
Recent Findings on Spawning Patterns and Recommendations for the Fishery Management of the Southern Red Snapper—Lutjanus purpureus (Poey, 1866)—On the Amazon Continental Shelf of Brazil †

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
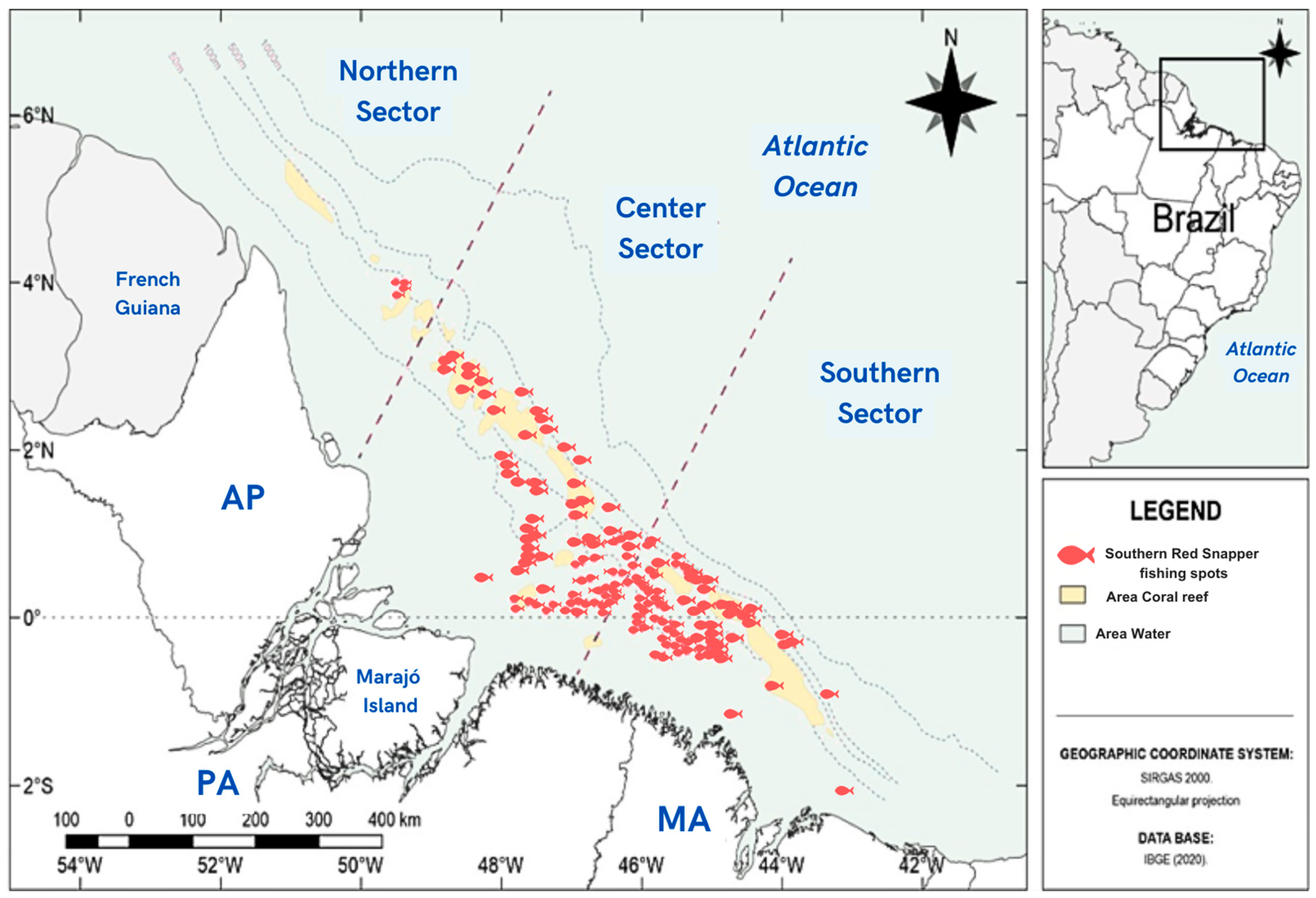
2.1. Study Area
2.2. Collection and Laboratory Screening of the Biological Samples
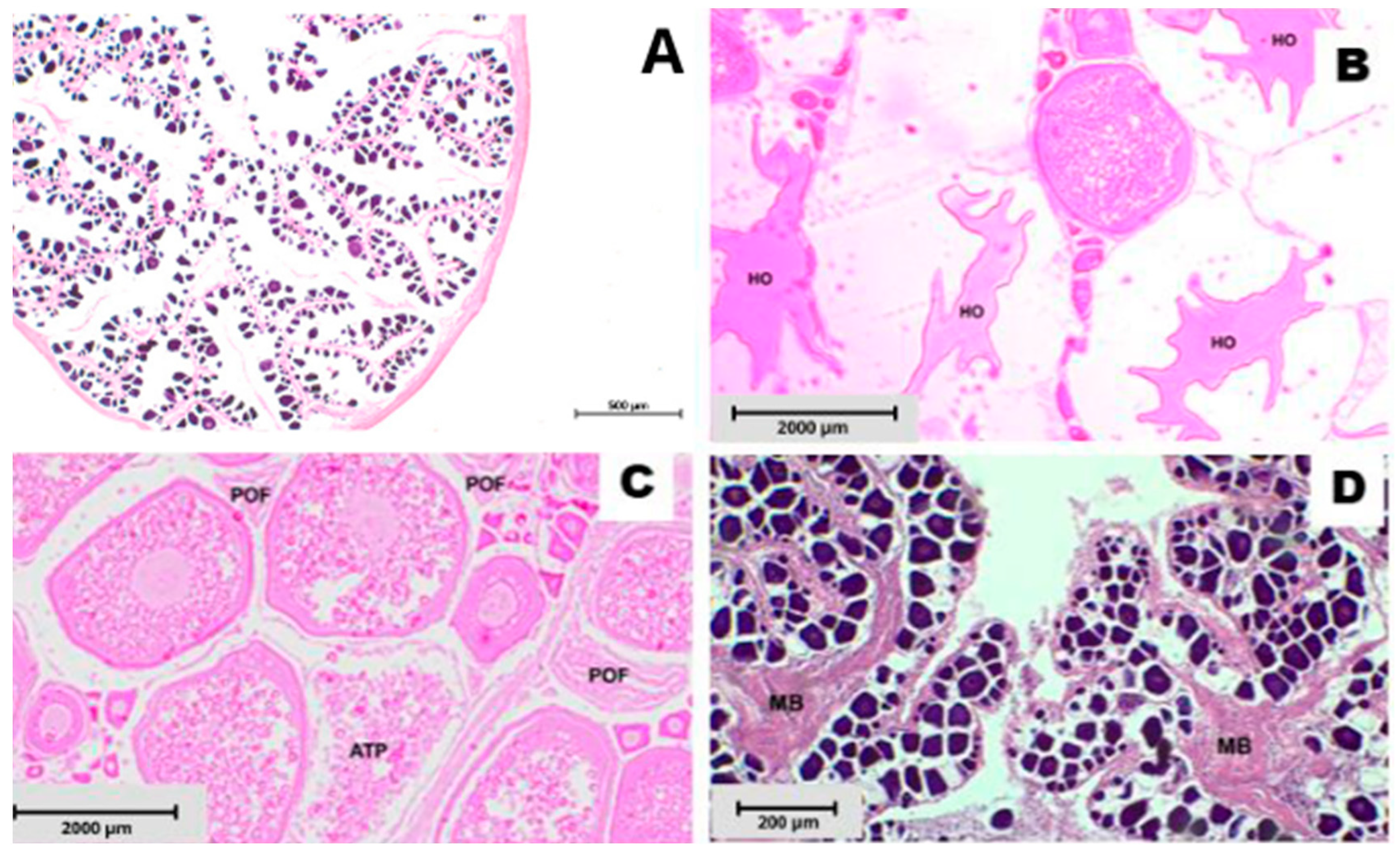
2.3. Histology and Reproductive Indicators
2.4. Data Analysis
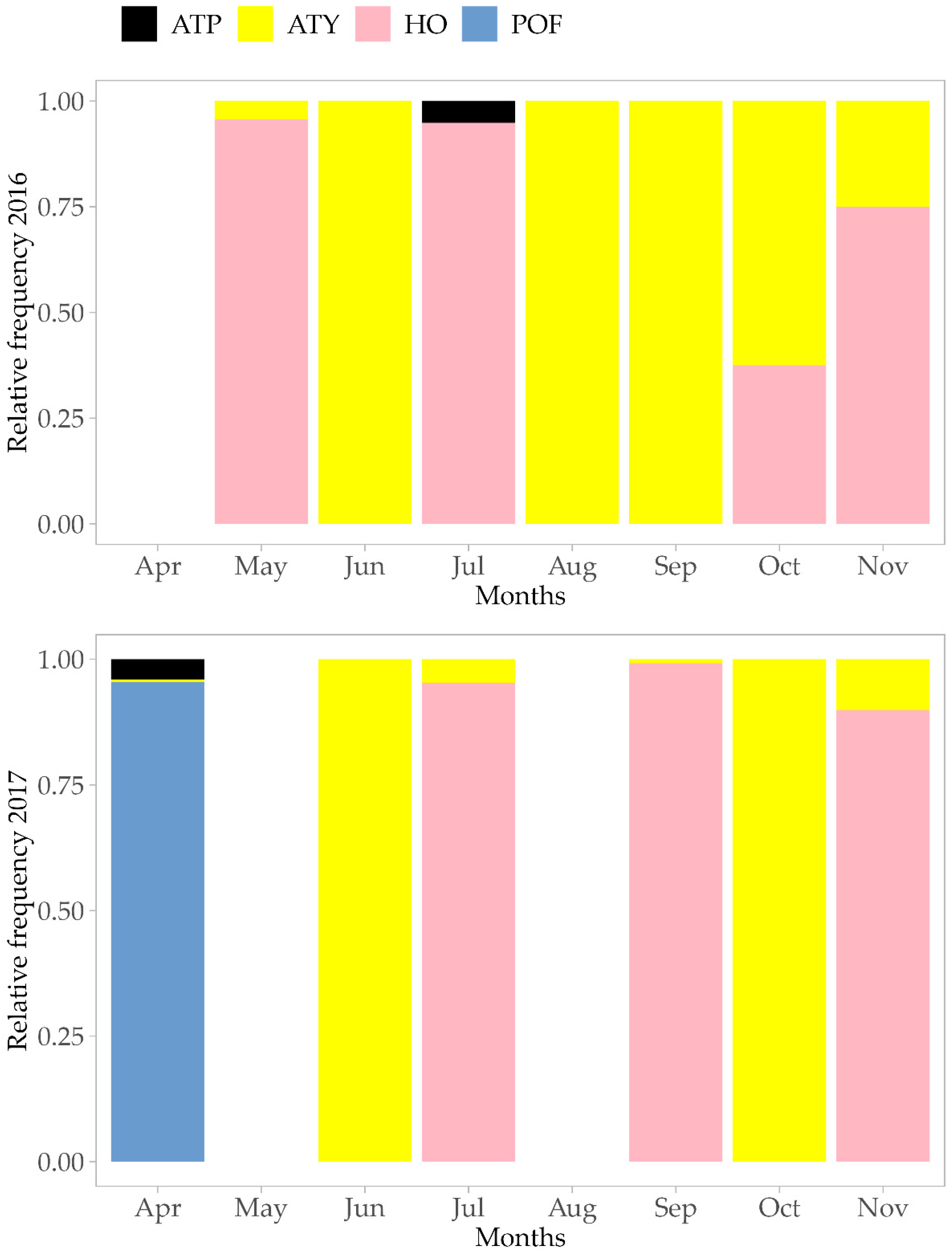
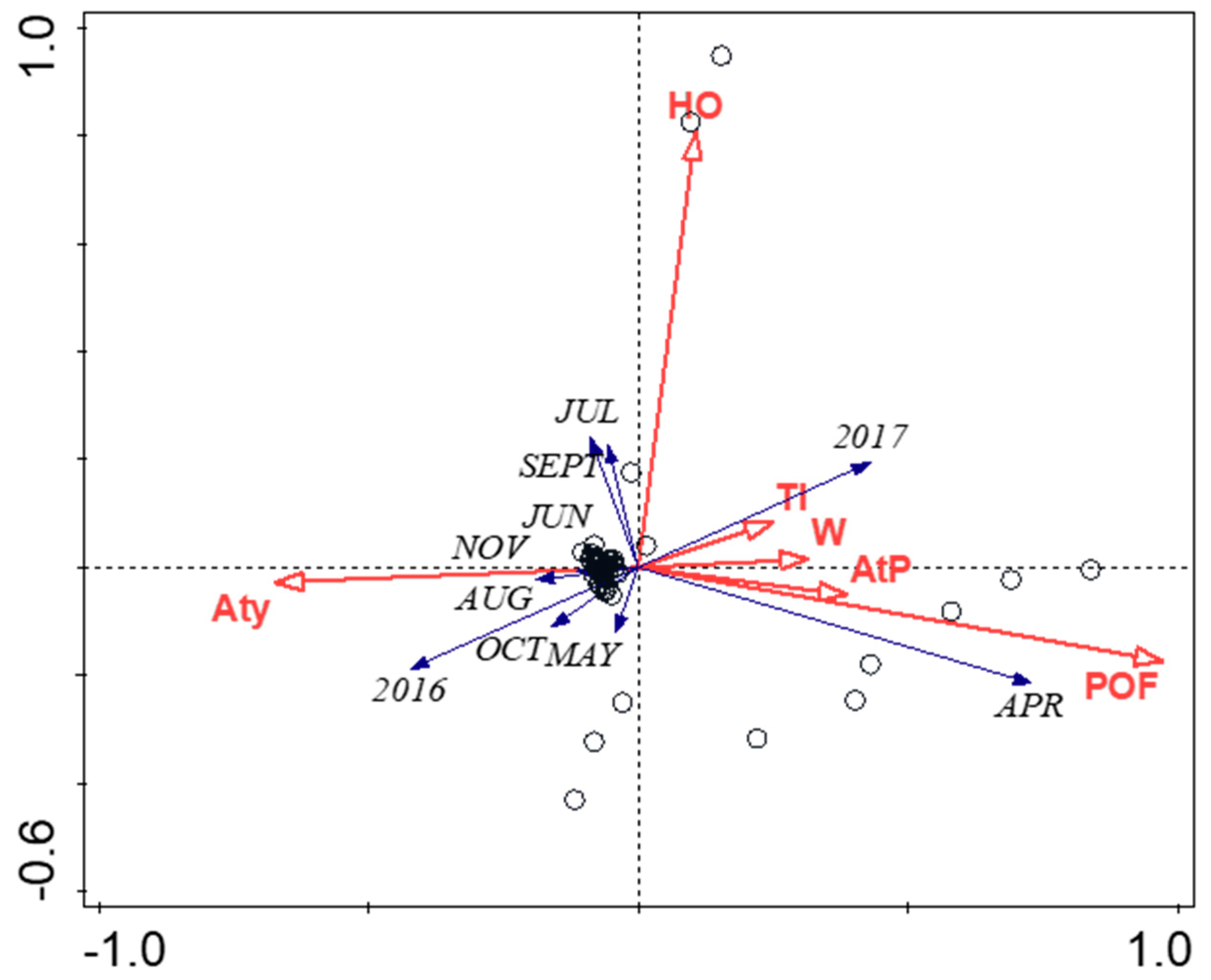
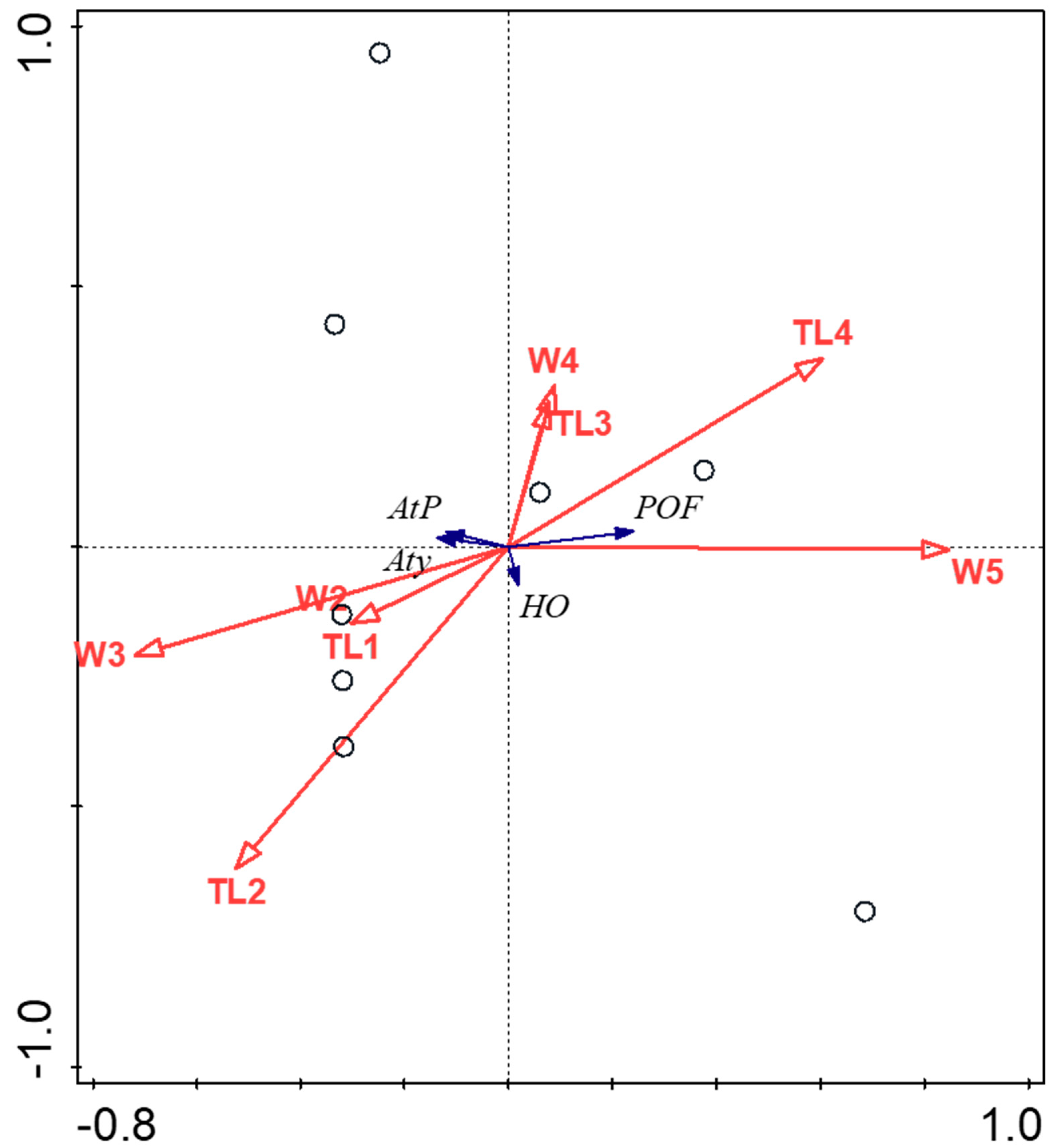
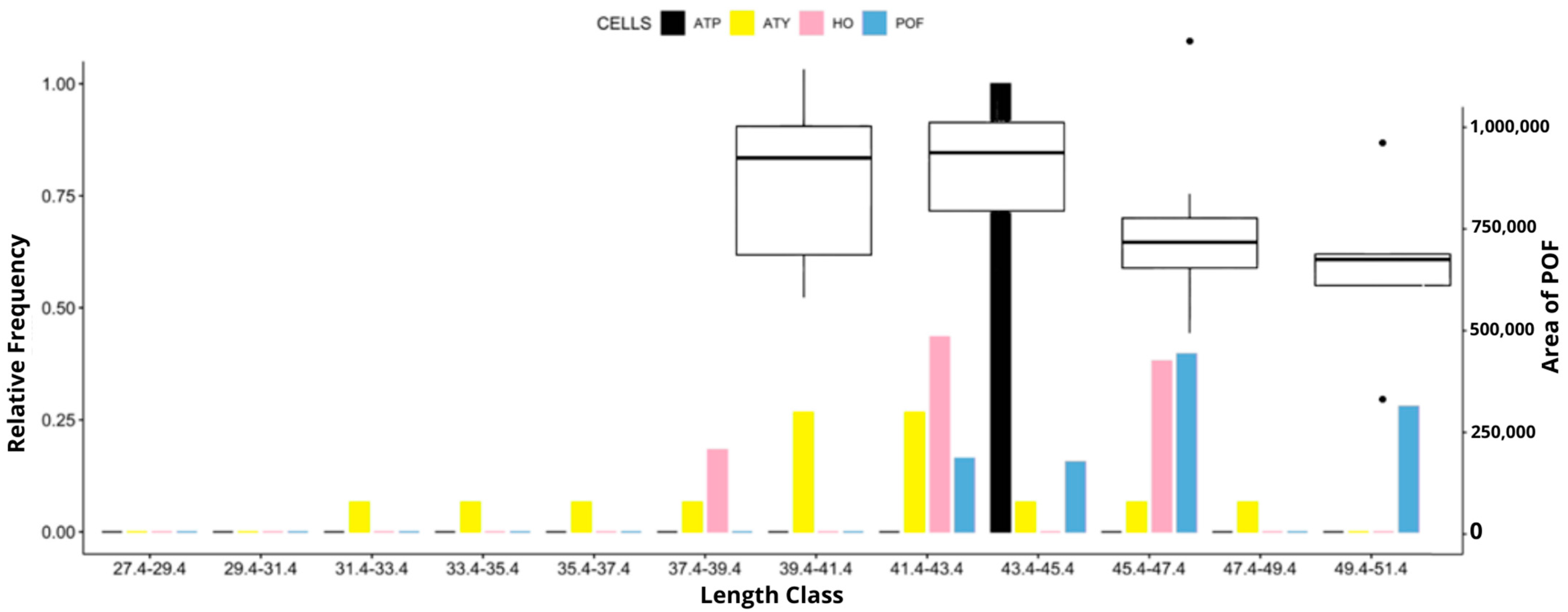
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fonteles Filho, A.A. Estudo sobre a Biologia da Pesca do Pargo, Lutjanus purpureus Poey, no Nordeste Brasileiro: Dados de 1970 e 1971. 1972. Available online: http://www.periodicos.ufc.br/arquivosdecienciadomar/article/view/32520 (accessed on 3 April 2024).
- Bentes, B.; Aragão, J.A.N.; Freire, J.L.; Lutz, Í.A.F.; Sarmento, G.C.; Gomes, T. Documento Sobre a Situação Atual das Pescarias do Pargo na Região Norte do Brasil; UFPA: Bragança, Portugal, 2017; Available online: www.fisheryprogress.org (accessed on 3 February 2021).
- Freire, J.L. Pesca, Reprodução e Crescimento de Lutjanus purpureus (Lutjanidae—Perciformes) na Costa Amazônica do Brasil. Tese de Doutorado. Doctoral Thesis, Universidade Federal do ParáUFPA, Belém, Brazil, 2019; Programa pós graduação em ecologia aquática e pesca (PPGEAP). UFPA. 2019. Tese (Doutorado em ecologia aquática e pesca) (PPGEAP).
- Martins, T.S.; Silva, R.D.F.D.; Veneza, I.B.; Santana, P.D.C.P.; Correa, R.R.; Brígida, N.C.N.S.; Gomes, G.F.E. Diversidade e abundância dos peixes comercializados no nordeste paraense, Amazônia costeira: O caso da feira livre de Bragança-PA. 2021. [Google Scholar] [CrossRef]
- Ministério da Economia. Siscomex, Sistema de Comércio Exterior. Available online: https://portalunico.siscomex.gov.br/portal/ (accessed on 22 August 2022).
- Araujo, L.S.; Magdalena, U.R.; Louzada, T.S.; Salomon, P.S.; Moraes, F.C.; Ferreira, B.P.; Paes, E.T.C.; Bastos, A.C.; Pereira, R.C.; Salgado, L.T.; et al. Growing industrialisation and poor conservation planning challenge natural resources’ management in the Amazon Shelf off Brazil. Mar. Policy 2021, 128, 104465. [Google Scholar] [CrossRef]
- Moura, R.L.; Amado-Filho, G.M.; Moraes, F.C.; Brasileiro, P.S.; Mahiques, M.M.; Bastos, A.C.; Almeida, M.G.; Silvia, J.M.; Araújo, B.F.; Brito, F.P.; et al. An extensive reef system at the Amazon River mouth. Sci. Adv. 2016, 2, e1501252. [Google Scholar] [CrossRef] [PubMed]
- Castello, J.P. Gestão sustentável dos recursos pesqueiros, isto é realmente possível. Pan-Am. J. Aquat. Sci. 2007, 2, 47–52. [Google Scholar]
- Diekert, F.K. Growth overfishing: The race to fish extends to the dimension of size. Environ. Resour. Econ. 2012, 52, 549–572. [Google Scholar] [CrossRef]
- Petersen, C.G.J. What is overfishing? J. Mar. Biol. Assoc. U. K. 1903, 6, 587–594. [Google Scholar] [CrossRef]
- Tagliarolo, M.; Cope, J.; Blanchard, F. Stock assessment on fishery-dependent data: Effect of data quality and parametrisation for a red snapper fishery. Fish. Manag. Ecol. 2021, 28, 592–603. [Google Scholar] [CrossRef]
- Benevides, E.A. Diversidade Genética, Conectividade Populacional e a Conservação do Mero (Epinephelus itajara; Percifomes Epinephelidae) na costa Atlântica da América do Sul. Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2011. [Google Scholar]
- Gesteira, T.C.; Ivo, C.T. Estudo da Reprodução e Fecundidade do Pargo, Lutjanus purpureus Poey, do norte e nordeste do Brasil. 1973. Available online: www.repositorio.ufc.br (accessed on 19 November 2020).
- Souza, R.F.; Ivo, C.T.; Souza, R.A. Aspectos da Reprodução do pargo, Lutjanus purpureus POEY, 1875 na costa Norte do Brasil. Bol Téc Cient CEPNOR 2003, 3, 107–121. [Google Scholar]
- Almeida, N.U.M. Estudos Preliminares sobre a primeira maturação sexual, época de desova e “sex-ratio” do pargo (Lutjanus aya) na costa nordeste do Brasil. Trab. Inst. Ocean. Univ. De Recife 1964, 5, 1477-158. [Google Scholar]
- Rocha, C.A.; Ivo, C.T.; Lopes, J.A. Análise da distribuição do esforço de pesca nas áreas de captura de pargo, Lutjanus purpureus Poey, no Norte e Nordeste do Brasil. Arq. Ciências Mar 1982, 22, 73–81. [Google Scholar]
- Morgan, M.J. Integrating reproductive biology into scientific advice for fisheries management. J. Northwest Atl. Fish. Sci. 2008, 41, 37–51. [Google Scholar] [CrossRef]
- Sousa, A.F.R.; Santos, N.B.; Neta, R.N.F.C.; Almeida, Z.D.S. Aspectos Reprodutivos Do Peixe Lutjanus synagris (Perciformes, Lutjanidae) Capturado Na Costa Nordeste Do Brasil. Rev. Bras. Eng. Pesca 2017, 10, 106–120. [Google Scholar] [CrossRef]
- Costa, E.F.S.; Dias, J.F.; Murua, H. Diplectrum radiale (Quoy & Gaimard, 1824) (Serranidae): A rare case of simultaneous hermaphroditism in a teleost fish. Acta Zool. 2018, 99, 244–257. [Google Scholar]
- Davis, T.L.O.; West, G.J. Maturation, Reproductive Seasonality, Fecundity, and Spawning Frequency in Lutjanus vittus (Quoy and Gaimard) from the North West Shelf of Australia. Fish. Bull. US 1993, 91, 224–236. [Google Scholar]
- De Santana, I. Ecologia Reprodutiva de Lutjanidae No Litoral Norte da Bahia, Brasil: Contribuições ao Manejo Sustentável da Pesca Artesanal. Doctoral Dissertation, Universidade de Vigo, Vigo, Spain, 2016. [Google Scholar]
- Dickerson, T.L.; Macewicz, B.J.; Hunter, J.R. Spawning frequency and batch fecundity of chub mackerel, Scomber japonicus, during 1985. CalCOFI Rep. 1992, 33, 130–140. [Google Scholar]
- Ganias, K.; Nunes, C.; Stratoudakis, Y. Degeneration of Postovulatory Follicles in the Iberian Sardine Sardina Pilchardus: Structural Changes and Factors Affecting Resorption. Fish. Bull. 2007, 105, 131–139. [Google Scholar]
- Ganias, K. Thirty years of using the postovulatory follicles method: Overview, problems and alternatives. Fish. Res. 2012, 117, 63–74. [Google Scholar] [CrossRef]
- Quagio-Grassiotto, I.; Wildner, D.D.; Ishiba, R. Gametogênese em peixes: Aspectos relevantes para o manejo reprodutivo. Rev. Bras. Reprodução Anim. 2013, 37, 181–191. [Google Scholar]
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; Macewicz, B.J.; Lowerre-Barbieri, S.K. A standardised terminology for describing reproductive development in fishes. Mar. Coast. Fish. 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Guaivira, D.A. Biologia Reprodutiva, Crescimento E Mortalidade. Tese de Doutorado. Instituto de Pesca. 2010. Available online: www.pesca.sp.gov.br (accessed on 1 April 2024).
- Romagosa, E.; Batlouni, S.R.; Borella, M.I.; Leonardo, A.F.G. Involução de folículos pós-ovulatórios em Pseudoplatystoma fasciatum (Pisces, Teleostei). Bol. Inst. Pesca 2005, 31, 129–135. [Google Scholar]
- Chaves, P.d.T.d.C.; Magalhaes, C. The oocyte development of Macrobrachium amazonicum (Heller, 1862), a freshwater shrimp of the Amazon Region (Crustacea: Decapoda: Palaemonidae). Acta Amaz. 1993, 23, 17–23. [Google Scholar] [CrossRef]
- Teixeira, S.F.; Ferreira, B.P.; Padovan, I.P. Aspects of fishing and reproduction of the black grouper Mycteroperca bonaci (Poey, 1860) (Serranidae: Epinephelinae) in the Northeastern Brazil. Neotrop. Ichthyol. 2004, 2, 19–30. [Google Scholar] [CrossRef]
- Gualberto, L.P.S.; EL-Robrini, M. Faciologia da cobertura sedimentar superficial da plataforma continental do Maranhão. Estud. Geológicos 2005, 15, 234–243. [Google Scholar]
- Silva, A.C.; Santos, M.D.; Araujo, M.C.; Bourlès, B. Observações hidrológicas e resultados de modelagem no espalhamento sazonal e espacial da pluma de água Amazônica. Acta Amaz. 2009, 39, 361–369. [Google Scholar] [CrossRef]
- Francini-Filho, R.B.; Asp, N.E.; Siegle, E.; Hocevar, J.; Lowyck, K.; D’Avila, N.; Vasconcelos, A.A.; Baitelo, R.; Rezende, C.E.; Omachi, C.Y.; et al. Perspectives on the Great Amazon Reef: Extension, biodiversity, and threats. Front. Mar. Sci. 2018, 5, 142. [Google Scholar] [CrossRef]
- Anderson, W.D.L. Volume 3: Bony fishes part 2 (Opistognathidae to Molidae), sea turtles and marine mammals. In The Living Marine Resources of the Western Central Atlantic, 1st ed.; Carpenter, K.E., Ed.; FAO: Rome, Italy, 2002; pp. 1479–1504. [Google Scholar]
- Allen, G.R. Vol. 6 Snappers of the world: An annotated and illustrated catalogue of the lutjanid species known to date. In FAO Species Catalogue; Food and Agriculture Organization of the United Nations: Rome, Italy, 1985. [Google Scholar]
- Cervigón, F.; Cipriane, R.; Fischer, W.; Garibaldi, L.; Hendrickx, M.; Lemus, A.J.; Rodriquez, B. Guia de Campo de las Espécies Comerciales marinas y de Aguas Salobres de la Costa Septentrional de Sur America; Fichas FAO de Identificacion de especies para los fines de la pesca; FAO: Rome, Italy, 1992; p. 350. [Google Scholar]
- Alves, M.I.M. Sobre a maturação sexual do pargo, Lutjanus purpureus Poey, do Nordeste Brasileiro. 1971. [Google Scholar]
- Vazzoler, A.E.A.M. Biologia da Reprodução de Peixes Teleósteos: Teoria e Prática; Eduem: Maringá, Brazil, 1996; Volume 169. [Google Scholar]
- West, G. Methods of assessing ovarian development in fishes: A Review. Aust. J. Mar. Freshwater Res. 1990, 41, 199–222. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.K.; Ganias, K.; Saborido-Rey, F.; Murua, H.; Hunter, J.R. Reproductive timing in marine fishes: Variability, temporal scales, and methods. Mar. Coast. Fish. 2011, 3, 71–91. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; W.H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Oksanen, J. “Package ‘vegan’.” Community Ecology Package, version 2.9; 2013; pp. 1–295. Available online: http://vegan.r-forger.r-project.org/ (accessed on 1 April 2024).
- Mangiafico, S. _rcompanion: Functions to Support Extension Education Program Evaluation. R Package Version 2.4.18. 2022. Available online: https://CRAN.R-project.org/package=rcompanion (accessed on 12 November 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 7 June 2022).
- Brasil, Portaria Sudepe N° 10, De 09 De Abril De 1984. Define o Tamanho Mínimo de Captura Para Pargo (Lutjanus purpureus), na Área Compreendida Entre o Norte do Amapá até a Divisa do Estado de Alagoas e Sergipe (Foz do Rio São Francisco). Available online: http://www.ibama.gov.br/component/legislacao/?view=legislacao&legislacao=91034 (accessed on 20 June 2022).
- Lucena-Frédou, F.; Eduardo, L.N.; Lira, A.S.; Pelage, L.; Passarone, R.; Frédou, T. Atividade pesqueira artesanal no nordeste do Brasil. In DO MAR; 2021; p. 374. Available online: http://www.researchgate.net (accessed on 1 April 2024).
- Oceana, Auditoria da Pesca Brasil. 2020. Available online: https://brasil.oceana.org/relatorios/auditoria-da-pesca-brasil-2020/ (accessed on 1 April 2024).
- Fonteles Filho, Antonio Adauto. Estudo sobre a biologia da Pesca do pargo, Lutjanus purpureus Poey, no Nordeste brasileiro: Dados de 1969. Arquivos ciência do mar.1. Available online: https://www.repositorio.ufc.br/handle/riufc/2003 (accessed on 8 April 2024).
- Ivo, C.T.C.; Hanson, A.J. Aspectos da biologia e dinâmica populacional do pargo Lutjanus purpureus Poey, no Norte e Nordeste do Brasil. Arq. Ciências Mar (UFCE Fortaleza) 1982, 22, 1–41. [Google Scholar]
- Freire, J.L.; Sarmento, G.C.; Lutz, Í.; Bentes, B.; Isaac, V.J. New insight into the reproductive biology and catch of juveniles of the Lutjanus purpureus in a portion of the Great Amazon reef system off the Northern Brazilian Coast. Front. Mar. Sci. 2022, 9, 804648. [Google Scholar] [CrossRef]
- IUCN. 2021. The IUCN Red List of Threatened Species. Version 2021-3. Available online: https://www.iucnredlist.org (accessed on 8 July 2022).
- Meador, M.R.; Brown, L.M. Life history strategies of fish species and biodiversity in eastern USA streams. Environ. Biol. Fishes 2015, 98, 663–677. [Google Scholar] [CrossRef]
- Stevenson, I.R.; Bryant, D.M. Climate change and constraints on breeding. Nature 2000, 406, 366–367. [Google Scholar] [CrossRef] [PubMed]
- Krabbenhoft, T.J.; Platania, S.P.; Turner, T.F. Interannual variation in reproductive phenology in a riverine fish assemblage: Implications for predicting the effects of climate change and altered flow regimes. Freshw. Biol. 2014, 59, 1744–1754. [Google Scholar] [CrossRef]
- Tao, J.; He, D.; Kennard, M.J.; Ding, C.; Bunn, S.E.; Liu, C.; Chen, Y. Strong evidence for changing fish reproductive phenology under climate warming on the Tibetan Plateau. Glob. Change Biol. 2018, 24, 2093–2104. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fu, H.; Wang, L.; Lin, L.; He, G.; Fu, P.; Kang, B. Fishery Status and Rebuilding of Major Economic Fishes in the Largest Freshwater Lake in China Based on Limited Data. Fishes 2022, 7, 47. [Google Scholar] [CrossRef]
- Trindade, D.G.; de Almeida de Gouveia, N.; da Mescouto NL, D.C.; de Sousa Moura HT, G.; da Silva UI, P.; Bentes, B. Effects of marine climate variability on the relative abundance of Lutjanus purpureus (POEY, 1866) on the Amazon Continental Shelf. Fish. Oceanogr. 2024, 33, e12662. [Google Scholar] [CrossRef]
- Mescouto, N.; Peixoto, U.I.; Trindade, D.G.; Moura, H.; Bentes, B. Caribean red snapper fishing performance indicators in Brazilian Amazon shelf: Is it the beginning of the end of a fishing system? PLoS ONE, 2024; in press. [Google Scholar]
- Perissi, I.; Bard, U.; El Asmar, T.; Lavacchi, A. Dynamic patterns of overexploitation in fisheries. Ecol. Model. 2017, 359, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Lotze, H.K.; Worm, B. Historical baselines for large marine animals. Trends Ecol. Evol. 2009, 24, 254–262. [Google Scholar] [CrossRef]
- Bayley, K.N. Factors Affecting Reproductive Investment by a Burying Beetle (Nicrophorus orbicollis). Doctoral Dissertation, Oklahoma State University, Stillwater, OK, USA, 2016. [Google Scholar]
- Núñez, J.; Duponchelle, F. Towards a universal scale to assess sexual maturation and related life history traits in oviparous teleost fishes. Fish Physiol. Biochem. 2009, 35, 167–180. [Google Scholar] [CrossRef]
- Miranda, A.C.L.; Bazzoli, N.; Rizzo, E.; Sato, Y. Ovarian follicular atresia in two teleost species: A histological and ultrastructural study. Tissue Cell 1999, 31, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Mcdonough, C.J.; Roumillat, W.A.; Wenner, C.A. Sexual differentiation and gonad development in striped mullet (Mugil cephalus L.) from South Carolina estuaries. Fish. Bull. 2005, 103, 601–619. [Google Scholar]
- González, L.W.; Lugo, T. Ovogenesis of Lutjanus purpureus Poey, 1867 (Pisces: Lutjanidae) in eastern Venezuela. Boletín Investig. Mar. Y Costeras-INVEMAR 1997, 26, 53–56. [Google Scholar]
- Brown-Peterson, N.J.; Burns, K.M.; Overstreet, R.M. Regional Differences in Florida Red Snapper Reproduction. 2009. Available online: http://aquadocs.org (accessed on 1 April 2024).
- Brulé, T.; Colás-marrufo, T.; Pérez-Díaz, E.; Sámano-Zapata, J.C. Red snapper reproductive biology in the southern Gulf of Mexico. Trans. Am. Fish. Soc. 2010, 139, 957–968. [Google Scholar] [CrossRef]
- Kulaw, D.H.; Cowan, J.H., Jr.; Jackson, M.W. Temporal and spatial comparisons of the reproductive biology of northern Gulf of Mexico (USA) red snapper (Lutjanus campechanus) collected a decade apart. PLoS ONE 2017, 12, e0172360. [Google Scholar] [CrossRef]
- Freitas, M.O.; Rocha, G.R.A.; Chaves, P.D.T.D.C.; De Moura, R.L. Reproductive biology of the lane snapper, Lutjanus synagris, and recommendations for its management on the Abrolhos Shelf, Brazil. J. Mar. Biol. Assoc. U. K. 2014, 94, 1711–1720. [Google Scholar] [CrossRef]
- Santamaría-Miranda, A.; Elorduy-Garay, J.F.; Villalejo-Fuerte, M.; Rojas-Herrera, A.A. Gonadal development and reproductive cycle of Lutjanus peru (Pisces: Lutjanidae) in Guerrero, Mexico. Rev. De Biol. Trop. 2003, 51, 489–501. [Google Scholar]
- Arellano-Martínez, M.; Rojas-Herrera, A.; García-Domínguez, F.; Ceballos-Vázquez, B.P.; Villalejo-Fuerte, M. Reproductive cycle of the spotted rose snapper Lutjanus guttatus (Steindachner, 1869) in the Guerrero coast, Mexico. Rev. De Biol. Mar. Y Oceanogr. 2001, 36, 1–8. [Google Scholar]
- Fernandes, C.A.F.; De Oliveira, P.G.; Travassos, P.E.; Hazin, F.H. Reproduction of the Brazilian snapper, Lutjanus alexandrei Moura & Lindeman, 2007 (Perciformes: Lutjanidae), off the northern coast of Pernambuco, Brazil. Neotrop. Ichthyol. 2012, 10, 587–592. [Google Scholar]
- Malafaia, P.N.; França, A.R.; Olavo, G. Spawning aggregation sites of the cubera snapper, Lutjanus cyanopterus, on the continental shelf of Bahia state, Northeastern Brazil. Fish. Res. 2021, 242, 106037. [Google Scholar] [CrossRef]
- Motta, F.S.; Freitas, M.O.; Rolim, F.A.; Abihoa, V.; Pereira, F.G.H. Direct evidence of a spawning aggregation of cubera snapper (Lutjanus cyanopterus) in southeastern Brazil and its management implications. Fish. Res. 2022, 252, 106339. [Google Scholar] [CrossRef]
- França, A.R.; Olavo, G. Indirect signals of spawning aggregations of three commercial reef fish species on the continental shelf of Bahia, east coast of Brazil. Braz. J. Oceanogr. 2015, 63, 289–301. [Google Scholar] [CrossRef]
- Nanninga, G.B.; Spaet, J.L.Y. Spawning aggregations of the Humpback red snapper, Lutjanus gibbus, in the Tuamotus, French Polynesia. Mar. Biodivers. 2017, 47, 375–376. [Google Scholar] [CrossRef]
- Johannes, R.E. Reproductive strategies of coastal marine fishes in the tropics. Environ. Biol. Fishes 1978, 3, 65–84. [Google Scholar] [CrossRef]
- Grimes, C.B. Reproductive Biology of the Lutjanidae: A Review; Westview Press: Boulder, CO, USA, 1987. [Google Scholar]
- Claro, R.; Lindeman, K.C. Spawning aggregation sites of snapper and grouper species (Lutjanidae and Serranidae) on the insular shelf of Cuba. Gulf Caribb. Res. 2003, 14, 91–106. [Google Scholar] [CrossRef]
- Fry, G.; Milton, D.A.; Van Der Velde, T.; Stobutzki, I.; Andamari, R.; Badrudin; Sumiono, B. Reproductive dynamics and nursery habitat preferences of two commercially important Indo-Pacific red snappers Lutjanus erythropterus and L. malabaricus. Fish. Sci. 2009, 75, 145–158. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.; Cabtree, L.; Switzer, T.; Burnsed, S.W.; Guenther, C. Assessing reproductive resilience: An example with South Atlantic red snapper Lutjanus campechanus. Mar. Ecol. Prog. Ser. 2015, 526, 125–141. [Google Scholar] [CrossRef]
- Jara, H.J.; Tam, J.; Reguero, B.G.; Ganoza, F.; Castillo, G.; Romero, C.Y.; Gévaudan, M.; Sánchez, A.A. Current and future socio-ecological vulnerability and adaptation of artisanal fisheries communities in Peru, the case of the Huaura province. Mar. Policy 2020, 119, 104003. [Google Scholar] [CrossRef]
- Heins, D.C.; Brown-Peterson, N.J. The reproductive biology of small fishes and the clutch concept: Combining macroscopic and histological approaches. Aquac. Fish Fish. 2022, 2, 253–264. [Google Scholar] [CrossRef]
- Honji, R.M.; Araújo, B.C.; Caneppele, D.; Nostro, F.L.; Moreira, R.G. Dynamics of ovarian maturation during the annual reproductive cycle of the endangered fish Steindachneridion parahybae (Siluriformes: Pimelodidae) in captivity. J. Fish Biol. 2022, 101, 55–68. [Google Scholar] [CrossRef]
- Constante-Pérez, L.G.; Castillo-Rivera, M.; Serrato-Díaz, A. Diversity, seasonal and diel distribution of snappers (Lutjanidae: Perciformes) in a tropical coastal inlet in the southwestern Gulf of Mexico. Neotrop. Ichthyol. 2022, 20, e220009. [Google Scholar] [CrossRef]
- Falfan, E.L.; Ordóñez, U.R.; Ornelas, M.A. Abundancia y Distribución de Larvas del Complejo Lutjanidae–Serranidae en la Plataforma de Yucatán. 2007. Available online: https://aquadocs.org/bitstream/handle/1834/28954/gcfi_58-20.pdf?sequence=1 (accessed on 8 July 2022).
- Viana, J.M.W. Pesca e Biologia Reprodutiva do Dentão(Lutjanus jocu) no Arquipélago de Fernando de Noronha. Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2018. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase | Previous Terminology | Macroscopic and Histological Features |
|---|---|---|
| Immature (never spawned) | Immature, virgin | Small ovaries, often transparent, with indistinct blood vessels. Only oogonia and PG oocytes present. No atresia or muscle bundles. Thin ovarian wall and little space between the oocytes |
| Developing (ovaries beginning to develop, but not ready to spawn) | Maturing, early developing, early maturation, mid-maturation, ripening, previtellogenic | Enlarging ovaries, blood vessels becoming more distinct. PG, CA, Vtg1, and Vtg2 oocytes present. No evidence of POFs or Vtg3 oocytes. Some atresia may be present. Only PG and CA oocytes are present in the early developing subphase. |
| Spawning capable (fish are developmentally and physiologically able to spawn) | Mature, late developing, late maturation, late ripening, total maturation, gravid, vitellogenic, ripe, partially spent, fully developed, prespawning, running ripe, final MO, spawning, gravid, ovulated | Large ovaries, blood vessels prominent. Individual oocytes visible macroscopically. Vtg3 oocytes or POFs present in batchers. Atresia of vitellogenic and/or hydrated oocytes may be present. Early stages of MO may be present. Actively spawning subphase: oocytes undergoing late GVM, GVBD, hydration or ovulation. |
| Regressing (spawning ceased) | Spent, regression, postspawning, recovering | Flaccid ovaries, blood vessels prominent. Atresia (any stage) and POFs present. Some CA and/or vitellogenic (Vtg1, Vtg2) oocytes present. |
| Regenerating (sexually mature, reproductively inactive) | Resting, regressed, recovering, inactive | Small ovaries. Blood vessels reduced, but present. Only oogonia and PG oocytes present. Muscle bundles, enlarged blood vessels, thick ovarian wall and/or gamma/delta atresia or old, degenerating POFs may be present. |
| Structure | Duration | Description |
|---|---|---|
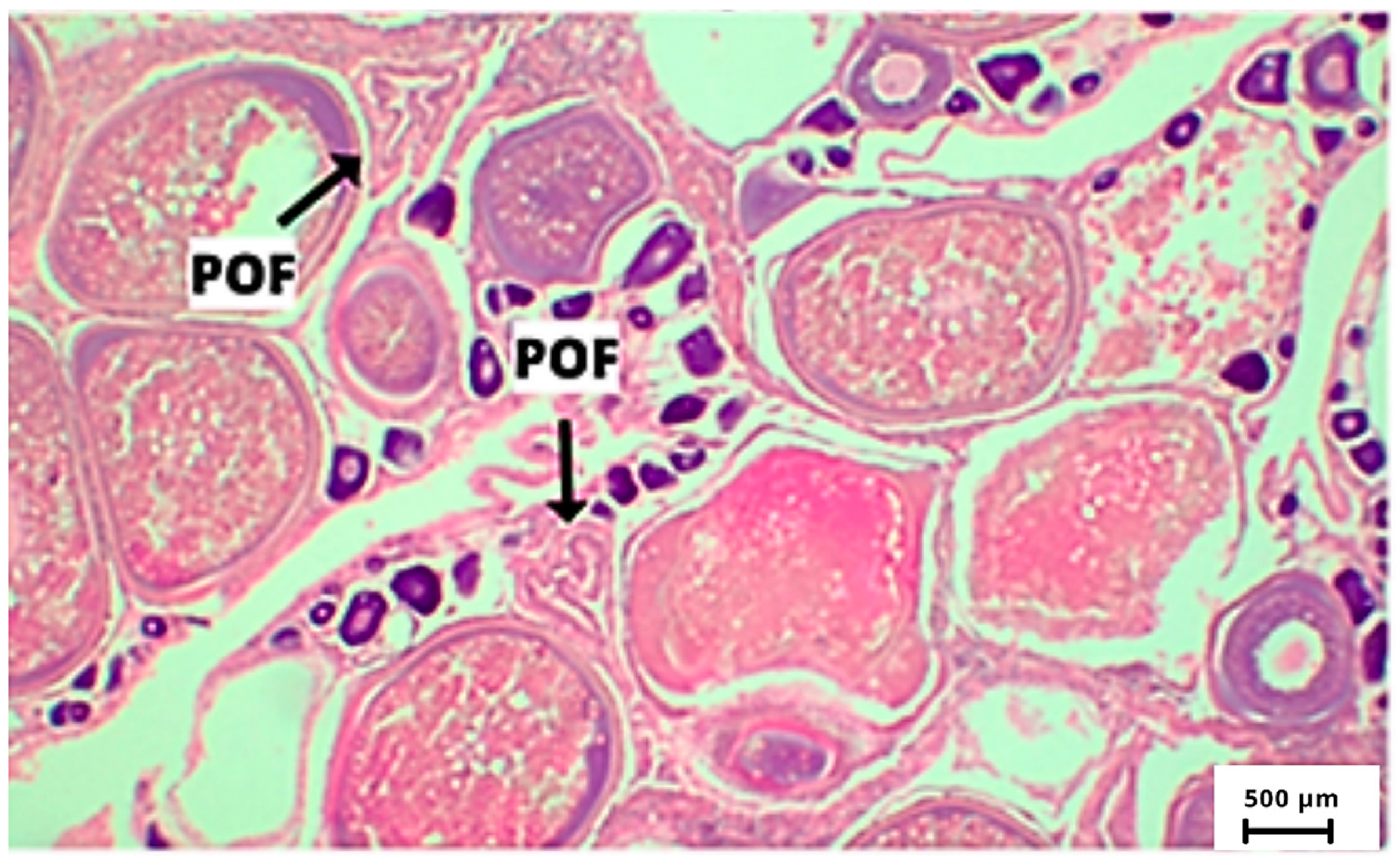
| Recent POF | 6 h | The remaining membranes (theca and granulosa) are easily distinguished from each other, with the basal membrane still intact. The follicular cells are shaped like a twisted cord. The lumen is clearly visible. |
| Intermediate POF | 12 h | Advanced degenerative process in the follicular cap, sinusoidal alignment still present, but with a considerable reduction of the lumen. Granulosa minor. |
| Old POF | 24 h | Lumen not visible, smaller follicles, absence of the sinusoidal alignment, follicular layers loosened. |
| Year | TL Class (cm) | Apr. | May | Jun. | Jul. | Aug. | Sep. | Oct. | Nov. | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 27–28 | 2 | 1 | 3 | ||||||
| 28–29 | 1 | 1 | 2 | |||||||
| 31–32 | 1 | 1 | ||||||||
| 32–33 | 1 | 1 | ||||||||
| 34–35 | 2 | 1 | 1 | 4 | ||||||
| 35–36 | 1 | 2 | 3 | |||||||
| 36–37 | 1 | 1 | ||||||||
| 37–38 | 1 | 1 | ||||||||
| 38–39 | 1 | 1 | 3 | 5 | ||||||
| 39–40 | 1 | 1 | 3 | 5 | ||||||
| 40–41 | 1 | 1 | 2 | 4 | ||||||
| 41–42 | 2 | 1 | 1 | 1 | 5 | |||||
| 42–43 | 1 | 2 | 2 | 5 | 10 | |||||
| 43–44 | 1 | 1 | 1 | 2 | 4 | 9 | ||||
| 44–45 | 1 | 5 | 1 | 7 | ||||||
| 45–46 | 2 | 1 | 3 | |||||||
| 47–48 | 1 | 1 | 1 | 1 | 4 | |||||
| 48–49 | 1 | 1 | 1 | 3 | ||||||
| 49–50 | 1 | 1 | 2 | |||||||
| 50–51 | 1 | 1 | ||||||||
| 54–55 | 1 | 1 | ||||||||
| 2017 | 27–28 | |||||||||
| 28–29 | ||||||||||
| 31–32 | ||||||||||
| 32–33 | 1 | 1 | ||||||||
| 33–34 | 1 | 1 | ||||||||
| 34–35 | 1 | 1 | ||||||||
| 35–36 | 1 | 1 | ||||||||
| 36–37 | 1 | 1 | ||||||||
| 37–38 | 2 | 2 | ||||||||
| 38–39 | 3 | 1 | 4 | |||||||
| 39–40 | 1 | 2 | 1 | 1 | 5 | |||||
| 40–41 | 1 | 1 | 1 | 3 | ||||||
| 41–42 | 1 | 1 | 2 | 2 | 1 | 7 | ||||
| 42–43 | 2 | 1 | 2 | 1 | 1 | 2 | 9 | |||
| 43–44 | 1 | 1 | 2 | |||||||
| 45–46 | 1 | 1 | 1 | 3 | ||||||
| 46–47 | 1 | 1 | ||||||||
| 47–48 | 2 | 2 | ||||||||
| 49–50 | 1 | 1 | ||||||||
| 50–51 | 1 | 1 | ||||||||
| Total | 13 | 3 | 7 | 19 | 21 | 14 | 17 | 27 | 120 |
| Dependent Variable | Factor | DF | SQ | H | p |
|---|---|---|---|---|---|
| HO | TLC | 3 | 642.1 | 31.883 | 0.3635 |
| Month | 7 | 1815.8 | 9.016 | 0.25151 | |
| TLC:Month | 17 | 5778.4 | 286.919 | 0.03748 | |
| Residual | 93 | 16,233.0 | |||
| ATY | TLC | 3 | 1562 | 17.818 | 0.6189 |
| Month | 7 | 3419 | 38.994 | 0.79129 | |
| TLC:Month | 171.243 | 14.175 | 0.65468 | 0.65438 | |
| Residual | 93 | 88,104 | |||
| POF | TLC | 3 | -- | 5.4868 | 0.1394 |
| Month | -- | -- | -- | -- | |
| TLC:Month | -- | -- | -- | -- | |
| ATP | TLC | -- | -- | -- | -- |
| Month | 7 | -- | 5.9117 | 0.5501 | |
| TLC:Month | -- | -- | -- | -- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Costa Pinheiro, L.; Ferreira, B.P.; Santos, P.E.; Peixoto, U.; Bentes, B. Recent Findings on Spawning Patterns and Recommendations for the Fishery Management of the Southern Red Snapper—Lutjanus purpureus (Poey, 1866)—On the Amazon Continental Shelf of Brazil. Fishes 2024, 9, 136. https://doi.org/10.3390/fishes9040136
da Costa Pinheiro L, Ferreira BP, Santos PE, Peixoto U, Bentes B. Recent Findings on Spawning Patterns and Recommendations for the Fishery Management of the Southern Red Snapper—Lutjanus purpureus (Poey, 1866)—On the Amazon Continental Shelf of Brazil. Fishes. 2024; 9(4):136. https://doi.org/10.3390/fishes9040136
Chicago/Turabian Styleda Costa Pinheiro, Larissa, Beatrice Padovani Ferreira, Paulo Emílio Santos, Ualerson Peixoto, and Bianca Bentes. 2024. "Recent Findings on Spawning Patterns and Recommendations for the Fishery Management of the Southern Red Snapper—Lutjanus purpureus (Poey, 1866)—On the Amazon Continental Shelf of Brazil" Fishes 9, no. 4: 136. https://doi.org/10.3390/fishes9040136