossRef]
- Bayne, C.J.; Gerwick, L. The acute phase response and innate immunity of fish. Dev. Comp. Immunol. 2001, 25, 725–743. [Google Scholar] [CrossRef] [PubMed]
- Stafford, J.L.; Belosevic, M. Transferrin and the innate immune response of fish: Identification of a novel mechanism of macrophage activation. Dev. Comp. Immunol. 2003, 27, 539–554. [Google Scholar] [CrossRef]
- Freedman, S.J. The role of alpha 2-macroglobulin in furunculosis: A comparison of rainbow trout and brook trout. Comp. Biochem. Physiol. B Comp. Biochem. 1991, 98, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, S.; Sahoo, P. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Hellio, C.; Pons, A.M.; Beaupoil, C.; Bourgougnon, N.; Le Gal, Y. Antibacterial, antifungal and cytotoxic activities of extracts from fish epidermis and epidermal mucus. Int. J. Antimicrob. Agents 2002, 20, 214–219. [Google Scholar] [CrossRef]
- Maier, V.H.; Dorn, K.V.; Gudmundsdottir, B.K.; Gudmundsson, G.H. Characterisation of cathelicidin gene family members in divergent fish species. Mol. Immunol. 2008, 45, 3723–3730. [Google Scholar] [CrossRef]
- Birkemo, G.A.; Lüders, T.; Andersen, Ø.; Nes, I.F.; Nissen-Meyer, J. Hipposin, a histone-derived antimicrobial peptide in Atlantic halibut (Hippoglossus hippoglossus L.). Biochim. Et Biophys. Acta BBA Proteins Proteom. 2003, 1646, 207–215. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Song, W.-C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cecconi, F.; Choi, A.M.; Chu, C.T.; Codogno, P.; Colombo, M.I. Molecular definitions of autophagy and related processes. EMBO J. 2017, 36, 1811–1836. [Google Scholar] [CrossRef]
- Goldsby, R.; Kindt, T.; Osborne, B.A.; Kuby, J.; Goldsby, R. Immunology, 5th, ed.; WH Freeman & Co: New York, NY, USA, 2002. [Google Scholar]
- Kaufman, J. Unfinished business: Evolution of the MHC and the adaptive immune system of jawed vertebrates. Annu. Rev. Immunol. 2018, 36, 383–409. [Google Scholar] [CrossRef]
- Radwan, J.; Babik, W.; Kaufman, J.; Lenz, T.L.; Winternitz, J. Advances in the evolutionary understanding of MHC polymorphism. Trends Genet. 2020, 36, 298–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Dijkstra, J.M. Major histocompatibility complex (MHC) genes and disease resistance in fish. Cells 2019, 8, 378. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, A.; Miller, N.; Clem, L. Phylogeny of immune recognition: Role of alloantigens in antigen presentation in channel catfish immune responses. Immunology 1991, 74, 165. [Google Scholar] [PubMed]
- Vallejo, A.N.; Miller, N.W.; Harvey, N.E.; Cuchens, M.A.; Warr, G.W.; Clem, L.W. Cellular pathway (s) of antigen processing and presentation in fish APC: Endosomal involvement and cell-free antigen presentation. Dev. Immunol. 1992, 3, 51–65. [Google Scholar] [CrossRef]
- Kumar, G.; Hummel, K.; Noebauer, K.; Welch, T.J.; Razzazi-Fazeli, E.; El-Matbouli, M. Proteome analysis reveals a role of rainbow trout lymphoid organs during Yersinia ruckeri infection process. Sci. Rep. 2018, 8, 1–13. [Google Scholar]
- Brattgjerd, S.; Evensen, Ø. A sequential light microscopic and ultrastructural study on the uptake and handling of Vibrio salmonicida in phagocytes of the head kidney in experimentally infected Atlantic salmon (Salmo salar L.). Vet. Pathol. 1996, 33, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Page, D.M.; Wittamer, V.; Bertrand, J.Y.; Lewis, K.L.; Pratt, D.N.; Delgado, N.; Schale, S.E.; McGue, C.; Jacobsen, B.H.; Doty, A. An evolutionarily conserved program of B-cell development and activation in zebrafish. Blood J. Am. Soc. Hematol. 2013, 122, e1–e11. [Google Scholar] [CrossRef]
- Stosik, M.P.; Tokarz-Deptuła, B.; Deptuła, W. Specific humoral immunity in Osteichthyes. Cent. Eur. J. Immunol. 2018, 43, 335–340. [Google Scholar] [CrossRef]
- Dijkstra, J.M.; Köllner, B.; Aoyagi, K.; Sawamoto, Y.; Kuroda, A.; Ototake, M.; Nakanishi, T.; Fischer, U. The rainbow trout classical MHC class I molecule Onmy-UBA* 501 is expressed in similar cell types as mammalian classical MHC class I molecules. Fish Shellfish. Immunol. 2003, 14, 1–23. [Google Scholar] [CrossRef]
- Chang, Y.-T.; Kai, Y.-H.; Chi, S.-C.; Song, Y.-L. Cytotoxic CD8α+ leucocytes have heterogeneous features in antigen recognition and class I MHC restriction in grouper. Fish Shellfish. Immunol. 2011, 30, 1283–1293. [Google Scholar] [CrossRef]
- Ackerman, A.L.; Cresswell, P. Cellular mechanisms governing cross-presentation of exogenous antigens. Nat. Immunol. 2004, 5, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, C.; Chaves-Pozo, E. Antigen Presentation and Autophagy in Teleost Adaptive Immunity. Int. J. Mol. Sci. 2022, 23, 4899. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, A.; Esteban, M.Á.; Meseguer, J. Cloning, distribution and up-regulation of the teleost fish MHC class II alpha suggests a role for granulocytes as antigen-presenting cells. Mol. Immunol. 2006, 43, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.L.; Del Cid, N.; Traver, D. Perspectives on antigen presenting cells in zebrafish. Dev. Comp. Immunol. 2014, 46, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Iliev, D.B.; Thim, H.; Lagos, L.; Olsen, R.; Jørgensen, J.B. Homing of antigen-presenting cells in head kidney and spleen–salmon head kidney hosts diverse APC types. Front. Immunol. 2013, 4, 137. [Google Scholar] [CrossRef]
- Yoshida, S.H.; Stuge, T.B.; Miller, N.W.; Clem, L.W. Phylogeny of lymphocyte heterogeneity: Cytotoxic activity of channel catfish peripheral blood leukocytes directed against allogeneic targets. Dev. Comp. Immunol. 1995, 19, 71–77. [Google Scholar] [CrossRef]
- Evans, D.L.; Smith, E.E., Jr.; Brown, F.E. Nonspectific cytotoxic cells in fish (Ictaluruspunctatus) VI. Flow cytometric analysis. Dev. Comp. Immunol. 1987, 11, 95–104. [Google Scholar] [CrossRef]
- Meseguer, J.; Esteban, M.A.; Lopez-ruiz, A.; Bielek, E. Ultrastructure of nonspecific cytotoxic cells in teleosts. I. Effector-target cell binding in a marine and a freshwater species (Seabream: Sparus aurata L., and Carp: Cyprinus carpio L.). Anat. Rec. 1994, 239, 468–474. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, Q.; Wang, Z.; Tang, J.; Lu, Y.; Qin, Q.; Cai, J.; Jian, J. Fish natural killer enhancing factor-A (NKEF-A) enhance cytotoxicity of nonspecific cytotoxic cells against bacterial infection. Mol. Immunol. 2021, 133, 184–193. [Google Scholar] [CrossRef]
- Jaso-Friedmann, L.; Leary, J.H., III; Evans, D.L. The non-specific cytotoxic cell receptor (NCCRP-1): Molecular organization and signaling properties. Dev. Comp. Immunol. 2001, 25, 701–711. [Google Scholar] [CrossRef]
- Praveen, K.; Evans, D.L.; Jaso-Friedmann, L. Evidence for the existence of granzyme-like serine proteases in teleost cytotoxic cells. J. Mol. Evol. 2004, 58, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.L.; Hogan, K.T.; Graves, S.S., Carlson; Carlson, R.L., Jr.; Floyd, E.; Dawe, D.L. Nonspecific cytotoxic cells in fish (Ictaluruspunctatus). III. Biophysical and biochemical properties affecting cytolysis. Dev. Comp. Immunol. 1984, 8, 599–610. [Google Scholar] [CrossRef]
- Shen, L.; Stuge, T.B.; Evenhuis, J.P.; Bengtén, E.; Wilson, M.; Chinchar, V.G.; Clem, L.W.; Miller, N.W. Channel catfish NK-like cells are armed with IgM via a putative FcμR. Dev. Comp. Immunol. 2003, 27, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Jaso-Friedmann, L.; Leary, J.H., III; Evans, D.L. NCCRP-1: A novel receptor protein sequenced from teleost nonspecific cytotoxic cells. Mol. Immunol. 1997, 34, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Praveen, K.; Evans, D.L.; Jaso-Friedmann, L. Constitutive expression of tumor necrosis factor-alpha in cytotoxic cells of teleosts and its role in regulation of cell-mediated cytotoxicity. Mol. Immunol. 2006, 43, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Jaso-Friedmann, L.; Ruiz, J.; Bishop, G.; Evans, D. Regulation of innate immunity in tilapia: Activation of nonspecific cytotoxic cells by cytokine-like factors. Dev. Comp. Immunol. 2000, 24, 25–36. [Google Scholar] [CrossRef]
- Secombes, C.; Fletcher, T. The role of phagocytes in the protective mechanisms of fish. Annu. Rev. Fish Dis. 1992, 2, 53–71. [Google Scholar] [CrossRef]
- Schoor, W.P.; Plumb, J.A. Induction of nitric oxide synthase in channel catfish Ictalurus punctatus by Edwardsiella ictaluri. Dis. Aquat. Org. 1994, 19, 153–155. [Google Scholar] [CrossRef]
- Stafford, J.; Neumann, N.; Belosevic, M. Products of proteolytic cleavage of transferrin induce nitric oxide response of goldfish macrophages. Dev. Comp. Immunol. 2001, 25, 101–115. [Google Scholar] [CrossRef]
- Neumann, N.F.; Stafford, J.L.; Barreda, D.; Ainsworth, A.J.; Belosevic, M. Antimicrobial mechanisms of fish phagocytes and their role in host defense. Dev. Comp. Immunol. 2001, 25, 807–825. [Google Scholar] [CrossRef]
- Abelli, L.; Picchietti, S.; Romano, N.; Mastrolia, L.; Scapigliati, G. Immunohistochemistry of gut-associated lymphoid tissue of the sea bassDicentrarchus labrax (L.). Fish Shellfish. Immunol. 1997, 7, 235–245. [Google Scholar] [CrossRef]
- Secombes, C.; Wang, T. The innate and adaptive immune system of fish. In Infectious Disease in Aquaculture; Elsevier: Amsterdam, The Netherlands, 2012; pp. 3–68. [Google Scholar]
- Arango Duque, G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanoni, I.; Ostuni, R.; Marek, L.R.; Barresi, S.; Barbalat, R.; Barton, G.M.; Granucci, F.; Kagan, J.C. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell 2011, 147, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C. The nonspecific immune system: Cellular defenses. Fish Immune Syst. Org. Pathog. Environ. 1996, 15, 63–103. [Google Scholar]
- Biller-Takahashi, J.D.; Urbinati, E.C. Fish Immunology. The modification and manipulation of the innate immune system: Brazilian studies. Anais Academia Brasileira Ciências 2014, 86, 1484–1506. [Google Scholar] [CrossRef]
- Toledo-Ibarra, G.A.; Rojas-Mayorquín, A.E.; Girón-Pérez, M.I. Influence of the cholinergic system on the immune response of teleost fishes: Potential model in biomedical research. Clin. Dev. Immunol. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Lazado, C.C.; Brinchmann, M.F.; Kiron, V. Transcription of selected immune-related genes in spleen cells of cod, Gadus morhua following incubation with alginic acid and β-glucan. J. Exp. Mar. Biol. Ecol. 2012, 416, 202–207. [Google Scholar] [CrossRef]
- Mokhtar, D.M. Characterization of the fish ovarian stroma during the spawning season: Cytochemical, immunohistochemical and ultrastructural studies. Fish Shellfish. Immunol. 2019, 94, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Hine, P. The granulocytes of fish. Fish Shellfish. Immunol. 1992, 2, 79–98. [Google Scholar] [CrossRef]
- Martins, M.L.; de Moraes, F.R.; de Moraes, J.R.E.; Malheiros, E.B. Falha na resposta do cortisol ao estresse por captura e por carragenina em Piaractus mesopotamicus Holmberg, 1887 (Osteichthyes: Characidae). Acta Scientiarum. Biol. Sci. 2000, 22, 545–552. [Google Scholar]
- Tavares-Dias, M.; Schalch, S.H.C.; Martins, M.L.; Silva, É.D.; Moraes, F.R.; Perecin, D. Hematologia de teleósteos brasileiros com infecção parasitária. I. Variáveis do Leporinus macrocephalus Garavelo e Britski, 1988 (Anostomidae) e Piaractus mesopotamicus Holmberg, 1887 (Characidae). Acta Scientiarum. Biol. Sci. 1999, 21, 337–342. [Google Scholar]
- Stuart, L.M.; Ezekowitz, R.A.B. Phagocytosis: Elegant complexity. Immunity 2005, 22, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, D.M. Fish Histology: From Cells to Organs; Apple Academic Press: Palm Bay, FL, USA, 2017. [Google Scholar]
- Sayed, R.K.; Zaccone, G.; Capillo, G.; Albano, M.; Mokhtar, D.M. Structural and Functional Aspects of the Spleen in Molly Fish Poecilia sphenops (Valenciennes, 1846): Synergistic Interactions of Stem Cells, Neurons, and Immune Cells. Biology 2022, 11, 779. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Zhu, L.-Y.; Nie, L.; Shi, W.; Dong, W.-R.; Xiang, L.-X.; Shao, J.-Z. Characterization of surface phenotypic molecules of teleost dendritic cells. Dev. Comp. Immunol. 2015, 49, 38–43. [Google Scholar] [CrossRef]
- Lugo-Villarino, G.; Balla, K.M.; Stachura, D.L.; Bañuelos, K.; Werneck, M.B.; Traver, D. Identification of dendritic antigen-presenting cells in the zebrafish. Proc. Natl. Acad. Sci. USA 2010, 107, 15850–15855. [Google Scholar] [CrossRef] [Green Version]
- Soleto, I.; Granja, A.G.; Simón, R.; Morel, E.; Díaz-Rosales, P.; Tafalla, C. Identification of CD8α+ dendritic cells in rainbow trout (Oncorhynchus mykiss) intestine. Fish Shellfish. Immunol. 2019, 89, 309–318. [Google Scholar] [CrossRef]
- Alesci, A.; Capillo, G.; Fumia, A.; Messina, E.; Albano, M.; Aragona, M.; Lo Cascio, P.; Spanò, N.; Pergolizzi, S.; Lauriano, E.R. Confocal characterization of intestinal dendritic cells from myxines to teleosts. Biology 2022, 11, 1045. [Google Scholar] [CrossRef]
- Kaattari, S.; Brown, G.; Kaattari, I.; Ye, J.; Haines, A.; Bromage, E. The Cellular and Developmental Biology of the Teleost Antibody Response. In Fish Defenses: Immunology; Zaccone, G., Meseguer, J., Garcia-Ayala, A., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2009; pp. 75–129. [Google Scholar]
- Nutt, S.L.; Hodgkin, P.D.; Tarlinton, D.M.; Corcoran, L.M. The generation of antibody-secreting plasma cells. Nat. Rev. Immunol. 2015, 15, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr. How the immune system works to protect the host from infection: A personal view. Proc. Natl. Acad. Sci. USA 2001, 98, 7461–7468. [Google Scholar] [CrossRef]
- Kurosaki, T.; Kometani, K.; Ise, W. Memory B cells. Nat. Rev. Immunol. 2015, 15, 149–159. [Google Scholar] [CrossRef]
- Castro, R.; Bernard, D.; Lefranc, M.; Six, A.; Benmansour, A.; Boudinot, P. T cell diversity and TcR repertoires in teleost fish. Fish Shellfish. Immunol. 2011, 31, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Gérard, A.; Khan, O.; Beemiller, P.; Oswald, E.; Hu, J.; Matloubian, M.; Krummel, M.F. Secondary T cell—T cell synaptic interactions drive the differentiation of protective CD8+ T cells. Nat. Immunol. 2013, 14, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Boschi, I.; Randelli, E.; Buonocore, F.; Casani, D.; Bernini, C.; Fausto, A.M.; Scapigliati, G. Transcription of T cell-related genes in teleost fish, and the European sea bass (Dicentrarchus labrax) as a model. Fish Shellfish. Immunol. 2011, 31, 655–662. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Katakura, F.; Someya, K.; Dijkstra, J.M.; Moritomo, T.; Nakanishi, T. Clonal growth of carp (Cyprinus carpio) T cells in vitro: Long-term proliferation of Th2-like cells. Fish Shellfish. Immunol. 2013, 34, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Koppang, E.O.; Fischer, U.; Moore, L.; Tranulis, M.A.; Dijkstra, J.M.; Köllner, B.; Aune, L.; Jirillo, E.; Hordvik, I. Salmonid T cells assemble in the thymus, spleen and in novel interbranchial lymphoid tissue. J. Anat. 2010, 217, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, D.M. Fish Histology: From Cells to Organs, 2nd ed.; Apple Academic Press: New york, NY, USA, 2021. [Google Scholar]
- Lauriano, E.R.; Calò, M.; Silvestri, G.; Zaccone, D.; Pergolizzi, S.; Cascio, P.L. Mast cells in the intestine and gills of the sea bream, Sparus aurata, exposed to a polychlorinated biphenyl, PCB 126. Acta Histochem. 2012, 114, 166–171. [Google Scholar] [CrossRef]
- Reite, O.B.; Evensen, Ø. Inflammatory cells of teleostean fish: A review focusing on mast cells/eosinophilic granule cells and rodlet cells. Fish Shellfish. Immunol. 2006, 20, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.; Mulero, I.; Mulero, V.; Noga, E.J. Detection of antimicrobial peptides related to piscidin 4 in important aquacultured fish. Dev. Comp. Immunol. 2010, 34, 331–343. [Google Scholar] [CrossRef]
- Zaccone, G.; Capillo, G.; Fernandes, J.M.O.; Kiron, V.; Lauriano, E.R.; Alesci, A.; Lo Cascio, P.; Guerrera, M.C.; Kuciel, M.; Zuwala, K. Expression of the Antimicrobial Peptide Piscidin 1 and Neuropeptides in Fish Gill and Skin: A Potential Participation in Neuro-Immune Interaction. Mar. Drugs 2022, 20, 145. [Google Scholar] [CrossRef]
- Capillo, G.; Zaccone, G.; Cupello, C.; Fernandes, J.M.O.; Viswanath, K.; Kuciel, M.; Zuwala, K.; Guerrera, M.C.; Aragona, M.; Icardo, J.M. Expression of acetylcholine, its contribution to regulation of immune function and O2 sensing and phylogenetic interpretations of the African butterfly fish Pantodon buchholzi (Osteoglossiformes, Pantodontidae). Fish Shellfish. Immunol. 2021, 111, 189–200. [Google Scholar] [CrossRef]
- Kageyama-Yahara, N.; Suehiro, Y.; Yamamoto, T.; Kadowaki, M. IgE-induced degranulation of mucosal mast cells is negatively regulated via nicotinic acetylcholine receptors. Biochem. Biophys. Res. Commun. 2008, 377, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Zaccone, D.; Icardo, J.M.; Kuciel, M.; Alesci, A.; Pergolizzi, S.; Satora, L.; Lauriano, E.R.; Zaccone, G. Polymorphous granular cells in the lung of the primitive fish, the bichir P olypterus senegalus. Acta Zool. 2017, 98, 13–19. [Google Scholar] [CrossRef]
- Reite, O.B. The rodlet cells of teleostean fish: Their potential role in host defence in relation to the role of mast cells/eosinophilic granule cells. Fish Shellfish. Immunol. 2005, 19, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Jordanova, M.; Miteva, N.; Rocha, E. A quantitative study of the hepatic eosinophilic granule cells and rodlet cells during the breeding cycle of Ohrid trout, Salmo letnica Kar.(Teloestei, Salmonidae). Fish Shellfish. Immunol. 2007, 23, 473–478. [Google Scholar] [CrossRef]
- Siderits, D.; Bielek, E. Rodlet cells in the thymus of the zebrafish Danio rerio (Hamilton, 1822). Fish Shellfish. Immunol. 2009, 27, 539–548. [Google Scholar] [CrossRef]
- Mokhtar, D.M. Histological and Ultrastructural Studies of the Unique Hemopoietic-Endocrine Organ of the Grass Carp, Ctenopharyngodon idella (Valenciennes, 1844). Microsc. Microanal. 2020, 26, 1264–1273. [Google Scholar] [CrossRef]
- Rideout, R.; Smith, S.; Morgan, M. High-density aggregations of rodlet cells in the gonads of Greenland halibut Reinhardtius hippoglossoides, a deep-water marine flatfish. J. Fish Biol. 2015, 86, 1630–1637. [Google Scholar] [CrossRef]
- Mendonça, I.; Matos, E.; Rodrigues, G.; Matos, P.; Casal, G.; Azevedo, C. Rodlet cells from the gills and kidneys of two brazilian freshwater fishes: An ultrastructural study. J. Morphol. Sci. 2017, 22, 0. [Google Scholar]
- Leino, R.L. Reaction of rodlet cells to a myxosporean infection in kidney of the bluegill, Lepomis macrochirus. Can. J. Zool. 1996, 74, 217–225. [Google Scholar] [CrossRef]
- Imagawa, T.; Hashimoto, Y.; Kon, Y.; Sugimura, M. Lectin histochemistry as special markers for rodlet cells in carp, Cyprinus carpio L. J. Fish Dis. 1990, 13, 537–540. [Google Scholar] [CrossRef]
- Bosi, G.; DePasquale, J.; Manera, M.; Castaldelli, G.; Giari, L.; Sayyaf Dezfuli, B. Histochemical and immunohistochemical characterization of rodlet cells in the intestine of two teleosts, Anguilla anguilla and Cyprinus carpio. J. Fish Dis. 2018, 41, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.D.; Landis, E.D.; Phillips, R.B. Discovery of a unique Ig heavy-chain isotype (IgT) in rainbow trout: Implications for a distinctive B cell developmental pathway in teleost fish. Proc. Natl. Acad. Sci. USA 2005, 102, 6919–6924. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Hamuro, K.; Suetake, H.; Saha, N.R.; Kikuchi, K.; Suzuki, Y. A teleost polymeric Ig receptor exhibiting two Ig-like domains transports tetrameric IgM into the skin. J. Immunol. 2007, 178, 5682–5689. [Google Scholar] [CrossRef]
- Wittamer, V.; Bertrand, J.Y.; Gutschow, P.W.; Traver, D. Characterization of the mononuclear phagocyte system in zebrafish. Blood J. Am. Soc. Hematol. 2011, 117, 7126–7135. [Google Scholar] [CrossRef]
- Bassity, E.; Clark, T.G. Functional identification of dendritic cells in the teleost model, rainbow trout (Oncorhynchus mykiss). PLoS ONE 2012, 7, e33196. [Google Scholar] [CrossRef] [Green Version]
- Langevin, C.; Aleksejeva, E.; Passoni, G.; Palha, N.; Levraud, J.-P.; Boudinot, P. The antiviral innate immune response in fish: Evolution and conservation of the IFN system. J. Mol. Biol. 2013, 425, 4904–4920. [Google Scholar] [CrossRef]
- Cone, R.A. Barrier properties of mucus. Adv. Drug Deliv. Rev. 2009, 61, 75–85. [Google Scholar] [CrossRef]
- Mokhtar, D.M.; Hussein, M.M.; Sayed, R.K. Novel Identification and Microscopy of the Intestinal Bulb of Molly Fish (Poecilia sphenops) with a Focus on Its Role in Immunity. Microsc. Microanal. 2022, 28, 1827–1839. [Google Scholar] [CrossRef]
- Arasu, A.; Kumaresan, V.; Sathyamoorthi, A.; Palanisamy, R.; Prabha, N.; Bhatt, P.; Roy, A.; Thirumalai, M.K.; Gnanam, A.J.; Pasupuleti, M. Fish lily type lectin-1 contains β-prism architecture: Immunological characterization. Mol. Immunol. 2013, 56, 497–506. [Google Scholar] [CrossRef]
- Charroux, B.; Capo, F.; Kurz, C.L.; Peslier, S.; Chaduli, D.; Viallat-Lieutaud, A.; Royet, J. Cytosolic and secreted peptidoglycan-degrading enzymes in Drosophila respectively control local and systemic immune responses to microbiota. Cell Host Microbe 2018, 23, 215–228.e214. [Google Scholar] [CrossRef] [PubMed]
- Arockiaraj, J.; Kumaresan, V.; Bhatt, P.; Palanisamy, R.; Gnanam, A.J.; Pasupuleti, M.; Kasi, M.; Chaurasia, M.K. A novel single-domain peptide, anti-LPS factor from prawn: Synthesis of peptide, antimicrobial properties and complete molecular characterization. Peptides 2014, 53, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ekman, D.; Skelton, D.; Davis, J.; Villeneuve, D.; Cavallin, J.; Schroeder, A.; Jensen, K.; Ankley, G.T.; Collette, T. Metabolite profiling of fish skin mucus: A novel approach for minimally-invasive environmental exposure monitoring and surveillance. Environ. Sci. Technol. 2015, 49, 3091–3100. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Gunasekaran, G. Fatty acids and amino acids composition in skin epidermal mucus of selected fresh water fish Mugil cephalus. World J. Pharm. Pharm. Sci. 2015, 4, 1275–1287. [Google Scholar]
- Rose, M.C.; Voynow, J.A. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol. Rev. 2006, 86, 245–278. [Google Scholar] [CrossRef]
- Perez-Vilar, J.; Hill, R.L. The Structure and Assembly of Secreted Mucins* 210. J. Biol. Chem. 1999, 274, 31751–31754. [Google Scholar] [CrossRef]
- Strous, G.J.; Dekker, J. Mucin-type glycoproteins. Crit. Rev. Biochem. Mol. Biol. 1992, 27, 57–92. [Google Scholar] [CrossRef]
- Desseyn, J.-L.; Tetaert, D.; Gouyer, V. Architecture of the large membrane-bound mucins. Gene 2008, 410, 215–222. [Google Scholar] [CrossRef]
- Jevtov, I.; Samuelsson, T.; Yao, G.; Amsterdam, A.; Ribbeck, K. Zebrafish as a model to study live mucus physiology. Sci. Rep. 2014, 4, 1–6. [Google Scholar] [CrossRef]
- Perez-Sanchez, J.; Estensoro, I.; Redondo, M.J.; Calduch-Giner, J.A.; Kaushik, S.; Sitja-Bobadilla, A. Mucins as diagnostic and prognostic biomarkers in a fish-parasite model: Transcriptional and functional analysis. PLoS ONE 2013, 8, e65457. [Google Scholar] [CrossRef]
- van der Marel, M.; Adamek, M.; Gonzalez, S.F.; Frost, P.; Rombout, J.H.; Wiegertjes, G.F.; Savelkoul, H.F.; Steinhagen, D. Molecular cloning and expression of two β-defensin and two mucin genes in common carp (Cyprinus carpio L.) and their up-regulation after β-glucan feeding. Fish Shellfish. Immunol. 2012, 32, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Malachowicz, M.; Wenne, R.; Burzynski, A. De novo assembly of the sea trout (Salmo trutta m. trutta) skin transcriptome to identify putative genes involved in the immune response and epidermal mucus secretion. PLoS ONE 2017, 12, e0172282. [Google Scholar] [CrossRef] [PubMed]
- Sveen, L.R.; Grammes, F.T.; Ytteborg, E.; Takle, H.; Jørgensen, S.M. Genome-wide analysis of Atlantic salmon (Salmo salar) mucin genes and their role as biomarkers. PLoS ONE 2017, 12, e0189103. [Google Scholar] [CrossRef]
- Marcos-López, M.; Calduch-Giner, J.A.; Mirimin, L.; MacCarthy, E.; Rodger, H.D.; O’Connor, I.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J.; Piazzon, M.C. Gene expression analysis of Atlantic salmon gills reveals mucin 5 and interleukin 4/13 as key molecules during amoebic gill disease. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Van der Marel, M.; Caspari, N.; Neuhaus, H.; Meyer, W.; Enss, M.L.; Steinhagen, D. Changes in skin mucus of common carp, Cyprinus carpio L., after exposure to water with a high bacterial load. J. Fish Dis. 2010, 33, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Estensoro, I.; Jung-Schroers, V.; Álvarez-Pellitero, P.; Steinhagen, D.; Sitjà-Bobadilla, A. Effects of Enteromyxum leei (Myxozoa) infection on gilthead sea bream (Sparus aurata)(Teleostei) intestinal mucus: Glycoprotein profile and bacterial adhesion. Parasitol. Res. 2013, 112, 567–576. [Google Scholar] [CrossRef]
- Rajanbabu, V.; Chen, J.-Y. Applications of antimicrobial peptides from fish and perspectives for the future. Peptides 2011, 32, 415–420. [Google Scholar] [CrossRef]
- Zaccone, G.; Kapoor, B.; Fasulo, S.; Ainis, L. Structural, histochemical and functional aspects of the epidermis of fishes. In Advances in Marine Biology; Southward, A., Tyler, P., Young, C., Fuiman, L., Eds.; Academic Press: London, UK, 2001; Volume 40, pp. 253–348. [Google Scholar]
- Salinas, I.; Fernández-Montero, Á.; Ding, Y.; Sunyer, J.O. Mucosal immunoglobulins of teleost fish: A decade of advances. Dev. Comp. Immunol. 2021, 121, 104079. [Google Scholar] [CrossRef]
- Tacchi, L.; Musharrafieh, R.; Larragoite, E.T.; Crossey, K.; Erhardt, E.B.; Martin, S.A.; LaPatra, S.E.; Salinas, I. Nasal immunity is an ancient arm of the mucosal immune system of vertebrates. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Xu, Z.; Takizawa, F.; Parra, D.; Gómez, D.; von Gersdorff Jørgensen, L.; LaPatra, S.E.; Sunyer, J.O. Mucosal immunoglobulins at respiratory surfaces mark an ancient association that predates the emergence of tetrapods. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Xu, Z.; Parra, D.; Gómez, D.; Salinas, I.; Zhang, Y.-A.; von Gersdorff Jørgensen, L.; Heinecke, R.D.; Buchmann, K.; LaPatra, S.; Sunyer, J.O. Teleost skin, an ancient mucosal surface that elicits gut-like immune responses. Proc. Natl. Acad. Sci. USA 2013, 110, 13097–13102. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-Y.; Kong, W.-G.; Xu, H.-Y.; Huang, Z.-Y.; Zhang, X.-T.; Ding, L.-G.; Dong, S.; Yin, G.-M.; Dong, F.; Yu, W. Convergent evolution of mucosal immune responses at the buccal cavity of teleost fish. Iscience 2019, 19, 821–835. [Google Scholar] [CrossRef]
- Løken, O.M.; Bjørgen, H.; Hordvik, I.; Koppang, E.O. A teleost structural analogue to the avian bursa of Fabricius. J. Anat. 2020, 236, 798–808. [Google Scholar] [CrossRef]
- Reardon, C.; Duncan, G.S.; Brüstle, A.; Brenner, D.; Tusche, M.W.; Olofsson, P.S.; Rosas-Ballina, M.; Tracey, K.J.; Mak, T.W. Lymphocyte-derived ACh regulates local innate but not adaptive immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 1410–1415. [Google Scholar] [CrossRef]
- Pan, J.; Zhang, L.; Shao, X.; Huang, J. Acetylcholine from tuft cells: The updated insights beyond its immune and chemosensory functions. Front. Cell Dev. Biol. 2020, 8, 606. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish. Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.K. Role of gastrointestinal microbiota in fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar] [CrossRef]
- Torrecillas, S.; Makol, A.; Betancor, M.B.; Montero, D.; Caballero, M.J.; Sweetman, J.; Izquierdo, M. Enhanced intestinal epithelial barrier health status on European sea bass (Dicentrarchus labrax) fed mannan oligosaccharides. Fish Shellfish. Immunol. 2013, 34, 1485–1495. [Google Scholar] [CrossRef]
- Fournier-Betz, V.; Quentel, C.; Lamour, F.; LeVen, A. Immunocytochemical detection of Ig-positive cells in blood, lymphoid organs and the gut associated lymphoid tissue of the turbot (Scophthalmus maximus). Fish Shellfish. Immunol. 2000, 10, 187–202. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Giovinazzo, G.; Lui, A.; Giari, L. Inflammatory response to Dentitruncus truttae (Acanthocephala) in the intestine of brown trout. Fish Shellfish. Immunol. 2008, 24, 726–733. [Google Scholar] [CrossRef]
- Inami, M.; Taverne-Thiele, A.J.; Schrøder, M.B.; Kiron, V.; Rombout, J.H. Immunological differences in intestine and rectum of Atlantic cod (Gadus morhua L.). Fish Shellfish. Immunol. 2009, 26, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Mulder, I.; Wadsworth, S.; Secombes, C. Cytokine expression in the intestine of rainbow trout (Oncorhynchus mykiss) during infection with Aeromonas salmonicida. Fish Shellfish. Immunol. 2007, 23, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.D.; Alder, M.N. The evolution of adaptive immune systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F. A cold-blooded view of adaptive immunity. Nat. Rev. Immunol. 2018, 18, 438–453. [Google Scholar] [CrossRef]
- Magadan, S.; Sunyer, O.J.; Boudinot, P. Unique features of fish immune repertoires: Particularities of adaptive immunity within the largest group of vertebrates. Pathog. Host Interact. Antigen. Var. Somat. Adapt. 2015, 57, 235–264. [Google Scholar]
- Mashoof, S.; Criscitiello, M.F. Fish immunoglobulins. Biology 2016, 5, 45. [Google Scholar] [CrossRef]
- Wu, L.; Qin, Z.; Liu, H.; Lin, L.; Ye, J.; Li, J. Recent advances on phagocytic B cells in teleost fish. Front. Immunol. 2020, 11, 824. [Google Scholar] [CrossRef]
- Acton, R.T.; Weinheimer, P.F.; Hall, S.J.; Niedermeier, W.; Shelton, E.; Bennett, J.C. Tetrameric immune macroglobulins in three orders of bony fishes. Proc. Natl. Acad. Sci. USA 1971, 68, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Flajnik, M.F. Comparative analyses of immunoglobulin genes: Surprises and portents. Nat. Rev. Immunol. 2002, 2, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Lobb, C.J.; Clem, L.W. Phylogeny of immunoglobulin structure and function. XI. Secretory immunoglobulins in the cutaneous mucus of the sheepshead, Archosargus probatocephalus. Dev. Comp. Immunol. 1981, 5, 587–596. [Google Scholar] [CrossRef]
- Lobb, C.J.; Clem, L.W. Phylogeny of immunoglobulin structure and function—XII. Secretory immunoglobulins in the bile of the marine teleost Archosargus probatocephalus. Mol. Immunol. 1981, 18, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Danilova, N.; Bussmann, J.; Jekosch, K.; Steiner, L.A. The immunoglobulin heavy-chain locus in zebrafish: Identification and expression of a previously unknown isotype, immunoglobulin Z. Nat. Immunol. 2005, 6, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Bengtén, E.; Clem, L.W.; Miller, N.W.; Warr, G.W.; Wilson, M. Channel catfish immunoglobulins: Repertoire and expression. Dev. Comp. Immunol. 2006, 30, 77–92. [Google Scholar] [CrossRef]
- Bradshaw, W.J.; Valenzano, D.R. Extreme genomic volatility characterizes the evolution of the immunoglobulin heavy chain locus in cyprinodontiform fishes. Proc. R. Soc. B 2020, 287, 20200489. [Google Scholar] [CrossRef] [PubMed]
- Magadán-Mompó, S.; Sánchez-Espinel, C.; Gambón-Deza, F. Immunoglobulin heavy chains in medaka (Oryzias latipes). BMC Evol. Biol. 2011, 11, 1–13. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, X.-J.; Chen, D.-D.; Sunyer, J.O.; Zhang, Y.-A. Molecular characterization and expression analysis of three subclasses of IgT in rainbow trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2017, 70, 94–105. [Google Scholar] [CrossRef]
- Wilson, M.; Bengtén, E.; Miller, N.W.; Clem, L.W.; Du Pasquier, L.; Warr, G.W. A novel chimeric Ig heavy chain from a teleost fish shares similarities to IgD. Proc. Natl. Acad. Sci. USA 1997, 94, 4593–4597. [Google Scholar] [CrossRef]
- Ohta, Y.; Flajnik, M. IgD, like IgM, is a primordial immunoglobulin class perpetuated in most jawed vertebrates. Proc. Natl. Acad. Sci. USA 2006, 103, 10723–10728. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Gomez, F.; Greene, W.; Rego, K.; Hansen, J.D.; Costa, G.; Kataria, P.; Bromage, E.S. Discovery and characterization of secretory IgD in rainbow trout: Secretory IgD is produced through a novel splicing mechanism. J. Immunol. 2012, 188, 1341–1349. [Google Scholar] [CrossRef]
- Zou, J.; Carrington, A.; Collet, B.; Dijkstra, J.M.; Yoshiura, Y.; Bols, N.; Secombes, C. Identification and bioactivities of IFN-γ in rainbow trout Oncorhynchus mykiss: The first Th1-type cytokine characterized functionally in fish. J. Immunol. 2005, 175, 2484–2494. [Google Scholar] [CrossRef]
- Zou, J.; Grabowski, P.S.; Cunningham, C.; Secombes, C.J. Molecular cloning of interleukin 1β from rainbow trout Oncorhynchus mykiss reveals no evidence of an ice cut site. Cytokine 1999, 11, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Laing, K.J.; Secombes, C.J. Chemokines. Dev. Comp. Immunol. 2004, 28, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Laing, K.J.; Wang, T.; Zou, J.; Holland, J.; Hong, S.; Bols, N.; Hirono, I.; Aoki, T.; Secombes, C.J. Cloning and expression analysis of rainbow trout Oncorhynchus mykiss tumour necrosis factor-α. Eur. J. Biochem. 2001, 268, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.; Zou, J.; Kono, T.; Sakai, M.; Dijkstra, J.M.; Secombes, C. Characterisation and expression analysis of interleukin 2 (IL-2) and IL-21 homologues in the Japanese pufferfish, Fugu rubripes, following their discovery by synteny. Immunogenetics 2005, 56, 909–923. [Google Scholar] [CrossRef]
- Bird, S.; Zou, J.; Savan, R.; Kono, T.; Sakai, M.; Woo, J.; Secombes, C. Characterisation and expression analysis of an interleukin 6 homologue in the Japanese pufferfish, Fugu rubripes. Dev. Comp. Immunol. 2005, 29, 775–789. [Google Scholar] [CrossRef]
- Zou, J.; Clark, M.S.; Secombes, C.J. Characterisation, expression and promoter analysis of an interleukin 10 homologue in the puffer fish, Fugu rubripes. Immunogenetics 2003, 55, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Bei, J.-X.; Suetake, H.; Araki, K.; Kikuchi, K.; Yoshiura, Y.; Lin, H.-R.; Suzuki, Y. Two interleukin (IL)-15 homologues in fish from two distinct origins. Mol. Immunol. 2006, 43, 860–869. [Google Scholar] [CrossRef]
- Redmond, A.K.; Zou, J.; Secombes, C.J.; Macqueen, D.J.; Dooley, H. Discovery of all three types in cartilaginous fishes enables phylogenetic resolution of the origins and evolution of interferons. Front. Immunol. 2019, 10, 1558. [Google Scholar] [CrossRef] [Green Version]
- Robertsen, B. The interferon system of teleost fish. Fish Shellfish. Immunol. 2006, 20, 172–191. [Google Scholar] [CrossRef]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809. [Google Scholar] [CrossRef]
- De Veer, M.J.; Holko, M.; Frevel, M.; Walker, E.; Der, S.; Paranjape, J.M.; Silverman, R.H.; Williams, B.R. Functional classification of interferon-stimulated genes identified using microarrays. J. Leukoc. Biol. 2001, 69, 912–920. [Google Scholar] [CrossRef]
- Roy, S.; Kumar, V.; Kumar, V.; Behera, B. Acute phase proteins and their potential role as an indicator for fish health and in diagnosis of fish diseases. Protein Pept. Lett. 2017, 24, 78–89. [Google Scholar] [CrossRef]
- Bird, S.; Wang, T.; Zou, J.; Cunningham, C.; Secombes, C.J. The first cytokine sequence within cartilaginous fish: IL-1β in the small spotted catshark (Scyliorhinus canicula). J. Immunol. 2002, 168, 3329–3340. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, K.; Shin, D.-H.; Nakao, M.; Yano, T. Molecular cloning and expression analysis of carp (Cyprinus carpio) interleukin-1β, high affinity immunoglobulin E Fc receptor γ subunit and serum amyloid A. Fish Shellfish. Immunol. 2000, 10, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Cunningham, C.; Secombes, C.J. The rainbow trout Oncorhynchus mykiss interleukin-1β gene has a different organization to mammals and undergoes incomplete splicing. Eur. J. Biochem. 1999, 259, 901–908. [Google Scholar] [CrossRef]
- Li, J.-Y.; Wang, Y.-Y.; Shao, T.; Fan, D.-D.; Lin, A.-F.; Xiang, L.-X.; Shao, J.-Z. The zebrafish NLRP3 inflammasome has functional roles in ASC-dependent interleukin-1β maturation and gasdermin E–mediated pyroptosis. J. Biol. Chem. 2020, 295, 1120–1141. [Google Scholar] [CrossRef] [PubMed]
- Angosto, D.; López-Castejón, G.; López-Muñoz, A.; Sepulcre, M.P.; Arizcun, M.; Meseguer, J.; Mulero, V. Evolution of inflammasome functions in vertebrates: Inflammasome and caspase-1 trigger fish macrophage cell death but are dispensable for the processing of IL-1β. Innate Immun. 2012, 18, 815–824. [Google Scholar] [CrossRef]
- Pelegrın, P.; Chaves-Pozo, E.; Mulero, V.; Meseguer, J. Production and mechanism of secretion of interleukin-1β from the marine fish gilthead seabream. Dev. Comp. Immunol. 2004, 28, 229–237. [Google Scholar] [CrossRef]
- Vojtech, L.N.; Scharping, N.; Woodson, J.C.; Hansen, J.D. Roles of inflammatory caspases during processing of zebrafish interleukin-1β in Francisella noatunensis infection. Infect. Immun. 2012, 80, 2878–2885. [Google Scholar] [CrossRef]
- Yin, Z.; Kwang, J. Carp interleukin-1β in the role of an immuno-adjuvant. Fish Shellfish. Immunol. 2000, 10, 375–378. [Google Scholar] [CrossRef]
- Taechavasonyoo, A.; Hirono, I.; Kondo, H. The immune-adjuvant effect of Japanese flounder Paralichthys olivaceus IL-1β. Dev. Comp. Immunol. 2013, 41, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.; Bird, S.; van Zwieten, R.; Secombes, C.J.; Wang, T. Cloning of the IL-1β3 gene and IL-1β4 pseudogene in salmonids uncovers a second type of IL-1β gene in teleost fish. Dev. Comp. Immunol. 2012, 38, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Li, R.; Xu, Q.; Secombes, C.J.; Wang, T. Two types of TNF-α exist in teleost fish: Phylogeny, expression, and bioactivity analysis of type-II TNF-α3 in rainbow trout Oncorhynchus mykiss. J. Immunol. 2013, 191, 5959–5972. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Zou, J.; Secombes, C.J.; Martin, S.A. Cortisol modulates the induction of inflammatory gene expression in a rainbow trout macrophage cell line. Fish Shellfish. Immunol. 2011, 30, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Maehr, T.; Vecino, J.L.G.; Wadsworth, S.; Wang, T.; Secombes, C.J. Four CISH paralogues are present in rainbow trout Oncorhynchus mykiss: Differential expression and modulation during immune responses and development. Mol. Immunol. 2014, 62, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Engelsma, M.Y.; Stet, R.J.; Saeij, J.P.; Verburg-van Kemenade, B.L. Differential expression and haplotypic variation of two interleukin-1β genes in the common carp (Cyprinus carpio L.). Cytokine 2003, 22, 21–32. [Google Scholar] [CrossRef]
- Hu, B.; Chen, B.; Mao, M.; Chen, M.; Liu, X.; Cui, Q.; Liu, Y.; Jiang, C. Molecular characterization and expression analysis of the interleukin 1b gene in Pacific cod (Gadus macrocephalus). Dev. Comp. Immunol. 2018, 88, 213–218. [Google Scholar] [CrossRef]
- Dinarello, C.A. IL-18: A TH1-inducing, proinflammatory cytokine and new member of the IL-1 family. J. Allergy Clin. Immunol. 1999, 103, 11–24. [Google Scholar] [CrossRef]
- Pérez-Cordón, G.; Estensoro, I.; Benedito-Palos, L.; Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Interleukin gene expression is strongly modulated at the local level in a fish–parasite model. Fish Shellfish. Immunol. 2014, 37, 201–208. [Google Scholar] [CrossRef]
- Venkatesh, B.; Lee, A.P.; Ravi, V.; Maurya, A.K.; Lian, M.M.; Swann, J.B.; Ohta, Y.; Flajnik, M.F.; Sutoh, Y.; Kasahara, M. Elephant shark genome provides unique insights into gnathostome evolution. Nature 2014, 505, 174–179. [Google Scholar] [CrossRef]
- Huising, M.O.; Stet, R.J.; Savelkoul, H.F.; Verburg-van Kemenade, B.L. The molecular evolution of the interleukin-1 family of cytokines; IL-18 in teleost fish. Dev. Comp. Immunol. 2004, 28, 395–413. [Google Scholar] [CrossRef]
- Zou, J.; Bird, S.; Truckle, J.; Bols, N.; Horne, M.; Secombes, C. Identification and expression analysis of an IL-18 homologue and its alternatively spliced form in rainbow trout (Oncorhynchus mykiss). Eur. J. Biochem. 2004, 271, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Rivers-Auty, J.; Daniels, M.J.; Colliver, I.; Robertson, D.L.; Brough, D. Redefining the ancestral origins of the interleukin-1 superfamily. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Biswas, G.; Kinoshita, S.; Kono, T.; Hikima, J.-I.; Sakai, M. Evolutionary evidence of tumor necrosis factor super family members in the Japanese pufferfish (Takifugu rubripes): Comprehensive genomic identification and expression analysis. Mar. Genom. 2015, 22, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Marín, I. Tumor necrosis factor superfamily: Ancestral functions and remodeling in early vertebrate evolution. Genome Biol. Evol. 2020, 12, 2074–2092. [Google Scholar] [CrossRef]
- Pleić, I.L.; Secombes, C.J.; Bird, S.; Mladineo, I. Characterization of three pro-inflammatory cytokines, TNFα1, TNFα2 and IL-1β, in cage-reared Atlantic bluefin tuna Thunnus thynnus. Fish Shellfish. Immunol. 2014, 36, 98–112. [Google Scholar] [CrossRef]
- Saeij, J.P.; Stet, R.J.; De Vries, B.J.; Van Muiswinkel, W.B.; Wiegertjes, G.F. Molecular and functional characterization of carp TNF: A link between TNF polymorphism and trypanotolerance? Dev. Comp. Immunol. 2003, 27, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Secombes, C.J.; Long, S.; Miller, N.; Clem, L.; Chinchar, V. Molecular identification and expression analysis of tumor necrosis factor in channel catfish (Ictalurus punctatus). Dev. Comp. Immunol. 2003, 27, 845–858. [Google Scholar] [CrossRef]
- Veenstra, K.A.; Wangkahart, E.; Wang, T.; Tubbs, L.; Arous, J.B.; Secombes, C.J. Rainbow trout (Oncorhynchus mykiss) adipose tissue undergoes major changes in immune gene expression following bacterial infection or stimulation with pro-inflammatory molecules. Dev. Comp. Immunol. 2018, 81, 83–94. [Google Scholar] [CrossRef]
- Mulero, V.; Meseguer, J. Functional characterisation of a macrophage-activating factor produced by leucocytes of gilthead seabream (Sparus aurataL.). Fish Shellfish. Immunol. 1998, 8, 143–156. [Google Scholar] [CrossRef]
- Tafalla, C.; Figueras, A.; Novoa, B. Viral hemorrhagic septicemia virus alters turbot Scophthalmus maximus macrophage nitric oxide production. Dis. Aquat. Org. 2001, 47, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.; Dios, S.; Novoa, B.; Figueras, A. Characterisation, expression and ontogeny of interleukin-6 and its receptors in zebrafish (Danio rerio). Dev. Comp. Immunol. 2012, 37, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, R.D.; Putoczki, T.L.; Griffin, M.D. Structural understanding of interleukin 6 family cytokine signaling and targeted therapies: Focus on interleukin 11. Front. Immunol. 2020, 11, 1424. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sultan, Y.; Sun, Y.; Zhang, S.; Liu, Y.; Li, X. Expression analysis of Hsp90α and cytokines in zebrafish caudal fin regeneration. Dev. Comp. Immunol. 2021, 116, 103922. [Google Scholar] [CrossRef]
- Eggestøl, H.Ø.; Lunde, H.S.; Knutsen, T.M.; Haugland, G.T. Interleukin-1 ligands and receptors in lumpfish (Cyclopterus lumpus L.): Molecular characterization, phylogeny, gene expression, and transcriptome analyses. Front. Immunol. 2020, 11, 502. [Google Scholar] [CrossRef]
- Veenstra, K.A.; Wang, T.; Alnabulsi, A.; Douglas, A.; Russell, K.S.; Tubbs, L.; Arous, J.B.; Secombes, C.J. Analysis of adipose tissue immune gene expression after vaccination of rainbow trout with adjuvanted bacterins reveals an association with side effects. Mol. Immunol. 2017, 88, 89–98. [Google Scholar] [CrossRef]
- Wang, T.; Holland, J.W.; Bols, N.; Secombes, C.J. Cloning and expression of the first nonmammalian interleukin-11 gene in rainbow trout Oncorhynchus mykiss. FEBS J. 2005, 272, 1136–1147. [Google Scholar] [CrossRef]
- Huising, M.O.; Kruiswijk, C.P.; Van Schijndel, J.E.; Savelkoul, H.F.; Flik, G.; Verburg-van Kemenade, B. Multiple and highly divergent IL-11 genes in teleost fish. Immunogenetics 2005, 57, 432–443. [Google Scholar] [CrossRef]
- Laing, K.; Cunningham, C.; Secombes, C. Genes for three different isoforms of transforming growth factor-β are present in plaice (Pleuronectes platessa) DNA. Fish Shellfish. Immunol. 2000, 10, 261–271. [Google Scholar] [CrossRef]
- Funkenstein, B.; Olekh, E.; Jakowlew, S.B. Identification of a novel transforming growth factor-β (TGF-β6) gene in fish: Regulation in skeletal muscle by nutritional state. BMC Mol. Biol. 2010, 11, 1–16. [Google Scholar] [CrossRef]
- Zhan, Y.; Jimmy, K. Molecular isolation and characterisation of carp transforming growth factor β1 from activated leucocytes. Fish Shellfish. Immunol. 2000, 10, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Yin, L.; Feng, S.; Wang, X.; Yang, K.; Zhang, A.; Zhou, H. Dual-parallel inhibition of IL-10 and TGF-β1 controls LPS-induced inflammatory response via NF-κB signaling in grass carp monocytes/macrophages. Fish Shellfish. Immunol. 2015, 44, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Maehr, T.; Costa, M.M.; Vecino, J.L.G.; Wadsworth, S.; Martin, S.A.; Wang, T.; Secombes, C.J. Transforming growth factor-β1b: A second TGF-β1 paralogue in the rainbow trout (Oncorhynchus mykiss) that has a lower constitutive expression but is more responsive to immune stimulation. Fish Shellfish. Immunol. 2013, 34, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Harms, C.; Kennedy-Stoskopf, S.; Horne, W.; Fuller, F.; Tompkins, W. Cloning and sequencing hybrid striped bass (Morone saxatilis x M. chrysops) transforming growth factor-β (TGF-β), and development of a reverse transcription quantitative competitive polymerase chain reaction (RT-qcPCR) assay to measure TGF-β mRNA of teleost fish. Fish Shellfish. Immunol. 2000, 10, 61–85. [Google Scholar] [PubMed]
- Schluesener, H.J.; Su, Y.; Ebrahimi, A.; Pouladsaz, D. Antimicrobial peptides in the brain: Neuropeptides and amyloid. Front. Biosci. Sch. 2012, 4, 1375–1380. [Google Scholar] [CrossRef]
- McDonald, M.; Mannion, M.; Pike, D.; Lewis, K.; Flynn, A.; Brannan, A.M.; Browne, M.J.; Jackman, D.; Madera, L.; Coombs, M.R.P. Structure–function relationships in histidine-rich antimicrobial peptides from Atlantic cod. Biochim. Biophys. Acta BBA Biomembr. 2015, 1848, 1451–1461. [Google Scholar] [CrossRef]
- Aidy, S.E.; Stilling, R.; Dinan, T.G.; Cryan, J.F. Microbiome to brain: Unravelling the multidirectional axes of communication. Microb. Endocrinol. Interkingdom Signal. Infect. Dis. Health 2016, 874, 301–336. [Google Scholar]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain–gut–microbe communication in health and disease. Front. Physiol. 2011, 2, 94. [Google Scholar] [CrossRef]
- Wang, H.-X.; Wang, Y.-P. Gut microbiota-brain axis. Chin. Med. J. 2016, 129, 2373–2380. [Google Scholar] [CrossRef]
- Kepka, M.; Verburg-van Kemenade, B.; Chadzinska, M. Neuroendocrine modulation of the inflammatory response in common carp: Adrenaline regulates leukocyte profile and activity. Gen. Comp. Endocrinol. 2013, 188, 102–109. [Google Scholar] [CrossRef]
- Zaccone, G.; Mauceri, A.; Maisano, M.; Giannetto, A.; Parrino, V.; Fasulo, S. Innervation and neurotransmitter localization in the lung of the Nile bichir Polypterus bichir bichir. Anat. Rec. Adv. Integr. Anat. Evol. Biol. Adv. Integr. Anat. Evol. Biol. 2007, 290, 1166–1177. [Google Scholar] [CrossRef] [PubMed]
- Aresti Sanz, J.; El Aidy, S. Microbiota and gut neuropeptides: A dual action of antimicrobial activity and neuroimmune response. Psychopharmacology 2019, 236, 1597–1609. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.B.; Mazmanian, S.K. The enteric network: Interactions between the immune and nervous systems of the gut. Immunity 2017, 46, 910–926. [Google Scholar] [CrossRef] [PubMed]
- Rességuier, J.; Dalum, A.S.; Du Pasquier, L.; Zhang, Y.; Koppang, E.O.; Boudinot, P.; Wiegertjes, G.F. Lymphoid tissue in teleost gills: Variations on a theme. Biology 2020, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Capillo, G.; Icardo, J.M.; Fernandes, J.M.O.; Kiron, V.; Kuciel, M.; Zuwala, K.; Guerrera, M.C.; Aragona, M.; Zaccone, G. Neuroepithelial cells (NECs) and mucous cells express a variety of neurotransmitters and neurotransmitter receptors in the gill and respiratory air-sac of the catfish Heteropneustes fossilis (Siluriformes, Heteropneustidae): A possible role in local immune defence. Zoology 2021, 148, 125958. [Google Scholar] [PubMed]
- Fernandes, J.M.; Ruangsri, J.; Kiron, V. Atlantic cod piscidin and its diversification through positive selection. PLoS ONE 2010, 5, e9501. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-W.; Su, B.-C.; Chen, J.-Y. Tilapia piscidin 4 (TP4) reprograms M1 macrophages to M2 phenotypes in cell models of Gardnerella Vaginalis-induced vaginosis. Front. Immunol. 2021, 12, 5123. [Google Scholar] [CrossRef]
- Branchfield, K.; Nantie, L.; Verheyden, J.M.; Sui, P.; Wienhold, M.D.; Sun, X. Pulmonary neuroendocrine cells function as airway sensors to control lung immune response. Science 2016, 351, 707–710. [Google Scholar] [CrossRef]
- Metz, J.R.; Huising, M.O.; Leon, K.; Verburg-van Kemenade, B.L.; Flik, G. Central and peripheral interleukin-1β and interleukin-1 Receptor I expression and their role in the acute stress response of common carp, Cyprinus carpio L. J. Endocrinol. 2006, 191, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Maier, S.F. Bi-Directional immune—brain communication: Implications for understanding stress, pain, and cognition. Brain Behav. Immun. 2003, 17, 69–85. [Google Scholar] [CrossRef]
- Khansari, A.R.; Parra, D.; Reyes-López, F.E.; Tort, L. Modulatory in vitro effect of stress hormones on the cytokine response of rainbow trout and gilthead sea bream head kidney stimulated with Vibrio anguillarum bacterin. Fish Shellfish. Immunol. 2017, 70, 736–749. [Google Scholar] [CrossRef]
- Weber, R.; Pérez Maceira, J.; Aldegunde, M.; Peleteiro, J.; García Martín, L.; Aldegunde, M. Effects of acute handling stress on cerebral monoaminergic neurotransmitters in juvenile Senegalese sole Solea senegalensis. J. Fish Biol. 2015, 87, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Butt, R.L.; Volkoff, H. Gut microbiota and energy homeostasis in fish. Front. Endocrinol. 2019, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Dolan, B.P.; Fisher, K.M.; Colvin, M.E.; Benda, S.E.; Peterson, J.T.; Kent, M.L.; Schreck, C.B. Innate and adaptive immune responses in migrating spring-run adult chinook salmon, Oncorhynchus tshawytscha. Fish Shellfish. Immunol. 2016, 48, 136–144. [Google Scholar] [CrossRef] [PubMed]
- McQuillan, H.J.; Lokman, P.M.; Young, G. Effects of sex steroids, sex, and sexual maturity on cortisol production: An in vitro comparison of chinook salmon and rainbow trout interrenals. Gen. Comp. Endocrinol. 2003, 133, 154–163. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the fish mucosal immunity.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mokhtar, D.M.; Zaccone, G.; Alesci, A.; Kuciel, M.; Hussein, M.T.; Sayed, R.K.A. Main Components of Fish Immunity: An Overview of the Fish Immune System. Fishes 2023, 8, 93. https://doi.org/10.3390/fishes8020093
AMA Style
Mokhtar DM, Zaccone G, Alesci A, Kuciel M, Hussein MT, Sayed RKA. Main Components of Fish Immunity: An Overview of the Fish Immune System. Fishes. 2023; 8(2):93. https://doi.org/10.3390/fishes8020093
Chicago/Turabian StyleMokhtar, Doaa M., Giacomo Zaccone, Alessio Alesci, Michal Kuciel, Manal T. Hussein, and Ramy K. A. Sayed. 2023. "Main Components of Fish Immunity: An Overview of the Fish Immune System" Fishes 8, no. 2: 93. https://doi.org/10.3390/fishes8020093