
Effects of Seasonal Variation on Nitrogen Use in Brazilian Cerrado Grass Communities
by
 , , and
, , and
Victor Camargo Keller
1,*,
Erico Fernando Lopes Pereira-Silva
2,3,* ,
,
Sergio Tadeu Meirelles
3 and
Elisa Hardt
1 1
Group of Landscape Ecology and Conservation Planning (LEPLAN), Department of Environmental Sciences, Universidade Federal de São Paulo, Rua Prof. Artur Riedel, 275, Diadema 09972-270, São Paulo State, Brazil
2
Center for Natural Sciences and Humanities, Universidade Federal do ABC, Rua Arcturus 03, Jardim Antres, São Bernardo do Campo 09606-070, São Paulo State, Brazil
3
Department of Ecology, Universidade de São Paulo, Rua do Matão, Trav. 14, 321, Cidade Universitária, São Paulo 05508-090, São Paulo State, Brazil
*
Authors to whom correspondence should be addressed.
Nitrogen 2024, 5(2), 373-385; https://doi.org/10.3390/nitrogen5020024
Submission received: 25 March 2024
/
Revised: 19 April 2024
/
Accepted: 22 April 2024
/
Published: 24 April 2024
(This article belongs to the Special Issue Nitrogen Signaling in Plants)
Abstract
:In savanna ecosystems, the seasonal effects of nitrogen forms and availability, as well as their utilization by plants, influence the abundance and distribution of herbaceous species in grassland communities. This study examines seasonal effects on nitrogen availability and utilization by native grass species in the Cerrado, a savanna ecosystem in Brazil. Ammonium and nitrate levels in soil, nitrate acquisition and transport, and Nitrate Reductase Activity (NRA) in different plant parts during dry and wet periods were assessed. Results indicated higher soil nitrate availability during the wet period, influenced by precipitation, with leaves showing a higher nitrate content compared to roots. There was seasonal modulation in nitrate reduction, with leaves being the primary site during the dry period and roots during the wet period. The studied grass species exhibited heterogeneous responses to seasonal nitrogen availability, potentially affecting community abundance patterns. Findings suggest that edaphoclimatic seasonality plays a crucial role in nitrogen distribution and utilization capacity by grass plants in the Cerrado, contributing to the understanding of these ecosystems’ ecology.
1. Introduction
Nitrogen (N) is an essential nutrient and is often a limiting factor for plant growth both in natural and cultivated landscapes [1]. This element is present in soil in organic forms, such as humic substances and amino acids, as well as in inorganic forms, including nitrate (NO3−) and ammonium (NH4+), which can be preferentially absorbed and assimilated by vegetation due to their high soil concentration and play a fundamental role in soil structure and fertility, contributing to nutrient availability for plants [2,3,4,5,6]. Plants can use different N forms, isolated or in combination, favoring biomass production and accumulation [7,8,9]. The NO3− and NH4+ availability for plant assimilation is directly related to the soil's physical, chemical, and biological attributes, as well as its water saturation levels [10]. Total N content tends to be low on old soil exposed to millions of years of weathering [11], like the soils in tropical regions of Brazil.
Cerrado soils are naturally acidic (pH < 5.0) and have low N availability [12,13], prevailing high NH4+ concentrations up to eleven times higher than NO3− levels [14]. When NH4+ concentration is high in the soil, plants can absorb it passively through the roots [15], promoting rhizosphere acidification through H+ leakage, favoring phosphorus capture by the roots [16], and reducing aluminum toxicity [17]. The absorbed NH4+ can be converted locally on the roots or transported to the leaves, where it can be converted to amino acids [15,18].
On calcareous (pH > 6.0) and well-aerated soils, like the ones in Mediterranean ecosystems [19], NO3− is the main inorganic N form available. Nitrate is highly soluble, being easily mobilized for the roots by mass flux and diffusion when water is available [20]. Nitrate can be assimilated on roots or transported via xylem, being accumulated or assimilated on the aerial parts [15,20,21]. To be assimilated, NO3− must be converted to NO2− by Nitrate Reductase (NR), an enzyme present in the cytosol, which is then converted to NH4+ by Nitrite Reductase (NiR) on chloroplasts and plastids [15,18]. Nitrate's high solubility favors its soil percolation, increasing its concentration in deep soil layers far from the superficial roots [22].
Because there are different energetic costs for NO3− or NH4+ absorption and assimilation [18,23,24], these processes are fine-regulated according to each available N form in the soil, its concentration inside the plant, and the availability of energy and carbon skeletons for amino acid production [15]. Nitrate Reductase Activity (NRA) is favored by the presence of light and substrate for reduction (NO3−) [25]. Ammonium assimilation is also promoted by the presence of light and sugars from photosynthesis [26]. Since water presence is essential both for NO3− uptake [20] and for the good function of the photosynthetic system [27], plant and soil hydraulic states can influence N absorption and assimilation, mainly for NO3−.
Soil fertilization in natural communities tends to increase local productivity [1,28] but can lead to species loss [28,29]. Changes in NH4+ and NO3− soil availability can alter the competitive balance between species [6,30,31], favoring biological invasions and the dominance of undesired species in the community [32,33]. Additionally, the loss of plant diversity, along with an increase in nitrogen concentration, can lead to greater NO3− percolation, contaminating water sheets and streams close to the affected community [22].
Plants' evolutionary history can also influence their ability to assimilate different nitrogen forms. Studies have evaluated relations between NRA and the vegetation in which one species occurs [34], its successional stage [5,35,36,37,38,39], below-ground functional traits [19], and its habit and capacity to fix nitrogen [21]. These papers suggest that NO3− reduction is higher on aerial parts of plants, with a high leaf NRA activity on dicotyledonous, herbaceous, heliophiles, and pioneering plants [21]. Despite this, none of these studies measure NRA on Cerrado’s native grasses, one of the most species-rich groups of open savannas.
Hence, the main goal of this study is to characterize nitrogen availability and its use by Cerrado’s native grasses, as well as to check for a climatic seasonality effect on nitrogen presence and use by native grasses. We also evaluated if there is an association between nitrogen use abilities and grass abundance in a natural grassland community. To access this information, we studied seven native grass species on their nitrate assimilation and content on roots and leaves over the dry and wet seasons. By studying these relationships, we intend to expand our knowledge of Cerrado’s native grass ecophysiology and its effects on grassland community assemblage.
2. Material and Methods
2.1. Study Site and Data Collection
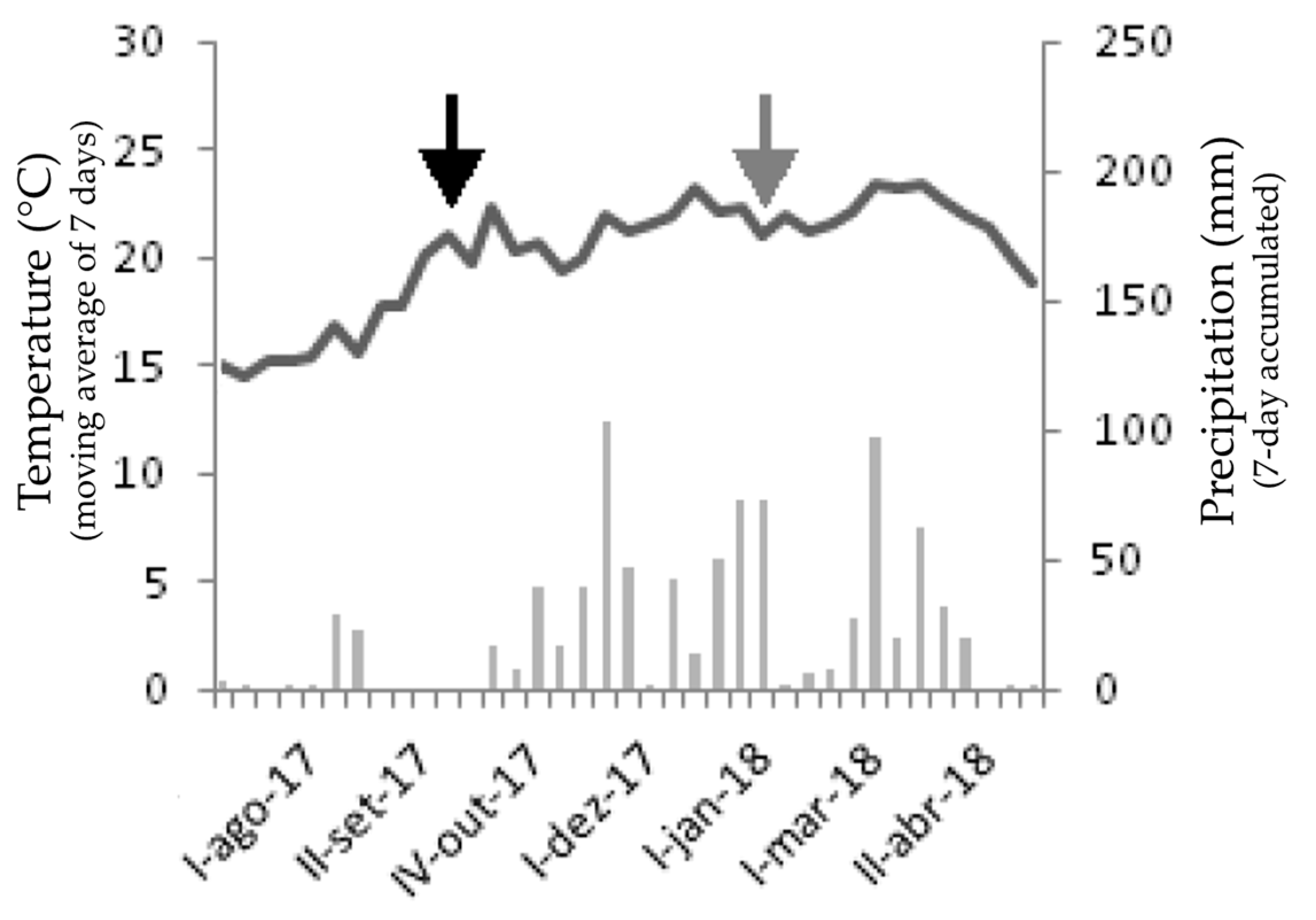
Our study was conducted at Juquery State Park (PEJY), a 2058 ha conservation unit covered with Atlantic Forest and grasslands, located at Franco da Rocha, São Paulo metropolitan region, Brazil. PEJY is located between 730 and 950 m above sea level, with a mean annual precipitation between 1200 and 2000 mm and a mean temperature range between 20 and 21 °C [40], with a Cfb Köppen–Geiger climate [41]. No rainfall was recorded during the dry period in the two weeks leading up to the collections, while the accumulated precipitation during the wet period totaled 137.9 mm (Figure 1). PEJY is the last grassland remnant of the original vegetation cover in the Sao Paulo metropolitan region, with an extremely rich and diverse plant community [42].
The plant and soil collection site is a flat hilltop, 810 m above sea level, within a 50 m radius of the coordinates 23°20′54.5″ S, 46°42′19.3″ O. This site has a homogeneous grassland cover, with herbaceous and sub-shrub species prevailing over a red–yellow/yellow argisol with the presence of philites and metasiltites [43,44].
We selected seven grass species (Table 1) with high or low abundance at the PEJY grasslands based on their community importance value (IV), which is a sum of their frequency (the number of plots the species occurred divided by the total number of plots × 100) and coverage (the sum of the canopy area of all individuals of a species divided by the total canopy area × 100) [45].
For each species, we collected five full-grown specimens out of the reproductive period for the dry season (27 September 2017) and five for the wet season (11 January 2018) (Figure 1). Individuals were extracted between 6:00 and 9:00 a.m. with their soil bulk attached to the roots. Samples were chilled in thermal cases with ice for transportation to the lab.
2.2. Edaphoclimatic Characterization
Local temperature, rainfall, and air relative humidity were obtained by a WatchDog® 2000 (Spectrum Technologies, Aurora, CO, USA) climatic station located at PEJY’s visitors center (data ceded by Meirelles, S. T.). We characterized the local climate at the moment of plant and soil collection based on these data.
Soil nitrate, calcium, and potassium content at the rhizosphere were determined using Horiba® LaquaTwin (Singapore) sensors, both in dry (7 samples—one for each species) and wet (35 samples—five for each species) seasons. Measures of pH (on water solution—pHH2O, and on soil solution—pHss) and conductivity were made with portable PeakTech® (PeakTech Prüf, Ahrensburg, Germany) sensors. To determine soil ammonium content, we used the phenol methodology [46], measured by spectrophotometry (λ = 625 nm). Absorbance values were converted to mg g−1 of fresh weight (FW) via one NH4Cl calibration curve.
Soil chemical fertility at the studied site was characterized through one compound soil sample for each plot of the superficial soil layer (0–10 cm depth), where most of the nutrients are located. This approach ensured a representative evaluation of soil fertility across the study area while minimizing variability within plots. This soil analysis was made by the Agronomical Institute of Campinas (IAC).
2.3. Plant Nitrogen Analysis
The determination of Nitrate Reductase (NADH) enzyme activity (ANR NRA—EC 1.7.1.1) was evaluated in vivo in leaves and roots following the method used by [47,48]. Plants were washed and chopped into pieces smaller than 1 cm. Fresh subsamples of leaves and roots from each specimen were used. These samples were separated into replicates of 0.1 g aliquots of fresh material and transferred to test tubes. The samples were then dark vacuum-incubated in a buffer solution (KH2PO4 0.05 M, pH = 7.5; 30 °C for 60 min) containing 1% (v/v) 1-propanol and 0.1 M KNO3. After incubation, the assay mixture was analyzed for nitrite formation using the Saltzman method [49]. A total of 1 mL of mM sulfanilamide and 1 mL of mM n-naphtylethylenediaminedihidrochloric solution were added to 1 mL of the assay mixture, and the induced azo dye was determined by spectrophotometry (λ = 540 nm) (Varian Cary® 50 UV–Vis spectrophotometer, Agilent Technologies, Inc., Santa Clara, CA, USA). The absorbance readings were then converted to pkat g−1FW (fresh weight), considering the incubation time, the concentration of the reagents, and the amount of plant material used.
For the determination of N–NO3− content, fresh leaf and root material prepared in MCW solution (methanol, chloroform, and water, in the ratio 12:5:3 v/v) were used, kept for 24 h at room temperature, and then stored in a freezer for later analysis, as described by [50]. N–NO3− content was determined using the salicylic acid nitration method [51] with spectrophotometer readings (λ = 410 nm) and conversion of the absorbance value to mg g−1 FW from a calibration curve with KNO3 solution.
2.4. Statistical Data Analysis
After checking the normality and homoscedasticity of the data by Shapiro–Wilk and Levene tests, respectively, tests were performed for comparisons between periods (dry and wet), compartments (leaf and root), and their interactions. For normal data, an ANOVA with Tukey’s a posteriori test was used. When the assumptions of normality and homoscedasticity were not met, Kruskal–Wallis and Scheirer–Ray–Hare’s H test, a nonparametric 2-factor analysis of variance [52], with Mann–Whitney’s a posteriori test, was applied. These analyses were performed in the virtual environment R [53], considering significance level α = 0.05, with the packages provided by Mangiafico S. S. (2016).
3. Results
3.1. Edaphoclimatic Characterization
It was possible to identify a dry period between August and October and a rainy period between October and April (Figure 1), highlighting that field samplings were conducted at representative moments of the region’s seasonal climate. In the fortnights preceding the collections, no rainfall was recorded during the dry period, and the accumulated precipitation during the wet period amounted to 137.9 mm.
Based on the provided data, relative frequency (RF) and relative cover (RC) are important measures in understanding the distribution and significance of species within a campo sujo environment (PEJY). For instance, the species Echinolaena inflexa, belonging to the tribe Paniceae, exhibits a high RC of 25.09 and an even more significant RF of 4.62, indicating not only frequent occurrence but also considerable coverage within the studied campo sujo.
The soil chemical fertility results showed low organic matter content (O.M. = 24 g/dm3), low phosphorus concentration (p = 2 mg/dm3), and high iron concentration (Fe = 64 mg/dm3) [55]. The potential acidity (H+Al) was 36 mmolc/dm3, the sum of bases (S.B.) was 11.4 mmolc/dm3, the cation exchange capacity (C.E.C.) was 47.4 mmolc/dm3, and the base saturation (V) was 24%, indicating an oligotrophic character for the soils in the study area.
The soil samples in the rhizosphere of the studied individuals showed an acidic character (pHH2O < 4), with low Ca2+ and medium–low K+ concentrations (Table 2). Ammonium content was higher than nitrate content in both the dry and wet seasons, with an ammonium/nitrate ratio greater than 2 in both seasons. Nitrate (F = 67.7; p < 0.001) and calcium (F = 17.6; p = 0.002) showed significantly higher values in the wet season. No significant differences in soil properties were observed between samples collected in the rhizosphere of the different species (Table 2).
The PCA of soil variables from these samples did not allow relationships to be established between the species studied and soil nutrient availability since there was no clustering of samples as a function of species (Figure 2A). The analysis shows little contribution of N–NO3− and a correlation between conductivity values and the concentration of Ca2+ (r = 0.52, p = 0.001) and K+ (r = 0.5, p = 0.001).
3.2. Nitrogen Usage by Native Grasses
The results of the analysis of variance with two factors (season and compartment) indicated no significant differences in NRA between the dry and wet seasons and between the aerial and underground plant compartments, although they did indicate an interaction between the two factors (Table 3). In the dry season, there were higher values in leaves (54.29 ± 5.33 pkat g−1FW) than in roots (29.66 ± 2.47 pkat g−1FW), while in the wet season, the values in roots (54.28 ± 5.49 pkat g−1FW) and leaves (46.72 ± 7.64 pkat g−1FW) did not differ significantly (Figure 3). Regarding the concentration of N–NO3−, the difference was only between the compartments, being always higher in the leaves (dry: 3.71 ± 0.17 mg g−1FW; wet: 2.91 ± 0.21 mg g−1 FW) than in the roots (dry: 0.94 ± 0.11 mg g−1 FW; wet: 0.99 ± 0.1 mg g−1 FW) (Table 3 and Figure 3).
The highest NRA values in the dry period were observed in the species E. inflexa (leaf = 94.57 ± 9.23 pkat g−1 FW; root = 45.85 ± 2.74 pkat g−1 FW) and A. recurvata (leaf = 85.77 ± 6.32 pkat g−1 FW; root = 45.96 ± 4.34 pkat g−1 FW) (Figure 3). The grass A. jubata showed the highest NRA values in the wet period (leaf = 137.9 ± 10.4 pkat g−1 FW; root = 100.68 ± 15.14 pkat g−1 FW). On the other hand, the lowest NRA values were observed in the A. lanata species in both the dry period (leaf = 23.8 ± 3.29 pkat g−1 FW; root = 12.04 ± 1.78 pkat g−1 FW) and the wet period (leaf = 10.76 ± 0.95 pkat g−1 FW; root = 26.86 ± 2.55 pkat g−1 FW) (Figure 3).
The same analysis of variance for NRA applied to each species indicated that only three had significant differences between the periods or between the compartments (Table 3). Between periods, in A. jubata and T. leiostachya, there was higher NRA in the wet period in both compartments, while in A. pressus, the highest NRA occurred in the dry period, both in leaves and roots (Figure 3). Between compartments, in both periods, there was higher NRA in the leaves of A. jubata and E. inflexa and in the roots of A. siccus (Figure 3). Of the species that showed significant interaction between factors, A. recurvata and A. lanata showed higher NRA in the leaves during the dry period, while in the wet period, the highest activity occurred in the roots (Figure 3).
Regarding the concentration of N–NO3− in plants, the comparison between compartments showed a definite pattern, being significantly higher in leaves than in roots in all species (Table 3). Only in A. siccus was a significant effect of the period observed, with a higher N–NO3− concentration in the dry period (Figure 3). The C3 grass E. inflexa showed the highest values of nitrate content in leaves during the wet period (mean = 4.21 ± 0.11 mg g−1 FW). On the other hand, the highest nitrate values in the root were observed in A. lanata in both the dry (mean = 1.83 ± 0.24 mg g−1 FW) and wet periods (mean = 1.69 ± 0.46 mg g−1 FW; Figure 3).
The PCA of the NRA and N–NO3− variables allowed the establishment of relationships between the studied species and nitrogen use since it aggregated most individuals of the same species (Figure 2B). Only the species A. recurvata and E. inflexa overlapped in the two-dimensional space, indicating similarities in nitrogen use patterns. The period (dry and wet) was more important for the NRA dispersal of individuals, while the compartment (leaf and root) was relevant for N–NO3−. In both periods, NRA in leaf and root were correlated and, together with N–NO3− content in roots, polarized the species E. inflexa and A. recurvata (high NRA and low N–NO3− concentration in roots) and A. lanata and A. siccus (low NRA and high N–NO3− concentration in roots) on opposite sides of Axis 1 (Figure 2B). NRA in the wet period had great weight in segregating the group of A. jubata individuals.
4. Discussion
Our results indicate that in the studied area, nitrogen is a scarce element that presents seasonal fluctuations, which may interfere in the strategies of acquisition and use of this resource by the vegetation.
In the Cerrado, the climate follows a seasonal pattern with distinct periods of dryness and rainfall. During the dry season, from July to October, there is little to no rainfall. Conversely, between October and April, a rainy season occurs with regular precipitation. Based on these climatic patterns, it is reasonable to infer that in the fortnights preceding the collections, there was no rainfall during the dry season, while a significant accumulation of precipitation occurred during the rainy season.
Observing Table 1, we can ascertain the variation in RF and RC values among different species within the campo sujo. These figures reflect species’ responses to specific environmental characteristics of the campo sujo, such as local soil and climatic conditions. Thus, the sensitivity of these metrics highlights their significance in elucidating the distribution and status of species within the studied area.
The soils analyzed around the sampling area and near the collected plants have an acidic character (pHH2O < 4) and can be considered hyper-dystrophic due to low base saturation (V < 30%) [56]. The low C.E.C. associated with the high potential acidity of the soil (H+Al) indicates a low availability of cations such as Ca2+ and K+ for plants. The soil P content can be considered low, as observed in other Cerrado areas [57,58]. The higher concentration of N–NO3− observed during the wet period indicates that precipitation may have favored the nitrification process [59]. The most abundant form of nitrogen in the soil was N–NH4+, which is expected for acidic soils [15], possibly being a nitrogenous form preferentially acquired by vegetation [2,3]. According to studies by [58], the low soil fertility of the studied area can be attributed to the low sum of bases and the acidic nature of the soil observed, which limits the decomposition process and results in a low organic matter content (O.M. < 60 g/dm3), characterizing soil with low fertility.
Although our data reveal novel patterns in nitrogen use by Cerrado grasses, measurements of other attributes would be needed to fully understand this process [23,36]. The species T. leiostachya, A. lanata, A. pressus, and A. siccus showed relatively low NRA (<50 pkat g−1 FW), similar to that observed in some Cerrado woody plants [14,34] but lower than that observed in another study in this same area [60]. On the other hand, the species of the genus Aristida and the C3 grass E. inflexa showed NRA values higher than 50 pkat g−1 FW and exceeding the value of 100 pkat g−1 FW in A. jubata. Although relatively higher, the NRA values observed in these grasses are still much lower than those observed in grasses in the Mediterranean [19] and in tree species in the Atlantic forest, which can exceed 1000 pkat g−1 FW [37]. The relatively low NRA values observed in native grasses and the lower soil N–NO3− values may indicate that plants use this nitrogenous form in a complementary manner to N–NH4+ [7,8,34], which may lead to greater biomass accumulation and a reduction in the effects of Al3+ toxicity on plants [2,17,18].
The seasonal climate of the Cerrado, characterized by distinct periods of drought and rainfall (Figure 1), was found to have a significant impact on the response of native grasses to soil nitrogen availability. The results revealed variations in the responses of grass species to these climatic conditions. The results indicate that climate seasonality can interfere in different ways with NRA in native Cerrado grasses. The highest NRA was expected during the wet season, when N–NO3− is more available in the soil [3]. However, only A. jubata showed a consistently increased NRA response in leaf and root during the wet season, indicating that this species may be more responsive to soil N–NO3− availability [6,39]. On the other hand, the opposite behavior was observed in A. pressus, with a lower NRA in both compartments during the wet period, despite the availability of N–NO3− in the soil. The species A. recurvata, A. lanata, and A. siccus showed variation in the preferential compartment of N–NO3− reduction according to the period analyzed. The NRA in these species was higher in the leaves during the dry period, while in the wet period, the roots were the preferred site for N–NO3− reduction. The higher NRA in the roots during the wet period suggests that grasses absorb and assimilate N–NO3− directly in the underground part, investing in root growth and function to maximize nutrient uptake during this period [18,61,62]. A. jubata showed a preference for reducing N–NO3− in the leaves in both periods, as observed in temperate grasses [36]. In contrast to NRA, the concentration of N–NO3− in leaves was higher compared to roots in both dry and wet periods. In A. recurvata, T. leiostachya, A. pressus, and E. inflexa, the concentration of N–NO3− in the leaves was 6–17 times higher than in the roots, indicating that the aerial part can act as a reservoir of NO3− that can be further assimilated into the leaves or roots [19,21,61].
The principal component analysis evidenced the presence of distinct N-use strategies among the sampled grasses. Different resource use strategies may be associated with the functional and ecological characteristics of the species [2,5,19,28,34,35,60,63], influencing its abundance in natural formations. Low NRA values are usually associated with tree species of late succession, slow growth, and nutrient-deficient environments [7,21]. On the other hand, high NRA activity may be associated with pioneer, colonizing, and fast-growing tree species [21,39]. In the case of grass species, although the NRA values observed in A. jubata were not as high as those observed in pioneer tree species [37], its behavior regarding N–NO3− use and its distribution in campo sujo, occurring abundantly in an area undergoing natural regeneration and road borders, and almost absent in the native field, suggest that this species could be a pioneer grass. The grass E. inflexa also showed high NRA values, but unlike A. jubata, it is the most abundant grass in the native field, with more than double the importance value of other species in the community structure. Grasses with C3 metabolism show higher N use efficiency than C4 grasses in dry environments [64], indicating that even low concentrations of N–NO3− in the soil may be sufficient for E. inflexa to have a competitive advantage over C4 grasses.
Competition and coexistence models indicate that the preference of a species for N–NH4+ or N–NO3− can lead to its exclusion, coexistence, or dominance in a community [31]. This means that, as stated by [31], plants may exhibit a preference for using nitrate, ammonium, or both as nitrogen sources. Within a community, the coexistence of species with different nitrogen utilization preferences can result in various dynamics of exclusion or dominance in the community, depending on the abundance of the species and their respective preferences for nitrogen resource utilization. In other words, species with similar nitrogen utilization preferences may interact to either facilitate or compete with each other, while species with distinct preferences for nitrogen utilization can generate diverse scenarios of exclusion, coexistence, or dominance in the community. These complex interactions among species and their nitrogen utilization preferences contribute to shaping the structure and dynamics of plant communities. In this study, we observed that the species T. leiostachya, A. lanata, A. pressus, and A. siccus have the lowest NRA values, possibly indicating a preference to use the available N–NH4+ in the soil [2,6,65]. Furthermore, these species present intermediate and very close values of importance in the community, indicating a balanced coexistence among them. On the other hand, the species A. jubata, A. recurvata, and E. inflexa, which presented the highest values of NRA, showed two distinct behaviors in the community: (i) the species of the genus Aristida are very little abundant in the native field, with A. jubata almost absent and A. recurvata occurring sparsely, with individuals significantly smaller than other species studied, and (ii) E. inflexa is the most abundant species in the native field, being the most frequent grass with greater coverage, besides presenting C3 photosynthesis, unlike the other species evaluated. The different N-use strategies of these grasses seem to influence their abundance in the native field, which may be contributing to the maintenance of grass diversity in this area.
5. Conclusions
Our results demonstrate that native grass species in the Cerrado exhibit seasonal modulation in nitrate assimilation, with clear differentiation between aboveground and underground plant parts. During the dry period, we observed higher N–NO3− reduction activity in the leaves, while in the wet period, this activity was more pronounced in the roots. This seasonal alternation suggests an adaptation of grasses to climatic variations, optimizing the utilization of available nitrogen. Furthermore, the relationship between nitrogen availability and species importance in the community reveals the direct influence of seasonality and nitrogen assimilation capacity on the distribution and abundance of grasses in the Cerrado campo sujo. These findings contribute to a better understanding of the ecology and physiology of native Cerrado grasses, highlighting the importance of climatic seasonality in nitrogen dynamics in this unique ecosystem.
This study presents some important limitations that need to be considered. Firstly, there is the limitation of sampling, which was restricted to a limited number of native grass species from the Cerrado campo sujo, despite the high richness of the ecosystem. Thus, future studies with other native grass species and even non-native ones may be necessary, especially considering the potential threat to local diversity. Additionally, another potential limitation is the temporal scale of the study, which may have been limited, hindering the observation of long-term trends or interannual variations in nitrogen dynamics. Therefore, it is suggested that further studies be conducted to address these limitations and gain a more comprehensive understanding of the processes under investigation.
Author Contributions
Conceptualization, V.C.K., E.F.L.P.-S. and E.H.; methodology, V.C.K., E.F.L.P.-S. and E.H.; software, V.C.K.; validation, V.C.K., E.F.L.P.-S., S.T.M. and E.H.; formal analysis, V.C.K., E.F.L.P.-S., S.T.M. and E.H.; investigation, V.C.K., E.F.L.P.-S., S.T.M. and E.H.; resources, E.F.L.P.-S.; data curation, V.C.K.; writing—original draft preparation, V.C.K., E.F.L.P.-S. and E.H.; writing—review and editing, V.C.K., E.F.L.P.-S. and E.H.; visualization, V.C.K.; supervision, E.F.L.P.-S. and E.H.; project administration, E.F.L.P.-S.; funding acquisition, E.H. All authors have read and agreed to the published version of the manuscript.
Funding
Support from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brazil (CAPES), research scholarship—Funding Code 001.
Data Availability Statement
Data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
This study received logistical support for field research from the Foundation for Forest Conservation and Production of the State of São Paulo (Fundação Florestal/SMA-SP) and research support from the laboratory facilities of the Universidade Federal do ABC, campus São Bernardo do Campo, SP.
Conflicts of Interest
The authors declare no conflict of interest.
References
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Falkengren-Grerup, U. Interspecies differences in the preference of ammonium and nitrate in vascular plants. Oecologia 1995, 102, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Houlton, B.Z.; Sigman, D.M.; Schuur, A.G.; Hedin, L.O. A climate-driven switch in plant nitrogen acquisition within tropical forest communities. Proc. Natl. Acad. Sci. USA 2007, 104, 8902–8906. [Google Scholar] [CrossRef] [PubMed]
- Harrison, K.A.; Bol, R.; Bardgett, R.D. Preferences for uptake of different nitrogen forms by co-existing plant species and soil microbes. Ecology 2007, 88, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.C.; da Silva, L.M.I.; de Freitas, L.D.; Debiasi, T.V.; Marchiori, N.M.; Aidar, M.P.M.; Bianchini, E.; Pimenta, J.A.; Stolf-Moreira, R. Nitrogen use strategies of seedlings from neotropical tree species of distinct successional groups. Plant Physiol. Biochem. 2017, 114, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, T.; Liu, X.; Nie, M.; Xu, X.; Zhou, S. Limited inorganic N niche partitioning by nine alpine plant species after long-term nitrogen addition. Sci. Total. Environ. 2020, 718, 137270. [Google Scholar] [CrossRef] [PubMed]
- Uscola, M.; Oliet, J.A.; Villar-Salvador, P.; Díaz-Pinés, E.; Jacobs, D.F. Nitrogen form and concentration interact to affect the performance of two ecologically distinct mediterranean forest trees. Eur. J. For. Res. 2014, 133, 235–246. [Google Scholar] [CrossRef]
- Andrews, M.; Maule, H.G.; Hodge, S.; Cherrill, A.; Raven, J.A. Seed dormancy, nitrogen nutrition and shade acclimation of impatiens glandulifera: Implications for successful invasion of deciduous woodland. Plant Ecol. Divers. 2009, 2, 145–153. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef]
- Eiten, G. The Cerrado vegetation of Brazil. Bot. Rev. 1972, 38, 201–341. [Google Scholar] [CrossRef]
- Furley, P.A.; Ratter, J.A. Soil Resources and Plant Communities of the Central Brazilian Cerrado and Their Development. J. Biogeogr. 1988, 15, 97. [Google Scholar] [CrossRef]
- Pereira-Silva, E.F.L.; Hardt, E.; dos Santos, J.E.; Aidar, M.P.M. Atividade de Nitrato Redutase e Conteúdo de Nitrogênio em Folhas de Espécies Arbóreas de Cerradão da Estação Ecológica de Jataí, Luiz Antônio, SP; EdUFSCar: São Carlos, Brazil, 2006; pp. 65–79. [Google Scholar]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Skrumsager Møller, I.; White, P. Functions of Macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; pp. 135–189. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A Re-evaluatin of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Shen, R.F.; Bin Sun, Q. Ammonium under solution culture alleviates aluminum toxicity in rice and reduces aluminum accumulation in roots compared with nitrate. Plant Soil 2008, 315, 107–121. [Google Scholar] [CrossRef]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Pereira-Silva, E.; Casals, P.; Sodek, L.; Delitti, W.; Vallejo, V. Post-fire nitrogen uptake and allocation by two resprouting herbaceous species with contrasting belowground traits. Environ. Exp. Bot. 2019, 159, 157–167. [Google Scholar] [CrossRef]
- Cramer, M.D.; Hawkins, H.-J.; Verboom, G.A. The importance of nutritional regulation of plant water flux. Oecologia 2009, 161, 15–24. [Google Scholar] [CrossRef]
- Smirnoff, N.; Stewart, G.R. Nitrate assimilation and translocation by higher plants: Comparative physiology and ecological consequences. Physiol. Plant. 1985, 64, 133–140. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M.; Palmborg, C.; Prinz, A.; Schulze, E.-D. The Role of Plant Diversity and Composition for Nitrate Leaching in Grasslands. Ecology 2003, 84, 1539–1552. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. Ecological significance and complexity of N-source preference in plants. Ann. Bot. 2013, 112, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Salsac, L.; Chaillou, S.; Morot-Gaudry, J.-F.; Lesaint, C.; Jolivet, E. Nitrate and ammonium nutrition in plants. Plant Physiol. Biochem. 1987, 25, 805–812. [Google Scholar]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2017, 217, 35–53. [Google Scholar] [CrossRef]
- Amancio, S.; Stulen, I. Nitrogen Acquisition and Assimilation in Higher Plants, 3rd ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Ghannoum, O. C4 photosynthesis and water stress. Ann. Bot. 2009, 103, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Lu, P.; Zhai, X.; Zhang, R.; Zheng, Y.; Wang, H.; Nie, B.; Bai, W.; Niu, S.; Shi, P.; et al. An integrated belowground trait-based understanding of nitrogen-driven plant diversity loss. Glob. Chang. Biol. 2022, 28, 3651–3664. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.M.; Cleland, E.E.; Collins, S.L.; Fargione, J.E.; Gough, L.; Gross, K.L.; Pennings, S.C.; Suding, K.N.; Grace, J.B. Environmental and plant community determinants of species loss following nitrogen enrichment. Ecol. Lett. 2007, 10, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Eller, C.B.; Oliveira, R.S. Effects of nitrogen availability on the competitive interactions between an invasive and a native grass from Brazilian cerrado. Plant Soil 2017, 410, 63–72. [Google Scholar] [CrossRef]
- Boudsocq, S.; Niboyet, A.; Lata, J.C.; Raynaud, X.; Loeuille, N.; Mathieu, J.; Blouin, M.; Abbadie, L.; Barot, S. Plant Preference for Ammonium versus Nitrate: A Neglected Determinant of Ecosystem Functioning? Am. Nat. 2012, 180, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, P.C.; Menendez, E.; da Silva, D.L.; Bonieck, D.; Ramírez-Bahena, M.H.; Resende-Stoianoff, M.A.; Peix, A.; Velázquez, E.; Mateos, P.F.; Scotti, M.R. Invasion of the Brazilian campo rupestre by the exotic grass Melinis minutiflora is driven by the high soil N availability and changes in the N cycle. Sci. Total. Environ. 2017, 577, 202–211. [Google Scholar] [CrossRef]
- Garcia, D.B.; Xavier, R.O.; Camargo, P.B.; Vieira, S.A.; Pivello, V.R. Can an invasive African grass affect carbon and nitrogen stocks in open habitats of the Brazilian Cerrado? Flora Morphol. Distrib. Funct. Ecol. Plants 2022, 286, 151968. [Google Scholar] [CrossRef]
- Stewart, G.R.; Joly, C.A.; Smirnoff, N. Partitioning of inorganic nitrogen assimilation between the roots and shoots of cerrado and forest trees of contrasting plant communities of South East Brasil. Oecologia 1992, 91, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Rice, E.L. Differences in nitrate reductase activity between species of different stages in old field succession. Oecologia 1983, 57, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Scheurwater, I.; Koren, M.; Lambers, H.; Atkin, O.K. The contribution of roots and shoots to whole plant nitrate reduction in fast- and slow-growing grass species. J. Exp. Bot. 2002, 53, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Aidar, M.P.M.; Schmidt, S.; Moss, G.; Stewart, G.R.; Joly, C.A. Nitrogen use strategies of neotropical rainforest trees in threatened Atlantic Forest. Plant Cell Environ. 2003, 26, 389–399. [Google Scholar] [CrossRef]
- Pereira-Silva, E.F.L.; Hardt, E.; Joly, C.A.; Aidar, M.P.M. Sucessão Ecológica E O Uso De Nitrogênio Em Florestas Tropicais. Interciência Soc. 2011, 1, 149–159. [Google Scholar]
- Pereira-Silva, E.F.L.; Hardt, E.; Fernandes, A.O. The soil-plant relationship of nitrogen use in three tropical tree species. Web Ecol. 2012, 12, 57–64. [Google Scholar] [CrossRef]
- Baitello, J.B.; de Aguiar, O.T.; Pastore, J.A.; Arzolla, F.A.R.D.P. Parque Estadual do Juquery: Refúgio do Cerrado no Domínio Atlântico. IF Ser. Regist. 2013, 50, 1–46. [Google Scholar]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Moraes, G.J.L.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Keller, V.C.; Pereira-Silva, E.F.L.; Hardt, E. High richness, new occurrences, and threatened species in a savanna grassland remnant in the largest Brazilian metropolis. Check List 2021, 17, 507–549. [Google Scholar] [CrossRef]
- Amorim, D.G.d.A.; Zaine, J.E.; Rodrigues, F.H. Avaliação de suscetibilidade à erosão e movimentação gravitacional de massa no Parque Estadual do Juquery, Franco da Rocha (SP). Rev. Inst. Geocièncias—USP 2017, 17, 3–21. [Google Scholar] [CrossRef]
- Rossi, M. Mapa Pedológico do Estado de São Paulo: Revisado e Expandido; Instituto Florestal: São Paulo, Brazil, 2017. [Google Scholar]
- Keller, V.C. Diversidade Taxonômica e Funcional em uma Comunidade Vegetal Campestre de Cerrado na Região Metropolitana de São Paulo. Master’s Thesis, Universidade Federal de São Paulo, Diadema, Brasil. Available online: https://repositorio.unifesp.br/items/b08eb344-2633-435c-bbac-7793a6b37798 (accessed on 18 April 2024).
- McCullough, H. The determination of ammonia in whole blood by a direct colorimetric method. Clin. Chim. Acta 1967, 17, 297–304. [Google Scholar] [CrossRef]
- Stewart, G.; Lee, J.; Orebamjo, T. Nitrogen metabolism of halophytes II. Nitrate availability and utilization. New Phytol. 1973, 72, 539–546. [Google Scholar] [CrossRef]
- Stewart, G.R.; Popp, M.; Holzapfel, I.; Stewart, J.A.; Dickie-Eskew, A. Localization of nitrate reduction in ferns and its relationship to environment and physiological characteristics. New Phytol. 1986, 104, 373–384. [Google Scholar] [CrossRef]
- Saltzman, B.E. Colorimetric microdetermination of nitrogen dioxide in atmosphere. Anal. Chem. 1954, 26, 1949–1955. [Google Scholar] [CrossRef]
- Bieleski, R.L.; Turner, N.A. Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Anal. Biochem. 1966, 17, 278–293. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry; Macmillan: New York, NY, USA, 1995. [Google Scholar]
- R-Core-Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2018. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- dos Santos, H.G.; Jacomine, P.K.T.; Dos Anjos, L.H.C.; De Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; de Almeida, J.A.; de Araujo Filho, J.C.; de Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos; Embrapa: Brasília, Brazil, 2018. [Google Scholar]
- Lepsch, I.F. Formação e Conservação dos Solos; Oficina de textos: São Paulo, Brazil, 2016. [Google Scholar]
- Soares, M.P.; Reys, P.; Pifano, D.S.; de Sá, J.L.; da Silva, P.O.; Santos, T.M.; Silva, F.G. Relationship Between Edaphic Factors and Vegetation in Savannas of the Brazilian Midwest Region. Rev. Bras. Ciência Solo 2015, 39, 821–829. [Google Scholar] [CrossRef]
- Pivello, V.R.; Oliveras, I.; Miranda, H.S.; Haridasan, M.; Sato, M.N.; Meirelles, S.T. Effect of fires on soil nutrient availability in an open savanna in Central Brazil. Plant Soil 2010, 337, 111–123. [Google Scholar] [CrossRef]
- Corre, M.D.; Schnabel, R.R.; Stout, W.L. Spatial and seasonal variation of gross nitrogen transformations and microbial biomass in a Northeastern US grassland. Soil Biol. Biochem. 2002, 34, 445–457. [Google Scholar] [CrossRef]
- Pereira-Silva, E.F.L.; Hardt, E.; Biral, M.B.; Keller, V.C.; Delitti, W.B.C. Effects of recent fire on soil conditions and nutrient use of a native and an invasive grass in the Brazilian savanna. Écoscience 2019, 26, 359–370. [Google Scholar] [CrossRef]
- Sarmiento, G. Adaptive strategies of perennial grasses in South American savannas. J. Veg. Sci. 1992, 3, 325–336. [Google Scholar] [CrossRef]
- Patterson, K.; Walters, L.A.; Cooper, A.M.; Olvera, J.G.; Rosas, M.A.; Rasmusson, A.G.; Escobar, M.A. Nitrate-regulated glutaredoxins control arabidopsis primary root growth. Plant Physiol. 2016, 170, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Xi, N.; Zhu, B.-R.; Zhang, D.-Y. Contrasting grass nitrogen strategies reflect interspecific trade-offs between nitrogen acquisition and use in a semi-arid temperate grassland. Plant Soil 2017, 418, 267–276. [Google Scholar] [CrossRef]
- Luo, W.; Wang, X.; Sardans, J.; Wang, Z.; Dijkstra, F.A.; Lü, X.-T.; Peñuelas, J.; Han, X. Higher capability of C3 than C4 plants to use nitrogen inferred from nitrogen stable isotopes along an aridity gradient. Plant Soil 2018, 428, 93–103. [Google Scholar] [CrossRef]
- Weigelt, A.; Bol, R.; Bardgett, R.D. Preferential uptake of soil nitrogen forms by grassland plant species. Oecologia 2005, 142, 627–635. [Google Scholar] [CrossRef]
Figure 1.
Climatic diagram of Juquery State Park between 7 August 2017 and 26 April 2018. Roman numerals (I, II, and IV) indicate the week of the corresponding month’s values. Black arrow = collection during the dry period. Gray arrow = collection during the wet period.
Figure 1.
Climatic diagram of Juquery State Park between 7 August 2017 and 26 April 2018. Roman numerals (I, II, and IV) indicate the week of the corresponding month’s values. Black arrow = collection during the dry period. Gray arrow = collection during the wet period.

Figure 2.
Principal component analysis (PCA) diagram: (A) Soil rhizosphere sample variables of the studied species—nitrate (N–NO3−), ammonium (N–NH4+), pHH2O, conductivity (cond.), calcium (Ca2+), and potassium (K+); (B) nitrogen use variables—Nitrate Reductase Activity (NRA) and nitrate content (NIT) in leaf (f) and root (r) of species during dry (s) and wet (u) periods. Aj = Aristida jubata; Ar = A. recurvata; Tl = Tristachya leiostachya; Al = Anthaenantia lanata; Ap = Axonopus pressus; As = A. siccus; Ei = Echinolaena inflexa.
Figure 2.
Principal component analysis (PCA) diagram: (A) Soil rhizosphere sample variables of the studied species—nitrate (N–NO3−), ammonium (N–NH4+), pHH2O, conductivity (cond.), calcium (Ca2+), and potassium (K+); (B) nitrogen use variables—Nitrate Reductase Activity (NRA) and nitrate content (NIT) in leaf (f) and root (r) of species during dry (s) and wet (u) periods. Aj = Aristida jubata; Ar = A. recurvata; Tl = Tristachya leiostachya; Al = Anthaenantia lanata; Ap = Axonopus pressus; As = A. siccus; Ei = Echinolaena inflexa.

Figure 3.
Box plot showing the values of Nitrate Reductase Activity (NRA) and nitrate (N–NO3−) in leaf (L) and root (R) of the species during dry and wet periods. Letters indicate intra-specific differences between compartments and periods (Mann–Whitney U test, p < 0.05).
Figure 3.
Box plot showing the values of Nitrate Reductase Activity (NRA) and nitrate (N–NO3−) in leaf (L) and root (R) of the species during dry and wet periods. Letters indicate intra-specific differences between compartments and periods (Mann–Whitney U test, p < 0.05).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Selected Poaceae species from PEJY were used for nitrogen use characterization.
| Tribe | Species | Acronym | PP | RF | RC | IV |
|---|---|---|---|---|---|---|
| Aristideae | Aristida jubata (Arechav.) Herter | Aj | C4 | 0.24 | 0.18 | 0.42 |
| Aristida recurvata Kunth | Ar | C4 | 2.89 | 0.67 | 3.56 | |
| Arundinelleae | Tristachya leiostachya Nees | Tl | C4 | 4.05 | 8.21 | 12.26 |
| Paniceae | Anthaenantia lanata (Kunth) Benth. | Al | C4 | 4.62 | 7.56 | 12.18 |
| Axonopus pressus (Nees ex Steud.) Parodi | Ap | C4 | 2.89 | 4.30 | 7.19 | |
| Axonopus siccus (Nees) Kuhlm. | As | C4 | 3.47 | 8.06 | 11.53 | |
| Echinolaena inflexa (Poir.) Chase | Ei | C3 | 4.62 | 25.09 | 29.71 |
PP: photosynthetic path; RF: relative frequency; RC: relative cover; IV: importance value.
Table 2.
Soil chemical parameters (mean ± standard error): (A) in dry and wet periods; (B) in rhizosphere samples of the studied species. Letters indicate significant differences between periods and among species (ANOVA, p < 0.05).
Table 2.
Soil chemical parameters (mean ± standard error): (A) in dry and wet periods; (B) in rhizosphere samples of the studied species. Letters indicate significant differences between periods and among species (ANOVA, p < 0.05).
| A | ||||||||
|---|---|---|---|---|---|---|---|---|
| Season | n | pH H2O | pHss | Ca2+ | K+ | N–NO3− | N–NH4+ | Condutiv |
| g/L | mg/g | µS/cm2 | ||||||
| Dry | 7 | 3.8 ± 0.1 a | 4.2 ± 0.1 a | 2.8 ± 0.2 a | 1.8 ± 0.3 a | 7.3 ± 0.5 a | 18.6 ± 3.3 a | 15.9 ± 1.8 a |
| Wet | 35 | 3.9 ± 0.1 a | 3.7 ± 0.1 a | 5.4 ± 0.6 b | 1.7 ± 0.2 a | 15.1 ± 0.8 b | 30.6 ± 3.1 A | 18.7 ± 1.9 a |
| B | ||||||||
| Species | ||||||||
| Aj | 5 | 3.8 ± 0.1 a | 3.5 ± 0.1 a | 6.6 ± 0.2 a | 1.0 ± 0.0 b | 16.4 ± 4.2 a | 39.2 ± 5.2 a | 20.4 ± 3.7 ab |
| Ar | 5 | 3.8 ± 0.1 a | 3.4 ± 0.1 a | 7.0 ± 3.2 ab | 1.8 ± 0.6 ab | 14.4 ± 3.1 a | 19.6 ± 6.0 A | 20.0 ± 5.6 ab |
| Tl | 5 | 3.6 ± 0.1 a | 3.6 ± 0.1 a | 6.6 ± 0.9 a | 2.8 ± 0.2 a | 12.8 ± 2.9 a | 38.9 ± 10.7 A | 25.3 ± 5.7 ab |
| Al | 5 | 4.0 ± 0.1 a | 3.7 ± 0.1 a | 4.4 ± 1.0 ab | 2.0 ± 0.8 ab | 14.8 ± 4.4 a | 41.1 ± 6.8 A | 24.0 ± 3.3 a |
| Ap | 5 | 3.8 ± 0.1 a | 3.7 ± 0.1 a | 3.8 ± 0.9 b | 1.6 ± 0.2 b | 15.6 ± 5.0 a | 33.1 ± 6.4 A | 15.2 ± 1.4 b |
| As | 5 | 3.9 ± 0.2 a | 3.5 ± 0.1 a | 5.2 ± 1.0 ab | 1.6 ± 0.2 b | 16.4 ± 2.8 a | 29.8 ± 8.4 A | 16.8 ± 1.2 ab |
| Ei | 5 | 3.9 ± 0.1 a | 3.6 ± 0.1 a | 6.8 ± 1.0 a | 1.6 ± 0.2 b | 11.6 ± 2.1 a | 20.7 ± 8.1 A | 20.0 ± 3.5 ab |
Aj = Aristida jubata, Ar = A. recurvata, Tl = Tristachya leiostachya, Al = Anthaenantia lanata, Ap = Axonopus pressus, As = A. siccus, and Ei = Echinolaena inflexa.
Table 3.
Comparison between dry and wet periods, leaf and root compartments, and their interaction for nitrate reductase (NRA) and nitrate (N–NO3−) variables (Scheirer–Ray–Hare Test, * p < 0.05, ** p < 0.01, and *** p < 0.001, ns = not significant): for all species; and for each grass species.
Table 3.
Comparison between dry and wet periods, leaf and root compartments, and their interaction for nitrate reductase (NRA) and nitrate (N–NO3−) variables (Scheirer–Ray–Hare Test, * p < 0.05, ** p < 0.01, and *** p < 0.001, ns = not significant): for all species; and for each grass species.
| All Species | Aj | Ar | Tl | Al | Ap | As | Ei | |
|---|---|---|---|---|---|---|---|---|
| NRA | ||||||||
| Period | ns | *** | ns | * | ns | *** | ns | ns |
| Compartment | ns | * | ns | ns | ns | ns | * | ** |
| Interaction | *** | ns | ** | * | *** | ns | * | ns |
| N–NO3− | ||||||||
| Period | ns | ns | ns | ns | ns | ns | ** | ns |
| Compartment | *** | *** | *** | *** | * | *** | * | *** |
| Interaction | ns | ns | ns | ns | ns | ns | ns | ns |
Aj = Aristida jubata, Ar = A. recurvata, Tl = Tristachya leiostachya, Al = Anthaenantia lanata, Ap = Axonopus pressus, As = A. siccus, and Ei = Echinolaena inflexa.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Keller, V.C.; Pereira-Silva, E.F.L.; Meirelles, S.T.; Hardt, E. Effects of Seasonal Variation on Nitrogen Use in Brazilian Cerrado Grass Communities. Nitrogen 2024, 5, 373-385. https://doi.org/10.3390/nitrogen5020024
AMA Style
Keller VC, Pereira-Silva EFL, Meirelles ST, Hardt E. Effects of Seasonal Variation on Nitrogen Use in Brazilian Cerrado Grass Communities. Nitrogen. 2024; 5(2):373-385. https://doi.org/10.3390/nitrogen5020024
Chicago/Turabian StyleKeller, Victor Camargo, Erico Fernando Lopes Pereira-Silva, Sergio Tadeu Meirelles, and Elisa Hardt. 2024. "Effects of Seasonal Variation on Nitrogen Use in Brazilian Cerrado Grass Communities" Nitrogen 5, no. 2: 373-385. https://doi.org/10.3390/nitrogen5020024