Brief Myofascial Intervention Modulates Visual Event-Related Potential Response to Emotional Photographic Contents: A Pilot Study



Abstract
:1. Introduction
1.1. The Present Study
2. Materials and Methods
2.1. Participants
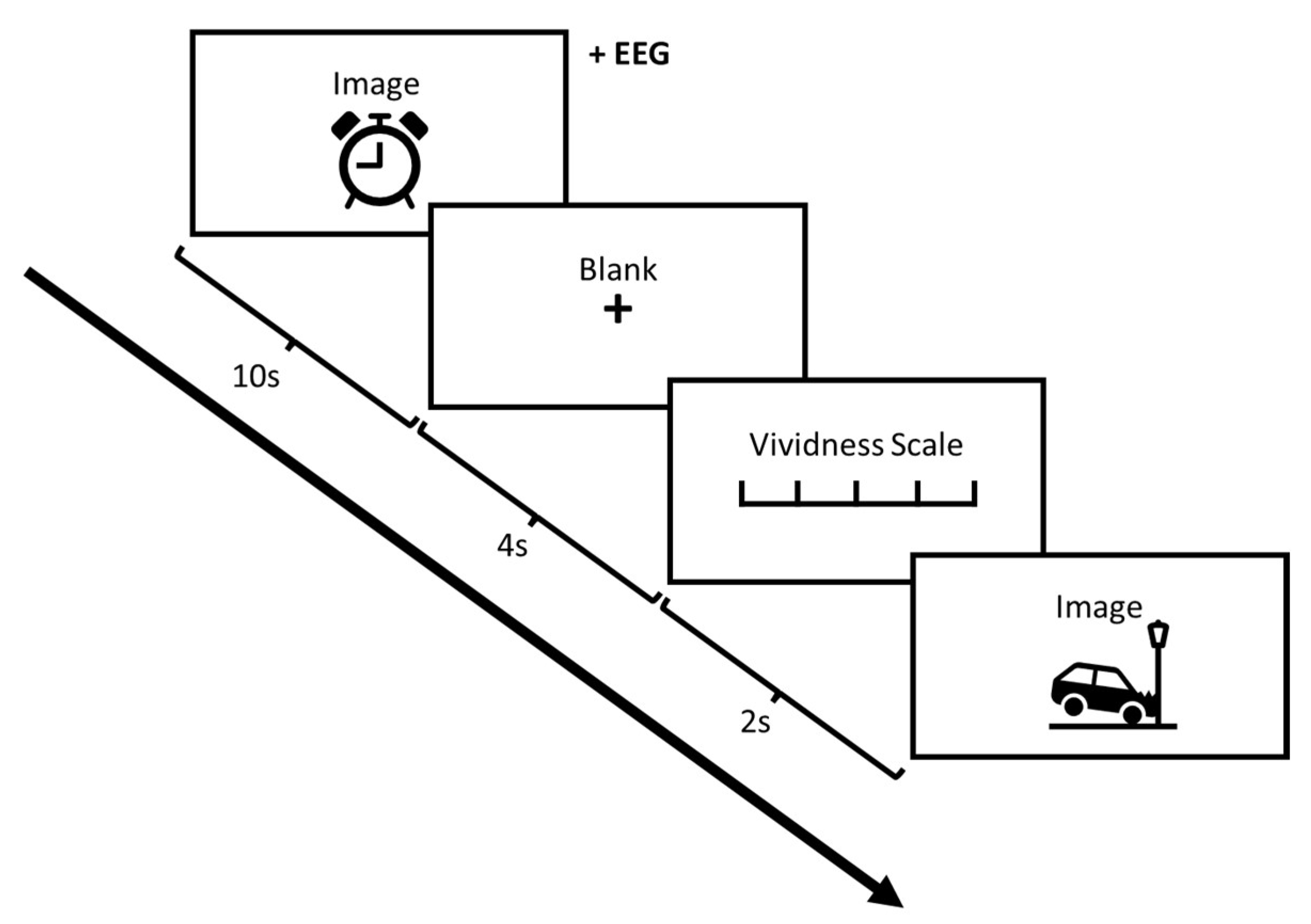
2.2. Procedure
2.3. Myofascial Intervention
2.4. Stimuli
2.5. Data Analysis
3. Results
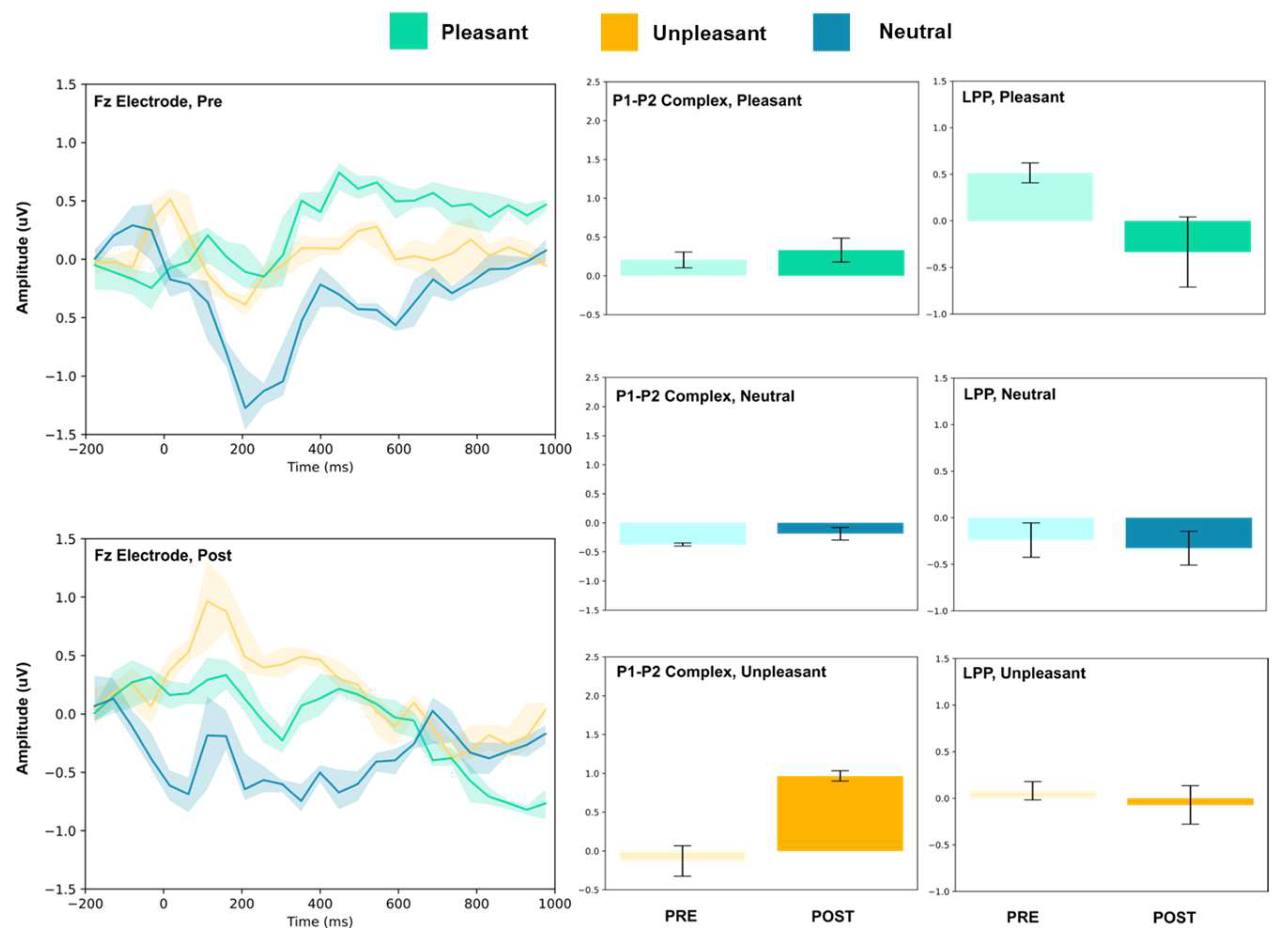
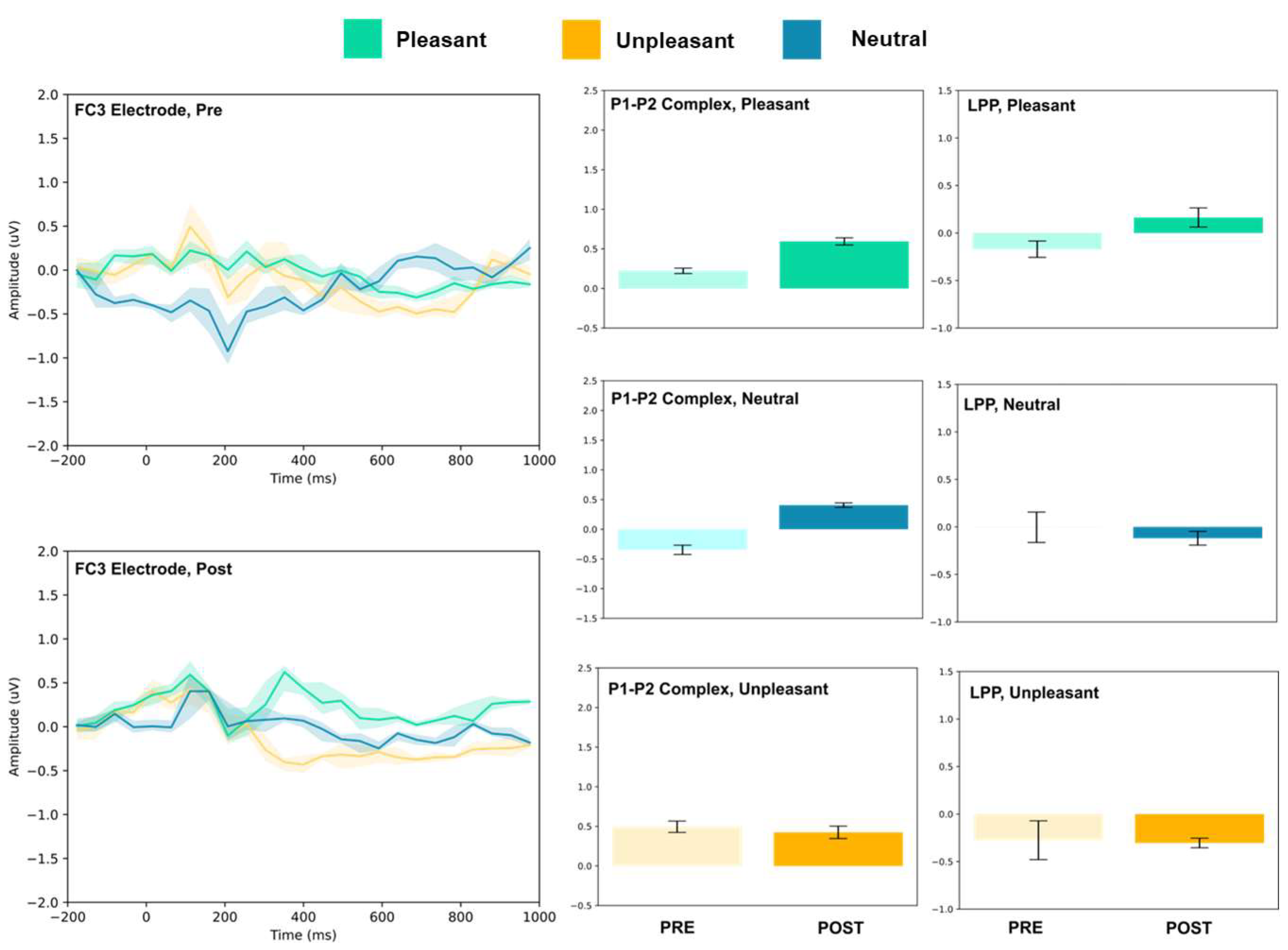
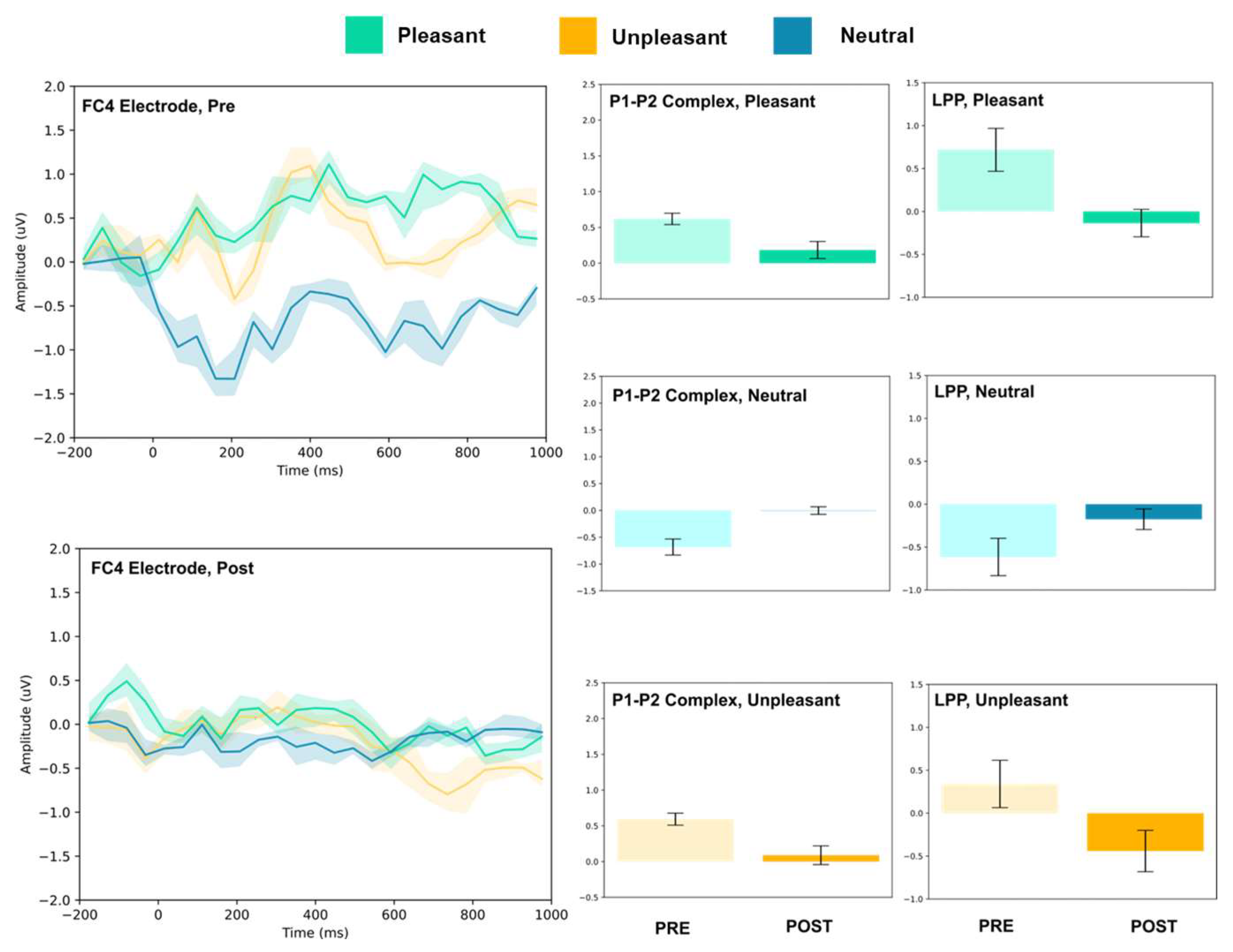
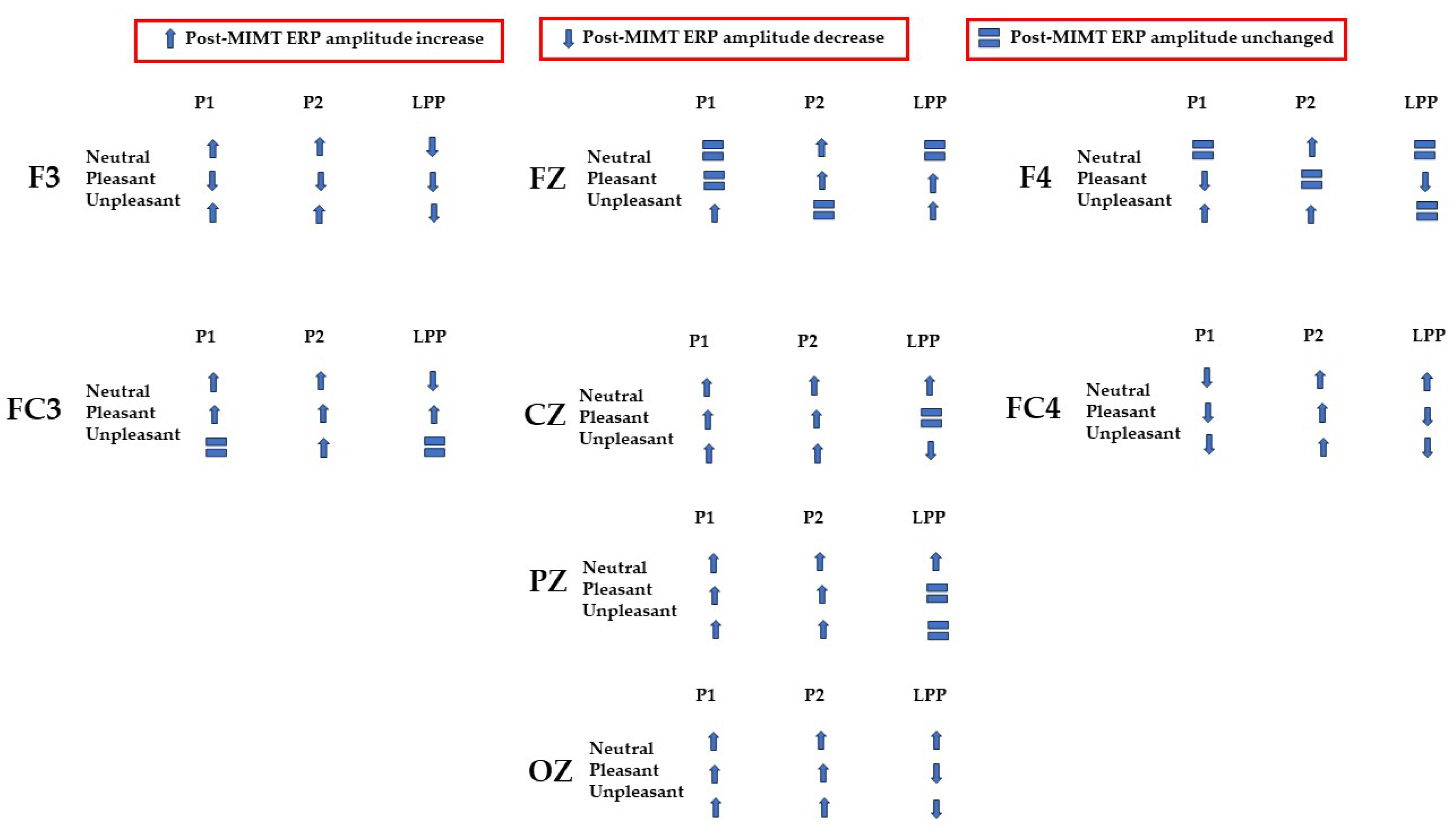
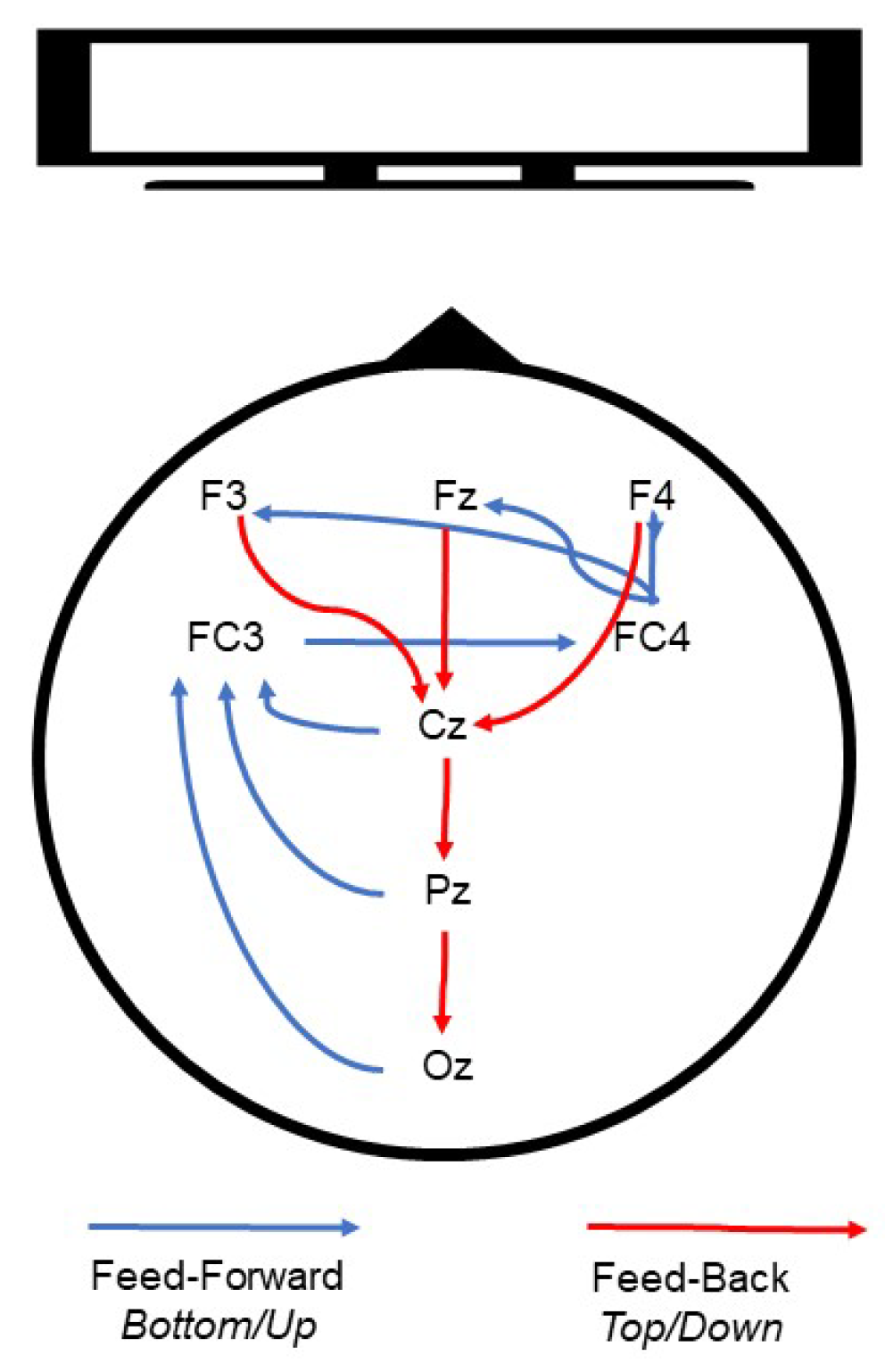
3.1. P1–P2 and LLP
3.1.1. Frontal Electrodes
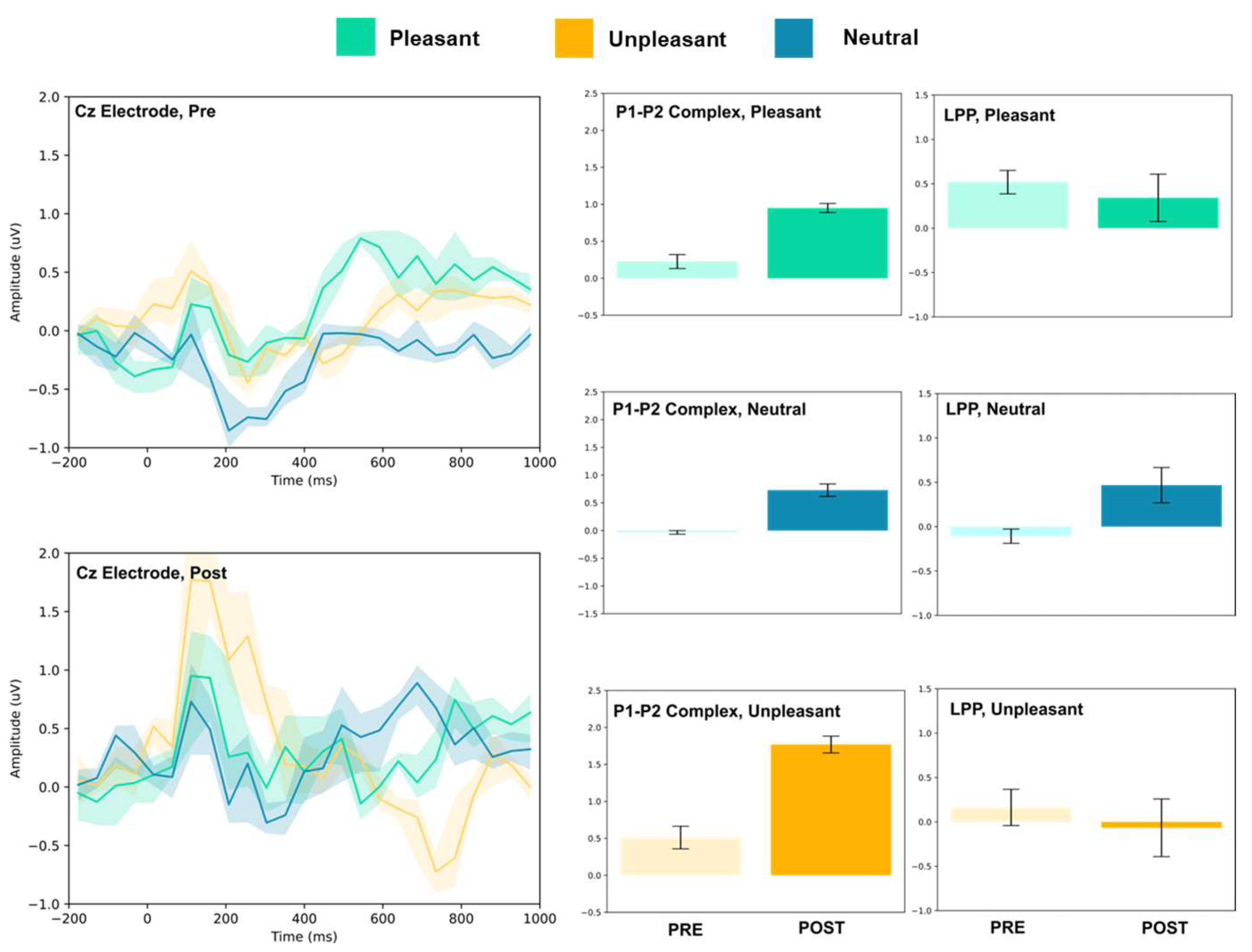
3.1.2. Frontocentral and Central Electrodes
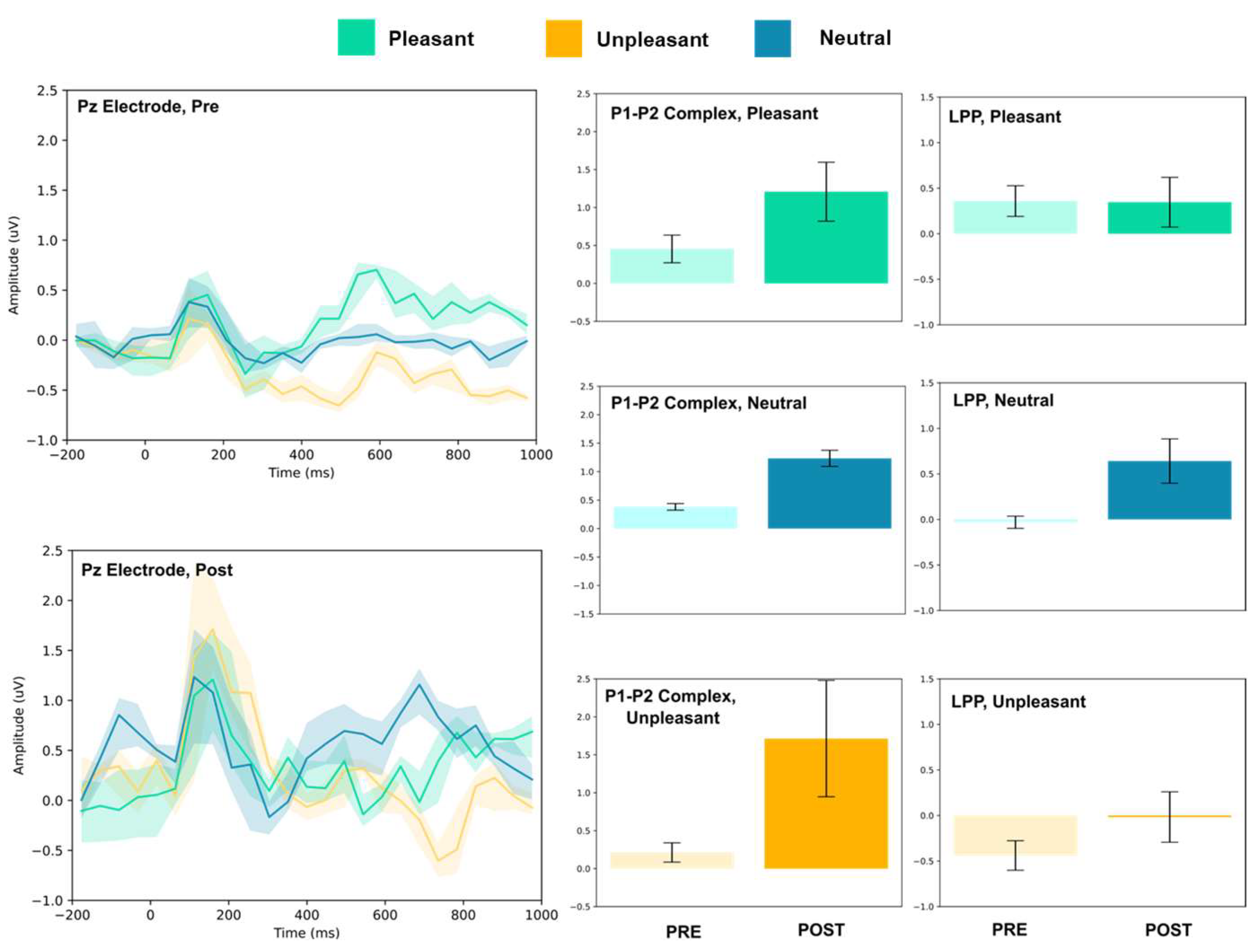
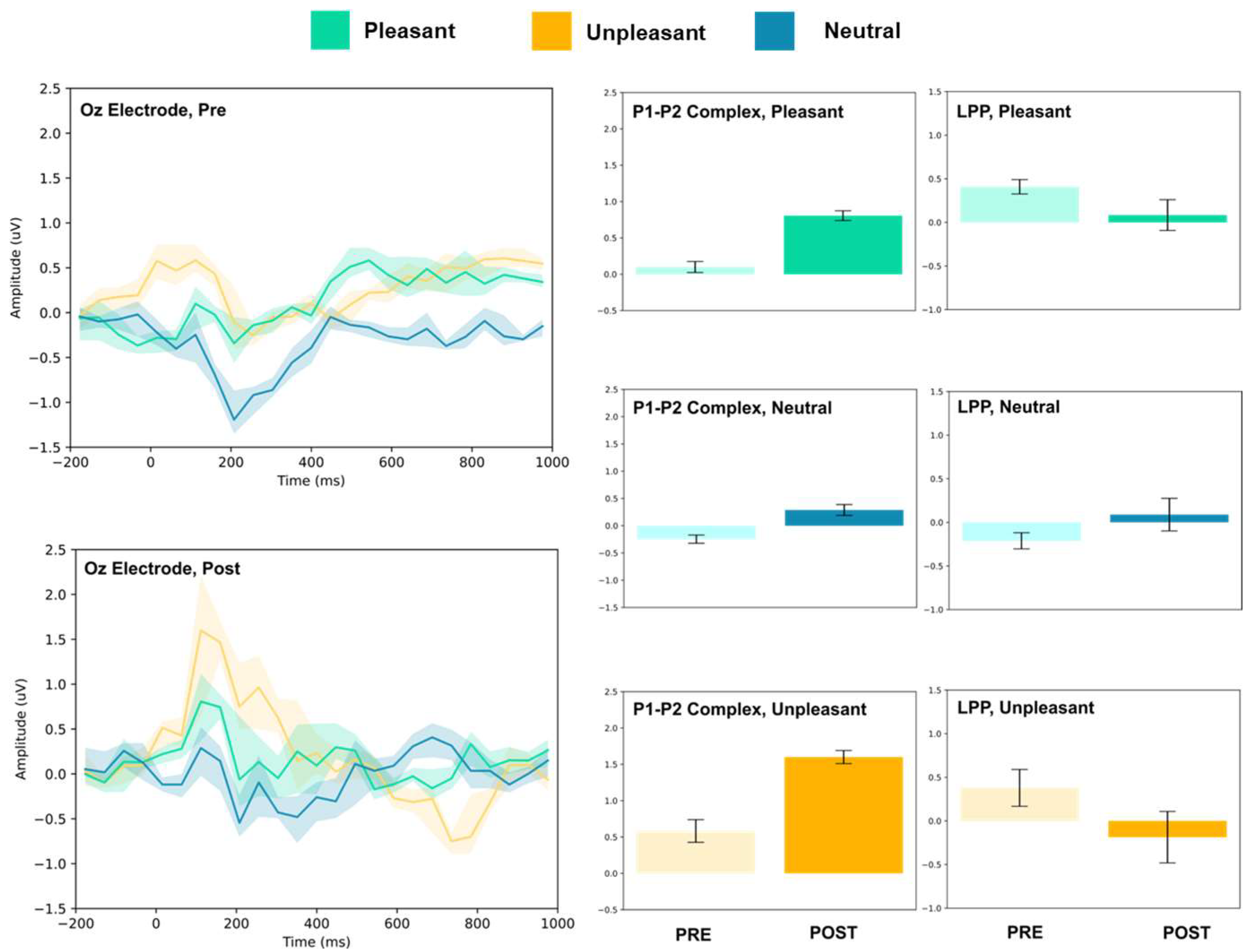
3.1.3. Parietal and Occipital Electrodes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawford, C.; Boyd, C.; Paat, C.F.; Price, A.; Xenakis, L.; Yang, E.; Zhang, W. The Impact of Massage Therapy on Function in Pain Populations—A Systematic Review and Meta-Analysis of Randomized Controlled Trials: Part I, Patients Experiencing Pain in the General Population. Pain Med. 2016, 17, 1353–1375. [Google Scholar] [CrossRef]
- Liu, C.; Chen, X.; Wu, S. The effect of massage therapy on pain after surgery: A comprehensive meta-analysis. Complement. Ther. Med. 2022, 71, 102892. [Google Scholar] [CrossRef]
- Anderson, P.G.; Cutshall, S.M. Massage Therapy: A Comfort Intervention for Cardiac Surgery Patients. Clin. Nurse Spec. 2007, 21, 161–165. [Google Scholar] [CrossRef]
- Piovesan, E.J.; Di Stani, F.; Kowacs, P.A.; Mulinari, R.A.; Radunz, V.H.; Utiumi, M.; Muranka, E.B.; Giublin, M.L.; Werneck, L.C. Massaging over the greater occipital nerve reduces the intensity of migraine attacks: Evidence for inhibitory trigemino-cervical convergence mechanisms. Arq. De Neuro-Psiquiatr. 2007, 65, 599–604. [Google Scholar] [CrossRef]
- Hart, S.; Field, T.; Hernandez-Reif, M.; Nearing, G.; Shaw, S.; Schanberg, S.; Kuhn, C. Anorexia Nervosa Symptoms are Reduced by Massage Therapy. Eat. Disord. 2001, 9, 289–299. [Google Scholar] [CrossRef]
- Kito, K.; Suzuki, K. Research on the Effect of the Foot Bath and Foot Massage on Residual Schizophrenia Patients. Arch. Psychiatr. Nurs. 2016, 30, 375–381. [Google Scholar] [CrossRef]
- Rapaport, M.; Schettler, P.; Larson, E.; Edwards, S.; Dunlop, B.; Rakofsky, J.; Kinkead, B. Acute Swedish Massage Monotherapy Successfully Remediates Symptoms of Generalized Anxiety Disorder: A Proof-of-Concept, Randomized Controlled Study. J. Clin. Psychiatry 2016, 77, e883–e891. [Google Scholar] [CrossRef]
- Diego, M.A.; Field, T. Moderate Pressure Massage Elicits a Parasympathetic Nervous System Response. Int. J. Neurosci. 2009, 119, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Sato, W. Inhibition of emotion-related autonomic arousal by skin pressure. Springerplus 2015, 4, 294. [Google Scholar] [CrossRef] [PubMed]
- Rea, P. Introduction to the Nervous System. In Clinical Anatomy of the Cranial Nerves; Rea, P., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. xv–xxix. [Google Scholar]
- Meier, M.; Unternaehrer, E.; Dimitroff, S.J.; Benz, A.B.E.; Bentele, U.U.; Schorpp, S.M.; Wenzel, M.; Pruessner, J.C. Standardized massage interventions as protocols for the induction of psychophysiological relaxation in the laboratory: A block randomized, controlled trial. Sci. Rep. 2020, 10, 14774. [Google Scholar] [CrossRef] [PubMed]
- Field, T.; Ironson, G.; Scafidi, F.; Nawrocki, T.; Goncalves, A.; Burman, I.; Pickens, J.; Fox, N.; Schanberg, S.; Kuhn, C. Massage Therapy Reduces Anxiety and Enhances Eeg Pattern of Alertness and Math Computations. Int. J. Neurosci. 1996, 86, 197–205. [Google Scholar] [CrossRef]
- Jones, N.A.; Field, T. Massage and music therapies attenuate frontal EEG asymmetry in depressed adolescents. Adolescence 1999, 34, 529–534. [Google Scholar]
- Jones, N.A.; Field, T.; Davalos, M. Massage therapy attenuates right frontal EEG asymmetry in one-month-old infants of depressed mothers. Infant Behav. Dev. 1998, 21, 527–530. [Google Scholar] [CrossRef]
- Sliz, D.; Smith, A.; Wiebking, C.; Northoff, G.; Hayley, S. Neural correlates of a single-session massage treatment. Brain Imaging Behav. 2012, 6, 77–87. [Google Scholar] [CrossRef]
- Ouchi, Y.; Kanno, T.; Okada, H.; Yoshikawa, E.; Shinke, T.; Nagasawa, S.; Minoda, K.; Doi, H. Changes in cerebral blood flow under the prone condition with and without massage. Neurosci. Lett. 2006, 407, 131–135. [Google Scholar] [CrossRef]
- Strauss, T.; Kämpe, R.; Hamilton, J.P.; Olausson, H.; Rottstädt, F.; Raue, C.; Croy, I. Deactivation of default mode network during touch. Sci. Rep. 2019, 9, 1293. [Google Scholar] [CrossRef]
- Taylor, A.G.; Goehler, L.E.; Galper, D.I.; Innes, K.E.; Bourguignon, C. Top-down and bottom-up mechanisms in mind-body medicine: Development of an integrative framework for psychophysiological research. Explore 2010, 6, 29–41. [Google Scholar] [CrossRef]
- Gilbert, C.D.; Li, W. Top-down influences on visual processing. Nat. Rev. Neurosci. 2013, 14, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Vuilleumier, P.; Driver, J. Modulation of visual processing by attention and emotion: Windows on causal interactions between human brain regions. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 837–855. [Google Scholar] [CrossRef]
- Bendall, R.C.A.; Eachus, P.; Thompson, C. A Brief Review of Research Using Near-Infrared Spectroscopy to Measure Activation of the Prefrontal Cortex during Emotional Processing: The Importance of Experimental Design. Front. Hum. Neurosci. 2016, 10, 529. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pérez, A.M.; Peralta-Ramírez, M.I.; Pilat, A.; Villaverde, C. Effects of myofascial induction techniques on physiologic and psychologic parameters: A randomized controlled trial. J. Altern. Complement. Med. 2008, 14, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Henley, C.E.; Ivins, D.; Mills, M.; Wen, F.K.; Benjamin, B.A. Osteopathic manipulative treatment and its relationship to autonomic nervous system activity as demonstrated by heart rate variability: A repeated measures study. Osteopath. Med. Prim. Care 2008, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Kaye, A.D.; Kaye, A.J.; Swinford, J.; Baluch, A.; Bawcom, B.A.; Lambert, T.J.; Hoover, J.M. The effect of deep-tissue massage therapy on blood pressure and heart rate. J. Altern. Complement. Med. 2008, 14, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Minasny, B. Understanding the process of fascial unwinding. Int. J. Ther. Massage Bodyw. 2009, 2, 10–17. [Google Scholar] [CrossRef]
- Luck, S.J.; Woodman, G.F.; Vogel, E.K. Event-related potential studies of attention. Trends Cogn. Sci. 2000, 4, 432–440. [Google Scholar] [CrossRef]
- Atchley, R.; Klee, D.; Memmott, T.; Goodrich, E.; Wahbeh, H.; Oken, B. Event-related potential correlates of mindfulness meditation competence. Neuroscience 2016, 320, 83–92. [Google Scholar] [CrossRef]
- Sharpe, L.; Perry, K.N.; Rogers, P.; Dear, B.F.; Nicholas, M.K.; Refshauge, K. A comparison of the effect of attention training and relaxation on responses to pain. Pain 2010, 150, 469–476. [Google Scholar] [CrossRef]
- Marmolejo-Ramos, F.; Hellemans, K.; Comeau, A.; Heenan, A.; Faulkner, A.; Abizaid, A.; D’Angiulli, A. Event-related potential signatures of perceived and imagined emotional and food real-life photos. Neurosci. Bull. 2015, 31, 317–330. [Google Scholar] [CrossRef]
- Cuthbert, B.N.; Schupp, H.T.; Bradley, M.M.; Birbaumer, N.; Lang, P.J. Brain potentials in affective picture processing: Covariation with autonomic arousal and affective report. Biol. Psychol. 2000, 52, 95–111. [Google Scholar] [CrossRef]
- Buchanan, D.M.; Grant, J.; D’Angiulli, A. Commercial wireless versus standard stationary EEG systems for personalized emotional brain-computer interfaces: A preliminary reliability check. Neurosci. Res. Notes 2019, 2, 7–15. [Google Scholar] [CrossRef]
- Smith, P.L.; Little, D.R. Small is beautiful: In defense of the small-N design. Psychon. Bull. Rev. 2018, 25, 2083–2101. [Google Scholar] [CrossRef] [PubMed]
- Bos, F.M.; Snippe, E.; de Vos, S.; Hartmann, J.A.; Simons, C.J.P.; van der Krieke, L.; de Jonge, P.; Wichers, M. Can We Jump from Cross-Sectional to Dynamic Interpretations of Networks Implications for the Network Perspective in Psychiatry. Psychother. Psychosom. 2017, 86, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, P.C.M. A Manifesto on Psychology as Idiographic Science: Bringing the Person Back into Scientific Psychology, This Time Forever. Meas. Interdiscip. Res. Perspect. 2004, 2, 201–218. [Google Scholar] [CrossRef]
- Molenaar, P.C. On the implications of the classical ergodic theorems: Analysis of developmental processes has to focus on intra-individual variation. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 2008, 50, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.V.; Grennan, G.; Zafar-Khan, M.; Alim, F.; Dey, S.; Ramanathan, D.; Mishra, J. Personalized machine learning of depressed mood using wearables. Transl. Psychiatry 2021, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Birkhoff, G.D. What is the ergodic theorem? Am. Math. Mon. 1942, 49, 222–226. [Google Scholar] [CrossRef]
- Nelson, C.A.; Thomas, K.M.; de Haan, M.D. Neuroscience of Cognitive Development: The Role of Experience and the Developing Brain; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Sporns, O. Networks of the Brain; MIT Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Medaglia, J.D.; Ramanathan, D.M.; Venkatesan, U.M.; Hillary, F.G. The challenge of non-ergodicity in network neuroscience. Netw. Comput. Neural Syst. 2011, 22, 148–153. [Google Scholar] [CrossRef]
- Atkinson, R.C.; Bower, G.H.; Crothers, E.J. Introduction to Mathematical Learning Theory; American Psychological Association: Washington, DC, USA, 1965. [Google Scholar]
- Vincent, S.B. The Functions of the Vibrissae in the Behavior of the White Rat; University of Chicago: Chicago, IL, USA, 1912; Volume 1. [Google Scholar]
- Bradley, M.M.; Lang, P.J. The International Affective Picture System (IAPS) in the study of emotion and attention. In Handbook of Emotion Elicitation and Assessment; Series in Affective Science; Oxford University Press: New York, NY, USA, 2007; pp. 29–46. [Google Scholar]
- Angold, A.; Costello, E.J.; Messer, S.C.; Pickles, A. Development of a short questionnaire for use in epidemiological studies of depression in children and adolescents. Int. J. Methods Psychiatr. Res. 1995, 5, 251–262. [Google Scholar]
- Tenssay, F.; Wang, H. Analysis of EEG Signals during Visual Processing: An ERP Study. In Proceedings of the 2019 IEEE International Conference on Signal Processing, Communications and Computing (ICSPCC), Dalian, China, 20–23 September 2019; pp. 1–5. [Google Scholar]
- Weiss, B.; Knakker, B.; Vidnyánszky, Z. Visual processing during natural reading. Sci. Rep. 2016, 6, 26902. [Google Scholar] [CrossRef]
- Ibanez, A.; Melloni, M.; Huepe, D.; Helgiu, E.; Rivera-Rei, A.; Canales-Johnson, A.; Baker, P.; Moya, A. What event-related potentials (ERPs) bring to social neuroscience? Soc. Neurosci. 2012, 7, 632–649. [Google Scholar] [CrossRef]
- Hegdé, J. Time course of visual perception: Coarse-to-fine processing and beyond. Prog. Neurobiol. 2008, 84, 405–439. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.; Yeh, W.-H.; Abizaid, A.; Hellemans, K.; Angiulli. Chopped Arms & Big Macs: ERP Correlates of Viewing and Imagining Aversive and Food Photos. Nat. Preced. 2010. [Google Scholar] [CrossRef]
- D’Angiulli, A.; Reeves, A. Generating visual mental images: Latency and vividness are inversely related. Mem. Cogn. 2002, 30, 1179–1188. [Google Scholar] [CrossRef]
- D’Angiulli, A.; Reeves, A. The relationship between self-reported vividness and latency during mental size scaling of everyday items: Phenomenological evidence of different types of imagery. Am. J. Psychol. 2007, 120, 521–551. [Google Scholar] [CrossRef] [PubMed]
- Pilat, A.; Batuecas Suárez, A.; González Nieto, J.L. Terapias Miofasciales: Inducción Miofascial; McGraw-Hill Interamericana: Mexico City, Mexico, 2003. [Google Scholar]
- Pilat, A. Chapter 36—Myofascial induction approaches. In Neck and Arm Pain Syndromes; Fernández de las Peñas, C., Cleland, J.A., Huijbregts, P.A., Eds.; Churchill Livingstone: Edinburgh, UK, 2011; pp. 455–475. [Google Scholar]
- D’Angiulli, A.; Pham, D.A.T.; Leisman, G.; Goldfield, G. Evaluating Preschool Visual Attentional Selective-Set: Preliminary ERP Modeling and Simulation of Target Enhancement Homology. Brain Sci. 2020, 10, 124. [Google Scholar] [CrossRef]
- D’Angiulli, A.; Lockman-Dufour, G.; Buchanan, D.M. Promise for Personalized Diagnosis? Assessing the Precision of Wireless Consumer-Grade Electroencephalography across Mental States. Appl. Sci. 2022, 12, 6430. [Google Scholar] [CrossRef]
- Agam, Y.; Sekuler, R. Interactions between working memory and visual perception: An ERP/EEG study. NeuroImage 2007, 36, 933–942. [Google Scholar] [CrossRef]
- Brown, S.; van Steenbergen, H.; Band, G.; de Rover, M.; Nieuwenhuis, S. Functional significance of the emotion-related late positive potential. Front. Hum. Neurosci. 2012, 6, 33. [Google Scholar] [CrossRef]
- Olofsson, J.K.; Nordin, S.; Sequeira, H.; Polich, J. Affective picture processing: An integrative review of ERP findings. Biol. Psychol. 2008, 77, 247–265. [Google Scholar] [CrossRef]
- Kanske, P.; Plitschka, J.; Kotz, S.A. Attentional orienting towards emotion: P2 and N400 ERP effects. Neuropsychologia 2011, 49, 3121–3129. [Google Scholar] [CrossRef]
- Maeno, T.; Gjini, K.; Iramina, K.; Eto, F.; Ueno, S. Event-related potential P2 derived from visual attention to the hemi-space. Source localization with LORETA. Int. Congr. Ser. 2004, 1270, 262–265. [Google Scholar] [CrossRef]
- Moser, J.S.; Hajcak, G.; Bukay, E.; Simons, R.F. Intentional modulation of emotional responding to unpleasant pictures: An ERP study. Psychophysiology 2006, 43, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Sambuco, N.; Bradley, M.M.; Lang, P.J. Narrative imagery: Emotional modulation in the default mode network. Neuropsychologia 2022, 164, 108087. [Google Scholar] [CrossRef] [PubMed]
- Tarsha, M.S.; Park, S.; Tortora, S. Body-Centered Interventions for Psychopathological Conditions: A Review. Front. Psychol. 2020, 10, 2907. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Thayer, J.F. From the heart to the mind: Cardiac vagal tone modulates top-down and bottom-up visual perception and attention to emotional stimuli. Front. Psychol. 2014, 5, 278. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Zhou, T.; Potter, T.; Zou, L.; Zhang, Y. The Cortical Network of Emotion Regulation: Insights from Advanced EEG-fMRI Integration Analysis. IEEE Trans. Med. Imaging 2019, 38, 2423–2433. [Google Scholar] [CrossRef] [PubMed]
- Nejati, V.; Majdi, R.; Salehinejad, M.A.; Nitsche, M.A. The role of dorsolateral and ventromedial prefrontal cortex in the processing of emotional dimensions. Sci. Rep. 2021, 11, 1971. [Google Scholar] [CrossRef]
- Poremba, A. Chapter 15—Neural and Behavioral Correlates of Auditory Short-Term and Recognition Memory. In Mechanisms of Sensory Working Memory; Jolicoeur, P., Lefebvre, C., Martinez-Trujillo, J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 187–200. [Google Scholar]
- Kensinger, E.A.; Choi, E.S. When side matters: Hemispheric processing and the visual specificity of emotional memories. J. Exp. Psychol. Learn. Mem. Cogn. 2009, 35, 247–253. [Google Scholar] [CrossRef]
- Chen, X.; Jin, J.-N.; Xiang, F.; Liu, Z.-P.; Yin, T. Frontal Eye Field Involvement in Color and Motion Feature-Based Attention: Single-Pulse Transcranial Magnetic Stimulation. Front. Hum. Neurosci. 2018, 12, 390. [Google Scholar] [CrossRef]
- Barceló, F.; Suwazono, S.; Knight, R.T. Prefrontal modulation of visual processing in humans. Nat. Neurosci. 2000, 3, 399–403. [Google Scholar] [CrossRef]
- Qian, Y.; Berenbaum, S.A.; Gilmore, R.O. Vision contributes to sex differences in spatial cognition and activity interests. Sci. Rep. 2022, 12, 17623. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-MIMT | Post-MIMT | ||
|---|---|---|---|
| T(198) * | |||
| Arousal | |||
| Neutral | 3.2 (0.3) | 3.5 (0.5) | 4.68 |
| Pleasant | 5.8 (0.4) | 6.1 (0.2) | 6.71 |
| Unpleasant | 6.8 (0.2) | 6.9 (0.1) | 4.47 |
| Valence | |||
| Neutral | 5.2 (0.1) | 5.4 (0.3) | 6.32 |
| Pleasant | 7.6 (0.3) | 7.8 (0.4) | 4.00 |
| Unpleasant | 2.4 (0.5) | 2.8 (0.6) | 5.12 |
| Condition | Electrode | P1–P2 Amplitude (uV) | T(5) | LPP (Mean uV) | T(5) | ||
|---|---|---|---|---|---|---|---|
| Pre ± SD | Post ± SD | Pre ± SD | Post ± SD | ||||
| Pleasant | Fz | 0.21 ± 0.10 | 0.33 ± 0.15 | 1.67 | 0.51 ± 0.11 | −0.33 ± 0.38 | 5.31 *** |
| F3 | 0.78 ± 0.11 | 0.04 ± 0.12 | 10.72 *** | 0.81 ± 0.21 | 0.19 ± 0.20 | 5.15 *** | |
| F4 | 0.41 ± 0.10 | 0.29 ± 0.06 | 2.40 | 0.69 ± 0.17 | −0.09 ± 0.21 | 7.11 *** | |
| FC3 | 0.22 ± 0.03 | 0.59 ± 0.04 | 16.10 *** | −0.17 ± 0.09 | 0.16 ± 0.10 | 6.16 *** | |
| FC4 | 0.62 ± 0.08 | 0.18 ± 0.11 | 7.45 *** | 0.72 ± 0.25 | −0.13 ± 0.16 | 7.05 *** | |
| Cz | 0.22 ± 0.10 | 0.95 ± 0.06 | 15.75 *** | 0.52 ± 0.13 | 0.34 ± 0.27 | 1.46 | |
| Pz | 0.45 ± 0.18 | 1.21 ± 0.39 | 4.31 ** | 0.36 ± 0.17 | 0.35 ± 0.27 | 0.10 | |
| Oz | 0.10 ± 0.08 | 0.81 ± 0.07 | 17.02 *** | 0.41 ± 0.08 | 0.08 ± 0.17 | 4.10 ** | |
| Neutral | Fz | −0.37 ± 0.03 | −0.18 ± 0.11 | 4.01 ** | −0.24 ± 0.18 | −0.33 ± 0.18 | 0.82 |
| F3 | −0.07 ± 0.05 | 0.25 ± 0.12 | 6.11 *** | 0.24 ± 0.44 | 0.06 ± 0.20 | 0.88 | |
| F4 | −0.35 ± 0.04 | −0.06 ± 0.10 | 6.78 *** | −0.34 ± 0.36 | 0.13 ± 0.10 | 3.13 | |
| FC3 | −0.35 ± 0.08 | 0.41 ± 0.04 | 21.32 *** | 0.00 ± 0.16 | −0.12 ± 0.07 | 1.61 | |
| FC4 | −0.68, ± 0.15 | 0.00 ± 0.07 | 10.04 *** | −0.61 ± 0.21 | −0.20 ± 0.12 | 4.33 ** | |
| Cz | −0.03 ± 0.03 | 0.72 ± 0.11 | 16.00 *** | −0.11 ± 0.08 | 0.47 ± 0.20 | 6.55 *** | |
| Pz | 0.38 ± 0.06 | 1.23 ± 0.14 | 13.64 *** | −0.03 ± 0.07 | 0.64 ± 0.24 | 6.51 *** | |
| Oz | −0.24 ± 0.07 | 0.29 ± 0.10 | 10.50 *** | −0.21 ± 0.09 | 0.09 ± 0.18 | 3.52 * | |
| Unpleasant | Fz | −0.13 ± 0.20 | 0.97 ± 0.07 | 12.96 *** | 0.08 ± 0.09 | −0.07 ± 0.21 | 1.62 |
| F3 | 0.33 ± 0.16 | 1.29 ± 0.07 | 13.42 *** | 0.52 ± 0.22 | 0.49 ± 0.32 | 0.11 | |
| F4 | 0.30 ± 0.10 | 0.89 ± 0.07 | 12.12 *** | 0.04 ± 0.12 | 0.14 ± 0. 29 | 0.75 | |
| FC3 | 0.49 ± 071 | 0.42 ± 0.08 | 1.52 | −0.27 ± 0.20 | −0.30 ± 0.05 | 0.33 | |
| FC4 | 0.59 ± 0.08 | 0.09 ± 0.13 | 7.94 *** | 0.34 ± 0.27 | −0.44 ± 0.24 | 5.22 *** | |
| Cz | 0.51 ± 0.15 | 1.77 ± 0.11 | 16.16 *** | 0.16 ± 0.20 | −0.07 ± 0.32 | 1.46 | |
| Pz | 0.21 ± 0.13 | 1.71 ± 0.76 | 4.73 | −0.44 ± 0.16 | −0.02 ± 0.28 | 3.22 * | |
| Oz | 0.58 ± 0.15 | 1.60 ± 0.09 | 13.84 *** | 0.38 ± 0.21 | −0.18 ± 0.29 | 3.82 ** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byczynski, G.; D’Angiulli, A. Brief Myofascial Intervention Modulates Visual Event-Related Potential Response to Emotional Photographic Contents: A Pilot Study. Vision 2023, 7, 77. https://doi.org/10.3390/vision7040077
Byczynski G, D’Angiulli A. Brief Myofascial Intervention Modulates Visual Event-Related Potential Response to Emotional Photographic Contents: A Pilot Study. Vision. 2023; 7(4):77. https://doi.org/10.3390/vision7040077
Chicago/Turabian StyleByczynski, Gabriel, and Amedeo D’Angiulli. 2023. "Brief Myofascial Intervention Modulates Visual Event-Related Potential Response to Emotional Photographic Contents: A Pilot Study" Vision 7, no. 4: 77. https://doi.org/10.3390/vision7040077