

Using Unoccupied Aerial Systems (UASs) to Determine the Distribution Patterns of Tamanend’s Bottlenose Dolphins (Tursiops erebennus) across Varying Salinities in Charleston, South Carolina

Abstract
:1. Introduction
2. Materials and Methods
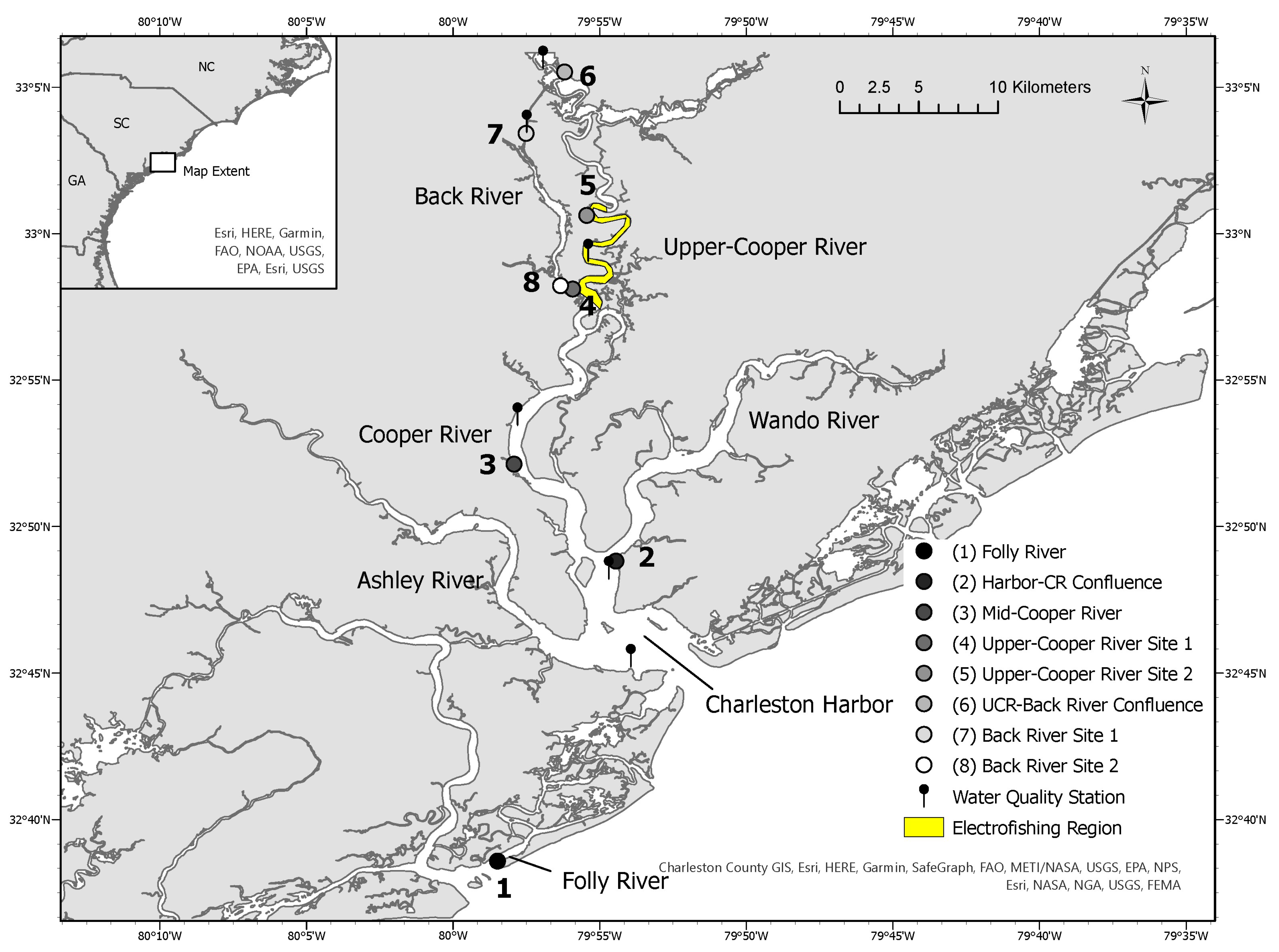
2.1. Study Area
- Folly River: Interior-side barrier island river that is adjacent to the Atlantic Ocean.
- Harbor–Cooper River Confluence: Deep, open area estuary confluence where the Charleston Harbor branches into the Cooper and Wando Rivers.
- Mid-Cooper River: Brackish riverine habitat.
- 4.
- Upper-Cooper River (UCR) Site 1: Brackish/freshwater riverine habitat.
- 5.
- Upper-Cooper River (UCR) Site 2: Brackish/freshwater riverine habitat.
- 6.
- Upper-Cooper River (UCR)/Back River Confluence: Open freshwater confluence where the Back River branches off the Cooper River.
- 7.
- Back River Site 1: Freshwater riverine habitat.
- 8.
- Back River Site 2: Freshwater riverine habitat; terminal end of the Back River.
2.2. UAS Survey Methodology
2.3. UAS Video Analysis
2.4. Environmental Data Collection
2.5. Statistical Analysis
2.5.1. Distribution across Study Sites
2.5.2. Seasonal Analysis
2.5.3. Environmental Variables
2.5.4. Prey Data Collection and Analysis
3. Results
3.1. UAS Survey Effort
3.2. Dolphin Distribution across Study Sites
3.3. Dolphin Distribution across Seasons
3.4. Dolphin Distribution in Relation to Environmental Variables
3.5. Prey Analysis
4. Discussion
4.1. Bottlenose Dolphin Distribution in the Charleston Estuary System (CES)
4.1.1. Distribution across High-Salinity Sites
4.1.2. Distribution across Low-Salinity Sites
4.2. Benefits and Limitations of UAS Methods to Study Bottlenose Dolphins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costa, A.P.B.; McFee, W.; Wilcox, L.A.; Archer, F.I.; Rosel, P.E. The common bottlenose dolphin (Tursiops truncatus) ecotypes of the western North Atlantic revisited: An integrative taxonomic investigation supports the presence of distinct species. Zool. J. Linn. Soc. 2022, 196, 1608–1636. [Google Scholar] [CrossRef]
- Wells, R.S.; Rhinehart, H.L.; Cunningham, P.; Whaley, J.; Baran, M.; Koberna, C.; Costa, D.P. Long distance offshore movements of bottlenose dolphins. Mar. Mammal Sci. 1999, 15, 1098–1114. [Google Scholar] [CrossRef]
- Wells, R.S.; Scott, M.D. Bottlenose dolphins (Tursiops truncatus). In Encyclopedia of Marine Mammals, 3rd ed.; Wursig, B., Thewissen, J.G.M., Kovacs, K., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 118–125. [Google Scholar]
- Hayes, S.A.; Josephson, E.; Maze-Foley, K.; Rosel, P.E.; Wallace, J.U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments 2021, NOAA Technical Memorandum NMFS-NE-271; U.S. Department of Commerce: Washington, DC, USA, 2022.
- Hornsby, F.; McDonald, T.; Balmer, B.; Speakman, T.; Mullin, K.; Rosel, P.; Wells, R.; Telander, A.; Marcy, P.; Schwacke, L. Using salinity to identify common bottlenose dolphin habitat in Barataria Bay, Louisiana, USA. Endanger. Species Res. 2017, 33, 181–192. [Google Scholar] [CrossRef]
- Takeshita, R.; Balmer, B.C.; Messina, F.; Zolman, E.S.; Thomas, L.; Wells, R.S.; Smith, C.R.; Rowles, T.K.; Schwacke, L.H. High site-fidelity in common bottlenose dolphins despite low salinity exposure and associated indicators of compromised health. PLoS ONE 2021, 16, e0258031. [Google Scholar] [CrossRef] [PubMed]
- Barco, S.G.; Swingle, W.M.; McLellan, W.A.; Harris, R.N.; Pabst, D.A. Local abundance and distribution of bottlenose dolphins (Tursiops truncatus) in the nearshore waters of Virginia Beach, Virginia. Mar. Mammal Sci. 1999, 15, 394–408. [Google Scholar] [CrossRef]
- Yeates, L.C.; Houser, D.S. Thermal tolerance in bottlenose dolphins (Tursiops truncatus). J. Exp. Biol. 2011, 211, 3249–3257. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.; Turrell, W.; Lutkebohle, T.; Thompson, P. Influence of the tidal cycle and a tidal intrusion front on the spatio-temporal distribution of coastal bottlenose dolphins. Mar. Ecol. Prog. Ser. 2002, 239, 221–229. [Google Scholar] [CrossRef]
- Fury, C.A.; Harrison, P.L. Impact of flood events on dolphin occupancy patterns. Mar. Mammal Sci. 2011, 27, E185–E205. [Google Scholar] [CrossRef]
- Torres, L.G.; Read, A.J.; Halpin, P. Fine-scale habitat modeling of a top marine predator: Do prey data improve predictive capacity? Ecol. Appl. 2008, 18, 1702–1717. [Google Scholar] [CrossRef]
- Eierman, L.; Connor, R. Foraging behavior, prey distribution, and microhabitat use by bottlenose dolphins, Tursiops truncatus, in a tropical atoll. Mar. Ecol. Prog. Ser. 2014, 503, 279–288. [Google Scholar] [CrossRef]
- Carmichael, R.H.; Graham, W.M.; Aven, A.; Worthy, G.; Howden, S. Were multiple stressors a “perfect storm” for Northern Gulf of Mexico bottlenose dolphins (Tursiops truncatus) in 2011? PLoS ONE 2012, 7, e41155. [Google Scholar] [CrossRef] [PubMed]
- McClain, A.M.; Daniels, R.; Gomez, F.M.; Ridgway, S.H.; Takeshita, R.; Jensen, E.D.; Smith, C.R. Physiological effects of low salinity exposure on bottlenose dolphins (Tursiops truncatus). J. Zool. Bot. Gard. 2020, 1, 61–75. [Google Scholar] [CrossRef]
- Marley, S.A.; Salgado Kent, C.P.; Erbe, C. Occupancy of Bottlenose Dolphins (Tursiops aduncus) in Relation to Vessel Traffic, Dredging, and Environmental Variables within a Highly Urbanised Estuary. Hydrobiologia 2017, 792, 243–263. [Google Scholar] [CrossRef]
- Pirotta, E.; Merchant, N.D.; Thompson, P.M.; Barton, T.R.; Lusseau, D. Quantifying the effect of boat disturbance on bottlenose dolphin foraging activity. Biol. Conserv. 2015, 181, 82–89. [Google Scholar] [CrossRef]
- Noke, W.D.; Odell, D.K. Interactions between the Indian River Lagoon blue crab fishery and the bottlenose dolphin, Tursiops truncatus. Mar. Mammal Sci. 2002, 18, 819–832. [Google Scholar] [CrossRef]
- Kucklick, J.; Schwacke, L.; Wells, R.; Hohn, A.; Guichard, A.; Yordy, J.; Hansen, L.; Zolman, E.; Wilson, R.; Litz, J.; et al. Bottlenose Dolphins as Indicators of Persistent Organic Pollutants in the Western North Atlantic Ocean and Northern Gulf of Mexico. Environ. Sci. Technol. 2011, 45, 4270–4277. [Google Scholar] [CrossRef]
- Rowles, T.K.; Schwacke, L.S.; Wells, R.S.; Saliki, J.T.; Hansen, L.; Hohn, A.; Townsend, F.; Sayre, R.A.; Hall, A.J. Evidence of Susceptibility to Morbillivirus Infection in Cetaceans from the United States. Mar. Mammal Sci. 2011, 27, 1–19. [Google Scholar] [CrossRef]
- Zolman, E.S. Residence Patterns of Bottlenose Dolphins (Tursiops truncatus) in the Stono River Estuary, Charleston County, South Carolina, USA. Mar. Mammal Sci. 2002, 18, 879–892. [Google Scholar] [CrossRef]
- Speakman, T.R.; Lane, S.M.; Schwacke, L.H.; Fair, P.A.; Zolman, E.S. Mark-Recapture Estimates of Seasonal Abundance and Survivorship for Bottlenose Dolphins (Tursiops truncatus) near Charleston, South Carolina, USA. J. Cetacean Res. Manag. 2010, 11, 153–162. [Google Scholar] [CrossRef]
- Bouchillon, H.; Levine, N.S.; Fair, P.A. GIS Investigation of the Relationship of Sex and Season on the Population Distribution of Common Bottlenose Dolphins (Tursiops truncatus) in Charleston, South Carolina. Int. J. Geogr. Inf. Syst. 2020, 34, 1552–1566. [Google Scholar] [CrossRef]
- Althausen, J.D.; Kjerfve, B. Distribution of Suspended Sediment in a Partially Mixed Estuary, Charleston Harbor, South Carolina, USA. Estuar. Coast. Shelf Sci. 1992, 35, 517–531. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration (NOAA). Final Damage Assessment and Restoration Plan and Environmental Assessment for the Koppers Site, Charleston, South Carolina, May 2017. Available online: https://pub-data.diver.orr.noaa.gov/admin-record/6217/Koppers_DARP_EA_FINAL_7-10-17.pdf (accessed on 20 November 2022).
- Schwacke, L.H.; Voit, E.O.; Hansen, L.J.; Wells, R.S.; Mitchum, G.B.; Hohn, A.A.; Fair, P.A. Probabilistic Risk Assessment of Reproductive Effects of Polychlorinated Biphenyls on Bottlenose Dolphins (Tursiops truncatus) from the Southeast United States Coast. Environ. Toxicol. Chem. 2002, 21, 2752–2764. [Google Scholar] [CrossRef]
- Fair, P.A.; White, N.D.; Wolf, B.; Arnott, S.A.; Kannan, K.; Karthikraj, R.; Vena, J.E. Persistent Organic Pollutants in Fish from Charleston Harbor and Tributaries, South Carolina, United States: A Risk Assessment. Environ. Res. 2018, 167, 598–613. [Google Scholar] [CrossRef] [PubMed]
- Fair, P.A.; Mitchum, G.; Hulsey, T.C.; Adams, J.; Zolman, E.; McFee, W.; Wirth, E.; Bossart, G.D. Polybrominated Diphenyl Ethers (PBDEs) in Blubber of Free-Ranging Bottlenose Dolphins (Tursiops truncatus) from Two Southeast Atlantic Estuarine Areas. Arch. Environ. Contam. Toxicol. 2007, 53, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Kjerfve, B. The Santee-Cooper: A Study of Estuarine Manipulations. In Estuarine Processes; Academic Press: Cambridge, MA, USA, 1976; pp. 44–56. [Google Scholar] [CrossRef]
- Bradley, P.M.; Kjerfve, B.; Morris, J.T. Rediversion Salinity Change in the Cooper River, South Carolina: Ecological Implications. Estuaries 1990, 13, 373–379. [Google Scholar] [CrossRef]
- Cheever, A. Examining the Environmental Variables Preceding Stranding by Common Bottlenose Dolphins in the Cooper and Back Rivers, Charleston, South Carolina. Master’s Thesis, The Graduate School of the University of Charleston, Charleston, SC, USA, 2021. [Google Scholar]
- Rosel, P.E.; Watts, H. Hurricane Impacts on Bottlenose Dolphins in the Northern Gulf of Mexico. Gulf Mex. Sci. 2008, 25, 7. [Google Scholar] [CrossRef]
- Deming, A.C.; Wingers, N.L.; Moore, D.P.; Rotstein, D.; Wells, R.S.; Ewing, R.; Hodanbosi, M.R.; Carmichael, R.H. Health Impacts and Recovery from Prolonged Freshwater Exposure in a Common Bottlenose Dolphin (Tursiops truncatus). Front. Vet. Sci. 2020, 7, 235. [Google Scholar] [CrossRef] [PubMed]
- Ewing, R.Y.; Mase-Guthrie, B.; McFee, W.; Townsend, F.; Manire, C.A.; Walsh, M.; Borkowski, R.; Bossart, G.D.; Schaefer, A.M. Evaluation of Serum for Pathophysiological Effects of Prolonged Low Salinity Water Exposure in Displaced Bottlenose Dolphins (Tursiops truncatus). Front. Vet. Sci. 2017, 4, 80. [Google Scholar] [CrossRef]
- Mullin, K.D.; Barry, K.P.; Sinclair, C.; Litz, J.; Maze-Foley, K.; Fougeres, E.; Mase, B.; Ewing, R.Y.; Gorgone, A.M.; Adams, J. Common Bottlenose Dolphins (Tursiops truncatus) in Lake Pontchartrain, Louisiana: 2007 to Mid–2014. In NOAA Technical Memorandum NMFS-SEFSC-673; 2015. Available online: https://repository.library.noaa.gov/view/noaa/4859 (accessed on 20 November 2022).
- Mase-Guthrie, B.; Townsend, F.; McFee, W.; Manire, C.; Ewing, R.Y. Cases of Prolonged Freshwater Exposure in Dolphins Along the Southeast United States. In Proceedings of the 16th Biennial Conference on the Biology of Marine Mammals, San Diego, CA, USA, 12–16 December 2005. [Google Scholar]
- Rice, K.C.; Hong, B.; Shen, J. Assessment of Salinity Intrusion in the James and Chickahominy Rivers as a Result of Simulated Sea-Level Rise in Chesapeake Bay, East Coast, USA. J. Environ. Manag. 2012, 111, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Carmen, M.; Berrow, S.D.; O’Brien, J.M. Foraging Behavior of Bottlenose Dolphins in the Shannon Estuary, Ireland as Determined through Static Acoustic Monitoring. J. Mar. Sci. Eng. 2021, 9, 275. [Google Scholar] [CrossRef]
- Fiori, L.; Doshi, A.; Martinez, E.; Orams, M.B.; Bollard-Breen, B. The Use of Unmanned Aerial Systems in Marine Mammal Research. Remote Sens. 2017, 9, 543. [Google Scholar] [CrossRef]
- Giles, A.B.; Butcher, P.A.; Colefax, A.P.; Pagendam, D.E.; Mayjor, M.; Kelaher, B.P. Responses of Bottlenose Dolphins (Tursiops spp.) to Small Drones. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 677–684. [Google Scholar] [CrossRef]
- Hodgson, A.; Peel, D.; Kelly, N. Unmanned Aerial Vehicles for Surveying Marine Fauna: Assessing Detection Probability. Ecol. Appl. 2017, 27, 1253–1267. [Google Scholar] [CrossRef] [PubMed]
- Linchant, J.; Lisein, J.; Semeki, J.; Lejeune, P.; Vermeulen, C. Are Unmanned Aircraft Systems (UASs) the Future of Wildlife Monitoring? A Review of Accomplishments and Challenges. Mammal Rev. 2015, 45, 239–252. [Google Scholar] [CrossRef]
- Colefax, A.P.; Butcher, P.A.; Pagendam, D.E.; Kelaher, B.P. Reliability of marine faunal detections in drone-based monitoring. Ocean Coast. Manag. 2019, 174, 108–115. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Colefax, A.P.; Tagliafico, A.; Bishop, M.J.; Giles, A.; Butcher, P.A. Assessing Variation in Assemblages of Large Marine Fauna off Ocean Beaches Using Drones. Mar. Freshw. Res. 2019, 71, 68–77. [Google Scholar] [CrossRef]
- Oliveira-da-Costa, M.; Marmontel, M.; da-Rosa, D.S.X.; Coelho, A.; Wich, S.; Mosquera-Guerra, F.; Trujillo, F. Effectiveness of Unmanned Aerial Vehicles to Detect Amazon Dolphins. Oryx 2020, 54, 696–698. [Google Scholar] [CrossRef]
- Torres, L.G.; Nieukirk, S.L.; Lemos, L.; Chandler, T.E. Drone Up! Quantifying Whale Behavior from a New Perspective Improves Observational Capacity. Front. Mar. Sci. 2018, 5, 319. [Google Scholar] [CrossRef]
- Stewart, J.D.; Durban, J.W.; Knowlton, A.R.; Lynn, M.S.; Fearnbach, H.; Barbaro, J.; Perryman, W.L.; Miller, C.A.; Moore, M.J. Decreasing Body Lengths in North Atlantic Right Whales. Curr. Biol. 2021, 31, 3174–3179.e3. [Google Scholar] [CrossRef]
- Cheney, B.J.; Dale, J.; Thompson, P.M.; Quick, N.J. Spy in the Sky: A Method to Identify Pregnant Small Cetaceans. Remote Sens. Ecol. Consev. 2022, 8, 492–505. [Google Scholar] [CrossRef]
- Fettermann de Oliveria, T. Unmanned Aerial Vehicle (UAV) Remote Sensing of Behaviour and Habitat Use of the Nationally Endangered Bottlenose Dolphin (Tursiops truncatus) off Great Barrier Island, New Zealand. Master’s Thesis, Auckland University of Technology, Auckland, New Zealand, 2018. [Google Scholar]
- Pinckney, J.; Dustan, P. Ebb-Tidal Fronts in Charleston Harbor, South Carolina: Physical and Biological Characteristics. Estuaries 1990, 13, 1–7. [Google Scholar] [CrossRef]
- Kjerfve, B.; Magill, K.E. Salinity changes in Charleston Harbor 1922–1987. J. Waterw. Port Coast. Ocean. Eng. 1990, 116, 153–168. [Google Scholar] [CrossRef]
- Van Dolah, R.F.; Wendt, P.H.; Wenner, E.L.; Sandifer, P.A. A Physical and Ecological Characterization of the Charleston Harbor Estuarine System; Executive Summary Submitted to the South Carolina Coastal Council: Charleston, SC, USA, 1990; pp. 1–12. [Google Scholar]
- Noren, S.R. Infant Carrying Behaviour in Dolphins: Costly Parental Care in an Aquatic Environment. Funct. Ecol. 2008, 22, 284–288. [Google Scholar] [CrossRef]
- U.S. Geological Survey Water Data for the Nation. Available online: https://waterdata.usgs.gov/nwis (accessed on 1 February 2022).
- Schemel, L.E. Simplified conversions between specific conductance and salinity units for use with data from monitoring stations. Interag. Ecol. Program Newsl. 2001, 14, 17–18. [Google Scholar]
- Conrads, P.A.; Darby, L.S. Development of a Coastal Drought Index Using Salinity Data. Bull. Am. Meteorol. Soc. 2017, 98, 753–766. [Google Scholar] [CrossRef]
- Petkewich, M.D.; McCloskey, B.J.; Rouen, L.F.; Conrads, P.A. Coastal Salinity Index for Monitoring Drought; U.S. Geological Survey Data Release; 2019. Available online: https://apps.usgs.gov/sawsc/csi/index.html (accessed on 1 February 2022).
- McCloskey, B. CSI: Coastal Salinity Index; R Package Version 0.0.1; 2022; Available online: https://rdrr.io/github/USGS-R/CSI/f/README.md (accessed on 1 February 2022).
- Shane, S.H. Behavior and Ecology of the Bottlenose Dolphin at Sanibel Island, Florida; Elsevier: Amsterdam, The Netherlands, 1990; pp. 245–265. [Google Scholar]
- Bossley, M.I.; Steiner, A.; Parra, G.J.; Saltré, F.; Peters, K.J. Dredging Activity in a Highly Urbanised Estuary Did Not Affect the Long-Term Occurrence of Indo-Pacific Bottlenose Dolphins and Long-Nosed Fur Seals. Mar. Pollut. Bull. 2022, 184, 114183. [Google Scholar] [CrossRef] [PubMed]
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression Models for Count Data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef]
- Zuur, A.F.; Saveliev, A.A.; Ieno, E.N. Zero Inflated Models and Generalized Linear Mixed Models with R; Highland Statistics Ltd.: Newburgh, UK, 2013; pp. 1–324. [Google Scholar]
- Potts, J.M.; Elith, J. Comparing Species Abundance Models. Ecol. Model. 2006, 199, 153–163. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Zero-Truncated and Zero-Inflated Models for Count Data. In Mixed Effects Models and Extensions in Ecology with R; Zuur, A.F., Ieno, E.N., Walker, N., Saveliev, A.A., Smith, G.M., Eds.; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; pp. 261–293. ISBN 978-0-387-87458-6. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Pate, S.M.; McFee, W.E. Prey Species of Bottlenose Dolphins (Tursiops truncatus) from South Carolina Waters. Southeast Nat. 2012, 11, 1–22. [Google Scholar] [CrossRef]
- Laska, D.; Speakman, T.; Fair, P.A. Community overlap of bottlenose dolphins (Tursiops truncatus) found in coastal waters near Charleston, South Carolina. JMATE 2011, 4, 10–18. [Google Scholar]
- Waring, G.T.; Josephson, E.; Maze-Foley, K.; Rosel, P.E.U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments 2010, NOAA Technical Memorandum, NMFS-NE-219; U.S. Department of Commerce: Washington, DC, USA, 2010.
- Tribble, C.; Monczak, A.; Transue, L.; Marian, A.; Fair, P.; Balmer, B.; Ballenger, J.; Baker, H.; Weinpress-Galipeau, M.; Robertston, A.; et al. Enhancing Interpretation of Cetacean Acoustic Monitoring: Investigating Factors that Influence Vocalization Patterns of Atlantic Bottlenose Dolphins in an Urbanized Estuary, Charleston Harbor, South Carolina, USA. Aquat. Mamm. 2023, 49, 519–549. [Google Scholar] [CrossRef]
- Young, R.F.; Phillips, H.D. Primary Production Required to Support Bottlenose Dolphins in a Salt Marsh Estuarine Creek System. Mar. Mammal Sci. 2002, 18, 358–373. [Google Scholar] [CrossRef]
- Bailey, H.; Thompson, P. Quantitative Analysis of Bottlenose Dolphin Movement Patterns and Their Relationship with Foraging. J. Anim. Ecol. 2006, 75, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Mintzer, V.J.; Fazioli, K.L. Salinity and Water Temperature as Predictors of Bottlenose Dolphin (Tursiops truncatus) Encounter Rates in Upper Galveston Bay, Texas. Front. Mar. Sci 2021, 8, 754686. [Google Scholar] [CrossRef]
- Ortiz, R.M. Osmoregulation in Marine Mammals. J. Exp. Biol. 2001, 204, 1831–1844. [Google Scholar] [CrossRef]
- Ridgway, S.; Venn-Watson, S. Effects of Fresh and Seawater Ingestion on Osmoregulation in Atlantic Bottlenose Dolphins (Tursiops truncatus). J. Comp. Physiol. B 2010, 180, 563–576. [Google Scholar] [CrossRef]
- Reif, J.S.; Schaefer, A.M.; Bossart, G.D.; Fair, P.A. Health and Environmental Risk Assessment Project for Bottlenose Dolphins Tursiops truncatus from the Southeastern USA. II. Environmental Aspects. Dis. Aquat. Org. 2017, 125, 155–166. [Google Scholar] [CrossRef]
- Fazioli, K.; Mintzer, V. Short-Term Effects of Hurricane Harvey on Bottlenose Dolphins (Tursiops truncatus) in Upper Galveston Bay, TX. Estuaries Coasts 2020, 43, 1013–1031. [Google Scholar] [CrossRef]
- McBride-Kebert, S.; Toms, C.N. Common Bottlenose Dolphin, Tursiops truncatus, Behavioral Response to a Record-Breaking Flood Event in Pensacola Bay, Florida. J. Zool. Bot. Gard. 2021, 2, 351–369. [Google Scholar] [CrossRef]
- NOAA Tides & Currents. Available online: https://tidesandcurrents.noaa.gov/sltrends/sltrends_station.shtml?id=8665530 (accessed on 1 January 2023).
- Chua, V.P.; Xu, M. Impacts of Sea-Level Rise on Estuarine Circulation: An Idealized Estuary and San Francisco Bay. J. Mar. Syst. 2014, 139, 58–67. [Google Scholar] [CrossRef]
- Fazioli, K.; Mintzer, V.; Guillen, G.; Loe, S. Texas’ estuarine bottlenose dolphins: Addressing knowledge gaps in Galveston Bay. In Proceedings of the 8th National Summit on Coastal and Estuarine Restoration, New Orleans, LA, USA, 10–15 December 2016. [Google Scholar]
- Moreno, M.P.T. Environmental Predictors of Bottlenose Dolphin Distribution and Core Feeding Densities in Galveston Bay, Texas. Ph.D. Dissertation, Texas A&M University, College Station, TX, USA, 2005. [Google Scholar]
- Ballenger, J.; (South Carolina Department of Natural Resources, Charleston, SC, USA). Personal communication, 2023.
- Scott, M.D.; Wells, R.S.; Irvine, A.B. A long-term study of bottlenose dolphins on the west coast of Florida. In The Bottlenose Dolphin; Leatherwood, S., Reeves, R.R., Eds.; Academic Press: Cambridge, MA, USA, 1989; pp. 235–244. [Google Scholar]
- Lang dos Santos, M.; Lemos, V.M.; Vieira, J.P. No mullet, no gain: Cooperation between dolphins and cast net fishermen in southern Brazil. Zoologica 2018, 35, 1–13. [Google Scholar] [CrossRef]
- McDonough, C.J.; Wenner, C.A. Growth, recruitment, and abundance of juvenile striped mullet (Mugil cephalus) in South Carolina Estuaries. Fish. Bull. 2003, 101, 343–357. [Google Scholar]
- Arnott, S.; Archambault, J.; Biondo, P.; DaVega, H.; Evitt, R.; Frazier, B.; Grosse, A.; Hein, J.; Johnson, J.; Levesque, E.; et al. Five Year Report to the Saltwater Recreational Fisheries Advisory Committee; SCDNR Inshore Fisheries Section: Charleston, SC, USA, 2013; pp. 1–146. [Google Scholar]
- Gerrodette, T.; Perryman, W.; Barlow, J. Calibrating group size estimates of dolphins in the Eastern Tropical Pacific Ocean. In National Marine Fisheries Service, National Oceanic and Atmospheric Administration, Southwest Fisheries Science Center Administrative Report LJ-02-08; 2002. Available online: https://swfsc-publications.fisheries.noaa.gov/publications/CR/1992/92105.PDF (accessed on 20 November 2022).
- Rosel, P.E.; Mullin, K.; Garrison, L.; Schwacke, L.; Adams, J.; Balmer, B.; Zolman, E.; Wells, R.; Wells, P.; Vollmer, N.; et al. Photo-Identification Capture-Mark-Recapture Techniques for Estimating Abundance of Bay, Sound, and Estuary Populations of Bottlenose Dolphins along the U.S. East Coast and Gulf of Mexico: A Workshop Report, NOAA Technical Memorandum NMFS-SEFSC-621); U.S. Department of Commerce: Washington, DC, USA, 2011.
- Aniceto, A.S.; Biuw, M.; Lindstrøm, U.; Solbø, S.A.; Broms, F.; Carroll, J. Monitoring Marine Mammals Using Unmanned Aerial Vehicles: Quantifying Detection Certainty. Ecosphere 2018, 9, e02122. [Google Scholar] [CrossRef]
- Fettermann, T.; Fiori, L.; Bader, M.; Doshi, A.; Breen, D.; Stockin, K.A.; Bollard, B. Behaviour Reactions of Bottlenose Dolphins (Tursiops truncatus) to Multirotor Unmanned Aerial Vehicles (UAVs). Sci. Rep. 2019, 9, 8558. [Google Scholar] [CrossRef]
- Ramos, E.A.; Maloney, B.; Magnasco, M.O.; Reiss, D. Bottlenose Dolphins and Antillean Manatees Respond to Small Multi-Rotor Unmanned Aerial Systems. Front. Mar. Sci. 2018, 5, 316. [Google Scholar] [CrossRef]
- Ramos, E.A. Adopting Small Unmanned Unmanned Aerial Systems for Behavioral Research with Coastal Marine Mammals. Ph.D. Dissertation, City University of New York, New York, NY, USA, 2022. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Total Months Surveyed | Total Flights Conducted | Total Flight Time (mins) | Area Covered (km2) | Temporal Data Gaps |
|---|---|---|---|---|---|
| Folly River (1) | 12 | 44 | 729.13 | 1.3 | August |
| Harbor–CR Confluence (2) | 13 | 55 | 944.77 | 2.2 | None |
| Mid-Cooper River (3) | 13 | 49 | 895.1 | 2.3 | None |
| Upper-Cooper River Site 1 (4) | 13 | 48 | 840.9 | 2.7 | None |
| Upper-Cooper River Site 2 (5) | 13 | 32 | 544.21 | 1.4 | None |
| UCR–Back River Confluence (6) | 9 | 21 | 332.26 | 1.5 | January 2021/2022, July, August |
| Back River Site 1 (7) | 11 | 20 | 307.62 | 0.2 | July, January 2022 |
| Back River Site 2 (8) | 13 | 27 | 404.04 | 0.5 | None |
| Study Site | Percentage of Flights w/Dolphins | Total Detections | Total Dolphins | Mom/Calf Pairs | Density (Total Dolphins/km2) | Mean (±SD) Group Size Per Detection |
|---|---|---|---|---|---|---|
| Folly River (1) | 54.5 | 27 | 49 | 8 | 37.7 | 3.27 (3.26) |
| Harbor–CR Confluence (2) | 49.1 | 34 | 70 | 7 | 31.8 | 2.59 (2.10) |
| Mid-Cooper River (3) | 34.7 | 23 | 48 | 2 | 20.9 | 2.53 (1.95) |
| Upper-Cooper River Site 1 (4) | 14.6 | 7 | 17 | 2 | 6.29 | 3.00 (1.83) |
| Upper-Cooper River Site 2 (5) | 9.4 | 3 | 5 | 2 | 3.57 | 2.33 (1.53) |
| UCR–Back River Confluence (6) | 0 | 0 | 0 | 0 | 0 | 0 |
| Back River Site 1 (7) | 0 | 0 | 0 | 0 | 0 | 0 |
| Back River Site 2 (8) | 0 | 0 | 0 | 0 | 0 | 0 |
| Overall | 26.4 | 94 | 189 | 21 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Principe, N.; McFee, W.; Levine, N.; Balmer, B.; Ballenger, J. Using Unoccupied Aerial Systems (UASs) to Determine the Distribution Patterns of Tamanend’s Bottlenose Dolphins (Tursiops erebennus) across Varying Salinities in Charleston, South Carolina. Drones 2023, 7, 689. https://doi.org/10.3390/drones7120689
Principe N, McFee W, Levine N, Balmer B, Ballenger J. Using Unoccupied Aerial Systems (UASs) to Determine the Distribution Patterns of Tamanend’s Bottlenose Dolphins (Tursiops erebennus) across Varying Salinities in Charleston, South Carolina. Drones. 2023; 7(12):689. https://doi.org/10.3390/drones7120689
Chicago/Turabian StylePrincipe, Nicole, Wayne McFee, Norman Levine, Brian Balmer, and Joseph Ballenger. 2023. "Using Unoccupied Aerial Systems (UASs) to Determine the Distribution Patterns of Tamanend’s Bottlenose Dolphins (Tursiops erebennus) across Varying Salinities in Charleston, South Carolina" Drones 7, no. 12: 689. https://doi.org/10.3390/drones7120689