Spatial Attention in Visual Working Memory Strengthens Feature-Location Binding


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experiment 1
2.1. Material and Methods
2.1.1. Participants
2.1.2. Apparatus
2.1.3. Stimuli
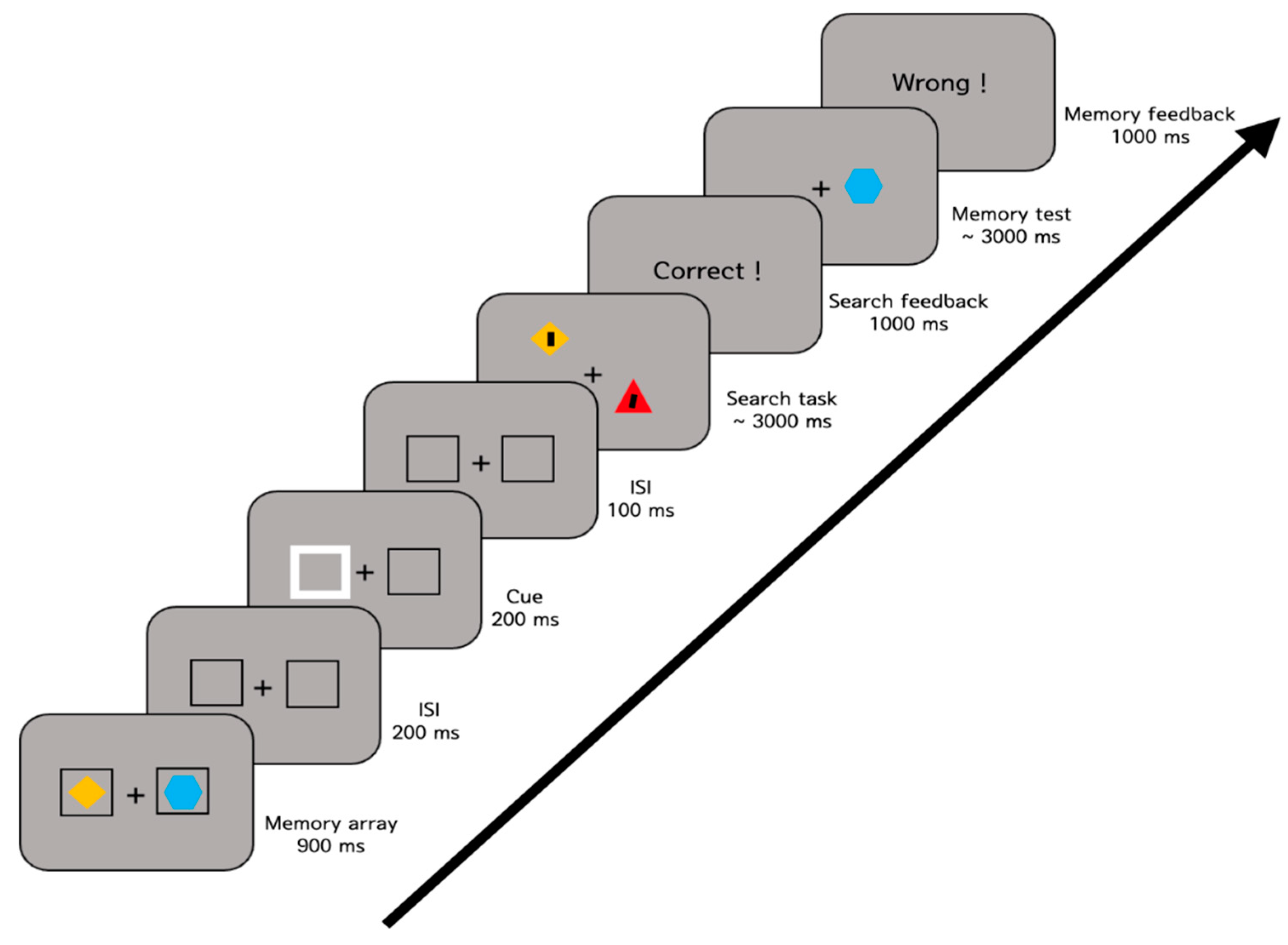
2.1.4. Procedure
2.2. Results
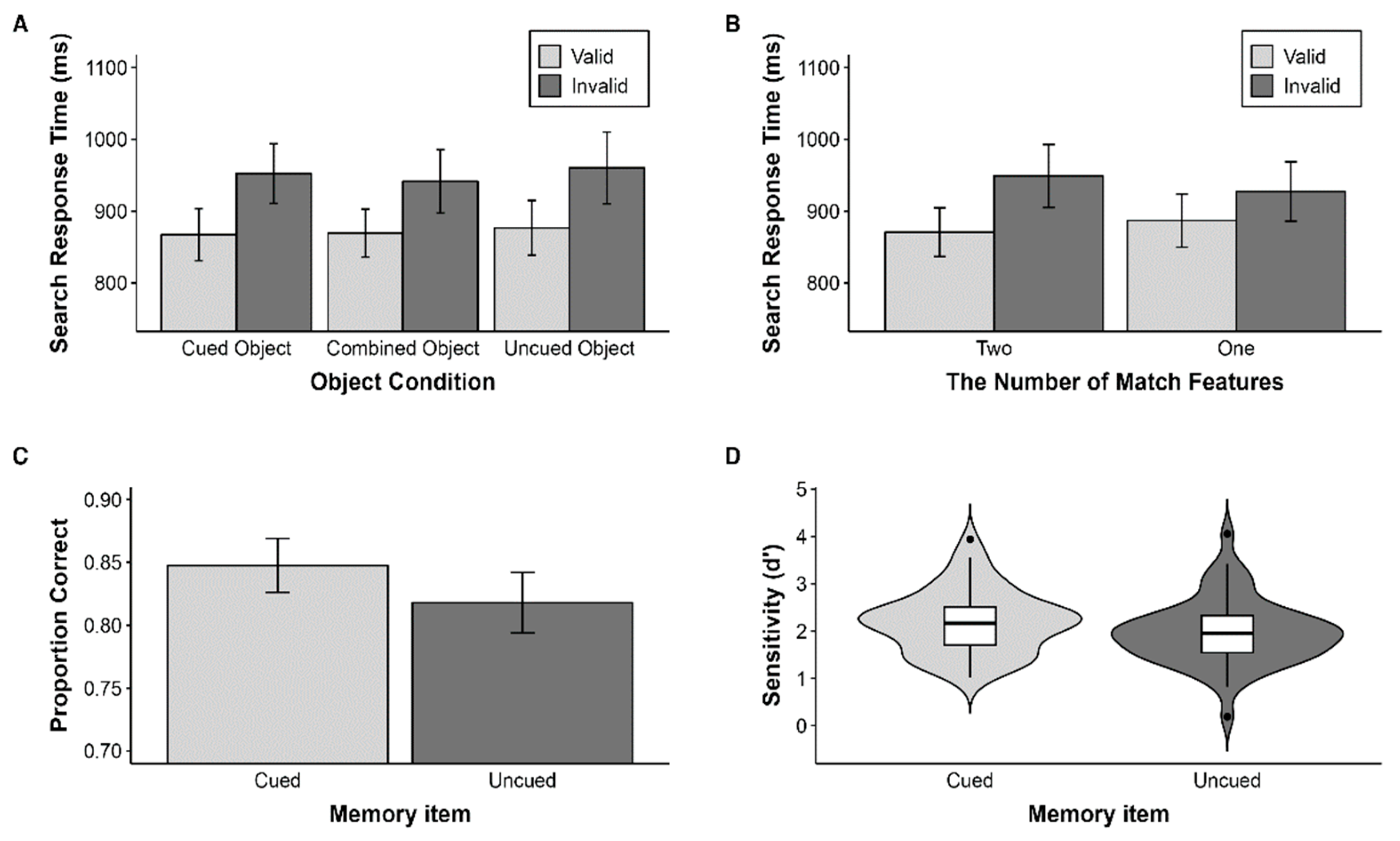
2.2.1. Search Task
2.2.2. Memory Task
2.3. Discussion
3. Experiment 2
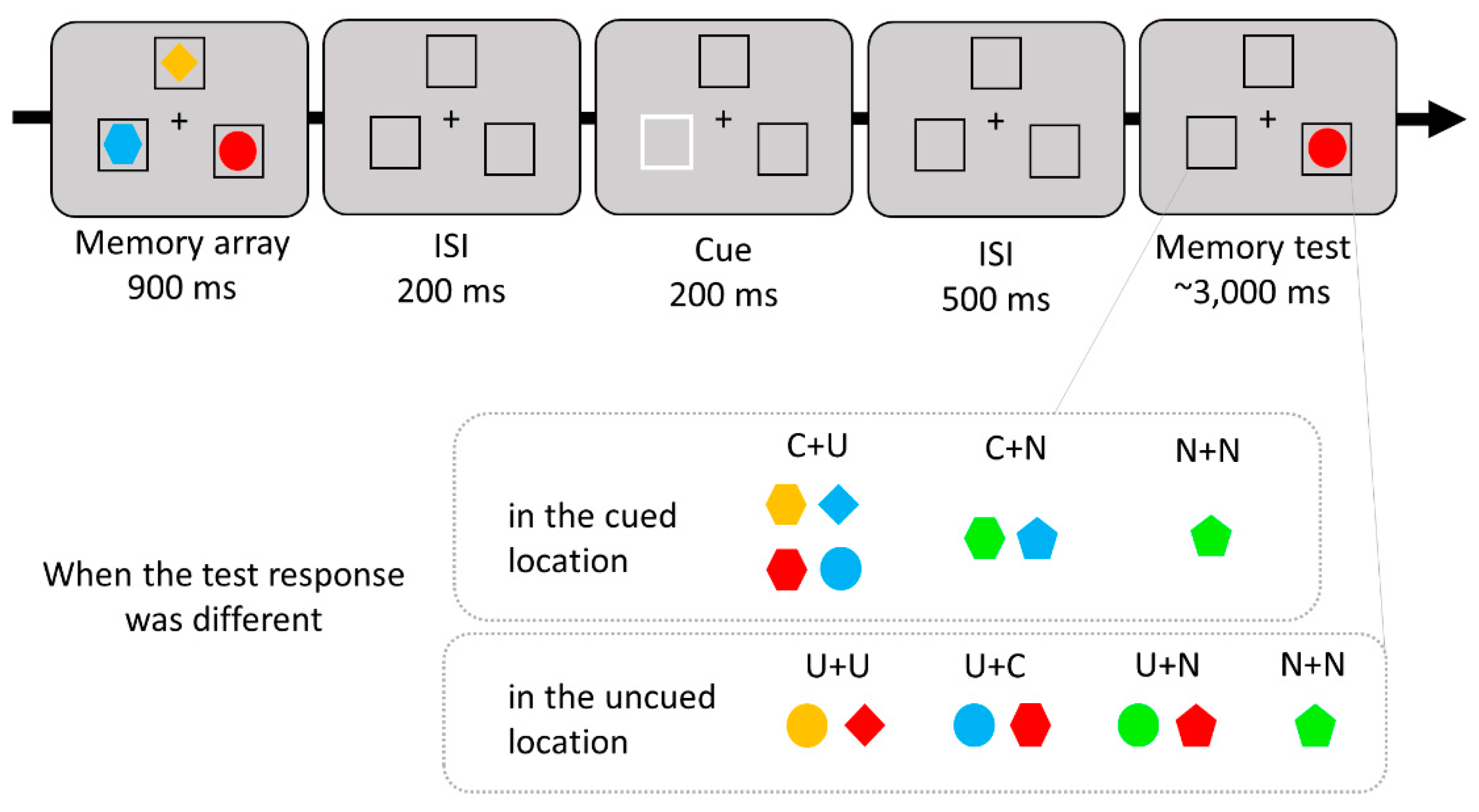
3.1. Material and Methods
3.1.1. Participants
3.1.2. Apparatus, Stimulus, and Procedure
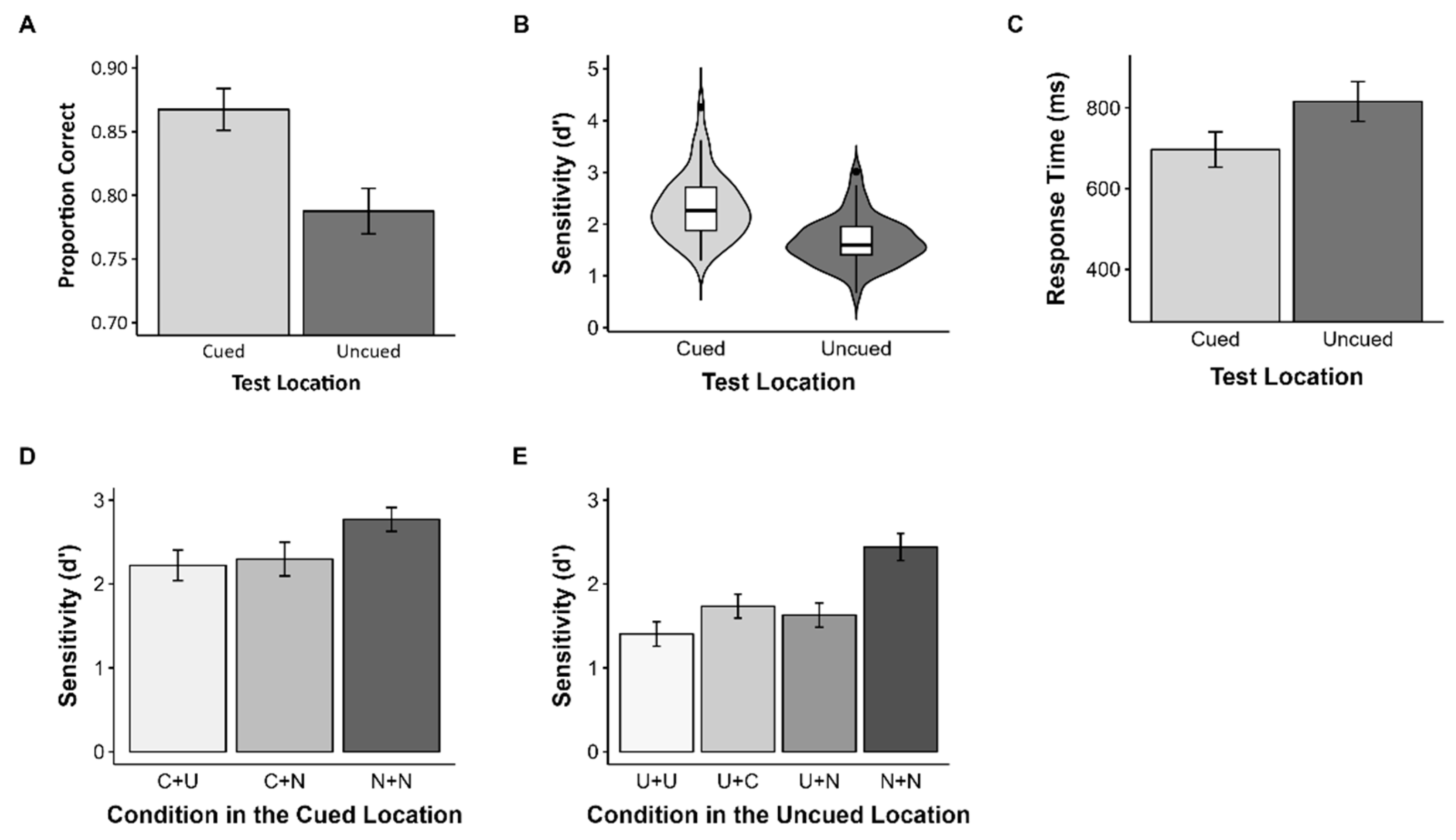
3.2. Results
3.3. Discussion
4. General Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Livingstone, M.; Hubel, D. Segregation of form, color, movement, and depth: Anatomy, physiology, and perception. Science 1988, 240, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Conway, B.R.; Chatterjee, S.; Field, G.D.; Horwitz, G.D.; Johnson, E.N.; Koida, K.; Mancuso, K. Advances in color science: From retina to behavior. J. Neurosci. 2010, 30, 14955–14963. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, G.D. Signals related to color in the early visual cortex. Annu. Rev. Vis. Sci. 2020, 6, 287–311. [Google Scholar] [CrossRef] [PubMed]
- Stefurak, D.L.; Boynton, R.M. Independence of memory for categorically different colors and shapes. Percept. Psychophys. 1986, 39, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Reeves, A.; Fuller, H.; Fine, E.M. The role of attention in binding shape to color. Vis. Res. 2005, 45, 3343–3355. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J.; Vogel, E.K. The capacity of visual working memory for features and conjunctions. Nature 1997, 390, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, G.A.; Cavanagh, P. The capacity of visual short-term memory is set both by visual information load and by number of objects. Psychol. Sci. 2004, 15, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Hardman, K.O.; Cowan, N. Remembering complex objects in visual working memory: Do capacity limits restrict objects or features? J. Exp. Psychol. Learn. Mem. Cogn. 2015, 41, 325. [Google Scholar] [CrossRef]
- Olson, I.R.; Jiang, Y. Is visual short-term memory object based? Rejection of the “strong-object” hypothesis. Percept. Psychophys. 2002, 64, 1055–1067. [Google Scholar] [CrossRef]
- Franconeri, S.L.; Alvarez, G.A.; Cavanagh, P. Flexible cognitive resources: Competitive content maps for attention and memory. Trends Cogn. Sci. 2013, 17, 134–141. [Google Scholar] [CrossRef]
- Wang, B.; Cao, X.; Theeuwes, J.; Olivers, C.N.; Wang, Z. Location-based effects underlie feature conjunction benefits in visual working memory. J. Vis. 2016, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Bays, P.M.; Wu, E.Y.; Husain, M. Storage and binding of object features in visual working memory. Neuropsychologia 2011, 49, 1622–1631. [Google Scholar] [CrossRef] [PubMed]
- Awh, E.; Jonides, J.; Reuter-Lorenz, P.A. Rehearsal in spatial working memory. J. Exp. Psychol. Hum. Percept. Perform. 1998, 24, 780. [Google Scholar] [CrossRef] [PubMed]
- Chun, M.M. Visual working memory as visual attention sustained internally over time. Neuropsychologia 2011, 49, 1407–1409. [Google Scholar] [CrossRef] [PubMed]
- Cowan, N. Attention and Memory: An Integrated Framework; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Olivers, C.N. Interactions between visual working memory and visual attention. Front. Biosci.-Landmark 2008, 13, 1182–1191. [Google Scholar] [CrossRef] [PubMed]
- Downing, P.E. Interactions between visual working memory and selective attention. Psychol. Sci. 2000, 11, 467–473. [Google Scholar] [CrossRef]
- Pan, Y. Attentional capture by working memory contents. Can. J. Exp. Psychol./Rev. Can. Psychol. Exp 2010, 64, 124. [Google Scholar] [CrossRef]
- Sasin, E.; Fougnie, D. Memory-driven capture occurs for individual features of an object. Sci. Rep. 2020, 10, 19499. [Google Scholar] [CrossRef]
- Thayer, D.D.; Bahle, B.; Hollingworth, A. Guidance of attention from visual working memory is feature-based, not object-based: Implications for models of feature binding. J. Exp. Psychol. Gen. 2022, 151, 1018. [Google Scholar] [CrossRef]
- Rangelov, D.; Zeki, S. Non-binding relationship between visual features. Front. Hum. Neurosci. 2014, 8, 749. [Google Scholar] [CrossRef]
- Kovacs, O.; Harris, I.M. The role of location in visual feature binding. Atten. Percept. Psychophys. 2019, 81, 1551–1563. [Google Scholar] [CrossRef] [PubMed]
- Schneegans, S.; Bays, P.M. Neural architecture for feature binding in visual working memory. J. Neurosci. 2017, 37, 3913–3925. [Google Scholar] [CrossRef] [PubMed]
- Leslie, A.M.; Xu, F.; Tremoulet, P.D.; Scholl, B.J. Indexing and the object concept: Developingwhat’andwhere’systems. Trends Cogn. Sci. 1998, 2, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Golomb, J.D.; Kupitz, C.N.; Thiemann, C.T. The influence of object location on identity: A “spatial congruency bias”. J. Exp. Psychol. Gen. 2014, 143, 2262. [Google Scholar] [CrossRef] [PubMed]
- Treisman, A.M.; Gelade, G. A feature-integration theory of attention. Cogn. Psychol. 1980, 12, 97–136. [Google Scholar] [CrossRef]
- Paul, L.; Schyns, P.G. Attention enhances feature integration. Vis. Res. 2003, 43, 1793–1798. [Google Scholar] [CrossRef]
- Treisman, A.; Schmidt, H. Illusory conjunctions in the perception of objects. Cogn. Psychol. 1982, 14, 107–141. [Google Scholar] [CrossRef]
- Brown, L.A.; Brockmole, J.R. The role of attention in binding visual features in working memory: Evidence from cognitive ageing. Q. J. Exp. Psychol. 2010, 63, 2067–2079. [Google Scholar] [CrossRef]
- Briand, K.A. Feature integration and spatial attention: More evidence of a dissociation between endogenous and exogenous orienting. J. Exp. Psychol. Hum. Percept. Perform. 1998, 24, 1243. [Google Scholar] [CrossRef]
- Macmillan, N.A.; Creelman, C.D. Detection Theory: A User’s Guide; Psychology Press: Mahwah, NJ, USA, 2004. [Google Scholar]
- Souza, A.S.; Skóra, Z. The interplay of language and visual perception in working memory. Cognition 2017, 166, 277–297. [Google Scholar] [CrossRef]
- Sense, F.; Morey, C.C.; Prince, M.; Heathcote, A.; Morey, R.D. Opportunity for verbalization does not improve visual change detection performance: A state-trace analysis. Behav. Res. Methods 2017, 49, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, A.; Johnson, W.; Logie, R.H. Aging and feature-binding in visual working memory: The role of verbal rehearsal. Psychol. Aging 2019, 34, 933. [Google Scholar] [CrossRef] [PubMed]
- Fougnie, D.; Alvarez, G.A. Object features fail independently in visual working memory: Evidence for a probabilistic feature-store model. J. Vis. 2011, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.S.; Oberauer, K. In search of the focus of attention in working memory: 13 years of the retro-cue effect. Atten. Percept. Psychophys. 2016, 78, 1839–1860. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Cave, K.R. Spatial attention in visual search for features and feature conjunctions. Psychol. Sci. 1995, 6, 376–380. [Google Scholar] [CrossRef]
- Kim, J.; Kim, M.-S.; Chun, M.M. Predictive spatial working memory content guides visual search. Vis. Cogn. 2010, 18, 574–590. [Google Scholar] [CrossRef]
- Han, S.-H.; Kim, M.-S. Visual search does not remain efficient when executive working memory is working. Psychol. Sci. 2004, 15, 623–628. [Google Scholar] [CrossRef]
- Han, S.W.; Kim, M.-S. Do the contents of working memory capture attention? Yes, but cognitive control matters. J. Exp. Psychol. Hum. Percept. Perform. 2009, 35, 1292. [Google Scholar] [CrossRef]
- Oh, S.-H.; Kim, M.-S. The role of spatial working memory in visual search efficiency. Psychon. Bull. Rev. 2004, 11, 275–281. [Google Scholar] [CrossRef]
- Soto, D.; Humphreys, G.W.; Heinke, D. Working memory can guide pop-out search. Vis. Res. 2006, 46, 1010–1018. [Google Scholar] [CrossRef]
- Park, S.; Kim, M.-S.; Chun, M.M. Concurrent working memory load can facilitate selective attention: Evidence for specialized load. J. Exp. Psychol. Hum. Percept. Perform. 2007, 33, 1062. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Kim, M.-S.; Chun, M.M. Concurrent working memory load can reduce distraction. Proc. Natl. Acad. Sci. USA 2005, 102, 16524–16529. [Google Scholar] [CrossRef] [PubMed]
- Scholl, B.J. Objects and attention: The state of the art. Cognition 2001, 80, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Hollingworth, A.; Beck, V.M. Memory-based attention capture when multiple items are maintained in visual working memory. J. Exp. Psychol. Hum. Percept. Perform. 2016, 42, 911. [Google Scholar] [CrossRef] [PubMed]
- Bays, P.M.; Husain, M. Dynamic shifts of limited working memory resources in human vision. Science 2008, 321, 851–854. [Google Scholar] [CrossRef]
- Pasternak, T.; Greenlee, M.W. Working memory in primate sensory systems. Nat. Rev. Neurosci. 2005, 6, 97–107. [Google Scholar] [CrossRef]
- Herwig, A.; Beisert, M.; Schneider, W.X. On the spatial interaction of visual working memory and attention: Evidence for a global effect from memory-guided saccades. J. Vis. 2010, 10, 8. [Google Scholar] [CrossRef]
- Dalmaso, M.; Castelli, L.; Scatturin, P.; Galfano, G. Working memory load modulates microsaccadic rate. J. Vis. 2017, 17, 6. [Google Scholar] [CrossRef]
- Schmidt, B.K.; Vogel, E.K.; Woodman, G.F.; Luck, S.J. Voluntary and automatic attentional control of visual working memory. Percept. Psychophys. 2002, 64, 754–763. [Google Scholar] [CrossRef]
- Golomb, J.D.; Nguyen-Phuc, A.Y.; Mazer, J.A.; McCarthy, G.; Chun, M.M. Attentional facilitation throughout human visual cortex lingers in retinotopic coordinates after eye movements. J. Neurosci. 2010, 30, 10493–10506. [Google Scholar] [CrossRef]
- Van der Stigchel, S.; Merten, H.; Meeter, M.; Theeuwes, J. The effects of a task-irrelevant visual event on spatial working memory. Psychon. Bull. Rev. 2007, 14, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Golomb, J.D.; L’Heureux, Z.E.; Kanwisher, N. Feature-binding errors after eye movements and shifts of attention. Psychol. Sci. 2014, 25, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Dowd, E.W.; Golomb, J.D. The Binding Problem after an eye movement. Atten. Percept. Psychophys. 2020, 82, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Robertson, L.C. Implicit representations of space after bilateral parietal lobe damage. J. Cogn. Neurosci. 2001, 13, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Rerko, L.; Oberauer, K. Focused, unfocused, and defocused information in working memory. J. Exp. Psychol. Learn. Mem. Cogn. 2013, 39, 1075. [Google Scholar] [CrossRef] [PubMed]
- Kahneman, D.; Treisman, A.; Gibbs, B.J. The reviewing of object files: Object-specific integration of information. Cogn. Psychol. 1992, 24, 175–219. [Google Scholar] [CrossRef]
- Noles, N.S.; Scholl, B.J.; Mitroff, S.R. The persistence of object file representations. Percept. Psychophys. 2005, 67, 324–334. [Google Scholar] [CrossRef]
- Soto, D.; Heinke, D.; Humphreys, G.W.; Blanco, M.J. Early, involuntary top-down guidance of attention from working memory. J. Exp. Psychol. Hum. Percept. Perform. 2005, 31, 248. [Google Scholar] [CrossRef]
- Zhang, B.; Huang, S.; Hou, Q. The priority of color in working-memory-driven ocular capture. Acta Psychol. Sin. 2014, 46, 17. [Google Scholar] [CrossRef]
- Rajsic, J.; Wilson, D.E. Asymmetrical access to color and location in visual working memory. Atten. Percept. Psychophys. 2014, 76, 1902–1913. [Google Scholar] [CrossRef]
- Gegenfurtner, K.R.; Sperling, G. Information transfer in iconic memory experiments. J. Exp. Psychol. Hum. Percept. Perform. 1993, 19, 845. [Google Scholar] [CrossRef] [PubMed]
- Vogel, E.K.; Woodman, G.F.; Luck, S.J. The time course of consolidation in visual working memory. J. Exp. Psychol. Hum. Percept. Perform. 2006, 32, 1436. [Google Scholar] [CrossRef] [PubMed]
- van Lamsweerde, A.E.; Johnson, J.S. Assessing the effect of early visual cortex transcranial magnetic stimulation on working memory consolidation. J. Cogn. Neurosci. 2017, 29, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- van de Ven, V.; Jacobs, C.; Sack, A.T. Topographic contribution of early visual cortex to short-term memory consolidation: A transcranial magnetic stimulation study. J. Neurosci. 2012, 32, 4–11. [Google Scholar] [CrossRef]
- Emrich, S.M.; Riggall, A.C.; LaRocque, J.J.; Postle, B.R. Distributed patterns of activity in sensory cortex reflect the precision of multiple items maintained in visual short-term memory. J. Neurosci. 2013, 33, 6516–6523. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joe, J.; Kim, M.-S. Spatial Attention in Visual Working Memory Strengthens Feature-Location Binding. Vision 2023, 7, 79. https://doi.org/10.3390/vision7040079
Joe J, Kim M-S. Spatial Attention in Visual Working Memory Strengthens Feature-Location Binding. Vision. 2023; 7(4):79. https://doi.org/10.3390/vision7040079
Chicago/Turabian StyleJoe, Juyeon, and Min-Shik Kim. 2023. "Spatial Attention in Visual Working Memory Strengthens Feature-Location Binding" Vision 7, no. 4: 79. https://doi.org/10.3390/vision7040079
APA StyleJoe, J., & Kim, M.-S. (2023). Spatial Attention in Visual Working Memory Strengthens Feature-Location Binding. Vision, 7(4), 79. https://doi.org/10.3390/vision7040079