
Nitrate/Ammonium Ratios and Nitrogen Deficiency Impact on Nutrient Absorption and Photosynthetic Efficiency of Cedrela odorata


Abstract
:1. Introduction
2. Materials and Methods
2.1. Species and Growth Conditions
2.2. Treatments and Experimental Design
2.3. Specific Procedures
2.3.1. Soil Collection, Characterization, and Fertilization
2.3.2. Seedlings, Containers, Irrigation, and Topdressing Fertilization
2.4. Experiment Evaluation
2.5. Photosynthetic Parameters
2.5.1. Gas Exchange
2.5.2. Chlorophyll Fluorescence
2.5.3. Relative Chlorophyll Index (RCI)
2.6. Growth Parameters
2.6.1. Plant Height and Diameter
2.6.2. Dry Matter Production
2.7. Nutritional Parameters
Nutritional Status
2.8. Data Analysis
3. Results
3.1. Foliar Nutrient Contents and Accumulation
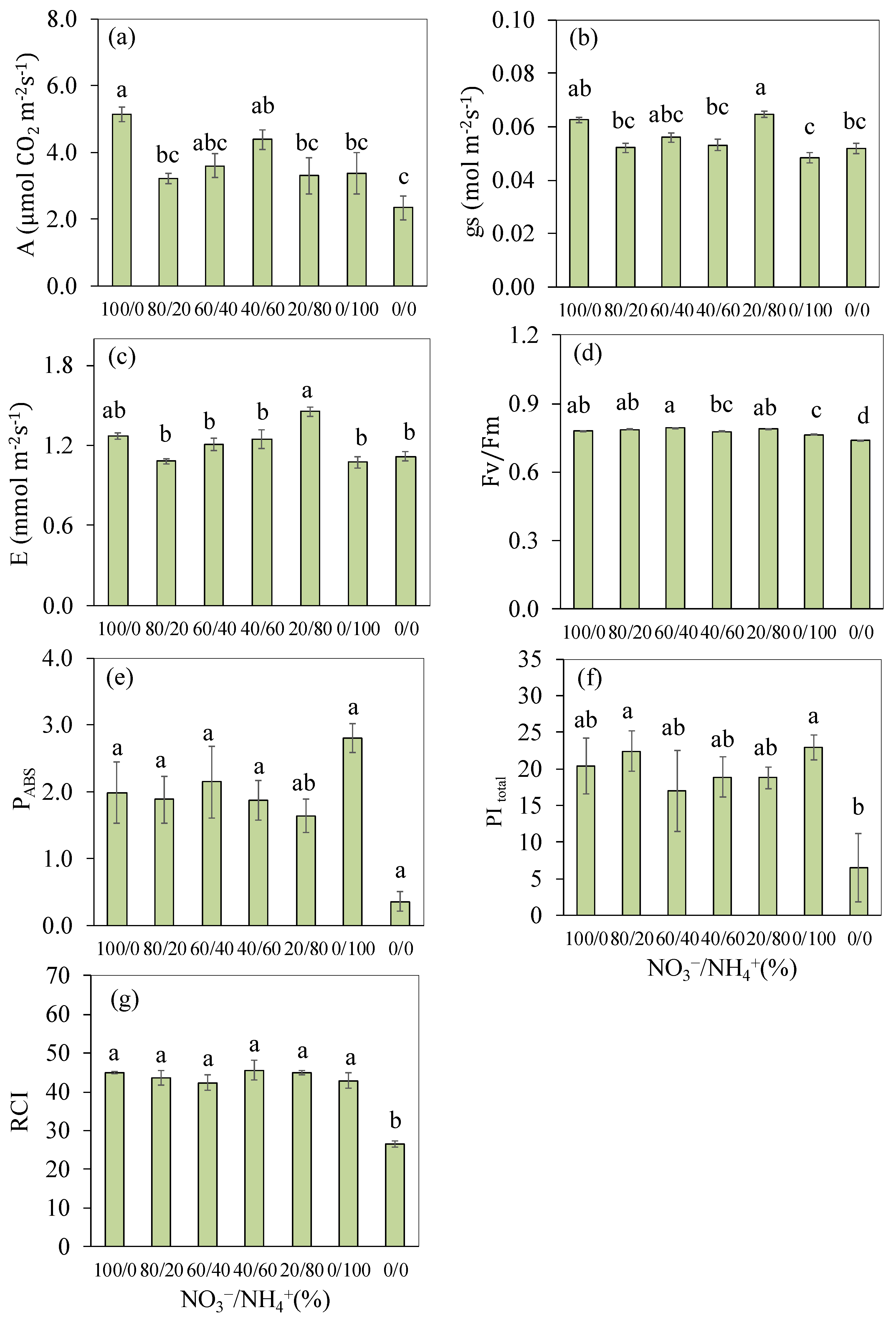
3.2. Photosynthetic Parameters
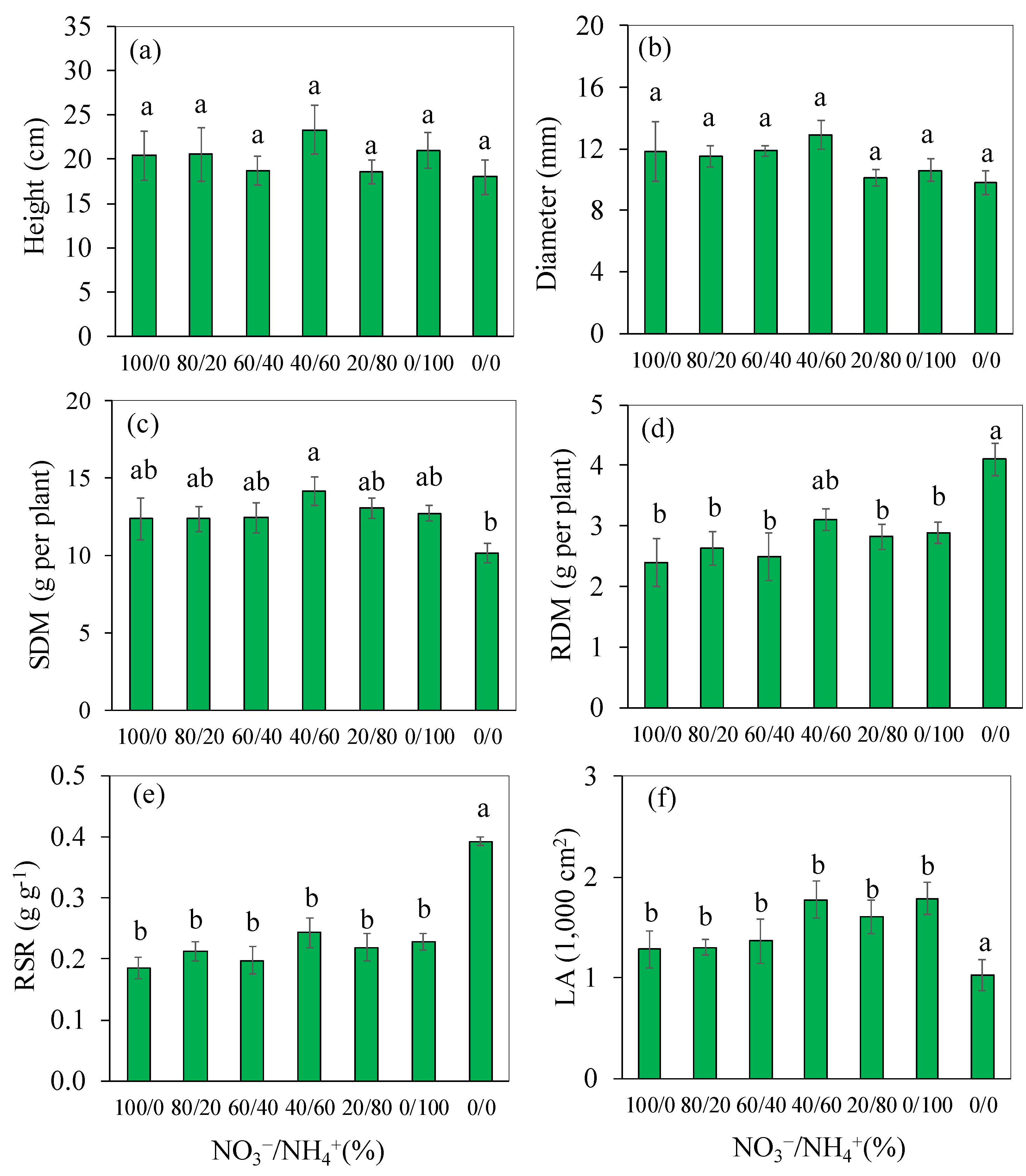
3.3. Plant Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | DF | N | P | K | Ca | Mg | S | Mn |
|---|---|---|---|---|---|---|---|---|
| Mean Square | ||||||||
| Treatment | 6 | 264.96 ** | 3.12 ** | 6.49 ns | 46.05 ** | 2.86 ** | 54.09 ** | 1466.45 ** |
| Error | 28 | 1.72 | 0.42 | 3.11 | 3.63 | 0.20 | 0.37 | 361.07 |
| CV (%) | - | 3.59 | 22.46 | 8.11 | 12.75 | 17.79 | 8.74 | 16.80 |
| Zn | B | Cu | Fe | N-ac | P-ac | K-ac | ||
| Treatment | 6 | 433.51 ** | 1909.71 ** | 4.56 * | 1278.59 ns | 0.047 ** | 0.000 ** | 0.004 ** |
| Error | 28 | 31.74 | 28.78 | 1.31 | 575.71 | 0.002 | 0.000 | 0.001 |
| CV (%) | - | 12.82 | 9.96 | 35.51 | 16.63 | 13.84 | 24.44 | 13.68 |
| Ca-ac | Mg-ac | S-ac | Mn-ac | Zn-ac | B-ac | Cu-ac | ||
| Treatment | 6 | 0.007 ** | 0.000 ** | 0.0024 ** | 0.304 ** | 0.061 ** | 0.082 ** | 0.0007 ** |
| Error | 28 | 0.001 | 0.000 | 0.0004 | 0.051 | 0.0088 | 0.005 | 0.0001 |
| CV (%) | - | 29.22 | 25.94 | 28.77 | 19.36 | 20.32 | 13.95 | 42.54 |
| Fe-ac | A | gs | E | Fv/Fm | PABS | PItotal | ||
| Treatment | 6 | 0.285 * | 4.05 ** | 0.000 ** | 0.09 ** | 0.000 ** | 2.72 ** | 152.60 ns |
| Error | 28 | 0.083 | 0.77 | 0.000 | 0.00 | 0.000 | 0.59 | 62.49 |
| CV (%) | - | 19.44 | 24.29 | 6.81 | 7.91 | 0.59 | 42.48 | 43.68 |
| RCI | Height | Diameter | SDM | RDM | RSR | LA | ||
| Treatment | 6 | 224.60 ** | 13.94 ns | 6.07 ns | 7.28 ns | 1.64 ** | 0.02 ** | 399,849.35 * |
| Error | 28 | 13.63 | 25.98 | 0.72 | 3.86 | 0.40 | 0.001 | 141,211.00 |
| CV (%) | - | 8.88 | 25.18 | 19.34 | 15.80 | 21.76 | 17.13 | 25.92 |
References
- Luchi, A.E. Quantitative feature so Cedrela odorata L. wood (Meliaceae). Braz. J. Bot. 2011, 34, 403–410. [Google Scholar] [CrossRef]
- Carrero, G.C.; Pereira, R.S.; Jacaúna, M.A.; Lima Junior, M.J. Trees of Amazon’s South: Guide of Species of Economic and Ecological Interest, 2nd ed.; IDESAM: Manaus, Brazil, 2014; 112p. [Google Scholar]
- Almeida, A.N.D.; Angelo, H.; Silva, J.C.G.L.D.; Hoeflich, V.A. Tropical sawn wood market: Substitution export demand. Acta Amaz. 2010, 40, 119–126. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 2012; p. 889. [Google Scholar]
- Peleja, V.L.; Peleja, P.L.; Lara, T.S.; Tribuzy, E.S.; de Moura, J.M.S. Seasonality and phosphate fertilization in carbohydrates storage: Carapa guianensis Aubl. Seedlings responses. Plants 2022, 11, 1956. [Google Scholar] [CrossRef] [PubMed]
- Freiberger, M.B.; Guerrini, I.A.; Galetti, G.; Fernandes, D.M.; Corrêa, J.C. Early growth and nutrition of cedar (Cedrela fissilis Vell.) as affected by nitrogen rates. Rev. Árvore 2013, 37, 385–392. [Google Scholar] [CrossRef]
- Jiang, L.; Tian, D.; Ma, S.; Zhou, X.; Xu, L.; Zhu, J.; Jing, X.; Zheng, C.; Shen, H.; Zhou, Z.; et al. The response of tree growth to nitrogen and phosphorus additions in a tropical montane rainforest. Sci. Tot. Environ. 2018, 618, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Bardy, L.R.; Debiasi, T.V.; Sanada, K.; Rondina, A.B.L.; Torezan, J.M.D.; Stolf-Moreira, R.; Bianchini, E.; Pimenta, J.A.; Oliveira, H.C. Effect of nitrogen addition to the soil on atlantic forest tree seedlings. Forests 2023, 14, 1111. [Google Scholar] [CrossRef]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting nitrogen metabolism and transport processes to improve plant nitrogen use efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar] [CrossRef]
- Al-Awad, O.A.S.; Prendergast, K.S.; Robson, A.; Rengel, Z. Screening canola genotypes for resistance to ammonium toxicity. Agronomy 2023, 13, 1150. [Google Scholar] [CrossRef]
- Castro-Rodríguez, V.; Cañas, R.A.; Torre, F.N.L.; Pascual, M.B.; Avila, C.; Cánovas, F.M. Molecular fundamentals of nitrogen uptake and transport in trees. J. Exp. Bot. 2017, 68, 2489–2500. [Google Scholar] [CrossRef]
- Hachiya, T.; Sakakibara, H. Interactions between nitrate and ammonium in their uptake, allocation, assimilation, and signaling in plants. J. Exp. Bot. 2017, 68, 2501–2512. [Google Scholar] [CrossRef]
- Huang, W.-T.; Xie, Y.-Z.; Chen, X.-F.; Zhang, J.; Chen, H.-H.; Ye, X.; Guo, J.; Yang, L.-T.; Chen, L.-S. Growth, mineral nutrients, photosynthesis and related physiological parameters of citrus in response to nitrogen deficiency. Agronomy 2021, 11, 1859. [Google Scholar] [CrossRef]
- Malavolta, E.; Moraes, M.F. Fundamentals of nitrogen and sulfur in the mineral nutrition of cultivated plants. In Nitrogen and sulfur in Brazilian Agriculture; Yamada, T., Abdalla, S.R.S., Vitti, G.C., Eds.; International Plant Nutrition Institute: Piracicaba, Brazil, 2007; pp. 189–249. [Google Scholar]
- Zhu, Y.; Qi, B.; Hao, Y.; Liu, H.; Sun, G.; Chen, R.; Song, S. Appropriate NH4+/NO3− ratio triggers plant growth and nutrient uptake of flowering Chinese cabbage by optimizing the pH value of nutrient solution. Front. Plant Sci. 2021, 12, 656144. [Google Scholar] [CrossRef]
- Araújo, J.L.; Faquin, V.; Vieira, N.M.B.; Oliveira, M.W.C.; Soares, A.A.; Rodrigues, C.R.; Mesquita, A.C. Rice growth and yield at different nitrate-ammonium ratios. Rev. Bras. Ciênc. Solo 2012, 36, 921–930. [Google Scholar] [CrossRef]
- Shi, L.; Liang, J.; Wang, R.; Wan, X.; Yan, B.; Zhang, Y.; Chen, M.; Liu, C.; Li, Q.; Wang, S.; et al. The ammonium/nitrate ratio affects the growth and shikonin accumulation in Arnebia euchroma. Agronomy 2023, 13, 1318. [Google Scholar] [CrossRef]
- Ying, J.; Li, X.; Wang, N.; Lan, Z.; He, J.; Bai, Y. Contrasting effects of nitrogen forms and soil pH on ammonia oxidizing microorganisms and their responses to long-term nitrogen fertilization in a typical steppe ecosystem. Soil Biol. Biochem. 2017, 107, 10–18. [Google Scholar] [CrossRef]
- Barrow, N.J.; Hartemink, A.E. The effects of pH on nutrient availability depend on both soils and plants. Plant Soil 2023, 487, 21–37. [Google Scholar] [CrossRef]
- Li, S.X.; Wang, Z.H.; Stewart, B.A. Responses of crop plants to ammonium and nitrate N. In Advances in Agronomy; Sparks, D.L., Ed.; Elsevier Academic Press: Waltham, MA, USA, 2013; pp. 205–398. [Google Scholar] [CrossRef]
- Máximo, W.P.F.; Santos, P.A.A.; Mendonça, E.G.; Santos, B.R.; Paiva, L.V. Nitrate (NO3−) and ammonium (NH4+) ratios for propagation of Eucalyptus hybrid in two different in vitro cultivation systems. Aust. J. Crop Sci. 2015, 12, 1242–1248. [Google Scholar]
- Shilpha, J.; Song, J.; Jeong, B.R. Ammonium phytotoxicity and tolerance: An insight into ammonium nutrition to improve crop productivity. Agronomy 2023, 13, 1487. [Google Scholar] [CrossRef]
- Chen, H.; Jia, Y.; Xu, H.; Wang, Y.; Zhow, Y.; Huang, Z.; Yang, L.; Li, Y.; Chen, L.-S.; Guo, J. Ammonium nutrition inhibits plant growth and nitrogen uptake in citrus seedlings. Sci. Hortic. 2020, 272, 109526. [Google Scholar] [CrossRef]
- Epstein, E.; Bloom, A. Mineral Plant Nutrition: Principles and Perspectives; Planta: Londrina, Brazil, 2006; pp. 184–200. [Google Scholar]
- Mantovani, C.; Prado, R.M.; Pivetta, K.F.L. Impact of nitrate and ammonium ratio on nutrition and growth of two epiphytic orchids. Ann. Braz. Acad. Sci. 2018, 90, 3423–3431. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, B.; Ni, X.; He, Y. Ammonium/nitrate ratio affects the growth and glucosinolates content of pakchoi. Hort. Bras. 2020, 38, 246–253. [Google Scholar] [CrossRef]
- Fageria, N.K. Soil fertility and plant nutrition research under controlled conditions: Basic principles and methodology. J. Plant Nutr. 2005, 28, 1975–1999. [Google Scholar] [CrossRef]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef] [PubMed]
- Cervi, A.C.; Linsingen, L.V.; Patricio, C.P. The genus Cedrela P. Browne (Meliaceae) in southern Brazil. Acta Biol. Parana. 2008, 37, 105–110. [Google Scholar] [CrossRef]
- Quesada, C.A.; Lloyd, J.; Anderson, L.O.; Fyllas, N.M.; Schwarz, M.; Czimczik, C.I. Soils of Amazonia with particular reference to the RAINFOR sites. Biogeosciences 2011, 8, 1415–1440. [Google Scholar] [CrossRef]
- Moreira, A.; Fageria, N.K. (2009) Soil Chemical Attributes of Amazonas State, Brazil. Commun. Soil Sci. Plant Anal. 2009, 40, 2912–2925. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; Mcmahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Soil Survey Staff. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; USDA-NRCS: Washington, DC, USA, 1999. [Google Scholar]
- EMBRAPA. Manual of Chemical Analysis of Soils, Plants, and Fertilizers; EMBRAPA Tecnological Information: Brasilia, Brazil, 2009. [Google Scholar]
- Allen, S.E.; Terman, G.L.; Clements, L.B. Greenhouse Techniques for Soil-Plant-Fertilizer Research; National Fertilizer Development Center: Muscle Shoals, AB, USA, 1976; p. 55. [Google Scholar]
- Malavolta, E. Elements of Mineral Nutrition of Plants; Agronômica Ceres: São Paulo, Brazil, 1980; p. 251. [Google Scholar]
- Melém Junior, N.; Mazza, A.; Dias, C.T.S.; Briske, E.G. Influence of nitrogen fertillzers on the acidification of a tipic paleudult soil cultivated with corn. Ciênc. Tecnol. 2001, 2, 75–89. [Google Scholar]
- Freire, J.C.; Ribeiro, M.A.V.; Bahia, V.G.; Lopes, A.S.; Aquino, L.E. Maize response in a greenhouse experiment to water levels in Lavras soils. Braz. J. Soil Sci. 1980, 4, 5–8. [Google Scholar]
- Santos Júnior, U.M.; Gonçalves, J.F.C.; Feldpausch, T.R. Growth, leaf nutrient concentration and photosynthetic nutrient use efficiency in tropical tree species planted in degraded areas in Central Amazonia. For. Ecol. Manag. 2006, 226, 299–309. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic chlorophyll a fluorescence transients in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Giang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Evaluation of Nutritional Plant Status: Principles and Applications, 2nd ed.; Potafos: Piracicaba, Brazil, 1997; p. 319. [Google Scholar]
- Rehmus, A.; Bigalke, M.; Valarezo, C.; Castillo, J.M.; Wilcke, W. Aluminum toxicity to tropical montane forest tree seedlings in southern Ecuador: Response of nutrient status to elevated Al concentrations. Plant Soil 2015, 388, 87–97. [Google Scholar] [CrossRef]
- Bittsánszky, A.; Pilinszky, K.; Gyulai, G.; Komives, T. Overcoming ammonium toxicity. Plant Sci. 2015, 231, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Figueiredo, F.A.M.A.; Carneiro, J.G.A.; Penchel, R.M.; Campostrini, E.; Thiebaut, J.T.L.; Barroso, D.G. Gas exchanges, water relations and photochemical efficiency in clonal rooted cuttings with prolonged times in tubes. Sci. For. 2014, 42, 533–542. [Google Scholar]
- Marenco, R.A.; Antezana-Vera, A.S.; Gouvêa, P.R.S.; Camargo, M.A.B.; Oliveira, M.F.; Santos, J.K.S. Physiology of Amazon tree species: Photosynthesis, respiration and water relations. Rev. Ceres 2014, 61, 786–789. [Google Scholar] [CrossRef]
- Ueda, Y.; Konishi, M.; Yanagisawa, S. Molecular basis of the nitrogen response in plants. Soil Sci. Plant Nutr. 2017, 63, 1–13. [Google Scholar] [CrossRef]
- Gonçalves, E.O.; Paiva, H.N.; Neves, J.C.L.; Gomes, J.M. Nutrition of red angico seedlings (Anadenanthera macrocarpa (Benth.) Brenan) under different macronutrient doses. J. Braz. For. Sci. 2012, 36, 219–228. [Google Scholar] [CrossRef]
- Moretti, B.S.; Furtini Neto, A.E.; Pinto, S.I.C.; Furtini, I.V.; Magalhães, C.A.S. Growth and mineral nutrition in seedlings of australian cedar (Toona ciliata) subjected to nutrient deprivation. Cerne 2011, 17, 453–463. [Google Scholar] [CrossRef]
- Guimarães, M.M.C.; Cairo, P.A.R.; Neves, O.S.C. Growth of Eucalyptus urophylla in hydroponic medium with different ratios of nitrate and ammonium. For. Environ. 2014, 21, 52–61. [Google Scholar] [CrossRef]
- Paiva, P.J.R.; Nicodemo, M.L.F. Sulfur in the Soil-Plant-Animal System; EMBRAPA-CNPGC: Campo Grande, MS, USA, 1994; pp. 16–19. [Google Scholar]
- Li, Q.; Gao, Y.; Yang, A. Sulfur homeostasis in plants. Int. J. Mol. Sci. 2020, 21, 8926. [Google Scholar] [CrossRef] [PubMed]
- Bont, L.; Donnay, N.; Couturier, J.; Rouhier, N. Redox regulation of enzymes involved in sulfate assimilation and in the synthesis of sulfur-containing amino acids and glutathione in plants. Front. Plant Sci. 2022, 13, 958490. [Google Scholar] [CrossRef]
- Narayan, O.P.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur nutrition and its role in plant growth and development. Plant Sign. Behav. 2022, 18, 2030082. [Google Scholar] [CrossRef] [PubMed]
- Piwpuan, N.; Jampeetong, A.; Brix, H. Ammonium tolerance and toxicity of Actinoscirpus grossus—A candidate species for use in tropical constructed wetland systems. Ecotoxicol. Environ. Saf. 2014, 107, 319–328. [Google Scholar] [CrossRef]
- Tsabarducas, V.; Chatzistathisa, T.; Theriosa, I.; Patakas, A. How nitrogen form and concentration affect growth, nutrient accumulation and photosynthetic performance of Olea europaea L. (cv. ‘Kalamon’). Sci. Hortic. 2017, 218, 23–29. [Google Scholar] [CrossRef]
- Bown, H.E.; Watt, M.S.; Clinton, P.W.; Mason, E.G. Influence of ammonium and nitrate supply on growth, dry matter partitioning, N uptake and photosynthetic capacity of Pinus radiata seedlings. Trees 2010, 24, 1097–1107. [Google Scholar] [CrossRef]
- Rufty Junior, T.W.; Huber, S.C.; Volk, R.J. Alterations in leaf carbohydrate metabolism in response to nitrogen stress. Plant Physiol. 1988, 88, 725–730. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2013. [Google Scholar]
- Desotgiu, R.; Pollastrini, M.; Cascio, C.; Gerosa, G.; Marzuoli, R.; Bussotti, F. Chlorophyll a fluorescence analysis along a vertical gradient of the crown in a poplar (Oxford clone) subjected to ozone and water stress. Tree Physiol. 2012, 32, 976–986. [Google Scholar] [CrossRef]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, X.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 13389. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, K.; Zhang, Z.; Cao, Q.; Lv, Z.; Yuan, Z.; Tian, Y.; Cao, W.; Zhu, Y. Canopy chlorophyll density based index for estimating nitrogen status and predicting grain yield in rice. Front. Plant Sci. 2017, 8, 1829. [Google Scholar] [CrossRef]
- Jia, M.; Colombo, R.; Roscsini, M.; Celesti, M.; Zhu, J.; Cogliati, S.; Cheng, T.; Tian, Y.; Zhu, Y.; Cao, W.; et al. Estimation of leaf nitrogen content and photosynthetic nitrogen use efficiency in wheat using sun-induced chlorophyll fluorescence at the leaf and canopy scales. Eur. J. Agron. 2021, 122, 126192. [Google Scholar] [CrossRef]
- Qin, L.; Walk, T.C.; Han, P.; Chen, L.; Zhang, S.; Li, Y.; Hu, X.; Xie, L.; Yang, Y.; Liu, J.; et al. Adaption of roots to nitrogen deficiency revealed by 3D quantification and proteomic analysis. Plant Physiol. 2019, 179, 329–347. [Google Scholar] [CrossRef]




| NH4+/NO3− (%) | N (g kg−1) | P (g kg−1) | K (g kg−1) | Ca (g kg−1) | Mg (g kg−1) | S (g kg−1) |
|---|---|---|---|---|---|---|
| 100/0 | 41.00 ± 1.00 a | 3.00 ± 0.00 abc | 20.00 ± 1.41 a | 15.00 ± 2.00 ab | 3.00 ± 0.50 a | 4.00 ± 0.60 e |
| 80/20 | 41.0.0 ± 1.41 a | 3.60 ± 0.55 a | 22.20 ± 2.68 a | 15.40 ± 2.07 ab | 2.80 ± 0.45 ab | 5.40 ± 0.55 cd |
| 60/40 | 39.40 ± 1.95 ab | 2.20 ± 0.84 bc | 21.20 ± 1.30 a | 9.80 ± 5.60 c | 1.00 ± 0.71 c | 5.00 ± 1.00 de |
| 40/60 | 39.00 ± 2.12 ab | 3.40 ± 0.89 ab | 23.20 ± 2.39 a | 16.80 ± 0.84 a | 3.00 ± 0.10 a | 7.00 ± 1.00 b |
| 20/80 | 38.20 ± 0.84 b | 2.00 ± 0.71 c | 21.80 ± 1.30 a | 12.20 ± 2.49 bc | 2.00 ± 0.10 b | 7.00 ± 2.00 b |
| 0/100 | 37.40 ± 0.89 b | 4.00 ± 0.71 a | 23.00 ± 1.22 a | 16.60 ± 1.40 a | 2.80 ± 0.84 ab | 6.40 ± 0.55 cd |
| 0/0 | 20.40 ± 0.55 c | 2.20 ± 0.45 bc | 21.00 ± 1.41 a | 18.80 ± 0.45 a | 3.0 0 ± 0.10 a | 14.00 ± 1.00 a |
| - | Mn (mg kg−1) | Zn (mg kg−1) | B (mg kg−1) | Cu (mg kg−1) | Fe (mg kg−1) | |
| 100/0 | 99.0 ± 7.4 b | 45.0 ± 1.2 a | 45.0 ± 2.0 b | 2.0 ± 0.0 b | 177.0 ± 5.5 a | |
| 80/20 | 127.0 ± 2 3.0 ab | 46.0 ± 3.4 b | 52.0 ± 3.0 b | 2.4 ± 0.9 b | 142.4 ± 37.4 ab | |
| 60/40 | 115.6 ± 20.7 ab | 61.2 ± 11 b | 41.4 ± 7.8 b | 4.8 ± 2.5 a | 138.4 ± 12.9 ab | |
| 40/60 | 116.6 ± 1.81 ab | 44.0 ± 1.6 b | 47.2 ± 0.8 b | 4.0 ± 1.0 ab | 143.2 ± 22.6 ab | |
| 20/80 | 132.0 ± 17.8 a | 39.8 ± 1.1 b | 47.0 ± 0.7 b | 3.4 ± 0.5 ab | 142.8 ± 27.2 ab | |
| 0/100 | 119.0 ± 25.8 ab | 41.6 ± 9.4b | 46.8 ± 4.3 b | 3.2 ± 0.8 ab | 142.6 ± 21.9 ab | |
| 0/0 | 82.2 ± 14.3 | 30.0 ± 2.1 c | 97.6 ± 0.6 a | 2.8 ± 0.4 ab | 123.8 ± 26.5 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosado, S.I.P.; Santos, J.Z.L.; Saraiva, I.S.D.A.; dos Santos, N.J.R.; Barbosa, T.M.B.; Araujo, J.L. Nitrate/Ammonium Ratios and Nitrogen Deficiency Impact on Nutrient Absorption and Photosynthetic Efficiency of Cedrela odorata. Nitrogen 2024, 5, 1-15. https://doi.org/10.3390/nitrogen5010001
Rosado SIP, Santos JZL, Saraiva ISDA, dos Santos NJR, Barbosa TMB, Araujo JL. Nitrate/Ammonium Ratios and Nitrogen Deficiency Impact on Nutrient Absorption and Photosynthetic Efficiency of Cedrela odorata. Nitrogen. 2024; 5(1):1-15. https://doi.org/10.3390/nitrogen5010001
Chicago/Turabian StyleRosado, Sulianne Idalior Paião, José Zilton Lopes Santos, Ives San Diego Amaral Saraiva, Nonato Junior Ribeiro dos Santos, Tainah Manuela Benlolo Barbosa, and Josinaldo Lopes Araujo. 2024. "Nitrate/Ammonium Ratios and Nitrogen Deficiency Impact on Nutrient Absorption and Photosynthetic Efficiency of Cedrela odorata" Nitrogen 5, no. 1: 1-15. https://doi.org/10.3390/nitrogen5010001