
Structural Characterization of Toxoplasma gondii Brain Cysts in a Model of Reactivated Toxoplasmosis Using Computational Image Analysis
 , , , , , , ,
, , , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Parasites
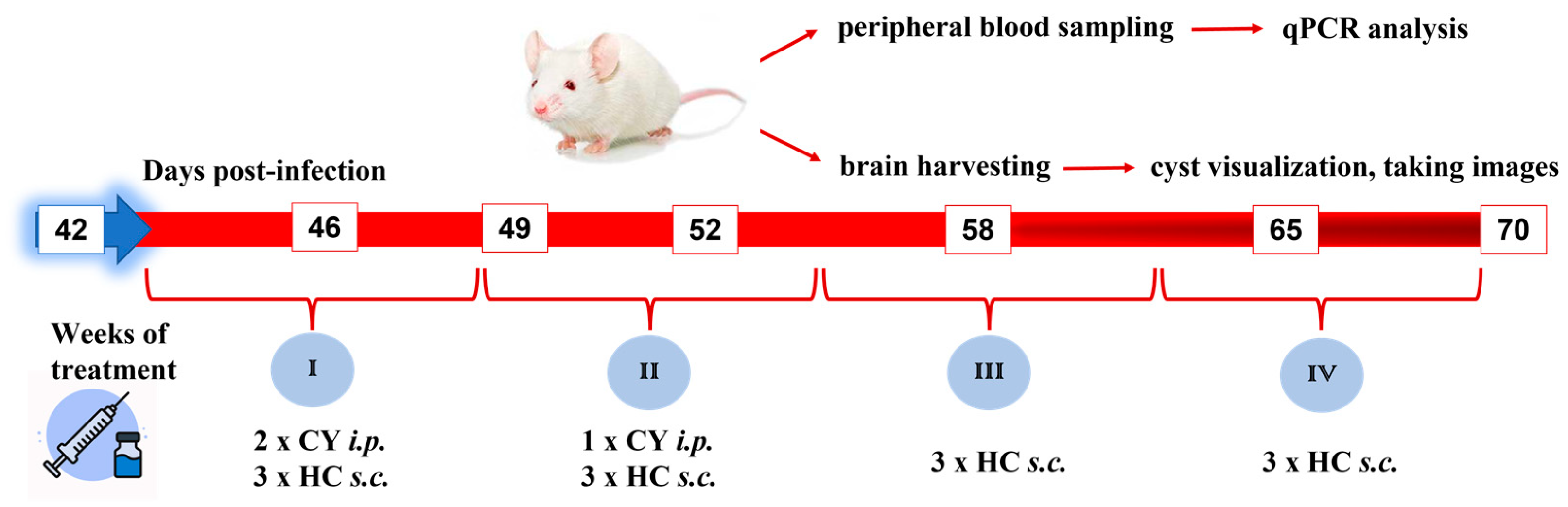
2.3. Experimental Design
2.4. Image Acquisition
2.5. Image Analysis
2.5.1. Morphological Analysis
2.5.2. Particle Analysis
2.5.3. Fractal Analysis
2.6. Statistical Analysis
2.7. Ethics Statement
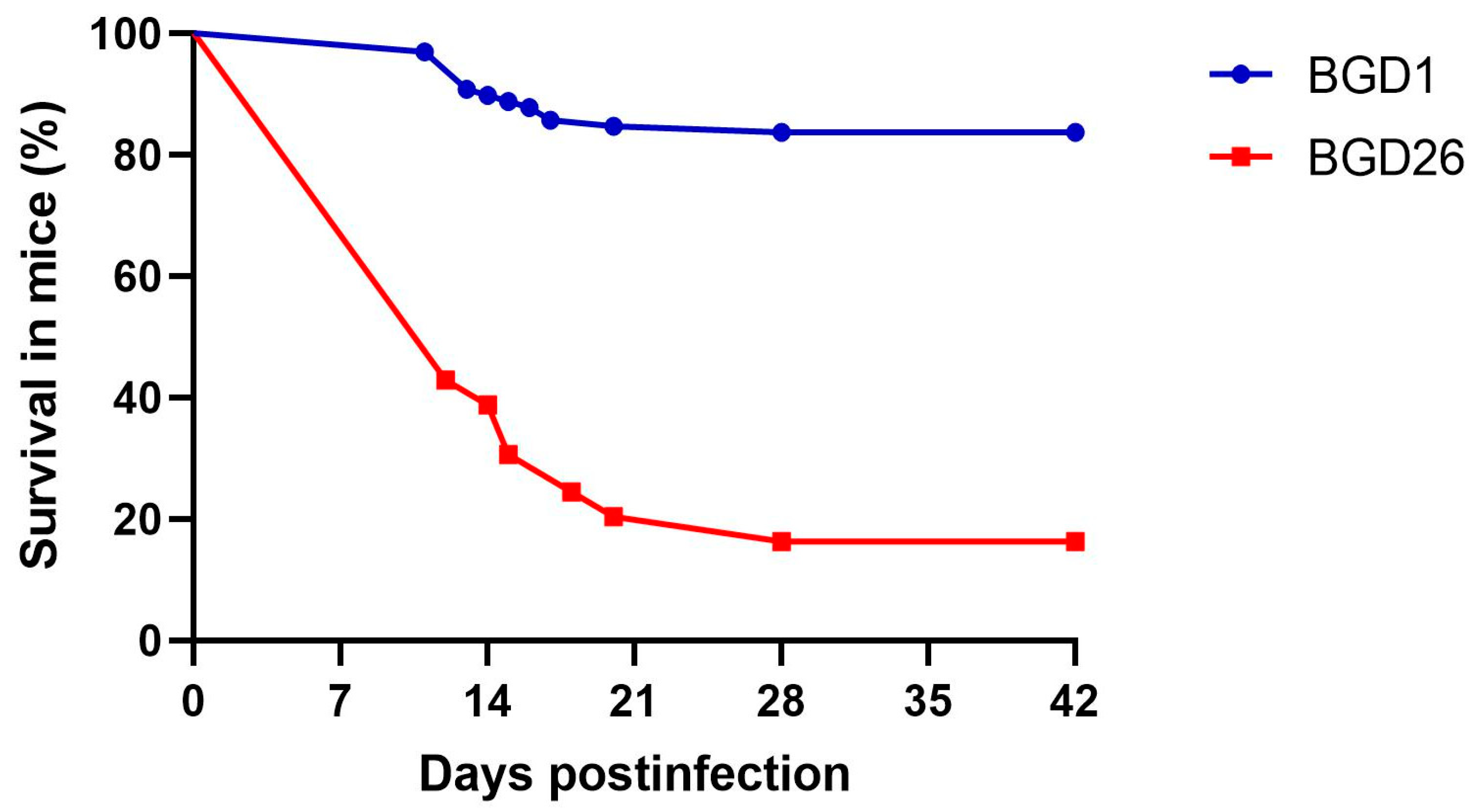
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dubey, J.P. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 1998, 28, 1019–1024. [Google Scholar] [CrossRef]
- Weiss, L.M.; Dubey, J.P. Toxoplasmosis: A history of clinical observations. Int. J. Parasitol. 2009, 39, 895–901. [Google Scholar] [CrossRef]
- Augusto, L.; Wek, R.C.; Sullivan, W.J., Jr. Host sensing and signal transduction during Toxoplasma stage conversion. Mol. Microbiol. 2021, 115, 839–848. [Google Scholar] [CrossRef]
- Martynowicz, J.; Doggett, J.S.; Sullivan, W.J., Jr. Efficacy of Guanabenz Combination Therapy against Chronic Toxoplasmosis across Multiple Mouse Strains. Antimicrob. Agents. Chemother. 2020, 64, e00539-20. [Google Scholar] [CrossRef]
- Hamie, M.; Najm, R.; Deleuze-Masquefa, C.; Bonnet, P.A.; Dubremetz, J.F.; El Sabban, M.; El Hajj, H. Imiquimod Targets Toxoplasmosis Through Modulating Host Toll-Like Receptor-MyD88 Signaling. Front. Immunol. 2021, 12, 629917. [Google Scholar] [CrossRef]
- Rodriguez, J.B.; Szajnman, S.H. An updated review of chemical compounds with anti-Toxoplasma gondii activity. Eur. J. Med. Chem. 2023, 262, 115885. [Google Scholar] [CrossRef]
- Angel, S.O.; Vanagas, L.; Ruiz, D.M.; Cristaldi, C.; Saldarriaga Cartagena, A.M.; Sullivan, W.J., Jr. Emerging Therapeutic Targets Against Toxoplasma gondii: Update on DNA Repair Response Inhibitors and Genotoxic Drugs. Front. Cell. Infect. Microbiol. 2020, 10, 289. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.D.; Teixeira, C.; Gomes, P.; Borges, M. Promising Drug Targets and Compounds with Anti-Toxoplasma gondii Activity. Microorganisms 2021, 9, 1960. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of toxoplasmosis: Historical perspective, animal models, and current clinical practice. Clin. Microbiol. Rev. 2018, 31, e00057-17. [Google Scholar] [CrossRef] [PubMed]
- Koundal, D.; Gupta, S. Advances in Computational Techniques for Biomedical Image Analysis; Academic Press: Cambridge, MA, USA, 2020; pp. 299–308. [Google Scholar]
- Madabhushi, A.; Lee, G. Image analysis and machine learning in digital pathology: Challenges and opportunities. Med. Image Anal. 2016, 33, 170–175. [Google Scholar] [CrossRef]
- Sbalzarini, I.F. Seeing is Believing: Quantifying is Convincing: Computational image analysis in biology. In Focus on Bio-Image Informatics. Advances in Anatomy, Embryology and Cell Biology; De Vos, W., Munck, S., Timmermans, J.P., Eds.; Springer: Cham, Switzerland, 2016; Volume 219. [Google Scholar] [CrossRef]
- AlSheikh, M.H.; Al-Saidi, N.M.G.; Ibrahim, R.W. Dental X-ray identification system based on association rules extracted by k-symbol fractional Haar functions. Fractal Fract. 2022, 6, 669. [Google Scholar] [CrossRef]
- Pantic, I.; Valjarevic, S.; Cumic, J.; Paunkovic, I.; Terzic, T.; Corridon, P.R. Gray level co-occurrence matrix, fractal and wavelet analyses of discrete changes in cell nuclear structure following osmotic stress: Focus on machine learning methods. Fractal Fract. 2023, 7, 272. [Google Scholar] [CrossRef]
- Djuričić, G.J.; Radulović, M.; Sopta, J.P.; Nikitović, M.; Milošević, N.T. Fractal and Gray Level Cooccurrence Matrix computational analysis of primary osteosarcoma magnetic resonance images predicts the chemotherapy response. Front. Oncol. 2017, 7, 246. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Vázquez, A.Y.; Cuautle-Estrada, A.; Grave-Capistrán, M.A.; Ramírez, O.; Torres-SanMiguel, C.R. Fractal analysis and FEM assessment of soft tissue affected by fibrosis. Fractal Fract. 2023, 7, 661. [Google Scholar] [CrossRef]
- Zaia, A.; Maponi, P. Mitochondrial DNA profiling by fractal lacunarity to characterize the senescent phenotype as normal aging or pathological aging. Fractal Fract. 2022, 6, 219. [Google Scholar] [CrossRef]
- Atupelage, C.; Nagahashi, H.; Yamaguchi, M.; Abe, T.; Hashiguchi, A.; Sakamoto, M. Computational grading of hepatocellular carcinoma using multifractal feature description. Comput. Med. Imaging Graph. 2013, 37, 61–71. [Google Scholar] [CrossRef]
- Abdelsalam, M.M.; Zahran, M.A. A novel approach of diabetic retinopathy early detection based on multifractal geometry analysis for OCTA macular images using support vector machine. IEEE Access 2021, 9, 22844–22858. [Google Scholar] [CrossRef]
- He, S.; Thangaraj, C.; Easwaramoorthy, D.; Muhiuddin, G. Multifractal analysis on age-based discrimination in X-ray images for sensing the severity of COVID-19 disease. Eur. Phys. J. Spec. Top. 2022, 231, 3663–3671. [Google Scholar] [CrossRef]
- Rani, P.; Kotwal, S.; Manhas, J.; Sharma, V.; Sharma, S. Machine Learning and Deep Learning Based Computational Approaches in Automatic Microorganisms Image Recognition: Methodologies, Challenges, and Developments. Arch. Comput. Methods. Eng. 2022, 29, 1801–1837. [Google Scholar] [CrossRef]
- Shambhu, S.; Koundal, D.; Das, P.; Hoang, V.T.; Tran-Trung, K.; Turabieh, H. Computational Methods for Automated Analysis of Malaria Parasite Using Blood Smear Images: Recent Advances. Comput. Intell. Neurosci. 2022, 2022, 3626726. [Google Scholar] [CrossRef]
- Jeckel, H.; Drescher, K. Advances and opportunities in image analysis of bacterial cells and communities. FEMS Microbiol. Rev. 2021, 45, fuaa062. [Google Scholar] [CrossRef]
- Karperien, A.L.; Jelinek, H.F. Box-counting fractal analysis: A primer for the clinician. In The Fractal Geometry of the Brain; Di Ieva, A., Ed.; Springer: New York, NY, USA, 2016; pp. 91–108. [Google Scholar] [CrossRef]
- Bauman, N.; Ilić, A.; Lijeskić, O.; Uzelac, A.; Klun, I.; Srbljanović, J.; Ćirković, V.; Bobić, B.; Štajner, T.; Djurković-Djaković, O. Computational image analysis reveals the structural complexity of Toxoplasma gondii tissue cysts. PLoS ONE 2020, 15, e0234169. [Google Scholar] [CrossRef] [PubMed]
- Passweg, J.R.; Baldomero, H.; Bader, P.; Bonini, C.; Cesaro, S.; Dreger, P.; Duarte, R.F.; Dufour, C.; Kuball, J.; Farge-Bancel, D.; et al. Hematopoietic stem cell transplantation in Europe 2014: More than 40000 transplants annually. Blood Marrow Transplant. 2016, 51, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Djurkovic-Djakovic, O.; Milenkovic, V. Murine model of drug induced reactivation of Toxoplasma gondii. Acta Protozool 2001, 40, 99–106. [Google Scholar]
- Carreras, E.; Dufour, C.; Mohty, M.; Kröger, N. (Eds.) The EBMT Handbook: Hematopoietic Stem Cell Transplantation and Cellular Therapies, 7th ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Silva, L.A.; Brandão, G.P.; Pinheiro, B.V.; Vitor, R.W. Immunosuppression with cyclophosphamide favors reinfection with recombinant Toxoplasma gondii strains. Parasite 2012, 19, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Yao, Y.; El-Ashram, S.; Tian, J.; Shen, J.; Ji, Y. The Neurotropic Parasite Toxoplasma gondii Induces Astrocyte Polarization through NFκB Pathway. Front. Med. 2019, 6, 267. [Google Scholar] [CrossRef] [PubMed]
- Djurković-Djaković, O.; Klun, I.; Khan, A.; Nikolić, A.; Knezević-Usaj, S.; Bobić, B.; Sibley, L.D. A human origin type II strain of Toxoplasma gondii causing severe encephalitis in mice. Microbes Infect. 2006, 8, 2206–2212. [Google Scholar] [CrossRef] [PubMed]
- Stajner, T.; Bobic, B.; Klun, I.; Nikolic, A.; Srbljanovic, J.; Uzelac, A.; Rajnpreht, I.; Djurkovic-Djakovic, O. Prenatal and early postnatal diagnosis of congenital toxoplasmosis in a setting with no systematic screening in pregnancy. Medicine 2016, 95, e2979. [Google Scholar] [CrossRef]
- Rajnpreht, I.; Klun, I.; Uzelac, A.; Bobić, B.; Nikolić, A.; Srbljanović, J.; Djurković-Djaković, O. Vero cells as an in vitro model system for the propagation of type-3 Toxoplasma gondii tachyzoites. In Proceedings of the 12th European Multicolloquium of Parasitology, Turku, Finland, 20–24 July 2016. [Google Scholar]
- Desmonts, G.; Remington, J.S. Direct agglutination test for diagnosis of Toxoplasma infection: Method for increasing sensitivity and specificity. J. Clin. Microbiol. 1980, 11, 562–568. [Google Scholar] [CrossRef]
- Huyan, X.H.; Lin, Y.P.; Gao, T.; Chen, R.Y.; Fan, Y.M. Immunosuppressive effect of cyclophosphamide on white blood cells and lymphocyte subpopulations from peripheral blood of Balb/c mice. Int. Immunopharmacol. 2011, 11, 1293–1297. [Google Scholar] [CrossRef]
- Stajner, T.; Vasiljević, Z.; Vujić, D.; Marković, M.; Ristić, G.; Mićić, D.; Pasić, S.; Ivović, V.; Ajzenberg, D.; Djurković-Djaković, O. Atypical strain of Toxoplasma gondii causing fatal reactivation after hematopoietic stem cell transplantion in a patient with an underlying immunological deficiency. J. Clin. Microbiol. 2013, 51, 2686–2690. [Google Scholar] [CrossRef]
- Singh, T.R.; Roy, S.; Singh, O.I.; Sinam, T.; Singh, K.M. A new local adaptive thresholding technique in binarization. Int. J. Comp. Sci. 2011, 8, 271–277. [Google Scholar] [CrossRef]
- Soille, P.; Vincent, L. Watersheds in digital spaces: An efficient algorithm based on immersion simulations. IEEE Trans. Pattern Anal. Mach. Intell. 1991, 13, 583–598. [Google Scholar] [CrossRef]
- Kornilov, A.S.; Safonov, I.V. An overview of watershed algorithm implementations in open source libraries. J. Imaging 2018, 4, 123. [Google Scholar] [CrossRef]
- Rougier, S.; Montoya, J.G.; Peyron, F. Lifelong persistence of toxoplasma cysts: A questionable dogma? Trends Parasitol. 2017, 33, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Watts, E.; Zhao, Y.; Dhara, A.; Eller, B.; Patwardhan, A.; Sinai, A.P. Novel approaches reveal that Toxoplasma gondii bradyzoites within tissue cysts are dynamic and replicating entities in vivo. mBio 2015, 6, e01155-15. [Google Scholar] [CrossRef]
- Armando, Q.; Graciela, J. Identification of Toxoplasma gondii cysts in samples from Colombia using digital image processing. In Proceedings of the 2014 IEEE ANDESCON, Cochabamba, Bolivia, 15–17 October 2014; p. 1. [Google Scholar] [CrossRef]
- Howe, D.K.; Sibley, L.D. Toxoplasma gondii comprises three clonal lineages: Correlation of parasite genotype with human disease. J. Infect. Dis. 1995, 172, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Szabo, E.K.; Finney, C.A.M. Toxoplasma gondii: One organism, multiple models. Trends Parasitol. 2017, 33, 113–127. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | IC | EG | Total | |
|---|---|---|---|---|
| PCR+ | PCR− | |||
| BGD1 | 27 | 7 | 21 | 55 |
| BGD26 | 10 | 15 | 10 | 35 |
| Strain | Group | D (μm) | C | PD (μm–2) | FD | L |
|---|---|---|---|---|---|---|
| BGD1 | IC | 45.80 ± 13.00 | 0.994 ± 0.003 | 0.533 ± 0.060 | 1.746 ± 0.070 | 0.259 ± 0.076 |
| EG PCR− | 44.55 ± 11.68 | 0.994 ± 0.002 | 0.604 ± 0.242 | 1.748 ± 0.069 | 0.259 ± 0.076 | |
| EG PCR+ | 46.32 ± 16.71 | 0.996 ± 0.001 | 0.553 ± 0.073 | 1.749 ± 0.069 | 0.252 ± 0.073 | |
| Total | 45.52 ± 12.85 | 0.994 ± 0.002 | 0.563 ± 0.159 | 1.747 ± 0.070 | 0.258 ± 0.076 | |
| BGD26 | IC | 32.97 ± 5.45 | 0.979 ± 0.011 | 0.415 ± 0.034 | 1.746 ± 0.071 | 0.289 ± 0.082 |
| EG PCR− | 41.02 ± 9.86 | 0.988 ± 0.006 | 0.452 ± 0.139 | 1.749 ± 0.070 | 0.285 ± 0.083 | |
| EG PCR+ | 34.29 ± 8.26 | 0.989 ± 0.007 | 0.480 ± 0.059 | 1.750 ± 0.067 | 0.294 ± 0.082 | |
| Total | 35.84 ± 8.55 | 0.986 ± 0.009 | 0.454 ± 0.087 | 1.749 ± 0.069 | 0.290 ± 0.082 | |
| p values | 0.0049 | <0.0001 | <0.0001 | 0.6341 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauman, N.; Srbljanović, J.; Čolović Čalovski, I.; Lijeskić, O.; Ćirković, V.; Trajković, J.; Bobić, B.; Ilić, A.Ž.; Štajner, T. Structural Characterization of Toxoplasma gondii Brain Cysts in a Model of Reactivated Toxoplasmosis Using Computational Image Analysis. Fractal Fract. 2024, 8, 175. https://doi.org/10.3390/fractalfract8030175
Bauman N, Srbljanović J, Čolović Čalovski I, Lijeskić O, Ćirković V, Trajković J, Bobić B, Ilić AŽ, Štajner T. Structural Characterization of Toxoplasma gondii Brain Cysts in a Model of Reactivated Toxoplasmosis Using Computational Image Analysis. Fractal and Fractional. 2024; 8(3):175. https://doi.org/10.3390/fractalfract8030175
Chicago/Turabian StyleBauman, Neda, Jelena Srbljanović, Ivana Čolović Čalovski, Olivera Lijeskić, Vladimir Ćirković, Jelena Trajković, Branko Bobić, Andjelija Ž. Ilić, and Tijana Štajner. 2024. "Structural Characterization of Toxoplasma gondii Brain Cysts in a Model of Reactivated Toxoplasmosis Using Computational Image Analysis" Fractal and Fractional 8, no. 3: 175. https://doi.org/10.3390/fractalfract8030175