
Boreal (Eemian) Transgression in the Northeastern White Sea Region: Multiproxy Evidence from Bychye-2 Section
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
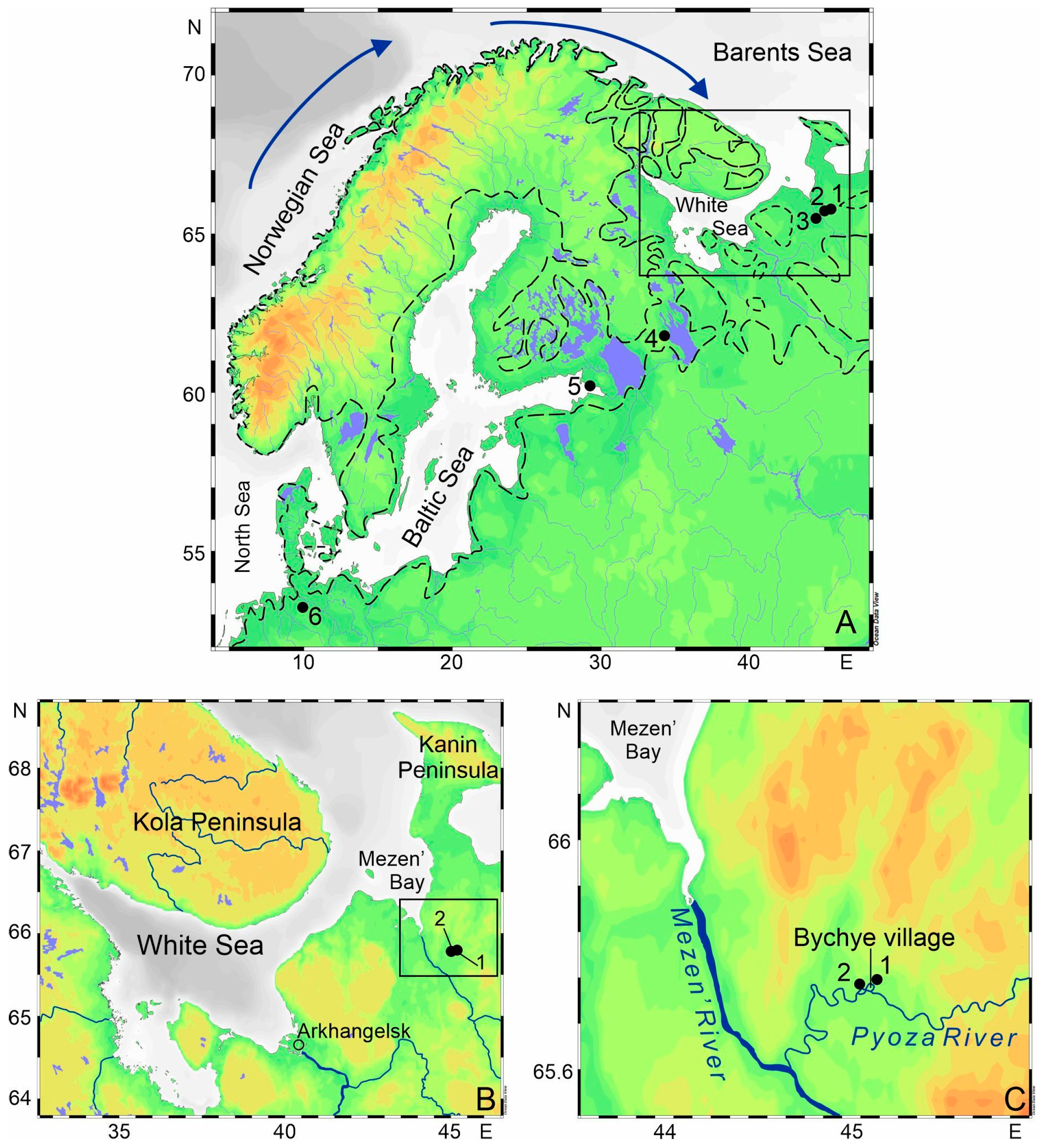
2. Materials and Methods
3. Results
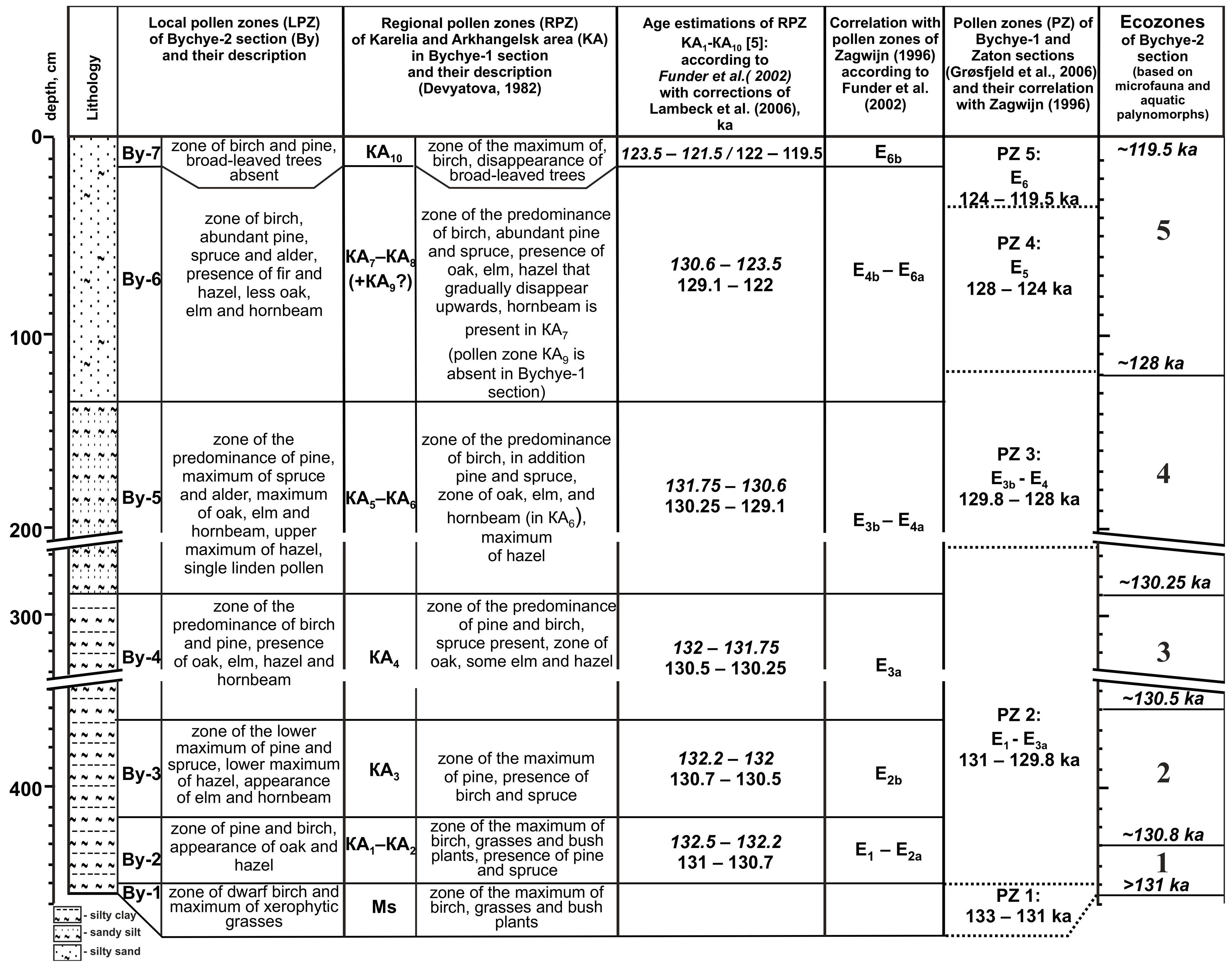
3.1. Pollen-Based Stratigraphy

3.2. Lithology
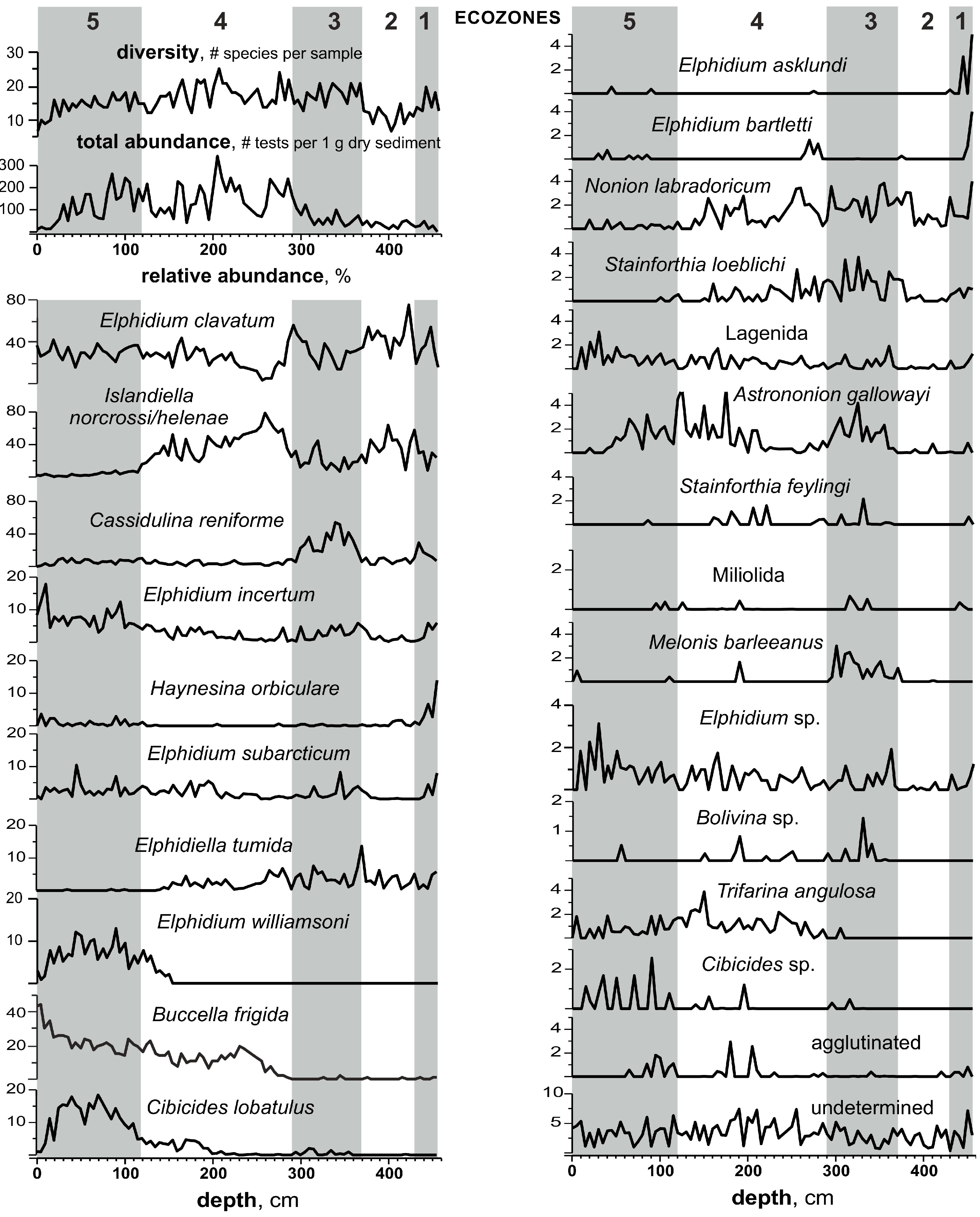
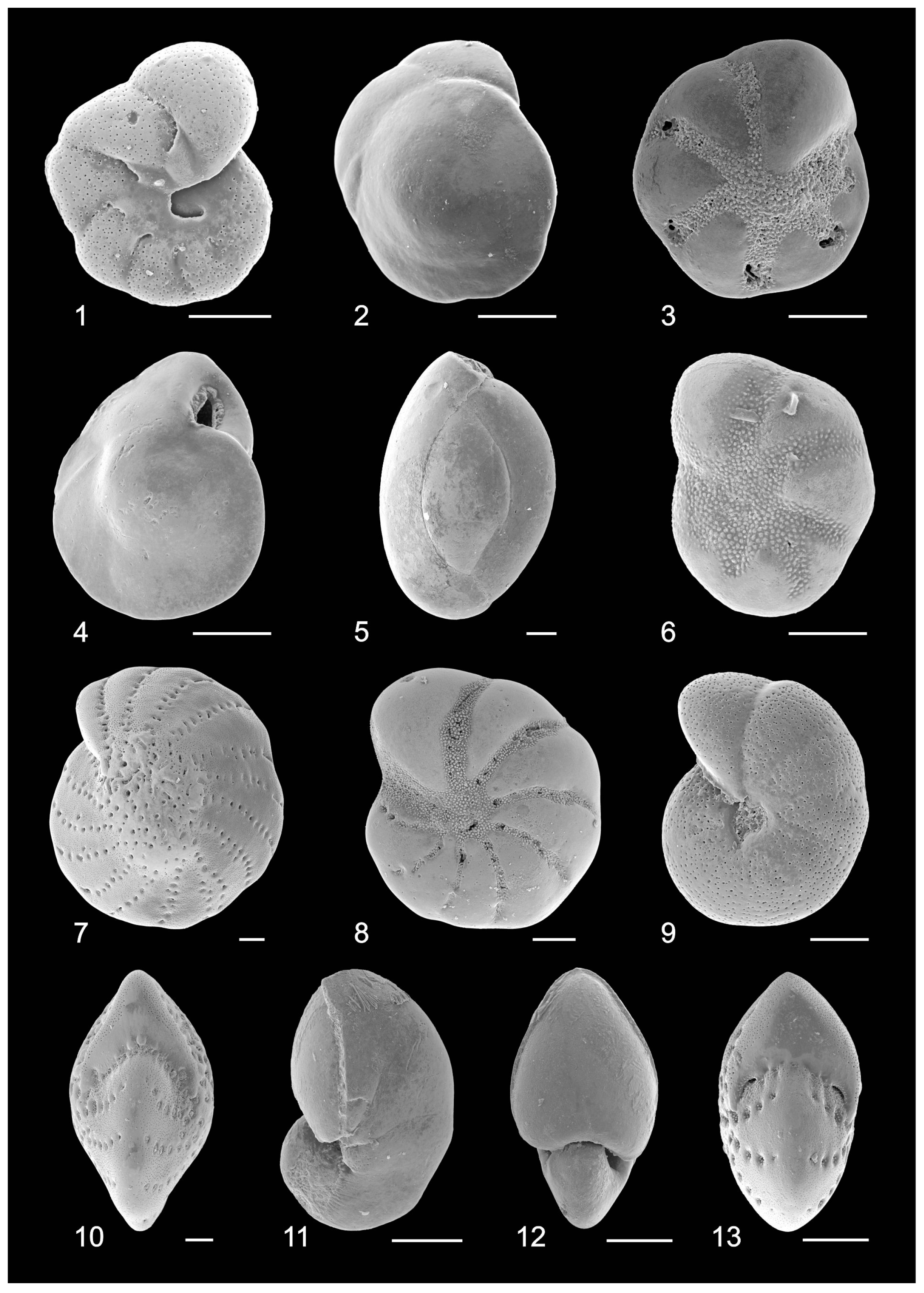
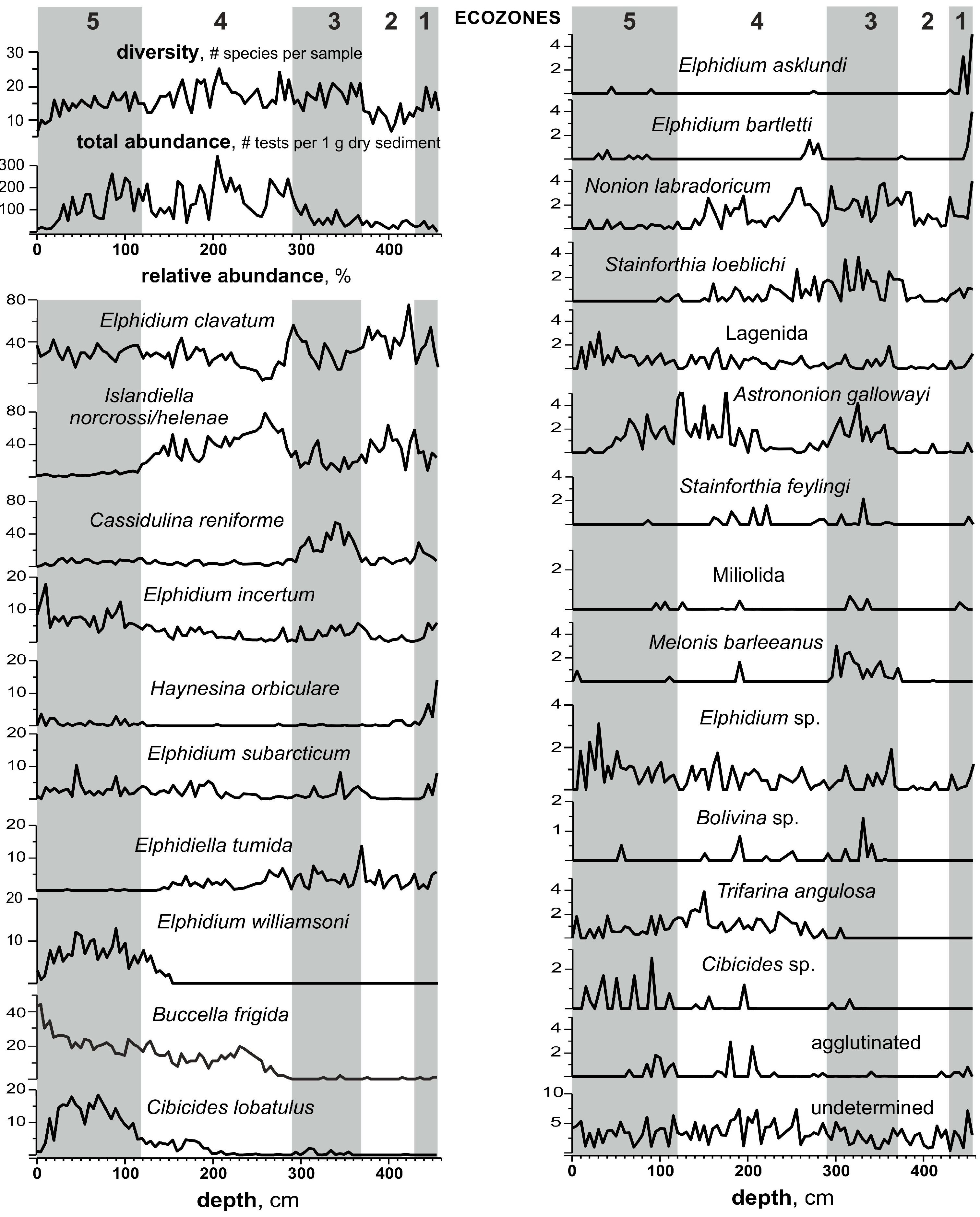
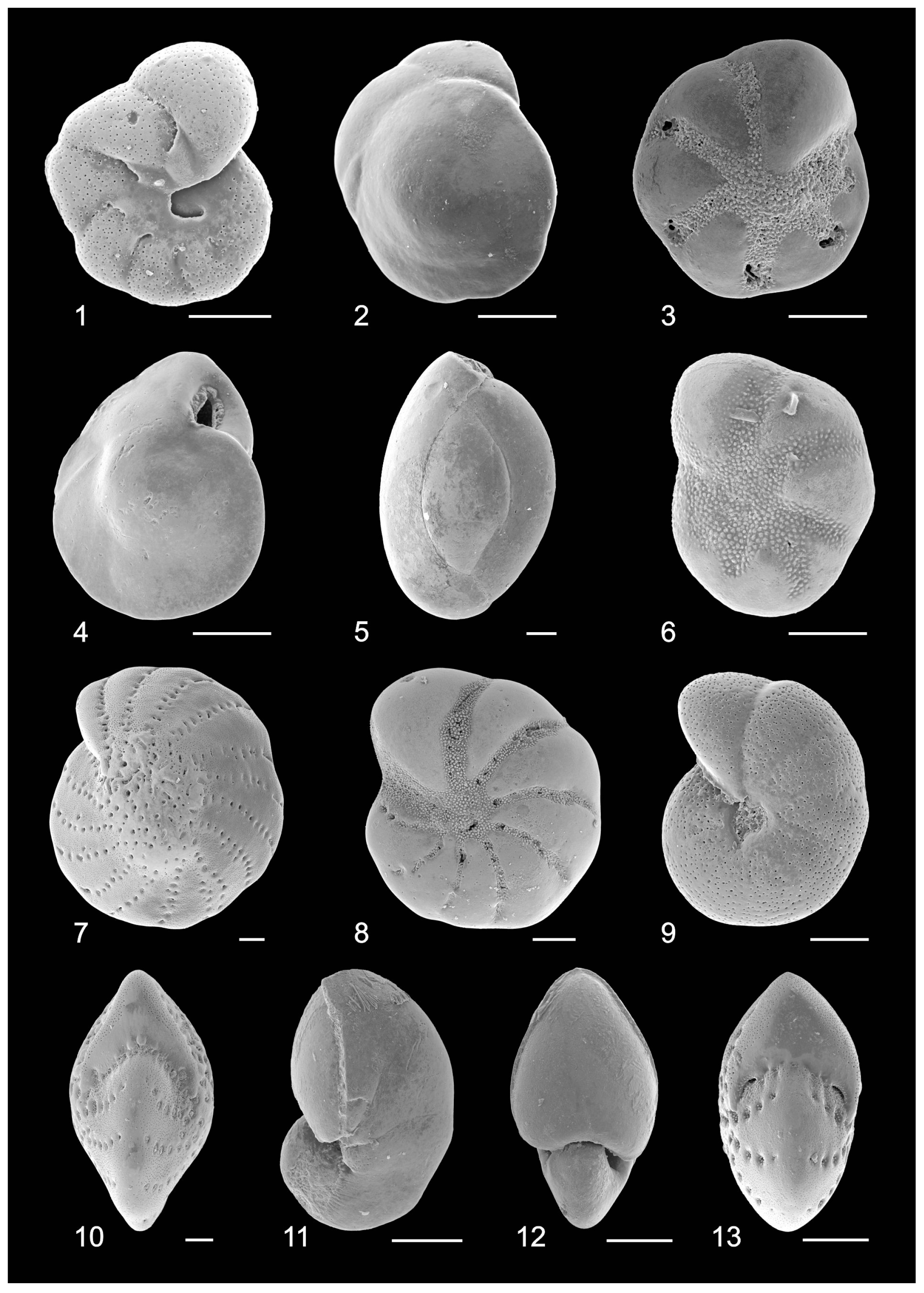
3.3. Benthic Foraminifers
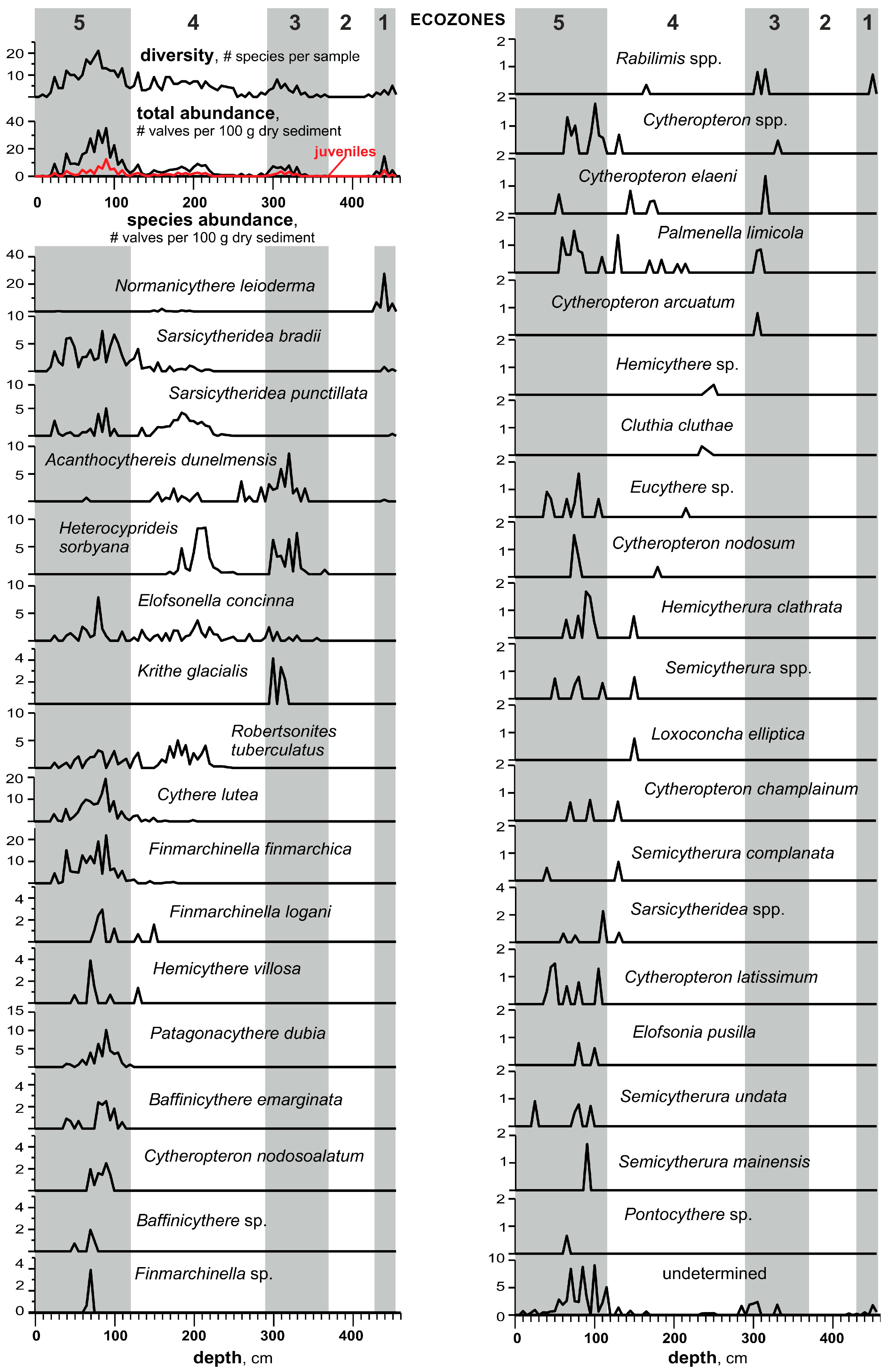
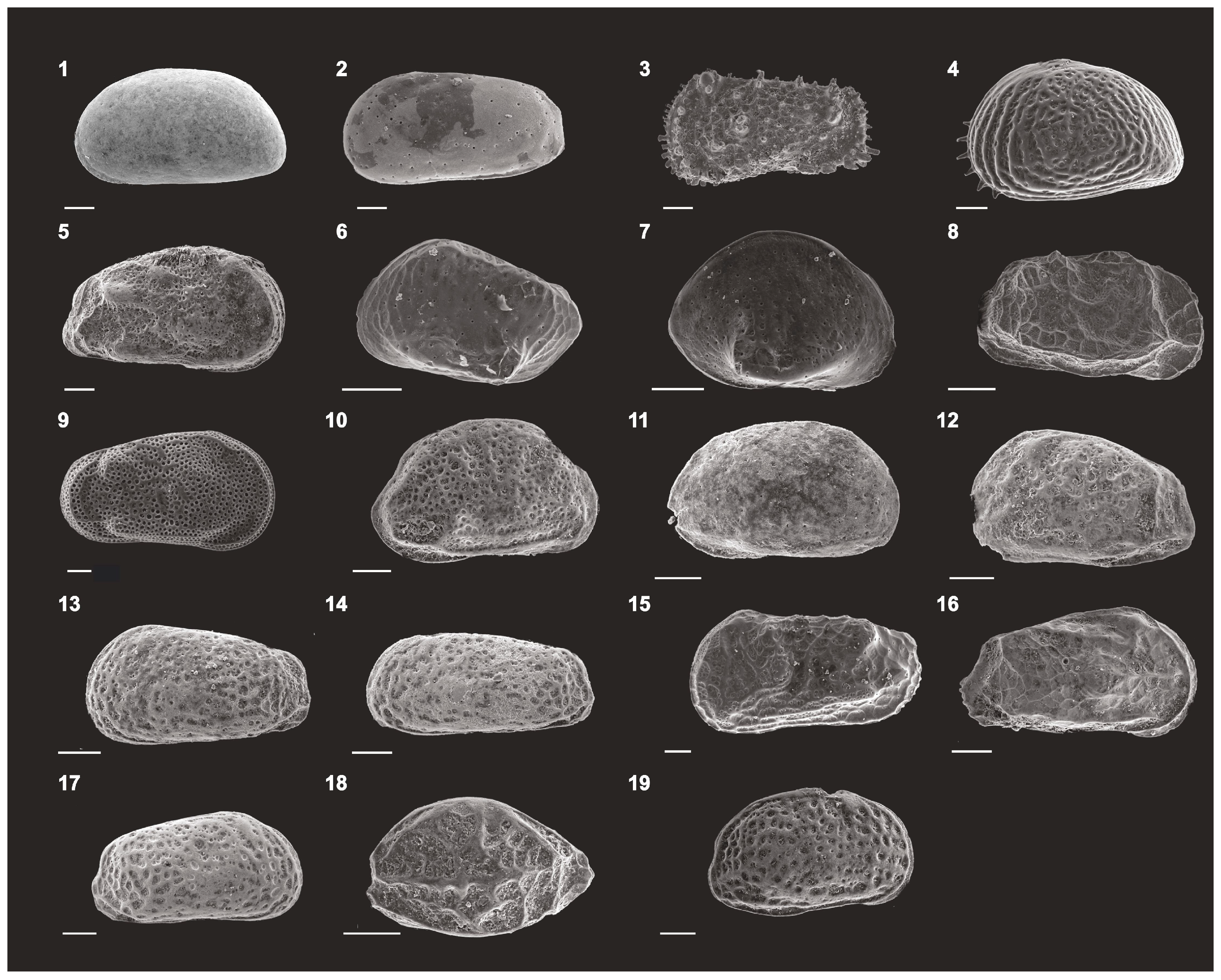
3.4. Ostracods
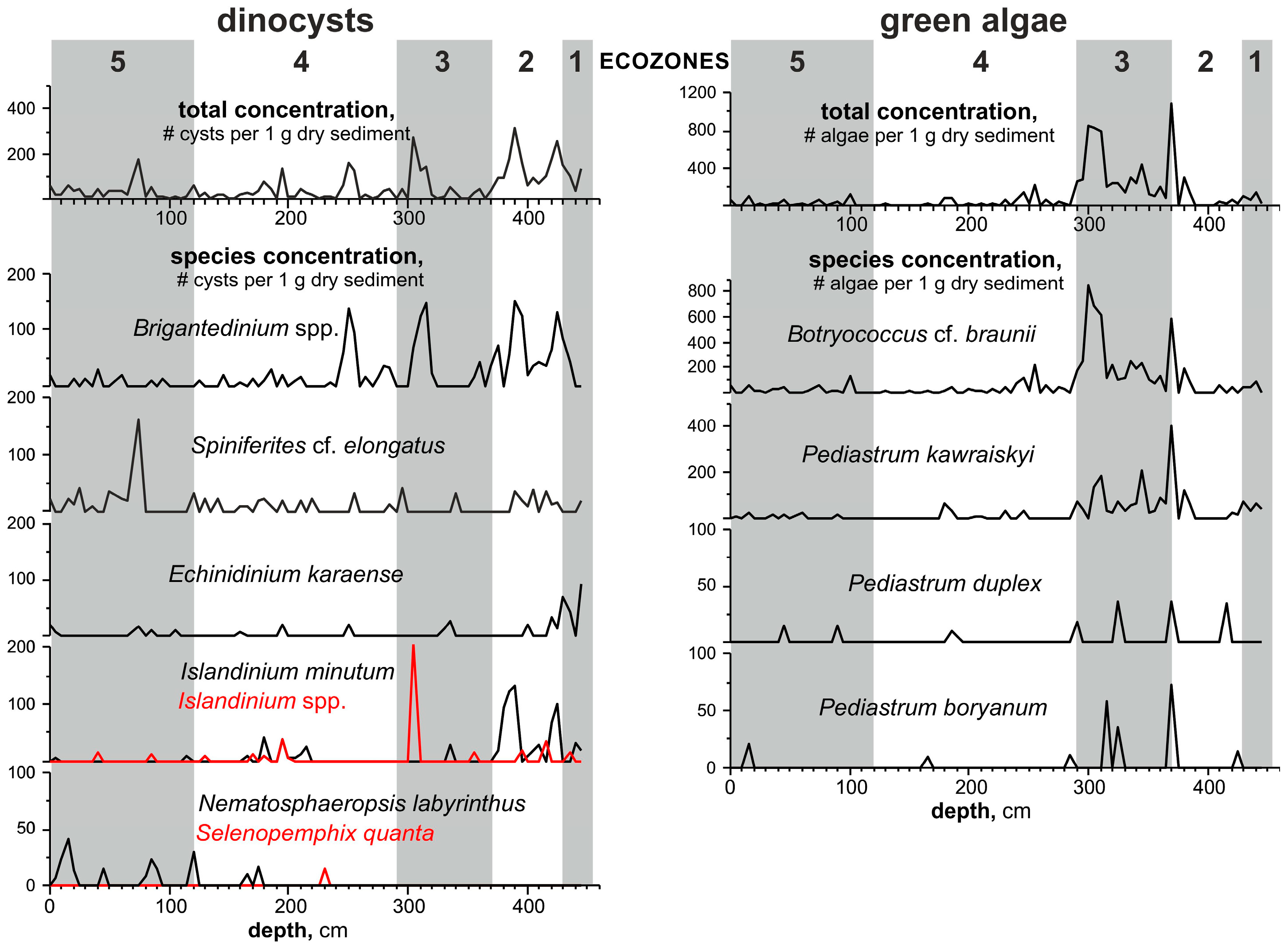
3.5. Aquatic Palynomorphs
4. Discussion
5. Conclusions
- Based on the high-resolution, multiproxy study of the reference Bychye-2 section, we reconstructed the course of the Boreal transgression in the northeastern White Sea region, which includes three major successive phases of paleoenvironmental changes: (1) the relatively deep coastal basin in the initial phases of flooding, with extensive sea ice cover and strong stratification (ecozones 1–2, >131–130.5 ka); (2) the deepest basin in the maximum phase of flooding (ecozone 3, 130.5–130.25 ka); and (3) the shallow-water basin with reduced seasonal sea ice cover (ecozones 4–5, 130.25–119.5 ka).
- The flooding by cold Arctic waters was rapid, and a relatively deep basin with high sedimentation rates existed for ~750 years, from >131 to 130.25 ka.
- According to the pollen zone comparison with other sites, the regression due to the regional isostatic rebound started around 130 ka.
- During the regressive stage, the occurrence of ostracods, which are characteristic of the Baltic Sea and presently absent in the Arctic seas, with the highest representation ~128–124 ka, allows us to assume that the connection between the White and Baltic seas existed for 6–7 thousand years, starting from >131 ka.
- The absence of the Atlantic water indicative species among benthic microfossils suggests that the subsurface Atlantic waters did not reach the studied region, which represented a shelf with water depths of less than 100 m, even during the period of its maximum deepening.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- CAPE-Last Interglacial Project Members. Last Interglacial Arctic warmth confirms polar amplification of climate change. Quat. Sci. Rev. 2006, 25, 1383–1400. [Google Scholar] [CrossRef]
- Dalton, A.S.; Gowan, E.J.; Mangerud, J.; Möller, P.; Lunkka, J.P.; Astakhov, V. Last Interglacial (MIS 5e) sea level proxies in the glaciated Northern Hemisphere. Earth Syst. Sci. Data 2022, 14, 1447–1492. [Google Scholar] [CrossRef]
- Lavrova, M.A. Correlation of the interglacial Boreal transgression of the northern USSR and Eemian transgression of the western Europe. Proc. Inst. Geol. Est. SSR Acad. Sci. 1961, VIII, 74–88. [Google Scholar]
- Gudina, V.I.; Evzerov, V.Y. Upper Pleistocene Stratigraphy and Foraminifers of the Kola Peninsula; Nauka: Novosibirsk, Russia, 1973; p. 146. [Google Scholar]
- Devyatova, E.I. Late Pleistocene Environment and Its Influence on Human Settlement in the Severnaya Dvina Basin and in Karelia; Karelia: Petrozavodsk, Russia, 1982; p. 156. [Google Scholar]
- Ikonen, L.; Ekman, I. Biostratigraphy of the Mikulino Interglacial Sediments in NW Russia: The Petrozavodsk Site and a Literature Review; Annales Academiae Scientiarum Fennicae A III Geologica-Geographica; Suomalainen Tiedeakatemia: Helsinki, Finland, 2001; Volume 161, p. 88. [Google Scholar]
- Houmark-Nielsen, M.; Demidov, I.; Funder, S.; Grøsfjeld, K.; Kjær, K.H.; Larsen, E.; Lavrova, N.; Lyså, A.; Nielsen, J.K. Early and Middle Valdaian glaciations, ice-dammed lakes and periglacial interstadials in northwest Russia: New evidence from the Pyoza River area. Glob. Planet. Chang. 2001, 31, 215–237. [Google Scholar] [CrossRef]
- Funder, S.; Demidov, I.; Yelovicheva, Y. Hydrography and mollusc faunas of the Baltic and the White Sea-North Sea seaway in the Eemian. Palaeogeogr. Palaeoclim. Palaeoecol. 2002, 184, 275–304. [Google Scholar] [CrossRef]
- Knudsen, K.L.; Jiang, H.; Gibbard, P.L.; Kristensen, P.; Seidenkrantz, M.-S.; Janczyk-Kopikowa, Z.; Marks, L. Environmental reconstructions of Eemian Stage interglacial marine records in the Lower Vistula area, southern Baltic Sea. Boreas 2012, 41, 209–234. [Google Scholar] [CrossRef]
- Molodkov, A.; Yevzerov, V. ESR/OSL ages of long-debated subtill fossil-bearing marine deposits from the southern Kola Peninsula: Stratigraphic implications. Boreas 2004, 33, 123–131. [Google Scholar]
- Grøsfjeld, K.; Funder, S.; Seidenkrantz, M.S.; Glaister, C. Last Interglacial marine environments in the White Sea region, northwestern Russia. Boreas 2006, 35, 493–520. [Google Scholar] [CrossRef]
- Lambeck, K.; Purcell, A.; Funder, S.; Kjær, K.H.; Larsen, E.; Möller, P. Constraints on the Late Saalian to early Middle Weichselian ice sheet of Eurasia from field data and rebound modelling. Boreas 2006, 35, 539–575. [Google Scholar] [CrossRef]
- Korsakova, O. Pleistocene marine deposits in the coastal areas of Kola Peninsula (Russia). Quat. Int. 2009, 206, 3–15. [Google Scholar] [CrossRef]
- Korsakova, O. Formal stratigraphy of the Neopleistocene (Middle and Upper/Late Pleistocene) in the Koola region, NW Russia. Quat. Int. 2019, 534, 42–59. [Google Scholar] [CrossRef]
- Miettinen, A.; Head, M.J.; Knudsen, K. Eemian sea-level highstand in the eastern Baltic Sea linked to long-duration White Sea connection. Quat. Sci. Rev. 2014, 86, 158–174. [Google Scholar] [CrossRef]
- Salonen, S.; Helmens, K.; Brendryen, J.; Kuosmanen, N.; Väliranta, M.; Goring, S.; Korpela, M.; Kylander, M.; Philip, A.; Plikk, A.; et al. Abrupt high-latitude climate events and decoupled seasonal trends during the Eemian. Nat. Commun. 2018, 9, 2851. [Google Scholar] [CrossRef] [PubMed]
- Zaretskaya, N.; Rybalko, A.; Repkina, T.; Shilova, O.; Krylov, A. Late Pleistocene in the southeastern White Sea and adjacent areas (Arkhangel’sk region, Russia): Stratigraphy and palaeoenvironments. Quat. Int. 2021, 605, 126–141. [Google Scholar] [CrossRef]
- Zaretskaya, N.E.; Taldenkova, E.E.; Ovsepyan, Y.S.; Ruchkin, M.V.; Baranov, D.V.; Rudenko, O.V.; Stepanova, A.Y. First data on the paleogeographical settings and chronology of the Last Interglacial on the Zimnii Coast of the White Sea. Dokl. Earth Sci. 2023, 512, 313–319. [Google Scholar] [CrossRef]
- Bauch, H.; Erlenkeuser, H.; Fahl, K.; Spielhagen, R.F.; Weinelt, M.S.; Andruleit, H.; Heinrich, R. Evidence for a steeper Eemian than Holocene sea surface temperature gradient between Arctic and sub-Arctic regions. Palaeogeogr. Palaeoclim. Palaeoecol. 1999, 145, 95–117. [Google Scholar] [CrossRef]
- Bauch, H.A.; Erlenkeuser, H.; Jung, S.J.A.; Thiede, J. Surface and deep water changes in the subpolar North Atlantic during termination II and the last interglaciation. Paleoceanography 2000, 15, 76–84. [Google Scholar] [CrossRef]
- Bauch, H.A.; Erlenkeuser, H. A “critical” climatic evaluation of last interglacial (MIS 5e) records from the Norwegian Sea. Polar Res. 2008, 27, 135–151. [Google Scholar] [CrossRef]
- Grichuk, V.P. Pleistocene History of Flora and Vegetation of the Russian Plain; Nauka: Moscow, Russia, 1989; p. 183. [Google Scholar]
- Zagwijn, W.H. An analysis of Eemian climate in western and central Europe. Quat. Sci. Rev. 1996, 15, 451–469. [Google Scholar] [CrossRef]
- Bauch, H.A. Interglacial climates and the Atlantic meridional overturning circulation: Is there and Arctic controversy? Quat. Sci. Rev. 2013, 63, 1–22. [Google Scholar] [CrossRef]
- Rasmussen, T.L.; Thomsen, E.; Kuijpers, A.; Wastegård, S. Late warming and early cooling of the sea surface in the Nordic seas during MIS 5e (Eemian Interglacial). Quat. Sci. Rev. 2003, 22, 809–821. [Google Scholar] [CrossRef]
- Nieuwenhove Van, N.; Bauch, H.A.; Eynaud, F.; Kandiano, E.; Cortijo, E.; Turon, J.L. Evidence for delayed poleward expansion of North Atlantic surface waters during the last interglacial (MIS 5e). Quat. Sci. Rev. 2011, 30, 934–946. [Google Scholar] [CrossRef]
- Zhuravleva, A.; Bauch, H.; Spielhagen, R. Atlantic water heat transfer through the Arctic gateway (Fram Strait) during the last interglacial. Glob. Planet. Chang. 2017, 157, 232–243. [Google Scholar] [CrossRef]
- Rudenko, O.V.; Taldenkova, E.E.; Bauch, H.A.; Ovsepyan, Y.S. Novye dannye k palinostratigrafii mikulinskikh (eemskikh) otlozhenii v nizov’yakh reki Peza (severo-vostok Belomorskogo regiona [New data on palynostratigraphy of the Mikulino (Eemian) deposits of the Lower Pyoza River (northeastern White Sea Region]. Probl. Arktiki I Antarkt. 2023, 69, 206–227. [Google Scholar] [CrossRef]
- Pokrovskaya, I.M. (Ed.) Paleopalynology; Nedra: Leningrad, Russia, 1966; Volume 1, pp. 34–39. [Google Scholar]
- NPP Database. Available online: https://non-pollen-palynomorphs.uni-goettingen.de/ (accessed on 26 September 2022).
- AlgaeBase; National University of Ireland: Galway, Ireland. 2020. Available online: https://www.algaebase.org (accessed on 17 August 2021).
- Komárek, J.; Jankovská, V. Review of the green algal genus Pediastrum: Implication for pollen-analytical research. Bibl. Phycol. 2001, 108, 127. [Google Scholar]
- Komárek, J.; Marvan, P. Morphological differences in natural populations of the genus Botryococcus (Chlorophyceae). Arch. Für Protistenkd. 1992, 141, 65–100. [Google Scholar] [CrossRef]
- Nieuwenhove van, N.; Pospelova, V.; Jordan, R.W. (Eds.) Taxonomy and Distribution of Modern Organic-Walled Dinoflagellate Cysts in Surface Sediments from the Northern Hemisphere: An Update of Rochon et al.; 1999. In Marine Micropaleontology; 2020; Volume 159, Available online: https://www.sciencedirect.com/journal/marine-micropaleontology/vol/159/suppl/C (accessed on 1 June 2022).
- Stockmarr, J. Tablets spores used in absolute pollen analysis. Pollen Spores 1971, 13, 616–621. [Google Scholar]
- Grimm, E.C. Computer Software, TILIA 2.0 Version b.4; Illinois State Museum, Research and Collections Center: Springfield, IL, USA, 1993.
- Grimm, E.C. TGView; Illinois State Museum, Research and Collections Center: Springfield, IL, USA, 2004. [Google Scholar]
- Müller, H. Pollenanalytische Untersuchungen und Jahresschichtenz ahlungen an der eem-zeitlichen Kieselgur von Bispingen/Luhe. Geol. Jahrb. 1974, A21, 19–169. [Google Scholar]
- McCulloch, M.T.; Esat, T. The coral record of last interglacial sea levels and sea surface temperatures. Chem. Geol. 2000, 169, 107–129. [Google Scholar] [CrossRef]
- Beets, D.J.; Beets, C.J.; Cleveringa, P. Age and climate of the late Saalian and early Eemian in the type-area, Amsterdam basin, The Netherlands. Quat. Sci. Rev. 2006, 25, 876–885. [Google Scholar] [CrossRef]
- Polyak, L.; Korsun, S.; Febo, L.; Stanovoy, V.; Khusid, T.; Hald, M.; Paulsen, B.E.; Lubinski, D.A. Benthic foraminiferal assemblages from the southern Kara Sea, a river-influenced arctic marine environment. J. Foraminifer. Res. 2002, 32, 252–273. [Google Scholar] [CrossRef]
- Steinsund, P.I. Benthic Foraminifera in the Surface Sediments of the Barents, Kara Seas: Modern and Late Quaternary Applications. Ph.D. Dissertation, University of Tromsø, Tromso, Norway, 1994. [Google Scholar]
- Klitgaard-Kristensen, D.; Sejrup, H.P.; Haflidason, H. Distribution of recent calcareous benthic foraminifera in the northern North Sea and relation to the environment. Polar Res. 2002, 21, 275–282. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006; p. 426. [Google Scholar]
- Stepanova, A.; Taldenkova, E.; Bauch, H.A. Recent Ostracoda of the Laptev Sea (Arctic Siberia): Taxonomic composition and some environmental implications. Mar. Micropal. 2003, 48, 23–48. [Google Scholar] [CrossRef]
- Stepanova, A.; Taldenkova, E.; Simstich, J.; Bauch, H.A. Comparison study of the modern ostracod associations in the Kara and Laptev seas: Ecological aspects. Mar. Micropal. 2007, 63, 111–142. [Google Scholar] [CrossRef]
- Frenzel, P.; Keyser, D.; Viehberg, A. An illustrated key and (palaeo)ecological primer for Postglacial to Recent Ostracoda (Crustacea) of the Baltic Sea. Boreas 2010, 39, 567–575. [Google Scholar] [CrossRef]
- Stepanova, A.; Obrochta, S.; Quintana Krupinski, N.B.; Hyttinen, O.; Kotilainen, A.; Andrén, T. Late Weichselian to Holocene history of the Baltic Sea as reflected in ostracod assemblages. Boreas 2019, 48, 761–778. [Google Scholar] [CrossRef]
- Hazel, J.E. Classification and distribution of the Recent Hemicytheridae and Trachyleberididae (Ostracoda) off northeastern North America. Geol. Surv. Prof. Pap. 1967, 564, 74. [Google Scholar]
- Hazel, J.E. Atlantic continental shelf and slope of the United States—Ostracode zoogeography in the southern Nova Scotian and northern Virginian faunal provinces. Geol. Surv. Prof. Pap. 1970, 529-E, 21. [Google Scholar]
- Brouwers, E.M. Systematic Paleontology of Quaternary Ostracode Assemblages from the Gulf of Alaska: Part 2. Families Trachyleberididae, Hemicytheridae, Loxoconchidae, Paracytheridae. U.S. Geol. Surv. Prof. Pap. 1993, 1531, 40. [Google Scholar]
- Brouwers, E.M.; Cronin, T.M.; Horne, D.J.; Lord, A.R. Recent shallow marine ostracods from high latitudes: Implications for late Pliocene and Quaternary palaeoclimatology. Boreas 2000, 29, 127–142. [Google Scholar] [CrossRef]
- Freiwald, A.; Mostafawi, N. Ostracods in a cold-temperate coastal environment, Western Troms, Northern Norway: Sedimentary aspects and assemblages. Facies 1998, 38, 255–273. [Google Scholar] [CrossRef]
- de Vernal, A.; Rochon, A.; Turon, J.-L.; Matthiessen, J. Organic-walled dinoflagellate cysts: Palynological tracers of seasurface conditions in middle to high latitude marine environments. Geobios 1997, 30, 905–920. [Google Scholar] [CrossRef]
- Marret, F.; Zonneveld, K. Atlas of modern organic-walled dinoflagellate cyst distribution. Rev. Palaeobot. Palynol. 2003, 125, 1–200. [Google Scholar] [CrossRef]
- Polyakova, E.; Dzhinoridze, R.; Novichkova, T.; Golovnina, E. Diatoms and palynomorphs in the White Sea sediments as indicators of ice and hydrological conditions. Oceanology 2003, 43, 144–158. [Google Scholar]
- Gemery, L.; Cronin, T.M.; Briggs, W.M.; Brouwers, E.M.; Schornikov, E.; Stepanova, A.; Wood, A.M.; Yasuhara, M. An Arctic and subarctic ostracode database: Biogeographic and paleoceanographic applications. Hydrobiologia 2015, 786, 59–95. [Google Scholar] [CrossRef]
- Ovsepyan, Y.S. Late Quaternary Foraminifers of the Laptev Sea and Reconstruction of Environmental Changes on the Basis of Paleoecological Analysis. Ph.D. Thesis, Geology and Mineralogy, Moscow, Russia, 2016; p. 240. [Google Scholar]
- Matthiessen, J.; Kunz-Pirrung, M.; Mudie, P.J. Freshwater chlorophycean algae in recent marine sediments of the Beaufort, Laptev and Kara Seas (Arctic Ocean) as indicators of river runoff. Int. J. Earth Sci. 2000, 89, 470–485. [Google Scholar] [CrossRef]
- Korsun, S.A.; Pogodina, I.A.; Tarasov, G.A.; Matishov, G.G. Foraminifers of the Barents Sea (Hydrobiology and Quaternary Paleoecology); KNTs RAN Publ.: Apatity, Russia, 1994; p. 136. [Google Scholar]
- Mackiewicz, A. Recent benthic Ostracoda from Hornsund, south Spitsbergen, Svalbard Archipelago. Pol. Polar Res. 2006, 27, 71–90. [Google Scholar]
- Szymańska, N.; Pawłowska, J.; Kucharska, M.; Kujawa, A.; Łącka, M.; Zajączkowski, M. Impact of shelf-transformed waters (STW) on foraminiferal assemblages in the outwash and glacial fjords of Adventfjorden and Hornsund, Svalbard. Oceanologia 2017, 59, 525–540. [Google Scholar] [CrossRef]
- Korsun, S.; Hald, M.; Golikova, E.; Yudina, A.; Kuznetsov, I.; Mikhailov, D.; Knyazeva, O. Intertidal foraminiferal fauna and the distribution of Elphidiidae at Chupa Inlet, western White Sea. Mar. Biol. Res. 2014, 10, 153–166. [Google Scholar] [CrossRef]
- Lubinski, D.J.; Polyak, L.; Forman, S.L. Freshwater and Atlantic water inflows to the deep northern Barents and Kara seas since ca 13 14C ka: Foraminifera and stable isotope. Quat. Sci. Rev. 2001, 20, 1851–1879. [Google Scholar] [CrossRef]
- Abrukina, R.E.; Krasil’nikova, G.N. Foraminifers of marine interglacial deposits of the Kola-Karelia region, their stratigraphic importance and evolutionary history. In Quaternary Geology and Geomorphology of the Eastern Baltic Shield. Proc; Institute of Geology: Karelian Branch, Russia, 1972; Volume 13, pp. 39–58. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taldenkova, E.; Ovsepyan, Y.; Rudenko, O.; Stepanova, A.; Bauch, H.A. Boreal (Eemian) Transgression in the Northeastern White Sea Region: Multiproxy Evidence from Bychye-2 Section. Quaternary 2024, 7, 3. https://doi.org/10.3390/quat7010003
Taldenkova E, Ovsepyan Y, Rudenko O, Stepanova A, Bauch HA. Boreal (Eemian) Transgression in the Northeastern White Sea Region: Multiproxy Evidence from Bychye-2 Section. Quaternary. 2024; 7(1):3. https://doi.org/10.3390/quat7010003
Chicago/Turabian StyleTaldenkova, Ekaterina, Yaroslav Ovsepyan, Olga Rudenko, Anna Stepanova, and Henning A. Bauch. 2024. "Boreal (Eemian) Transgression in the Northeastern White Sea Region: Multiproxy Evidence from Bychye-2 Section" Quaternary 7, no. 1: 3. https://doi.org/10.3390/quat7010003
APA StyleTaldenkova, E., Ovsepyan, Y., Rudenko, O., Stepanova, A., & Bauch, H. A. (2024). Boreal (Eemian) Transgression in the Northeastern White Sea Region: Multiproxy Evidence from Bychye-2 Section. Quaternary, 7(1), 3. https://doi.org/10.3390/quat7010003