The Impact of Biosurfactants on Microbial Cell Properties Leading to Hydrocarbon Bioavailability Increase





Abstract
1. Introduction
2. Hydrocarbon Emulsification and Desorption with Biosurfactants
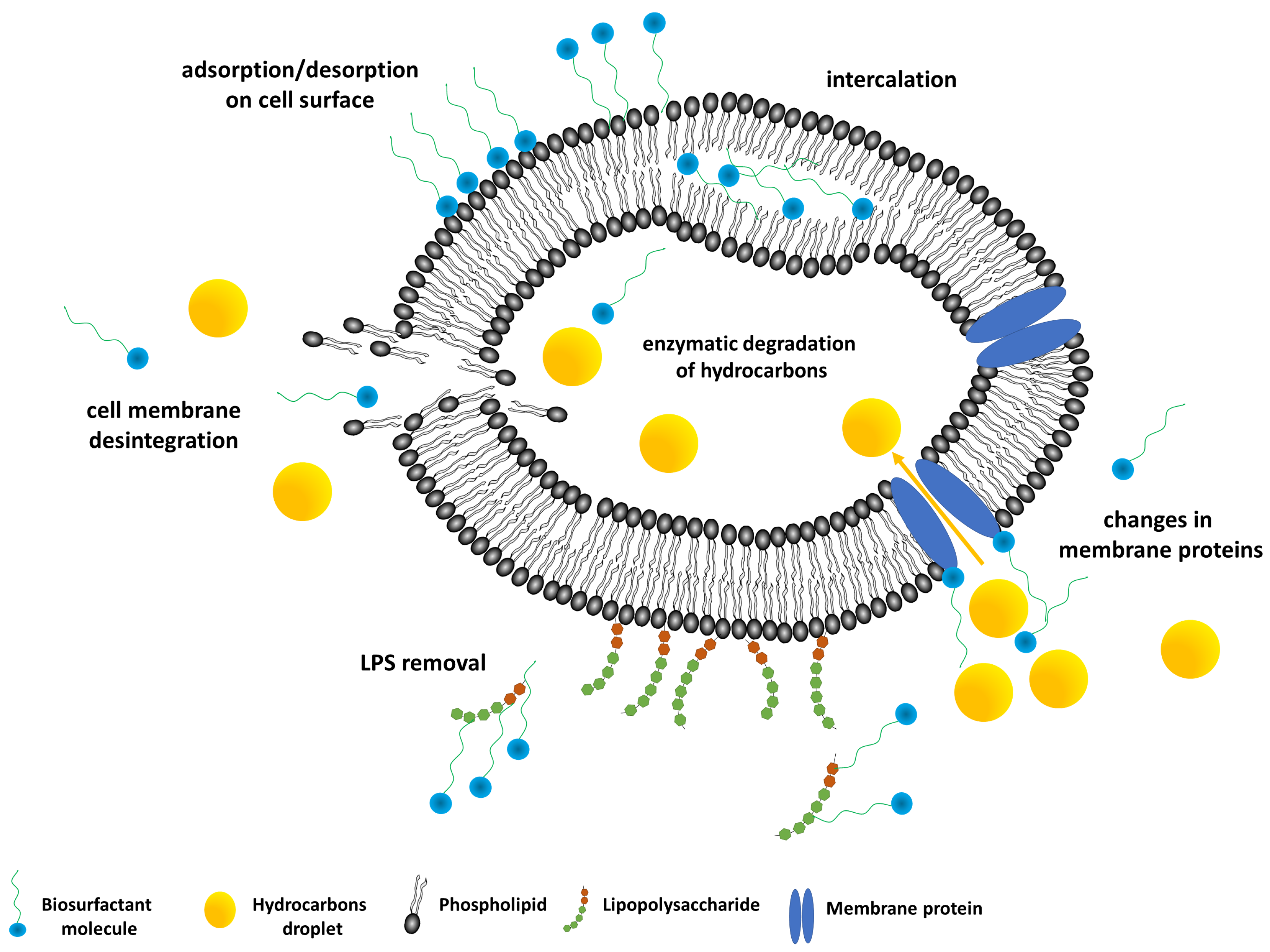
3. Biosurfactant–Bacteria Interactions
3.1. Impact on Microbial Cell Surface Properties
3.2. Impact on Microbial Cell Membranes
4. Biosurfactants-Enhanced Hydrocarbons Biodegradation
5. Conclusions
Funding
Conflicts of Interest
References
- De Almeida, D.G.; De Cássia, R.D.C.F.S.; Luna, J.M.; Rufino, R.D.; Santos, V.A.; Banat, I.M. Biosurfactants: Promising Molecules for Petroleum Biotechnology Advances. Front Microbiol. 2016, 7, 1718. [Google Scholar] [CrossRef] [PubMed]
- Nwankwegu, A.S.; Onwosi, C.O. Bioremediation of gasoline contaminated agricultural soil by bioaugmentation. Environ. Technol. Innov. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Liu, R.; Jadeja, R.N.; Zhou, Q.; Liu, Z. Treatment and Remediation of Petroleum-Contaminated Soils Using Selective Ornamental Plants. Environ. Eng. Sci. 2012, 29, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Cocârţă, D.; Stoian, M.; Karademir, A. Crude Oil Contaminated Sites: Evaluation by Using Risk Assessment Approach. Sustainability 2017, 9, 1365. [Google Scholar] [CrossRef]
- Bezza, F.A.; Nkhalambayausi Chirwa, E.M. Biosurfactant-enhanced bioremediation of aged polycyclic aromatic hydrocarbons (PAHs) in creosote contaminated soil. Chemosphere 2016, 144, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Maier, R.M. Bioavailability and Its Importance to Bioremediation. In Bioremediation; Valdes, J.J., Ed.; Springer: Dordrecht, The Netherlands, 2000; pp. 59–78. ISBN 978-94-015-9425-7. [Google Scholar]
- Megharaj, M.; Ramakrishnan, B.; Venkateswarlu, K.; Sethunathan, N.; Naidu, R. Bioremediation approaches for organic pollutants: A critical perspective. Environ. Int. 2011, 37, 1362–1375. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zeng, G.; Tang, L.; Wang, J.; Wan, J.; Liu, Y.; Yu, J.; Yi, H.; Ye, S.; Deng, R. Sorption, transport and biodegradation—An insight into bioavailability of persistent organic pollutants in soil. Sci. Total Environ. 2018, 610–611, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Liu, Y.; Zhong, H.; Xiao, R.; Zeng, G.; Liu, Z.; Cheng, M.; Lai, C.; Zhang, C.; Liu, G.; et al. Mechanisms for rhamnolipids-mediated biodegradation of hydrophobic organic compounds. Sci. Total Environ. 2018, 634, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.F.B.; Gudiña, E.J.; Costa, R.; Vitorino, R.; Teixeira, J.A.; Coutinho, J.A.P.; Rodrigues, L.R. Optimization and characterization of biosurfactant production by Bacillus subtilis isolates towards microbial enhanced oil recovery applications. Fuel 2013, 111, 259–268. [Google Scholar] [CrossRef]
- Bajaj, A.; Mayilraj, S.; Mudiam, M.K.R.; Patel, D.K.; Manickam, N. Isolation and functional analysis of a glycolipid producing Rhodococcus sp. strain IITR03 with potential for degradation of 1,1,1-trichloro-2,2-bis(4-chlorophenyl)ethane (DDT). Bioresour. Technol. 2014, 167, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, E.; Fodelianakis, S.; Korkakaki, E.; Kalogerakis, N. Biosurfactant production from marine hydrocarbon-degrading consortia and pure bacterial strains using crude oil as carbon source. Front. Microbiol. 2015, 6, 274. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Rufino, R.; Luna, J.; Santos, V.; Sarubbo, L. Biosurfactants: Multifunctional biomolecules of the 21st century. Int. J. Mol. Sci. 2016, 17, 401. [Google Scholar] [CrossRef] [PubMed]
- Mnif, I.; Chaabouni, S.E.; Ghribi, D. Glycolipid Biosurfactants, Main Classes, Functional Properties and Related Potential Applications in Environmental Biotechnology. J. Polym. Environ. 2018, 26, 2192–2206. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Das, K. Microbial Surfactants and Their Potential Applications: An Overview. Biosurfactants 2010, 672, 54–64. [Google Scholar]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Piotrowska-Seget, Z.; Cameotra, S.S. Environmental applications of biosurfactants: Recent advances. Int. J. Mol. Sci. 2011, 12, 633–654. [Google Scholar] [CrossRef] [PubMed]
- Mnif, I.; Ghribi, D. Lipopeptides biosurfactants: Mean classes and new insights for industrial, biomedical, and environmental applications. Biopolymers 2015, 104, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Irfan-Maqsood, M.; Seddiq-Shams, M. Rhamnolipids: Well-Characterized Glycolipids with Potential Broad Applicability as Biosurfactants. Ind. Biotechnol. 2014, 10, 4. [Google Scholar] [CrossRef]
- Fakruddin, M. Biosurfactant: Production and Application. J. Pet. Environ. Biotechnol. 2012, 3, 124. [Google Scholar] [CrossRef]
- Ali, S.; Hameed, S.; Imran, A.; Iqbal, M.; Lazarovits, G. Genetic, physiological and biochemical characterization of Bacillus sp. strain RMB7 exhibiting plant growth promoting and broad spectrum antifungal activities. Microb. Cell Fact. 2014, 13, 144. [Google Scholar] [CrossRef] [PubMed]
- Passed, A.; Lang, S.; Wagner, F.; Wray, V. Marine Biosurfactants, II. Production and Characterization of an Anionic Trehalose Tetraester from the Marine Bacterium Arthrobacter sp. EK 1. Zeitschrift für Naturforschung C 2014, 46, 204–209. [Google Scholar]
- Bezza, F.A.; Chirwa, E.M.N. Bioremediation of Polycyclic Aromatic Hydrocarbon Contaminated Soil by a Microbial Consortium through Supplementation of Biosurfactant Produced by Pseudomonas aeruginosa Strain. Polycycl. Aromat. Compd. 2016, 36. [Google Scholar] [CrossRef]
- Franzetti, A.; Tamburini, E.; Banat, I.M. Applications of Biological Surface Active Compounds in Remediation Technologies. Biosurfactants 2010, 672, 121–134. [Google Scholar]
- Singh, R.; Glick, B.R.; Rathore, D. Biosurfactants as a Biological Tool to Increase Micronutrient Availability in Soil: A Review. Pedosphere 2018, 28, 170–189. [Google Scholar] [CrossRef]
- Noordman, W.H.; Wachter, J.H.J.; De Boer, G.J.; Janssen, D.B. The enhancement by surfactants of hexadecane degradation by Pseudomonas aeruginosa varies with substrate availability. J. Biotechnol. 2002, 94, 195–212. [Google Scholar] [CrossRef]
- Patowary, K.; Patowary, R.; Kalita, M.C.; Deka, S. Development of an Efficient Bacterial Consortium for the Potential Remediation of Hydrocarbons from Contaminated Sites. Front. Microbiol. 2016, 7, 1092. [Google Scholar] [CrossRef] [PubMed]
- Jemil, N.; Hmidet, N.; Ben Ayed, H.; Nasri, M. Physicochemical characterization of Enterobacter cloacae C3 lipopeptides and their applications in enhancing diesel oil biodegradation. Process Saf. Environ. Prot. 2018, 117, 399–407. [Google Scholar] [CrossRef]
- Schippers, C.; Geßner, K.; Müller, T.; Scheper, T. Microbial degradation of phenanthrene by addition of a sophorolipid mixture. J. Biotechnol. 2000, 83, 189–198. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Q.; Liu, W.; Liu, X.; Hou, J.; Teng, Y.; Luo, Y.; Christie, P. Chemosphere Biosurfactant-producing microorganism Pseudomonas sp. SB assists the phytoremediation of DDT-contaminated soil by two grass species. Chemosphere 2017, 182, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Mnif, I.; Sahnoun, R.; Ellouze-chaabouni, S. Evaluation of B. subtilis SPB1 biosurfactants’ potency for diesel-contaminated soil washing: Optimization of oil desorption using Taguchi design. Environ. Sci. Pollut. 2014, 21, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, S.; Krishna, K.C.B.; Sarukkalige, R. Surfactant-enhanced remediation of polycyclic aromatic hydrocarbons: A review. J. Environ. Mana. 2017, 199, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Attwood, D.; Florence, A.T. Surfactant Systems: Their Chemistry, Pharmacy and Biology, Aspects of Surfactant Toxicity; Chaman & Hall: London, UK, 1983. [Google Scholar]
- Kaczorek, E.; Urbanowicz, M.; Olszanowski, A. The influence of surfactants on cell surface properties of Aeromonas hydrophila during diesel oil biodegradation. Colloid Surf. B 2010, 81, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Bueno-Montes, M.; Springael, D.; Ortega-Calvo, J.J. Effect of a nonionic surfactant on biodegradation of slowly desorbing PAHs in contaminated soils. Environ. Sci. Technol. 2011, 45, 3019–3026. [Google Scholar] [CrossRef] [PubMed]
- Paria, S. Surfactant-enhanced remediation of organic contaminated soil and water. Adv. Colloids Interface Sci. 2008, 138, 24–58. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Aitken, M.D. Surfactant-enhanced desorption and biodegradation of polycyclic aromatic hydrocarbons in contaminated soil. Environ. Sci. Technol. 2010, 44, 7260–7265. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. Bioavailability of organic compounds solubilized in nonionic surfactant micelles. Appl. Microbiol. Biotechnol. 2011, 89, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.; Wang, H.Q. Uptake and trans-membrane transport of petroleum hydrocarbons by microorganisms. Biotechnol. Biotechnol. Equip. 2014, 28, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Bishop, P.L. Influence of nonionic surfactant on attached biofilm formation and phenanthrene bioavailability during simulated surfactant enhanced bioremediation. Environ. Sci. Technol. 2007, 41, 7107–7113. [Google Scholar] [CrossRef] [PubMed]
- Kar, T.; Destain, J.; Thonart, P.; Delvigne, F. Scale-down assessment of the sensitivity of Yarrowia lipolytica to oxygen transfer and foam management in bioreactors: Investigation of the underlying physiological mechanisms. J. Ind. Microbiol. Biotechnol. 2012, 39, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Rafati Atri, M.; Ashrafizadeh, S.N. The importance of foams and antifoaming in bioprocesses. Pak. J. Biotechnol. 2010, 7, 19–39. [Google Scholar]
- Luna, J.M.; Rufino, R.D.; Sarubbo, L.A.; Campos-Takaki, G.M. Characterisation, surface properties and biological activity of a biosurfactant produced from industrial waste by Candida sphaerica UCP0995 for application in the petroleum industry. Colloid Surf. B 2013, 102, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Suganthi, S.H.; Murshid, S.; Sriram, S.; Ramani, K. Enhanced biodegradation of hydrocarbons in petroleum tank bottom oil sludge and characterization of biocatalysts and biosurfactants. J. Environ. Manag. 2018, 220, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Jemil, N.; Ben Ayed, H.; Hmidet, N.; Nasri, M. Characterization and properties of biosurfactants produced by a newly isolated strain Bacillus methylotrophicus DCS1 and their applications in enhancing solubility of hydrocarbon. World J. Microbiol. Biotechnol. 2016, 32, 175. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.Y.; Sun, M.Y.; Dong, W.; He, C.Q.; Chen, F.L.; Ma, Y.L. Effects of nutrition optimization strategy on rhamnolipid production in a Pseudomonas aeruginosa strain DN1 for bioremediation of crude oil. Biocatal. Agric. Biotechnol. 2016, 6, 144–151. [Google Scholar] [CrossRef]
- Peele, K.A.; Ch, V.R.T.; Kodali, V.P. Emulsifying activity of a biosurfactant produced by a marine bacterium. 3 Biotech. 2016, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.G.V.A.O.; Nitschke, M.; Lépine, F.; Déziel, E.; Contiero, J. Structure, properties and applications of rhamnolipids produced by Pseudomonas aeruginosa L2-1 from cassava wastewater. Process Biochem. 2010, 45, 1511–1516. [Google Scholar] [CrossRef]
- Lee, D.W.; Lee, H.; Kwon, B.O.; Khim, J.S.; Yim, U.H.; Kim, B.S.; Kim, J.J. Biosurfactant-assisted bioremediation of crude oil by indigenous bacteria isolated from Taean beach sediment. Environ. Pollut. 2018, 241, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Mukherji, S. Surfactant aided biodegradation of NAPLs by Burkholderia multivorans: Comparison between Triton X-100 and rhamnolipid JBR-515. Colloid Surf. B 2013, 102, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, J.; Dick, R.P.; Li, H.; Shen, D.; Gao, Y.; Waigi, M.G.; Ling, W. Rhamnolipid influences biosorption and biodegradation of phenanthrene by phenanthrene-degrading strain Pseudomonas sp. Ph6. Environ. Pollut. 2018, 240, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Congiu, E.; Ortega-Calvo, J. Role of Desorption Kinetics in the Rhamnolipid-Enhanced Biodegradation of Polycyclic Aromatic Hydrocarbons. Environ. Sci. Technol. 2014, 48, 10869–10877. [Google Scholar] [CrossRef] [PubMed]
- Bezza, F.A.; Chirwa, E.M.N. The role of lipopeptide biosurfactant on microbial remediation of aged polycyclic aromatic hydrocarbons (PAHs)-contaminated soil. Chem. Eng. J. 2017, 309, 563–576. [Google Scholar] [CrossRef]
- Bezza, F.A.; Nkhalambayausi-Chirwa, E.M. Desorption kinetics of polycyclic aromatic hydrocarbons (Pahs) from contaminated soil and the effect of biosurfactant supplementation on the rapidly desorbing fractions. Biotechnol. Biotechnol. Equip. 2015, 29, 680–688. [Google Scholar] [CrossRef]
- Yao, Y.; Huang, G.H.; An, C.J.; Cheng, G.H.; Wei, J. Effects of freeze-thawing cycles on desorption behaviors of PAH-contaminated soil in the presence of a biosurfactant: A case study in western Canada. Environ. Sci. Proc. Imp. 2017, 19, 874–882. [Google Scholar] [CrossRef]
- Crampon, M.; Cebron, A.; Portet-Koltalo, F.; Uroz, S.; Le Derf, F.; Bodilis, J. Low effect of phenanthrene bioaccessibility on its biodegradation in diffusely contaminated soil. Environ. Pollut. 2017, 225, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Huang, G.H.; Xiao, H.; Wang, L.; Chen, W. Combined effects of DOM and biosurfactant enhanced biodegradation of polycylic armotic hydrocarbons (PAHs) in soil-water systems. Environ. Sci. Pollut. Res. 2014, 21, 10536–10549. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, S.F.; Li, Y.; Xu, H.; Qin, L.; Tay, J.H. The influence of cell and substratum surface hydrophobicities on microbial attachment. J. Biotechnol. 2004, 110, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhong, H.; Yang, X.; Liu, Y.; Shao, B.; Liu, Z. Advances in applications of rhamnolipids biosurfactant in environmental remediation: A review. Biotechnol. Bioeng. 2018, 115, 796–814. [Google Scholar] [CrossRef] [PubMed]
- Silva-Dias, A.; Miranda, I.M.; Branco, J.; Monteiro-Soares, M.; Pina-Vaz, C.; Rodrigues, A.G. Adhesion, biofilm formation, cell surface hydrophobicity, and antifungal planktonic susceptibility: Relationship among Candida spp. Front Microbiol. 2015, 6, 205. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Krasowska, A.; Sigler, K. How microorganisms use hydrophobicity and what does this mean for human needs? Front. Cell. Infect. Microbiol. 2014, 4, 112. [Google Scholar] [CrossRef] [PubMed]
- Kaczorek, E.; Smułek, W.; Zdarta, A.; Sawczuk, A.; Zgoła-Grześkowiak, A. Influence of saponins on the biodegradation of halogenated phenols. Ecotoxicol. Environ. Saf. 2016, 131, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Moreno, A.M.; Navarro-Pérez, M.L.; Vadillo-Rodríguez, V.; Bruque, J.M.; González-Martín, M.L. Insights into bacterial contact angles: Difficulties in defining hydrophobicity and surface Gibbs energy. Colloid Surf. B 2011, 88, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Van der Mei, H.C.; Bos, R.; Busscher, H.J. A reference guide to microbial cell surface hydrophobicity based on contact angles. Colloid Surf. B 1998, 11, 213–221. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, L.; Li, F. Influences and mechanisms of surfactants on pyrene biodegradation based on interactions of surfactant with a Klebsiella oxytoca strain. Bioresour. Technol. 2013, 142, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Liu, S.; Tong, L.; Zhang, Y.; Yang, J.; Liu, W.; Guo, C.; Xie, Y.; Lu, G.; Dang, Z. Effects of rhamnolipids on the cell surface characteristics of Sphingomonas sp. GY2B and the biodegradation of phenanthrene. RSC Adv. 2017, 7, 24321–24330. [Google Scholar] [CrossRef]
- Kaczorek, E.; Olszanowski, A. Uptake of hydrocarbon by Pseudomonas fluorescens (P1) and Pseudomonas putida (K1) strains in the presence of surfactants: A cell surface modification. Water Air Soil Pollut. 2011, 214, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Selvam, A.; Wong, J.W.-C. Effects of rhamnolipids on cell surface hydrophobicity of PAH degrading bacteria and the biodegradation of phenanthrene. Bioresour. Technol. 2011, 102, 3999–4007. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, C.; Rappazzo, A.C.; Michaud, L.; De Domenico, E.; Rochera, C.; Camacho, A.; Lo Giudice, A. Efficiency in hydrocarbon degradation and biosurfactant production by Joostella sp. A8 when grown in pure culture and consortia. J. Environ. Sci. 2018, 67, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.V.; Karegoudar, T.B.; Nayak, A.S. Enhanced utilization of fluorene by Paenibacillus sp. PRNK-6: Effect of rhamnolipid biosurfactant and synthetic surfactants. Ecotoxicol. Environ. Saf. 2018, 151, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Al-Tahhan, R.A.; Sandrin, T.R.; Bodour, A.A.; Maier, R.M. Rhamnolipid-induced removal of lipopolysaccharide from Pseudomonas aeruginosa: Effect on cell surface properties and interaction with hydrophobic substrates. Appl. Environ. Microbiol. 2000, 66, 3262–3268. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; Wang, S.; Abuduaini, R.; Zhang, M.; Zhu, X.; Zhao, Y. Rhamnolipid-aided biodegradation of carbendazim by Rhodococcus sp. D-1: Characteristics, products, and phytotoxicity. Sci. Total Environ. 2017, 590–591, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Sotirova, A.V.; Spasova, D.I.; Galabova, D.N.; Karpenko, E.; Shulga, A. Rhamnolipid–biosurfactant permeabilizing effects on gram-positive and gram-negative bacterial strains. Curr. Microbiol. 2008, 56, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Sotirova, A.; Spasova, D.; Vasileva-Tonkova, E.; Galabova, D. Effects of rhamnolipid-biosurfactant on cell surface of Pseudomonas aeruginosa. Microbiol. Res. 2009, 164, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Chen, J.; Lun, S.; Wang, X. Influence of biosurfactants produced by Candida antarctica on surface properties of microorganism and biodegradation of n-alkanes. Water Res. 2003, 37, 4143–4150. [Google Scholar] [CrossRef]
- Sze, A.; Erickson, D.; Ren, L.; Li, D. Zeta-potential measurement using the Smoluchowski equation and the slope of the current-time relationship in electroosmotic flow. J. Colloid Interface Sci. 2003, 261, 402–410. [Google Scholar] [CrossRef]
- Martienssen, M.; Reichel, O.; Kohlweyer, U. Surface Properties of Bacteria from Different Wastewater Treatment Plants. Acta Biotechnol. 2001, 21, 207–225. [Google Scholar] [CrossRef]
- Kaczorek, E. Effect of External Addition of Rhamnolipids Biosurfactant on the Modification of Gram Positive and Gram Negative Bacteria Cell Surfaces during Biodegradation of Hydrocarbon Fuel Contamination. Polish J. Environ. Stud. 2012, 21, 901–909. [Google Scholar]
- Soni, K.A.; Balasubramanian, A.K.; Beskok, A.; Pillai, S.D. Zeta potential of selected bacteria in drinking water when dead, starved, or exposed to minimal and rich culture media. Curr. Microbiol. 2008, 56, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Wang, F.; Chen, H.; Li, M.; Qiao, F.; Liu, Z.; Hou, Y.; Wu, C.; Fan, Y.; Liu, L.; et al. Selective Antimicrobial Activities and Action Mechanism of Micelles Self-Assembled by Cationic Oligomeric Surfactants. ACS Appl. Mater. Interface 2016, 8, 4242–4249. [Google Scholar] [CrossRef] [PubMed]
- Koley, D.; Bard, A.J. Triton X-100 concentration effects on membrane permeability of a single HeLa cell by scanning electrochemical microscopy (SECM). Proc. Natl. Acad. Sci. USA 2010, 107, 16783–16787. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Jasmine, J.; Mukherji, S. Practical considerations and challenges involved in surfactant enhanced bioremediation of oil. Biomed Res. Int. 2013, 2013, 328608. [Google Scholar] [CrossRef] [PubMed]
- Ivanković, T.; Hrenović, J. Surfactants in the environment. Arch. Ind. Hyg. Toxicol. 2010, 61, 95–110. [Google Scholar]
- Scott, M.J.; Jones, M.N. The biodegradation of surfactants in the environment. Biochim. Biophys. Acta 2000, 1508, 235–251. [Google Scholar] [CrossRef]
- Shao, B.; Liu, Z.; Zhong, H.; Zeng, G.; Liu, G.; Yu, M.; Liu, Y.; Yang, X.; Li, Z.; Fang, Z.; et al. Effects of rhamnolipids on microorganism characteristics and applications in composting: A review. Microbiol. Res. 2017, 200, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Sperandeo, P.; Lau, F.K.; Carpentieri, A.; De Castro, C.; Molinaro, A.; Deho, G.; Silhavy, T.J.; Polissi, A. Functional Analysis of the Protein Machinery Required for Transport of Lipopolysaccharide to the Outer Membrane of Escherichia coli. J. Bacteriol. 2008, 190, 4460–4469. [Google Scholar] [CrossRef] [PubMed]
- Bharali, P.; Saikia, J.P.; Ray, A.; Konwar, B.K. Rhamnolipid (RL) from Pseudomonas aeruginosa OBP1: A novel chemotaxis and antibacterial agent. Colloid Surf. B 2013, 103, 502–509. [Google Scholar] [CrossRef] [PubMed]
- De Aguiar Cordeiro, R.; Weslley Caracas Cedro, E.; Raquel Colares Andrade, A.; Serpa, R.; José de Jesus Evangelista, A.; Sales de Oliveira, J.; Santos Pereira, V.; Pereira Alencar, L.; Bruna Leite Mendes, P.; Cibelle Soares Farias, B.; et al. Inhibitory effect of a lipopeptide biosurfactant produced by Bacillus subtilis on planktonic and sessile cells of Trichosporon spp. Biofouling 2018, 34, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Sana, S.; Datta, S.; Biswas, D.; Sengupta, D. Assessment of synergistic antibacterial activity of combined biosurfactants revealed by bacterial cell envelop damage. Biochim. Biophys. Acta 2018, 1860, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Vasileva-Tonkova, E.; Sotirova, A.; Galabova, D. The effect of rhamnolipid biosurfactant produced by Pseudomonas fluorescens on model bacterial strains and isolates from industrial wastewater. Curr. Microbiol. 2011, 62, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Vasileva-Tonkova, E.; Galabova, D.; Karpenko, E.; Shulga, A. Biosurfactant-rhamnolipid effects on yeast cells. Lett. Appl. Microbiol. 2001, 33, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Dusane, D.H.; Dam, S.; Nancharaiah, Y.V.; Kumar, A.; Venugopalan, V.P.; Zinjarde, S.S. Disruption of Yarrowia lipolytica biofilms by rhamnolipid biosurfactant. Aquat. Biosyst. 2012, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Teruel, J.A.; Aranda, F.J.; Ortiz, A. Molecular mechanism of membrane permeabilization by the peptide antibiotic surfactin. Biochim. Biophys. Acta 2003, 1611, 91–97. [Google Scholar] [CrossRef]
- Kim, L.H.; Jung, Y.; Kim, S.-J.; Kim, C.-M.; Yu, H.-W.; Park, H.-D.; Kim, I.S. Use of rhamnolipid biosurfactant for membrane biofouling prevention and cleaning. Biofouling 2015, 31, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, A.; Aranda, F.J.; Espuny, M.J.; Teruel, J.A.; Marqués, A.; Manresa, Á.; Ortiz, A. Mechanism of Membrane Permeabilization by a Bacterial Trehalose Lipid Biosurfactant Produced by Rhodococcus sp. Langmuir 2009, 25, 7892–7898. [Google Scholar] [CrossRef] [PubMed]
- Smułek, W.; Zdarta, A.; Pacholak, A.; Zgoła-Grześkowiak, A.; Marczak, Ł.; Jarzębski, M.; Kaczorek, E. Saponaria officinalis L. extract: Surface active properties and impact on environmental bacterial strains. Colloid Surf. B 2017, 150, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Zdarta, A.; Dudzińska-Bajorek, B.; Nowak, A.; Guzik, U.; Kaczorek, E. Impact of potent bioremediation enhancing plant extracts on Raoultella ornithinolytica properties. Ecotoxicol. Environ. Saf. 2017, 145, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Teruel, J.A.; Manresa, Á.; Espuny, M.J.; Marqués, A.; Aranda, F.J. Effects of a bacterial trehalose lipid on phosphatidylglycerol membranes. Biochim. Biophys. Acta 2011, 1808, 2067–2072. [Google Scholar] [CrossRef] [PubMed]
- Amro, N.A.; Kotra, L.P.; Wadu-Mesthrige, K.; Bulychev, A.; Mobashery, S.; Liu, G. High-Resolution Atomic Force Microscopy Studies of the Escherichiacoli Outer Membrane: Structural Basis for Permeability. Langmuir 2000, 16, 2789–2796. [Google Scholar] [CrossRef]
- Kallimanis, A.; Frillingos, S.; Drainas, C.; Koukkou, A.I. Taxonomic identification, phenanthrene uptake activity, and membrane lipid alterations of the PAH degrading Arthrobacter sp. strain Sphe3. Appl. Microbiol. Biotechnol. 2007, 76, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Murínová, S.; Dercová, K.; Čertík, M.; Lászlová, K. The adaptation responses of bacterial cytoplasmic membrane fluidity in the presence of environmental stress factors—Polychlorinated biphenyls and 3-chlorobenzoic acid. Biologia (Bratisl) 2014, 69. [Google Scholar] [CrossRef]
- Li, F.; Zhu, L. Surfactant-modified fatty acid composition of Citrobacter sp. SA01 and its effect on phenanthrene transmembrane transport. Chemosphere 2014, 107, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Tecon, R.; Van Der Meer, J.R. Effect of two types of biosurfactants on phenanthrene availability to the bacterial bioreporter Burkholderia sartisoli strain RP037. App. Microbiol. Biotechnol. 2010, 84, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Otzen, D.E. Biosurfactants and surfactants interacting with membranes and proteins: Same but different? Biochim. Biophys. Acta 2017, 1859, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Aranda, F.J.; Teruel, J.A.; Espuny, M.J.; Marqués, A.; Manresa, Á.; Ortiz, A. Permeabilization of biological and artificial membranes by a bacterial dirhamnolipid produced by Pseudomonas aeruginosa. J. Colloid Interface Sci. 2010, 341, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Heipieper, H.J.; Neumann, G.; Cornelissen, S.; Meinhardt, F. Solvent-tolerant bacteria for biotransformations in two-phase fermentation systems. Appl. Microbiol. Biotechnol. 2007, 74, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Kastell, A.; Knorr, D.; Smetanska, I. Exudation: An expanding technique for continuous production and release of secondary metabolites from plant cell suspension and hairy root cultures. Plant Cell Rep. 2012, 31, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Yuniati, M.D. Bioremediation of petroleum-contaminated soil: A Review. IOP Conf. Ser. Earth Environ. Sci. 2018, 118, 012063. [Google Scholar] [CrossRef]
- Mao, X.; Jiang, R.; Xiao, W.; Yu, J. Use of surfactants for the remediation of contaminated soils: A review. J. Hazard. Mater. 2015, 285, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Liu, G.; Jiang, Y.; Yang, J.; Liu, Y.; Yang, X.; Liu, Z.; Zeng, G. Transport of bacteria in porous media and its enhancement by surfactants for bioaugmentation: A review. Biotechnol. Adv. 2017, 35, 490–504. [Google Scholar] [CrossRef] [PubMed]
- Perfumo, A.; Smyth, T.J.P.; Marchant, R.; Banat, I.M. Production and roles of biosurfactants and bioemulsifiers in accessing hydrophobic substrates. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin Heidelberg, Germany, 2010; pp. 1501–1512. [Google Scholar]
- Zhang, G.; Wu, Y.; Qian, X.; Meng, Q. Biodegradation of crude oil by Pseudomonas aeruginosa in the presence of rhamnolipids. J. Zhejiang Univ. Sci. 2005, 6B, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Miller, R.M. Effect of rhamnolipid (biosurfactant) structure on solubilization and biodegradation of n-alkanes. Appl. Environ. Microbiol. 1995, 61, 2247–2251. [Google Scholar] [PubMed]
- Bramwell, D.A.P.; Laha, S. Effects of surfactant addition on the biomineralization and microbial toxicity of phenanthrene. Biodegradation 2000, 11, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Urum, K.; Pekdemir, T. Evaluation of biosurfactants for crude oil contaminated soil washing. Chemesphere 2004, 57, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, I.; Mukherji, S. Diverse effect of surfactants on pyrene biodegradation by a Pseudomonas strain utilizing pyrene by cell surface hydrophobicity induction. Int. Biodeterior. Biodegrad. 2016, 108, 67–75. [Google Scholar] [CrossRef]
- Tian, W.; Yao, J.; Liu, R.; Zhu, M.; Wang, F.; Wu, X.; Liu, H. Effect of natural and synthetic surfactants on crude oil biodegradation by indigenous strains. Ecotoxicol. Environ. Saf. 2016, 121, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Thavasi, R.; Jayalakshmi, S.; Banat, I.M. Application of biosurfactant produced from peanut oil cake by Lactobacillus delbrueckii in biodegradation of crude oil. Bioresour. Technol. 2011, 102, 3366–3372. [Google Scholar] [CrossRef] [PubMed]
- Sobrinho, H.B.S.; Rufino, R.D.; Luna, J.M.; Salgueiro, A.A.; Campos-Takaki, G.M.; Leite, L.F.C.; Sarubbo, L.A. Utilization of two agroindustrial by-products for the production of a surfactant by Candida sphaerica UCP0995. Process Biochem. 2008, 43, 912–917. [Google Scholar] [CrossRef]
- Shavandi, M.; Mohebali, G.; Haddadi, A.; Shakarami, H.; Nuhi, A. Emulsification potential of a newly isolated biosurfactant-producing bacterium, Rhodococcus sp. strain TA6. Colloid Surf. B 2011, 82, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Beal, R.; Betts, W.B. Role of rhamnolipid biosurfactants in the uptake and mineralization of hexadecane in Pseudomonas aeruginosa. J. Appl. Microbiol. 2000, 89, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Liu, Z.; Zhong, H.; Li, J.; Yuan, X.; Fu, H.; Ding, Y.; Wang, J.; Zhou, M. Effect of monorhamnolipid on the degradation of n-hexadecane by Candida tropicalis and the association with cell surface properties. Appl. Microbiol. Biotechnol. 2011, 90, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K.S.M.; Rahman, T.J.; Kourkoutas, Y.; Petsas, I.; Marchant, R.; Banat, I.M. Enhanced bioremediation of n-alkane in petroleum sludge using bacterial consortium amended with rhamnolipid and micronutrients. Bioresour. Technol. 2003, 90, 159–168. [Google Scholar] [CrossRef]
- Zhang, H.F.; Yang, X.H.; Wang, Y. Microwave assisted extraction of secondary metabolites from plants: Current status and future directions. Trends Food Sci. Technol. 2011, 22, 672–688. [Google Scholar] [CrossRef]
- Whang, L.M.; Liu, P.W.G.; Ma, C.C.; Cheng, S.S. Application of rhamnolipid and surfactin for enhanced diesel biodegradation-Effects of pH and ammonium addition. J. Hazard. Mater. 2009, 164, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Yan, J.; Wen, J.; Hu, Y.; Chen, Y.; Wu, W. Rhamnolipid-enhanced aerobic biodegradation of triclosan (TCS) by indigenous microorganisms in water-sediment systems. Sci. Total Environ. 2016, 571, 1304–1311. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Singh, H.; Raj, M. Ecotoxicology and Environmental Safety Rhamnolipid mediated enhanced degradation of chlorpyrifos by bacterial consortium in soil-water system. Ecotoxicol. Environ. Saf. 2016, 134, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Price, N.P.J.; Ray, K.J.; Vermillion, K.E.; Dunlap, C.A.; Kurtzman, C.P. Structural characterization of novel sophorolipid biosurfactants from a newly identified species of Candida yeast. Carbohydr. Res. 2012, 348, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Kim, Y.B.; Shin, J.D.; Kim, E.K. Enhanced biodegradation of hydrocarbons in soil by microbial biosurfactant, sophorolipid. Appl. Biochem. Biotechnol. 2010, 160, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.G.; Soares da Silva, R.D.C.F.; Brasileiro, P.P.F.; de Luna, J.M.; Da Silva, M.D.G.C.; Rufino, R.D.; Costa, A.F.S.; Santos, V.A.; Sarubbo, L.A. Application of a biosurfactant from Candida tropicalis ucp 0996 produced in low-cost substrates for hydrophobic contaminants removal. Chem. Eng. Trans. 2018, 64, 541–546. [Google Scholar] [CrossRef]
- Sajna, K.V.; Sukumaran, R.K.; Gottumukkala, L.D.; Pandey, A. Crude oil biodegradation aided by biosurfactants from Pseudozyma sp. NII 08165 or its culture broth. Bioresour. Technol. 2015, 191, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.; Durán, N.; Diez, M.C. Biosurfactants are useful tools for the bioremediation of contaminated soil: A review. J. Soil Sci. Plant Nutr. 2012, 12, 667–687. [Google Scholar] [CrossRef]
- Swaathy, S.; Kavitha, V.; Pravin, A.S.; Mandal, A.B.; Gnanamani, A. Microbial surfactant mediated degradation of anthracene in aqueous phase by marine Bacillus licheniformis MTCC 5514. Biotechnol. Rep. 2014, 4, 161–170. [Google Scholar] [CrossRef] [PubMed]
- McKew, B.A.; Coulon, F.; Yakimov, M.M.; Denaro, R.; Genovese, M.; Smith, C.J.; Osborn, A.M.; Timmis, K.N.; McGenity, T.J. Efficacy of intervention strategies for bioremediation of crude oil in marine systems and effects on indigenous hydrocarbonoclastic bacteria. Environ. Microbiol. 2007, 9, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.H.; Zhan, X.H.; Wang, S.M.; Lin, Y.S.; Zhou, L.X. Effects of a Biosurfactant and a Synthetic Surfactant on Phenanthrene Degradation by a Sphingomonas Strain. Pedosphere 2010, 20, 771–779. [Google Scholar] [CrossRef]
- Nazari, M.; Kurdi, M.; Heerklotz, H. Classifying Surfactants with Respect to Their Effect on Lipid Membrane Order. Biophys. J. 2012, 102, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Kowalewski, E.; Rueslåtten, I.; Steen, K.H.; Bødtker, G.; Torsæter, O. Microbial improved oil recovery—Bacterial induced wettability and interfacial tension effects on oil production. J. Pet. Sci. Eng. 2006, 52, 275–286. [Google Scholar] [CrossRef]
- Kuppusamy, S.; Thavamani, P.; Venkateswarlu, K.; Lee, Y.B.; Naidu, R.; Megharaj, M. Remediation approaches for polycyclic aromatic hydrocarbons (PAHs) contaminated soils: Technological constraints, emerging trends and future directions. Chemosphere 2017, 168, 944–968. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, B.; Chen, B.; Cai, Q.; Lin, W. Biosurfactant Production by Marine-Originated Bacteria Bacillus Subtilis and Its Application for Crude Oil Removal. Water. Air. Soil Pollut. 2016, 227. [Google Scholar] [CrossRef]
- Vilela, W.F.D.; Fonseca, S.G.; Oliveira, V.M.; Nitschke, M. Production and Properties of a Surface-Active Lipopeptide Produced by a New Marine Brevibacterium luteolum Strain. Appl. Biochem. Biotechnol. 2014, 2245–2256. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, G.J.; Ciapina, E.M.P.; de Barros Gomes, E.; Pereira Junior, N. Biosurfactant production by Rhodococcus erythropolis and its application to oil removal. Braz. J. Microbiol. 2010, 41, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Makkar, R.S.; Cameotra, S.S. Biosurfactant production by microorganisms on unconventional carbon sources. J. Surfactants Deterg. 1999, 2, 237–241. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Class | Subclass | Examples of Producers |
|---|---|---|
| Glycolipids | Rhamnolipids | Pseudomonas aeruginosa Pseudomonas sp. |
| Trehalolipids | Mycobacterium tuberculosis Nocardia sp. Corynebacterium sp. Rhodococcus erythropolis Micrococcus luteus | |
| Sophorolipids | Candida bombicola Candida magnolia Candida apicola Candida bogoriensis Torulopsis bombicola Torulopsis petrophilum Torulopsis apicola | |
| Xylolipids | Lactococcus lactis Pichia caribbica | |
| Cellobiolipid | Cryptococcus humicola | |
| Lipopeptides | Surfactin | Bacillus subtilis |
| Iturin | Bacillus subtilis | |
| Fengycin | Bacillus subtilis | |
| Lichenysin | Bacillus licheniformis | |
| Viscosinamid | P. fluorescens | |
| Viscosin | P. libanensis | |
| Flavolipid | Flavobacterium sp. | |
| Fatty Acid Biosurfactant | Arthobacter sp. Pseudomonas aeruginosa | |
| Polymeric Biosurfactants | Emulsan | Acinetobacter calcoaceticus |
| Biodispersan | Acinetobacter calcoaceticus | |
| Alasan | Acinetobacter radioresistens | |
| Liposan | Candida lipolytica |
| Biosurfactant | Biourfactant Source | Surfactant Concentration | Affected Microorganisms | Effect | Method | Additional Factors | Reference | |
|---|---|---|---|---|---|---|---|---|
| Cell surface Hydrophobicity | Rhamnolipid | N/D | 0–8 CMC | Klebsiella oxytoca |
| BATH | - | [65] |
| Biosurfactant | Joostella sp. A8 | N/D | Joostella sp. A8 Pseudomonas A6 Joostella sp. A8 Alcanivorax A53 consortium |
| BATH | cultures supplemented with diesel oil | [69] | |
| Rhamnolipids (Purity ≥ 98%) | Huzhou Zijin Biological Technology Company | 0; 0.2; 1 and 4 CMC | Sphingomonas sp. GY2B |
| MATH | cells analyzed after 24 h phenanthrene degradation | [66] | |
| Rhamnolipid | Pseudoxanthomo-nas sp. PNK-04 | 25 mg/L | Paenibacillus sp. PRNK-6 |
| BATH | - | [70] | |
| Biosurfactant BS-UC | Candida antarctica | 0–3% | Candida antarctica |
| MATH | cells cultured with 8% (v/v) of undecane, hexadecane, soybean oil, or glucose | [75] | |
| Rhamnolipid (90%) | AGAE Technologies | 0; 5; 50; 100; 200 and 400 mg L−1 | Pseudomonas sp. Ph6 |
| MATH | cells cultured with phenanthrene addition (50 mg L−1) | [50] | |
| Rhamnolipid | Jeneil Biosurfactant Company, USA | 2 CMC | Burkholderia multivorans |
| water contact angle | addition of 0.1% (v/v) NAPL | [49] | |
| Rhamnolipids JBR 425 | Jeneil Biosurfactant Company, USA | 6; 30; 60; 120; 150; 240; 360 mg L−1 | Pseudomonas fluorescens (P1) Pseudomonas putida K1 |
| MATH | different carbon sources with surfactants or surfactants only | [67] | |
| Rhamnolipids | P. aeruginosa ATCC9027 | 0, 20, 40, 120 and 400 mg L−1 | B. subtilis BUM P. aeruginosa P-CG3 |
| nitrocellulose filter test | single or mixed bacterial strains before and after PAH degradation | [68] | |
| Rhamnolipid-Biosurfactant PS | Pseudomonas sp. PS-17 | 50; 300 μg mL−1 | Pseudomonas aeruginosa NBIMCC 1390 |
| BATH | - | [74] | |
| Rhamnolipids RL (90% Purity) | Gemking Biotechnology Ltd. (Huzhou, China) | 0; 50; 150 ppm | Rhodococcus sp. D-1 |
| BATH and water contact angle | Cells cultured with 200 ppm of Carbendazim | [72] | |
| Biomorphology changes | Rhamnolipid-Biosurfactant PS | Pseudomonas sp. PS-17 | 50; 300 μg mL−1 | Pseudomonas aeruginosa NBIMCC 1390 |
| TEM | - | [74] |
| Rhamnolipid-Biosurfactant PS (Rhamnolipid + Alginate) | Pseudomonas sp. PS-17 | 0.5% | Bacillus subtilis 168 |
| SEM | - | [73] | |
| Rhamnolipids RL (90% Purity) | Gemking Biotechnology Ltd. (Huzhou, China) | 0; 50; 150 ppm | Rhodococcus sp. D-1 |
| SEM | cells cultured with 200 ppm of Carbendazim | [72] | |
| Rhamnolipid (90%) | AGAE Technologies | 0; 5; 50; 100; 200 and 400 mg L−1 | Pseudomonas sp. Ph6 |
| TEM | cells cultured with phenanthrene addition (50.0 mg L−1) | [50] | |
| Rhamnolipids (Purity ≥ 98%) | Huzhou Zijin Biological Technology Company | 0, 0.2; 1 and 4 CMC | Sphingomonas sp. GY2B |
| SEM | cells analyzed after 24 and 48 h phenanthrene degradation | [66] | |
| Zeta potential | Rhamnolipids (Purity ≥ 98%) | Huzhou Zijin Biological Technology Company | 0; 0.2; 1 and 4 CMC | Sphingomonas sp. GY2B |
| laser Doppler velocimetry | cells analyzed after 24 h phenanthrene degradation | [66] |
| Biosurfactants BS-UC and MEL | C. antarctica | 0; 1%; 2% and 3% | C. antarctica |
| dynamic light scattering | - | [75] | |
| Rhamnolipid (90%) | AGAE Technologies | 0; 5; 50; 100; 200 and 400 mg L−1 | Pseudomonas sp. Ph6 |
| Zetaphore-meter (Les Essarts-le- Roi, France) | cells cultured with phenanthrene addition (50 mg L−1) | [50] | |
| Rhamnolipid JBR 515 | Jeneil Biosurfactant Company, USA | 2 CMC | Burkholderia multivorans |
| Zeta potential analyzer (Zeta Pals) | Addition of 0.1% (v/v) NAPL | [49] | |
| Rhamnolipids RL (90% Purity) | Gemking Biotechnology Ltd. (Huzhou, China) | 0; 50; 150 ppm | Rhodococcus sp. D-1 |
| ZetaSizer Nano-ZS Zen 3600 | Cells cultured with 200 ppm of Carbendazim | [72] | |
| Cell surface Functional Groups | Rhamnolipids (Purity ≥ 98%) | Huzhou Zijin Biological Technology Company | 0; 0.2 and 1 CMC | Sphingomonas sp. GY2B |
| FTIR | freeze dried cells analyzed after 24h phenanthrene degradation | [66] |
| Rhamnolipid (90%) | AGAE Technologies | 0; 5; 50; 100; 200 and 400 mg L−1 | Pseudomonas sp. Ph6 |
| FTIR | cells cultured with phenanthrene addition (50 mg L−1) | [50] | |
| Rhamnolipidsrl (90% Purity) | Gemking Biotechnology Ltd. (Huzhou, China) | 0; 50; 150 ppm | Rhodococcus sp. D-1 |
| FTIR | Cells cultured with 200 ppm of Carbendazim | [72] | |
| Rhamnolipid JBR 515 | Jeneil Biosurfactant Company, USA | 2 CMC | Burkholderia multivorans |
| surface complexation modeling (potentiometric titration) | Addition of 0.1% (v/v) NAPL | [49] |
| Type of BS | Source of BS | Microorganism Tested | Method of Analysis | Impact of BS on CMP | References |
|---|---|---|---|---|---|
| Rhamnolipids | Pseudomonas aeruginosa C2 | E. coli | REP, CV | + | [89] |
| S, aureus | REP, CV | + | |||
| Lipopeptide Type BS15 | Bacillus stratosphericus A15 | E. coli | REP, CV | + | |
| S. aureus | REP, CV | + | |||
| Biosurfactant PS | Pseudomonas sp. S-17 | Saccharomyces cerevisiae 83-20 | REP & enzymatic activity | + | [91] |
| Rhamnolipids | N/D | Y. lipolytica NCIM 3589 | The methylene blue dye exclusion assay | Weak + | [92] |
| Surfactin, Iturin and Fengicin (Mixture) | Bacillus subtilis | Trichosporon spp. | REP NuclA | 0 + | [88] |
| Biosurfactant PS | Pseudomonas sp. S-17 | Bacillus subtilis 168 | REP | + (up to 0,01% of BS) − (above 0,01% of BS) | [75] |
| Pseudomonas aeruginosa NBIMCC 1390 | REP | + | |||
| Rhamnolipids | Pseudomonas fluorescens HW-6 | Bacillus sp. HW-4, Arthrobacter sp. HW-7 Streptococcus sp. HW-9 Micrococcus sp. HW-11 Pseudomonas sp. HW-1 Pseudomonas sp. HW-10 Pseudomonas sp. HW-12 Escherichia sp. HW-13 | REP | + | [90] |
| Rhamnolipids | N/D | P. aeruginosa P60 | Fluorescence | + | [94] |
| Rhamnolipids | P. aeruginosa OBP1 | Klebsiella pneumoniae Staphylococcus aureus | REP, CV | + | [87] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczorek, E.; Pacholak, A.; Zdarta, A.; Smułek, W. The Impact of Biosurfactants on Microbial Cell Properties Leading to Hydrocarbon Bioavailability Increase. Colloids Interfaces 2018, 2, 35. https://doi.org/10.3390/colloids2030035
Kaczorek E, Pacholak A, Zdarta A, Smułek W. The Impact of Biosurfactants on Microbial Cell Properties Leading to Hydrocarbon Bioavailability Increase. Colloids and Interfaces. 2018; 2(3):35. https://doi.org/10.3390/colloids2030035
Chicago/Turabian StyleKaczorek, Ewa, Amanda Pacholak, Agata Zdarta, and Wojciech Smułek. 2018. "The Impact of Biosurfactants on Microbial Cell Properties Leading to Hydrocarbon Bioavailability Increase" Colloids and Interfaces 2, no. 3: 35. https://doi.org/10.3390/colloids2030035
APA StyleKaczorek, E., Pacholak, A., Zdarta, A., & Smułek, W. (2018). The Impact of Biosurfactants on Microbial Cell Properties Leading to Hydrocarbon Bioavailability Increase. Colloids and Interfaces, 2(3), 35. https://doi.org/10.3390/colloids2030035