
Improving Seed Germination by Cold Atmospheric Plasma
by
 , , ,
, , ,
Dayun Yan
1,* ,
,
Li Lin
1,
Michelle Zvansky
2,
Leat Kohanzadeh
2,
Shannon Taban
2,
Sabrina Chriqui
2 and
Michael Keidar
1,* 1
Department of Mechanical and Aerospace Engineering, George Washington University, Washington, DC 20052, USA
2
Department of Life Sciences, University of California, Los Angeles, CA 90095, USA
*
Authors to whom correspondence should be addressed.
Plasma 2022, 5(1), 98-110; https://doi.org/10.3390/plasma5010008
Submission received: 30 November 2021
/
Revised: 26 January 2022
/
Accepted: 7 February 2022
/
Published: 9 February 2022
(This article belongs to the Special Issue Dielectric Barrier Discharges)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Cold atmospheric plasma (CAP) is a tunable source of reactive species and other physical factors. It exerts luxuriant biochemical effects on diverse cells, including bacterial cells, mammalian cells, and plant cells. Over the past decade, CAP has shown promising application in modern agriculture. Here, we focused on the state of the art of plasma agriculture, particularly the improvement of seed germination rates. Typical plasma sources, underlying physical principles, and the chemical and cellular mechanism of plasma’s effect on plants seeds have been discussed in depth.
1. Cold Atmospheric Plasma (CAP): Sources and Physics
As a phase of matter, plasma can easily be found in many places, from household objects to deep space, such as fluorescent lamps, lightning, nuclear fusion, the ionosphere, the sun, and all other stars [1]. The first definition of plasma was introduced by Irving Langmuir, who described plasma as an ionized gas containing ions and electrons with a roughly equal charge as a whole. In other words, plasma is an ionized gas with a quasi-neutrality [2]. Ionizations can be achieved by particle collisions with energies higher than their ionization thresholds.
Heating and applying an electric field are two common ways of energizing collisions. The former strengthens the random motion of particles, while the latter accelerates naturally existed “seed” electrons to knock out more electrons from atoms. However, the electrons with lower mass are usually accelerated prior to ions at the beginning of applying an external electric field. The higher drift motion of these electrons causes a higher random motion (thermal motion, also called “Brownian motion”) during collisions, resulting in a different temperature distribution from other species [3]. Such plasma is called thermal nonequilibrium plasma. In contrast, the accelerated electrons in thermal equilibrium plasma deliver their kinetic energy to other particles during collisions until all of them share the same temperature profile [4].
Cold atmospheric plasma (CAP) is a thermal nonequilibrium plasma with all heavy particles at near room temperature (average temperature) because the collision frequency of electrons with other particles or species is not high enough to make the plasma reach thermal equilibrium before those free electrons are lost during the diffusion, the recombination, or the attachment with atoms [5].
CAP has been studied for many years, and multiple generators have been invented. The most popular hardware includes the Cold Atmospheric Plasma Jet (CAPJ), Dielectric Barrier Discharge (DBD) reactor, radiofrequency (RF) reactor, and surface DBD source [6]. To avoid arc discharge, either the anode or cathode will be covered by a layer of dielectric materials such as glass, quartz, ceramic, or others [7]. Noble gases such as helium (He) and argon (Ar) are used to trigger the stable and nonthermal glow-like discharge in CAP sources [8]. For the CAP jet, the samples do not involve the discharge process (Figure 1). Thus, the CAP jet is an indirect discharge source [9]. In contrast, the DBD source is a direct discharge source, in which samples such as seeds are involved in the discharge process [10]. Compared with the CAP jet, DBD plasma has a short length but can affect a much larger area simultaneously. However, the nature of the CAP jet makes it so that it has a high aspect ratio and can even reach 11 mm of length [11]. Thus, the DBD reactor is more suitable for a large area treatment compared to the CAP jet. Similarly, surface DBD is a unique DBD configuration, in which two electrodes have been integrated in one surface [12]. Such a design overcomes the sample areas’ three-dimensional space limitation. Even a large sample can also be treated by a surface DBD source. The discharge in a radiofrequency (RF) plasma reactor is based on the RF power input between two electrodes, rather than AC or DC power [10]. Typically, RF plasma generates in a chamber, where samples such as seeds are set in the gaps between electrodes.
Here, we shortly explained the physical mechanism of CAP formation using the CAP jet as an example. Specifically, taking the CAP jet as an example, a single electrode or a pair of high voltage (several kilovolts) electrodes are separated from a noble gas flow by dielectric material such as a glass tube. The electric field is thus applied on the noble gas, and results in an ionization wave (IW), which is defined as the propagation of an ionization rate peak [13]. The wave can thus propagate to the areas that the noble gas flow covers. The photon emission during the IW propagation luminates the ionization region called the ‘plasma bullet’ [14]. The bullets are usually observed in an Intensified Charge-Coupled Device (ICCD).
2. Reactive Species in CAP
The electron impact ionizations and excitations are not the only result of electron collisions during dicharge. An abundance of reactive species are also formed in CAP [15]. Due to the emission, many excited species can be identified from spectra, such as an Optical Emission Spectrum (OES). A typical OES of plasma in DBD was shown in Figure 2a, in which N2 or N2+ usually have the top peak. It is generally understood that the reactive species in CAP build the foundation for the interaction between CAP and cells, including bacterial, fungal, mammalian, and plant cells [16,17]. The electrons can be captured by the reactive species with strong negativity such as oxygen. The electrons can also recombine with ions, such as N2+, O2+, NO+, and He2+. Besides the electron collisions, other reactive species also collide with each other to form new species. For example, the penning ionization of metastable helium (He(23S)) and nitrogen collide to form N2+ and He. Some reactions will form important chemicals such as NO, N2O, NO2, as well as OH. These chemicals may play a key role during the interaction between CAP and cells. Some typical reactions formulae are shown at here as the examples. O + O2− => O3 + e, N + O2 => NO + O, N2(A3Σu+) + O2 => N2O + O, O2 + N2O+ => NO+ + NO2, N2(A3Σu+) + H2O => H + OH + N2, H2O+ + O2− => O2 + OH + H, N+ + H2O => NO+ + H2, etc. Note that the hydrogen element comes from the humidity components in air [18]. The reaction rate coefficients of these reactions are significantly affected by the electric field, electron temperature, gas bulk temperature, as well as the densities of reactants. The initial concentration of these species is also important for these reactions. The initial mole faction of He, N2, and O2 can be controlled by the flow rate of a carrying gas such as He and the geometry of the nozzle [19]. In addition, the humidity of air is another key but hardly controllable factor for the formation of reactive species in plasma [18].
Reactive species in the gas phase will have complex CAP-aqueous solution interactions when plasma touches solution (Figure 2b). The main reactive species in CAP can be divided into two categories: short-lived and long-lived reactive species. Short-lived reactive species, such as OH. and 1O2, may only affect the cells near cold plasma over a short-time scale [21]. In contrast, long-lived reactive species, such as H2O2, O3, NO, and NO2−, may be capable of affecting cells from the site even far from plasma through the diffusion in aqueous environments over a long-time scale [22]. In addition, many aqueous components, such as amino acids in medium, are modified during CAP treatment [23]. The pH in the CAP-treated solutions may also be changed; however, such a change is negligible because of the existence of buffering chemicals in solutions, such as phosphate monobasic (H2PO4−), phosphate dibasic (HPO42−), and bicarbonate (HCO3−) [24,25]. All these chemical changes may finally contribute to the biological impact of CAP on plant cells.
3. Plasma Agriculture, an Overview
The chemical effect of CAP may build the foundation for plasma agriculture [26,27]. The pathogen-based disease therapy, the resistance to abiotic stress, food sterilization during storage, as well as the improvement of germination rates are challenges in modern agriculture. CAP may help humans to overcome all these challenges through abundant and controllable reactive species generation [28,29,30].
Pathogen-based plant diseases are profoundly affecting crops worldwide [31]. Naturally, H2O2 production in plants can kill pathogens directly or induce defense genes to limit the infection [32,33,34]. CAP is a powerful source of ROS including H2O2 and others. A CAP jet was capable of curing the fungus-infected plant leaves and inhibiting the spread of infection (Figure 3) [35]. The leaves with the small (<2 mm) black spots infected with fungal cells could be completely recovered from the infection [35]. A CAP jet with a relatively high plasma density could completely kill the tomato pathogen C. fulvum and decrease the rotting rate of the infected tomato seeds within a short treatment time [31]. Aspergillus niger is a dangerous pathogen of date-palm fruits due to the production of fumonisin B2 (FB2) and ochratoxin A (OTA) [36]. A double atmospheric pressure argon cold plasma (DAPACP) jet system showed promising results in the inhibition of mycotoxin release by Aspergillus niger [36]. The pathogenic microorganisms of diverse sprout seeds play a crucial role in the pathogenesis of sprout-related outbreaks [37]. Three minutes of corona discharge plasma jet (CDPJ) treatment showed strong killing capability on aerobic bacteria, models and yeast, Bacillus cereus, Escherichia coli, Salmonella spp. in rapeseeds [37].
Furthermore, CAP is a safe tool for food decontamination. Diverse pathogens are large threats to the modern food industry. The abundant reactive species components in CAP make it a flexible and low-cost food processing technology for vegetables, fruits, poultry, as well as meats [38,39,40,41]. For example, a nitrogen CAP inactivated S. typhimurium on potato, strawberry, and lettuce [42]. CAP treatment is also desirable for in-package decontamination. CAP significantly eliminated the postprocess contamination while retaining the essential quality characteristics of strawberries [43]. The foods’ cross-contamination from persistent pathogen reservoirs is a risk in processing environments. CAP rapidly decontaminated the food surface with salmonella biofilms [44]. CAP treatment increased the tomato’s resistance to R. solanacearum with an efficacy of 25%, which might be due to the regulation of peroxidase, polyphenol oxidase, and phenylalanine ammonia lyase in tomato [45]. CAP also effectively inactivated the spoilage bacteria P. fluorescens and M. caseolyticus in packages [46]. Moreover, a short- pulsed CAP has shown its strong effect of eliminating foodborne pathogens salmonella and E. coli O157:H7 on almonds [47]. Compared with the dry air, nitrogen, as a carrying gas, resulted in a worse antimicrobial efficacy [47].
Additionally, CAP treatment can trigger the tolerance of plants to various abiotic and biotic stress. CAP was used to help oilseed rape seedlings resist the damage caused by the drought stress tomato seeds [45,48,49]. Under drought stress, the direct CAP treatment significantly improved seeding growth characteristics, including shoot and root dry weights, shoot and root lengths, and lateral root number in a drought-sensitive oilseed [48]. ROS and RNS in CAP may mainly contribute to the tolerance of plants to diverse stresses. Naturally, the H2O2 production in plants also induces the resistance to various stresses, including ultraviolet radiation (UV), salt stress, drought stress, light stress, metal stress, and high or low temperature [32]. H2O2 is involved in the abscisic acid (ABA)-induced stomatal opening and closing [32,34,50]. NO has been identified as an important signaling molecule in plant immune response [51]. NO is also involved in the ABA-induced stomatal closure in epidermal peels [52]. NO can also strongly counteract many ROS-mediated cytotoxic processes in plants [53]. Furthermore, NO possesses antioxidant properties and might act as a signal in activating the ROS-scavenging enzyme activities under abiotic stress, such as salt, drought, temperature, UV, and heavy metal stress [54,55,56]. Several NO donors, including sodium nitroprusside, S-nitroso-N-acetylpenicillamine, and a mixed solution of ascorbic acid and NaNO2, can prevent chlorophyll loss [57].
In addition to the direct CAP treatment on plants or seeds, aqueous solutions (particularly, water) have been used as the carriers for the CAP-originated reactive species through CAP treatment on these solutions [39,58,59,60,61,62,63,64,65,66,67,68,69]. CAP-stimulated solution (PSS) can be widely used in many circumstances independent of CAP devices (Figure 4). PSS was also named as CAP-activated solution (PAS) or water (PAW) in some references [46,70,71,72]. PSS contains an abundance of reactive species and has promising applications in agriculture. PSS has been shown to have a strong killing effect on bacteria [46,70,71,72] and viruses [41,73,74].
4. Seeds Germination
4.1. Germination Mechanism
Improving the seed germination rate is a main challenge in agriculture [75]. In agriculture, most crop seeds are dispersed in a dry and mature state during storage. When environmental conditions are favorable, seeds will initiate a complex process of germination [76,77]. Seed germination is a complex physiological process involving strictly regulated signaling pathways affected by a combination of environmental and endogenous factors [33,77]. The early germination phase culminates in the testa rupture, which is followed by the late germination phase and endosperm rupture [76]. Abscisic acid (ABA) and gibberellins acids (GAs), two widely investigated plant hormones, regulate the germination process [76]. ABA maintains dormancy, while GA promotes germination [77]. ROS and RNS may exert significant impacts on seed germination by interfering with ABA-related signaling, and stimulate GA-related signaling, which will modify redox balance and other downstream events in seeds to further trigger germination [33,77]. Specifically, OH. facilitates the loosening process of the cell wall, along with the protrusion and the elongation of radicles, which are the necessary steps for germination [33,78]. H2O2 stimulates the seed germination of cereal plants such as pea, apple, barley, wheat, and rice by facilitating the removal of ethanol-soluble compounds from the seeds with pericarp or by the oxidation or decomposition of the germination inhibitors in pericarp [33,79,80]. More importantly, H2O2 directly interferes with ABA and GA signaling pathways, breaks down ABA, modifies redox status, and induces protein carbonylation [33,77,80,81,82]. Several RNS, such as NO, NO2−, NO3−, NO2, or NO donor, such as sodium nitroprusside (SNP) also promote the release of seed dormancy and seed germination via affecting metabolism, oxidation state, as well as other signaling pathways, such as ABA/GAs-based pathways in seeds [55,77,81,82,83,84,85,86,87,88].
4.2. CAP’s Capability
As a controllable ROS/RNS source, CAP treatment has significantly improved the seed germination rate, speed, water uptake, seed vigor, as well as several key characteristics of seedling growth, including the shoot length, shoot dry weight, root length, and root dry weight of several seeds [89]. These effects were observed in chickpea seeds [75], cotton seeds [90], soybean seeds [91], brown rice seed [92], mung bean seeds [93], rapeseed [37], erythrina velutina seeds [94], sweet basil seeds [95], wheat seed [96,97], spinach seed [98], hemp cultivars [99], wheat seeds [100], tomato seeds [45,48,49], watermelon seeds [60], as well as cultivar seeds [48]. In short, CAP treatment is an alternative to conventional pre-germination treatments.
The improved germination rate and the increased growth of the hypocotyl and the radicle of mung beans were strongly dependent on the feed gases used to generate CAP and CAP dose (Figure 5) [93]. The CAP using air as feed gases showed a stronger effect on the seed germination rate than the CAP using O2, N2, and He did [93]. In addition, a moderate-intensity DBD treatment increased the seedling growth by 24.0%, 28.0%, and 35.5% after a 4 min of air plasma, N2 plasma, and Ar plasma treatment, respectively [96]. The O2 plasma treatment did not cause the observable enhancement on germination [96].
Though the key role of ROS/RNS in the seed germination process has been demonstrated in plant biology, the dominating factors for the improved germination rate and seedling growth have not been strictly demonstrated yet. The acidification and formation of H2O2 or nitrogen compounds may mainly contribute to these performances by promoting the leathering of seed chaps and the activation of catalase expression [93]. However, the enhanced germination rate may not be simply due to the chemical factors such as ROS or RNS. It was also found that a radiofrequency (5.28 MHz) electromagnetic radiation increased the germination rate of freshly harvested R. smirnowii seeds about 70% [101]. The electromagnetic effect in CAP may also exert an impact on seeds [102,103].
So far, the limited studies demonstrated that CAP treatment indeed triggered an observable cellular and molecular response in seeds [102,103,104]. An argon plasma promoted soybean seed germination and sprout growth by regulating the demethylation of adenosine triphosphate (ATP), target of rapamycin (TOR), and growth-regulating factor (GRF) [105]. The level of GA3 hormone, mRNA expression of amylolytic enzyme-related gene, chlorophyll, as well as the total polyphenols in spinach seedlings have been increased by a CAP treatment [98]. It was also found that the superoxide dismutase and catalase activities have been increased by 17.71% and 16.25% in CAP-treated oilseed, respectively [48]. The expression of heat shock proteins HSP101 and HSP70 in maize grains has been increased by a CAP treatment [106]. Thus, CAP treatment may improve oilseed rape drought tolerance by improving antioxidant enzyme activities, increasing osmotic-adjustment products, and reducing lipid peroxidation [48].
Seed germination is a complicated physiological process that begins with the seeds’ water absorption (imbibition) and ends with the radicle emerging [28,62,107]. It is known that the seeds’ kinetics and quantity water uptake efficacy are affected by their surface properties (e.g., morphology, composition, and structure) and surrounding environment, such as moisture content and temperature. The modification of CAP on the seed’s surface properties have been explored over the past decade. Some CAP-treated seeds, either experiencing a direct treatment or a soak of CAP-treated solution, showed better hydrophilicity and better water update efficacy [108]. For example, the seeds of a common bean (Phaseolus vulgaris L.) were treated by low-pressure oxygen plasma triggered by an inductively coupled radiofrequency (RF) discharge [109]. As shown in Figure 6, the CAP treatment drastically increased the wettability of bean seeds via changing their surface properties such as surface chemistry, surface roughness, and surface morphology. Promisingly, the improved wettability significantly increased the water uptake of these bean seeds. More attractively, these improvements were achieved by just a short treatment length (a few seconds). In short, CAP is scalable to large systems in the real agricultural applications.
PSS also improves the germination rate of seeds. The CAP-stimulated tap water, demineralized water, and liquid fertilizer have shown promising effects on increasing the germination rate and the stem elongation rate of lentils [39,59,60,61,62,63,64,65,66,67,68,69,110]. Reactive species in the aqueous solutions mainly contribute to the observed increased germination rate and stem growth (Figure 7) [59]. The endogenous production of NO radicals in seeds may be activated and facilitate the release of seeds from dormancy [59]. A cylindrical double DBD reactor was used for water activation and a plate-to-plate double DBD reactor was employed for the treatment of the seeds of radish, tomato, and sweet pepper [66]. The activated water has been acidified (pH ≈ 3) due to the absence of a buffer [66]. PSS showed a significant impact on germination rate as well as plant growth for all three types of seeds [66]. The stem length increased about 60% when the PSS treatment and the direct CAP treatment were combined [66].
Furthermore, CAP treatment also improves the germination rate by the inactivation of microorganisms grown on the surface of seeds [84,85,99]. The control of seedborne rice seedling diseases in the seed beds is key to avoid epidemics in rice nurseries and paddies [111]. Two seedborne rice seedling diseases, bakanae disease caused by the fungal pathogen Fusarium fujikuroi, and the bacterial seedling blight caused by Burkholderia plantarii, can be effectively inhibited by the CAP treatment on infected rice seeds [71]. A surface DBD can effectively decrease microbial contamination without reducing the viability of the seeds of sweet basil (Ocimum basilicum L.) [95]. The low-pressure radiofrequency (RF) oxygen and air plasmas also showed a strong anti-bacteria effect on the seed-borne bacteria [72].
5. Conclusions
Over the past decade, CAP has shown its attractive potential as a novel and safe modality in agriculture. The complex reactive chemical nature of CAP builds the foundation of plasma agriculture, including the pathogen-based plant disease therapy, the resistance to abiotic stress, food sterilization, and the improvement of seed germination rates. Reactive species either kill microorganisms on seeds’ surface or trigger cellular and molecular pathways in seeds to facilitate the germination process and further development such as seedling. CAP also shows a strong potential to modify the surface properties of seeds, causing an improvement in the water uptake and later development. The CAP-stimulated (treated, activated) solutions recently attracted widespread attention due to their convenience during storage and the potential for massive use in farms. Moreover, a CAP source can consume electricity and use air to generate reactive species via careful design. Thus, plasma agriculture is an environmentally friendly, green technology.
To date, the demonstration of these practical perspectives is mainly limited to the lab level. It is far from an extensive application of CAP technology in modern agriculture. Though we know many positive outcomes of plasma agriculture, such as improved seed germination, we do not know whether CAP treatment could affect the quality of these important crops, such as rice production, and potential side effects. Furthermore, it is entirely unknown whether plasma agriculture could contribute to some significant challenges in agriculture, such as the growth of crops in saline-alkali land. Will the CAP-treated seeds give the final crop a solid resistance to diseases? In terms of efficacy and economy, will plasma agriculture be competitive compared to existing breeding technologies such as fertilizers? Many challenges are waiting to be solved before the final realization of plasma agriculture.
Author Contributions
Conceptualization, D.Y. and M.K.; methodology, writing—original draft preparation, review and editing, D.Y., L.L., M.Z., L.K., S.T., S.C. and M.K.; funding acquisition, M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation grant, grant number 1747760.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Samukawa, S.; Hori, M.; Rauf, S.; Tachibana, K.; Bruggeman, P.; Kroesen, G.; Whitehead, J.C.; Murphy, A.B.; Gutsol, A.F.; Starikovskaia, S.; et al. The 2012 Plasma Roadmap. J. Phys. D Appl. Phys. 2012, 45, 253001. [Google Scholar] [CrossRef]
- Attri, P.; Arora, B.; Choi, E.H. Utility of Plasma: A New Road from Physics to Chemistry. RSC Adv. 2013, 3, 12540–12567. [Google Scholar] [CrossRef]
- Laroussi, M.; Akan, T. Arc-Free Atmospheric Pressure Cold Plasma Jets: A Review. Plasma Process. Polym. 2007, 4, 777–788. [Google Scholar] [CrossRef]
- Fridman, A.; Chirokov, A.; Gutsol, A. Non-Thermal Atmospheric Pressure Discharges. J. Phys. D Appl. Phys. 2005, 38, R1. [Google Scholar] [CrossRef]
- Lin, L.; Keidar, M. A Map of Control for Cold Atmospheric Plasma Jets: From Physical Mechanisms to Optimizations. Appl. Phys. Rev. 2021, 8, 011306. [Google Scholar] [CrossRef]
- Sakudo, A.; Yagyu, Y.; Onodera, T. Disinfection and Sterilization Using Plasma Technology: Fundamentals and Future Perspectives for Biological Applications. Int. J. Mol. Sci. 2019, 20, 5216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Sherman, J.H.; Keidar, M. Cold Atmospheric Plasma, a Novel Promising Anti-Cancer Treatment Modality. Oncotarget 2017, 8, 15977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keidar, M. Plasma for Cancer Treatment. Plasma Sources Sci. Technol. 2015, 24, 033001. [Google Scholar] [CrossRef]
- Pawłat, J.; Starek, A.; Sujak, A.; Terebun, P.; Kwiatkowski, M.; Budzeń, M.; Andrejko, D. Effects of atmospheric pressure plasma jet operating with DBD on Lavatera thuringiaca L. seeds’ germination. PLoS ONE 2018, 13, 0194349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Ramírez, A.; López-Santos, C.; Cantos, M.; García, J.L.; Molina, R.; Cotrino, J.; Espinós, J.P.; González-Elipe, A.R. Surface Chemistry and Germination Improvement of Quinoa Seeds Subjected to Plasma Activation. Sci. Rep. 2017, 7, 5924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Jiang, Z.; Xiong, Q.; Tang, Z.; Hu, X.; Pan, Y. An 11 Cm Long Atmospheric Pressure Cold Plasma Plume for Applications of Plasma Medicine. Appl. Phys. Lett. 2008, 92, 2006–2008. [Google Scholar]
- Waskow, A.; Howling, A.; Furno, I. Advantages and Limitations of Surface Analysis Techniques on Plasma-Treated Arabidopsis Thaliana Seeds. Front. Mater. Sci. 2021, 8, 123. [Google Scholar] [CrossRef]
- Lu, X.P.; Ostrikov, K.K. Guided Ionization Waves: The Physics of Repeatability. Appl. Phys. Rev. 2018, 5, 031102. [Google Scholar] [CrossRef]
- Lu, X.P.; Naidis, G.V.; Laroussi, M.; Ostrikov, K. Guided Ionization Waves: Theory and Experiments. Phys. Rep. 2014, 540, 123–166. [Google Scholar] [CrossRef]
- Yan, D.; Horkowitz, A.; Keidar, M. On the Selective Killing of Cold Atmospheric Plasma Cancer Treatment: Status and Beyond. Plasma Process Polym. 2021, 18, 2100020. [Google Scholar] [CrossRef]
- Graves, D.B. The Emerging Role of Reactive Oxygen and Nitrogen Species in Redox Biology and Some Implications for Plasma Applications to Medicine and Biology. J. Phys. D Appl. Phys. 2012, 45, 263001–263042. [Google Scholar] [CrossRef]
- Motyka-Pomagruk, A.; Dzimitrowicz, A.; Orlowski, J.; Babinska, W.; Terefinko, D.; Rychlowski, M.; Prusinski, M.; Pohl, P.; Lojkowska, E.; Jamroz, P.; et al. Implementation of a Non-Thermal Atmospheric Pressure Plasma for Eradication of Plant Pathogens from a Surface of Economically Important Seeds. Int. J. Mol. Sci. 2021, 22, 9256. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Lyu, Y.; Trink, B.; Canady, J.; Keidar, M. Cold Atmospheric Helium Plasma Jet in Humid Air Environment. J. Appl. Phys. 2019, 125, 153301. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Y.; Lin, L.; Gjika, E.; Lee, T.; Keidar, M. Mathematical Modeling and Control for Cancer Treatment with Cold Atmospheric Plasma Jet. J. Phys. D Appl. Phys. 2019, 52, 185202. [Google Scholar] [CrossRef]
- Perinban, S.; Orsat, V.; Raghavan, V. Nonthermal Plasma-Liquid Interactions in Food Processing: A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1985–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malyavko, A.; Yan, D.; Wang, Q.; Klein, A.L.; Patel, K.C. Cold Atmospheric Plasma Cancer Treatment, Direct versus Indirect Approaches. Mater. Adv. 2020, 1, 1494–1505. [Google Scholar] [CrossRef]
- Tanaka, H.; Bekeschus, S.; Yan, D.; Hori, M.; Keidar, M.; Laroussi, M. Plasma-Treated Solutions (PTS) in Cancer Therapy. Cancers 2021, 13, 1737. [Google Scholar] [CrossRef] [PubMed]
- Takai, E.; Kitamura, T.; Kuwabara, J.; Ikawa, S.; Yoshizawa, S.; Shiraki, K.; Kawasaki, H.; Arakawa, R.; Kitano, K. Chemical Modification of Amino Acids by Atmospheric-Pressure Cold Plasma in Aqueous Solution. J. Phys. D Appl. Phys. 2014, 47, 285403. [Google Scholar] [CrossRef]
- Panngom, K.; Baik, K.Y.; Nam, M.K.; Han, J.H.; Rhim, H.; Choi, E.H. Preferential Killing of Human Lung Cancer Cell Lines with Mitochondrial Dysfunction by Nonthermal Dielectric Barrier Discharge Plasma. Cell Death Dis. 2013, 4, e642. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, N.; Kumar, N.; Kim, C.H.; Kaushik, N.K.; Choi, E.H. Dielectric Barrier Discharge Plasma Efficiently Delivers an Apoptotic Response in Human Monocytic Lymphoma. Plasma Process. Polym. 2014, 11, 1175–1187. [Google Scholar] [CrossRef]
- Adhikari, B.; Pangomm, K.; Veerana, M.; Mitra, S.; Park, G. Plant Disease Control by Non-Thermal Atmospheric-Pressure Plasma. Front. Plant Sci. 2020, 11, 77. [Google Scholar] [CrossRef]
- Holubová, L.; Kyzek, S.; Ďurovcová, I.; Fabová, J.; Horváthová, E.; Ševčovičová, A.; Gálová, E. Non-Thermal Plasma—A New Green Priming Agent for Plants? Int. J. Mol. Sci. 2020, 21, 9466. [Google Scholar] [CrossRef] [PubMed]
- Starič, P.; Vogel-Mikuš, K.; Mozetič, M.; Junkar, I. Effects of Nonthermal Plasma on Morphology, Genetics and Physiology of Seeds: A Review. Plants 2020, 9, 1736. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, P.; Veerana, M.; Lim, J.S.; Mumtaz, S.; Shrestha, B.; Kaushik, N.K.; Park, G.; Choi, E.H. Low-Temperature Plasma-Assisted Nitrogen Fixation for Corn Plant Growth and Development. Int. J. Mol. Sci. 2021, 22, 5360. [Google Scholar] [CrossRef] [PubMed]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M. Plasma Agriculture from Laboratory to Farm: A Review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, D.; Song, Y.; Zhou, R.; Niu, J. Inactivation of the Tomato Pathogen Cladosporium Fulvum by an Atmospheric-Pressure Cold Plasma Jet. Plasma Process. Polym. 2014, 11, 1028–1036. [Google Scholar] [CrossRef]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen Peroxide in Plants: A Versatile Molecule of the Reactive Oxygen Species Network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef]
- Bailly, C. Active Oxygen Species and Antioxidants in Seed Biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, D.; Zhou, R.; Song, Y.; Sun, Y.; Zhang, Q.; Niu, J.; Fan, H.; Yang, S.Z. Atmospheric Cold Plasma Jet for Plant Disease Treatment. Appl. Phys. Lett. 2014, 104, 043702. [Google Scholar] [CrossRef]
- Ouf, S.A.; Basher, A.H.; Mohamed, A.A.H. Inhibitory Effect of Double Atmospheric Pressure Argon Cold Plasma on Spores and Mycotoxin Production of Aspergillus Niger Contaminating Date Palm Fruits. J. Sci. Food Agric. 2015, 95, 3204–3210. [Google Scholar] [CrossRef] [PubMed]
- Puligundla, P.; Kim, J.; Mok, C. Effect of Corona Discharge Plasma Jet Treatment on Decontamination and Sprouting of Rapeseed (Brassica Napus L.) Seeds. Food Control 2017, 71, 376–382. [Google Scholar] [CrossRef]
- Bourke, P.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Keener, K. The Potential of Cold Plasma for Safe and Sustainable Food Production. Trends Biotechnol. 2018, 36, 615–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirumdas, R.; Kothakota, A.; Annapure, U.; Siliveru, K.; Blundell, R.; Gatt, R.; Valdramidis, V.P. Plasma Activated Water (PAW): Chemistry, Physico-Chemical Properties, Applications in Food and Agriculture. Trends Food Sci. Technol. 2018, 77, 21–31. [Google Scholar] [CrossRef]
- Hertwig, C.; Meneses, N.; Mathys, A. Cold Atmospheric Pressure Plasma and Low Energy Electron Beam as Alternative Nonthermal Decontamination Technologies for Dry Food Surfaces: A Review. Trends Food Sci. Technol. 2018, 77, 131–142. [Google Scholar] [CrossRef]
- Filipić, A.; Primc, G.; Zaplotnik, R.; Mehle, N.; Gutierrez-Aguirre, I.; Ravnikar, M.; Mozetič, M.; Žel, J.; Dobnik, D. Cold Atmospheric Plasma as a Novel Method for Inactivation of Potato Virus Y in Water Samples. Food Environ. Virol. 2019, 11, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Fernández, A.; Noriega, E.; Thompson, A. Inactivation of Salmonella Enterica Serovar Typhimurium on Fresh Produce by Cold Atmospheric Gas Plasma Technology. Food Microbiol. 2013, 33, 24–29. [Google Scholar] [CrossRef]
- Misra, N.N.; Moiseev, T.; Patil, S.; Pankaj, S.K.; Bourke, P.; Mosnier, J.P.; Keener, K.M.; Cullen, P.J. Cold Plasma in Modified Atmospheres for Post-Harvest Treatment of Strawberries. Food Bioproc. Technol. 2014, 7, 3045–3054. [Google Scholar] [CrossRef]
- Niemira, B.A.; Boyd, G.; Sites, J. Cold Plasma Rapid Decontamination of Food Contact Surfaces Contaminated with Salmonella Biofilms. J. Food Sci. 2014, 79, M917–M922. [Google Scholar] [CrossRef]
- Jiang, J.; Lu, Y.; Li, J.; Li, L.; He, X.; Shao, H.; Dong, Y. Effect of Seed Treatment by Cold Plasma on the Resistance of Tomato to Ralstonia Solanacearum (Bacterial Wilt). PLoS ONE 2014, 9, e97753. [Google Scholar]
- Wang, J.; Zhuang, H.; Zhang, J. Inactivation of Spoilage Bacteria in Package by Dielectric Barrier Discharge Atmospheric Cold Plasma Treatment Time Effects. Food Bioproc. Technol. 2016, 9, 1648–1652. [Google Scholar] [CrossRef]
- Niemira, B.A. Cold Plasma Reduction of Salmonella and Escherichia Coli O157: H7 on Almonds Using Ambient Pressure Gases. J. Food Sci. 2012, 77, 171–175. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Shen, M.; Zhang, C.; Dong, Y. Cold Plasma Treatment Enhances Oilseed Rape Seed Germination under Drought Stress. Sci. Rep. 2015, 5, 13033. [Google Scholar] [CrossRef]
- Bormashenko, E.; Shapira, Y.; Grynyov, R.; Whyman, G.; Bormashenko, Y.; Drori, E. Interaction of Cold Radiofrequency Plasma with Seeds of Beans (Phaseolus Vulgaris). J. Exp. Bot. 2015, 66, 4013–4021. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.; Cheung, M.K.; Bright, J.; Henson, D.; Hancock, J.T.; Neill, S.J. ABA, Hydrogen Peroxide and Nitric Oxide Signalling in Stomatal Guard Cells. J. Exp. Bot. 2004, 55, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Lum, H.K.; Butt, Y.K.C.; Lo, S.C.L. Hydrogen Peroxide Induces a Rapid Production of Nitric Oxide in Mung Bean (Phaseolus Aureus). Nitric Oxide Biol. Chem. 2002, 6, 205–213. [Google Scholar] [CrossRef]
- Wilson, I.D.; Neill, S.J.; Hancock, J.T. Nitric Oxide Synthesis and Signalling in Plants. Plant Cell Environ. 2008, 31, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Beligni Maria, V.; Lamattina, L. Nitric Oxide Counteracts Reactive Oxygen Species Actions in Plant Tissues. Planta 1999, 208, 337–344. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Role of Nitric Oxide in Tolerance of Plants to Abiotic Stress. Protoplasma 2011, 248, 447–455. [Google Scholar] [CrossRef]
- Kopyra, M.; Gwóźdź, E.A. Nitric Oxide Stimulates Seed Germination and Counteracts the Inhibitory Effect of Heavy Metals and Salinity on Root Growth of Lupinus Luteus. Plant Physiol. Biochem. 2003, 41, 1011–1017. [Google Scholar] [CrossRef]
- Shi, S.; Wang, G.; Wang, Y.; Zhang, L.; Zhang, L. Protective Effect of Nitric Oxide against Oxidative Stress under Ultraviolet-B Radiation. Nitric Oxide: Biol. Chem. 2005, 13, 1–9. [Google Scholar] [CrossRef]
- Beligni, M.V.; Lamattina, L. Nitric Oxide Protects against Cellular Damage Produced by Methylviologen Herbicides in Potato Plants. Nitric Oxide Biol. Chem. 1999, 3, 199–208. [Google Scholar] [CrossRef]
- Lu, P.; Boehm, D.; Cullen, P.; Bourke, P. Controlled Cytotoxicity of Plasma Treated Water Formulated by Open-Air Hybrid Mode Discharge. Appl. Phys. Lett. 2017, 110, 264102. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Rousseau, A.; Dufour, T. Promoting Lentil Germination and Stem Growth by Plasma Activated Tap Water, Demineralized Water and Liquid Fertilizer. RSC Adv. 2017, 7, 31244–31251. [Google Scholar] [CrossRef] [Green Version]
- Lotfy, K. Effects of Cold Atmospheric Plasma Jet Treatment on the Seed Germination and Enhancement Growth of Watermelon. Open J. Appl. Sci. 2017, 7, 705–719. [Google Scholar] [CrossRef] [Green Version]
- Kostoláni, D.; Ndiffo Yemeli, G.B.; Švubová, R.; Kyzek, S.; Machala, Z. Physiological Responses of Young Pea and Barley Seedlings to Plasma-Activated Water. Plants 2021, 10, 1750. [Google Scholar] [CrossRef]
- Adhikari, B.; Adhikari, M.; Ghimire, B.; Park, G.; Choi, E.H. Cold Atmospheric Plasma-Activated Water Irrigation Induces Defense Hormone and Gene Expression in Tomato Seedlings. Sci. Rep. 2019, 9, 16080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Than, H.A.Q.; Pham, T.H.; Nguyen, D.K.V.; Pham, T.H.; Khacef, A. Non-Thermal Plasma Activated Water for Increasing Germination and Plant Growth of Lactuca Sativa L. Plasma Chem. Plasma Process. 2021; in press. [Google Scholar]
- Fan, L.; Liu, X.; Ma, Y.; Xiang, Q. Effects of Plasma-Activated Water Treatment on Seed Germination and Growth of Mung Bean Sprouts. J. Taibah Univ. Sci. 2020, 14, 823–830. [Google Scholar] [CrossRef]
- Chen, D.; Chen, P.; Cheng, Y.; Peng, P.; Liu, J.; Ma, Y.; Liu, Y.; Ruan, R. Deoxynivalenol Decontamination in Raw and Germinating Barley Treated by Plasma-Activated Water and Intense Pulsed Light. Food Bioproc. Technol. 2019, 12, 246–254. [Google Scholar] [CrossRef]
- Sivachandiran, L.; Khacef, A. Enhanced Seed Germination and Plant Growth by Atmospheric Pressure Cold Air Plasma: Combined Effect of Seed and Water Treatment. RSC Adv. 2017, 7, 1822–1832. [Google Scholar] [CrossRef] [Green Version]
- El Shaer, M.; El Welily, H.; Zaki, A.; Arafa, H.; Elsebaei, A.; Eldaly, M.; Mobasher, M. Germination of Wheat Seeds Exposed to Cold Atmospheric Plasma in Dry and Wet Plasma-Activated Water and Mist. Plasma Med. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Terebun, P.; Kwiatkowski, M.; Hensel, K.; Kopacki, M.; Pawłat, J. Influence of Plasma Activated Water Generated in a Gliding Arc Discharge Reactor on Germination of Beetroot and Carrot Seeds. Appl. Sci. 2021, 11, 6164. [Google Scholar] [CrossRef]
- Herianto, S.; Shih, M.K.; Lin, C.M.; Hung, Y.C.; Hsieh, C.W.; Wu, J.S.; Chen, M.H.; Chen, H.L.; Hou, C.Y. The Effects of Glazing with Plasma-Activated Water Generated by a Piezoelectric Direct Discharge Plasma System on Whiteleg Shrimp (Litopenaeus Vannamei). LWT 2022, 154, 112547. [Google Scholar] [CrossRef]
- Ikawa, S.; Kitano, K.; Hamaguchi, S. Effects of PH on Bacterial Inactivation in Aqueous Solutions Due to Low-Temperature Atmospheric Pressure Plasma Application. Plasma Process. Polym. 2010, 7, 33–42. [Google Scholar] [CrossRef]
- Ochi, A.; Konishi, H.; Ando, S.; Sato, K.; Yokoyama, K.; Tsushima, S.; Yoshida, S.; Morikawa, T.; Kaneko, T.; Takahashi, H. Management of Bakanae and Bacterial Seedling Blight Diseases in Nurseries by Irradiating Rice Seeds with Atmospheric Plasma. Plant Pathol. 2017, 66, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Ono, R.; Uchida, S.; Hayashi, N.; Kosaka, R.; Soeda, Y. Inactivation of Bacteria on Plant Seed Surface by Low-Pressure RF Plasma Using a Vibrating Stirring Device. Vaccum 2016, 136, 214–220. [Google Scholar] [CrossRef]
- Aboubakr, H.A.; Gangal, U.; Youssef, M.M.; Goyal, S.M.; Bruggeman, P.J. Inactivation of Virus in Solution by Cold Atmospheric Pressure Plasma: Identification of Chemical Inactivation Pathways. J. Phys. D Appl. Phys. 2016, 49, 204001. [Google Scholar] [CrossRef]
- Filipić, A.; Dobnik, D.; Tušek Žnidarič, M.; Žegura, B.; Štern, A.; Primc, G.; Mozetič, M.; Ravnikar, M.; Žel, J.; Gutierrez Aguirre, I. Inactivation of Pepper Mild Mottle Virus in Water by Cold Atmospheric Plasma. Front. Microbiol. 2021, 12, 618209. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Li, Y.F.; Klämpfl, T.G.; Shimizu, T.; Jeon, J.; Morfill, G.E.; Zimmermann, J.L. Inactivation of Surface-Borne Microorganisms and Increased Germination of Seed Specimen by Cold Atmospheric Plasma. Food Bioproc. Technol. 2014, 7, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the Mark: Early Seed Germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular Aspects of Seed Dormancy. Annu. Rev. Plant. Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, K.; Linkies, A.; Vreeburg, R.A.M.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In Vivo Cell Wall Loosening by Hydroxyl Radicals during Cress Seed Germination and Elongation Growth. Plant Physiol. 2009, 150, 1855–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, K.; Iwabuchi, M. A Mechanism for Promoting the Germination of Zinnia Elegans Seeds by Hydrogen Peroxide. Plant Cell Physiol. 2001, 42, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Barba-Espin, G.; Diaz-Vivancos, P.; Clemente-Moreno, M.J.; Albacete, A.; Faize, L.; Faize, M.; Pérez-Alfocea, F.; Hernández, J.A. Interaction between Hydrogen Peroxide and Plant Hormones during Germination and the Early Growth of Pea Seedlings. Plant Cell Environ. 2010, 33, 981–994. [Google Scholar] [CrossRef] [PubMed]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative Signaling in Seed Germination and Dormancy. Plant Signal Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, M.; Garcia, Q. Reactive Oxygen Species and Seed Germination. Biologia 2013, 68, 351–357. [Google Scholar] [CrossRef]
- Zhang, H.; Shen, W.-B.; Xu, L.-L. Effects of Nitric Oxide on the Germination of Wheat Seeds and Its Reactive Oxygen Species Metabolisms under Osmotic Stress. Acta Bot. Sin. 2003, 45, 901–905. [Google Scholar]
- Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Liu, W.; Jing, Q.; Cao, W. Exogenous Nitric Oxide Improves Seed Germination in Wheat against Mitochondrial Oxidative Damage Induced by High Salinity. Environ. Exp. Bot. 2009, 67, 222–227. [Google Scholar] [CrossRef]
- Hu, K.D.; Hu, L.Y.; Li, Y.H.; Zhang, F.Q.; Zhang, H. Protective Roles of Nitric Oxide on Germination and Antioxidant Metabolism in Wheat Seeds under Copper Stress. Plant Growth Regul. 2007, 53, 173–183. [Google Scholar] [CrossRef]
- Sarath, G.; Bethke, P.C.; Jones, R.; Baird, L.M.; Hou, G.; Mitchell, R.B. Nitric Oxide Accelerates Seed Germination in Warm-Season Grasses. Planta 2006, 223, 1154–1164. [Google Scholar] [CrossRef] [Green Version]
- Bethke, P.C.; Libourel, I.G.L.; Jones, R.L. Nitric Oxide Reduces Seed Dormancy in Arabidopsis. J. Exp. Bot. 2006, 57, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, S.B.; Taylorson, R.B. Promotion of Seed Germination by Nitrate, Nitrite, Hydroxylamine, and Ammonium Salts. Plant Physiol. 1974, 54, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, B.; Adhikari, M.; Park, G. The Effects of Plasma on Plant Growth, Development, and Sustainability. Appl. Sci. 2020, 10, 6045. [Google Scholar] [CrossRef]
- de Groot, G.J.J.B.; Hundt, A.; Murphy, A.B.; Bange, M.P.; Mai-Prochnow, A. Cold Plasma Treatment for Cotton Seed Germination Improvement. Sci. Rep. 2018, 8, 14372. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Jiang, J.; Li, J.; Shen, M.; He, X.; Shao, H.; Dong, Y. Effects of Cold Plasma Treatment on Seed Germination and Seedling Growth of Soybean. Sci. Rep. 2014, 4, 5859. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Chia, H.; Kuo, Y.; Lun, C.; Yi, S.; Sheng, Y. An Improved Process for High Nutrition of Germinated Brown Rice Production: Low-Pressure Plasma. Food Chem. 2016, 191, 120–127. [Google Scholar]
- Zhou, R.; Zhou, R.; Zhang, X.; Zhuang, J.; Yang, S.; Bazaka, K.; Ostrikov, K. Effects of Atmospheric-Pressure N2, He, Air, and O2 Microplasmas on Mung Bean Seed Germination and Seedling Growth. Sci. Rep. 2016, 6, 32603. [Google Scholar] [CrossRef] [Green Version]
- Junior, C.A.; de Oliveira Vitoriano, J.; da Silva, D.L.S.; de Lima Farias, M.; de Lima Dantas, N.B. Water Uptake Mechanism and Germination of Erythrina Velutina Seeds Treated with Atmospheric Plasma. Sci. Rep. 2016, 6, 33722. [Google Scholar] [CrossRef] [PubMed]
- Ambrico, P.F.; Šimek, M.; Morano, M.; de Miccolis Angelini, R.M.; Minafra, A.; Trotti, P.; Ambrico, M.; Prukner, V.; Faretra, F. Reduction of Microbial Contamination and Improvement of Germination of Sweet Basil (Ocimum Basilicum L.) Seeds via Surface Dielectric Barrier Discharge. J. Phys. D Appl. Phys. 2017, 50, 305401. [Google Scholar] [CrossRef]
- Meng, Y.; Qu, G.; Wang, T.; Sun, Q.; Liang, D.; Hu, S. Enhancement of Germination and Seedling Growth of Wheat Seed Using Dielectric Barrier Discharge Plasma with Various Gas Sources. Plasma Chem. Plasma Process. 2017, 37, 1105–1119. [Google Scholar] [CrossRef]
- Lotfy, K.; Al-Harbi, N.A.; Abd El-Raheem, H. Cold Atmospheric Pressure Nitrogen Plasma Jet for Enhancement Germination of Wheat Seeds. Plasma Chem. Plasma Process. 2019, 39, 897–912. [Google Scholar] [CrossRef]
- Ji, S.H.; Choi, K.H.; Pengkit, A.; Im, J.S.; Kim, J.S.; Kim, Y.H.; Park, Y.; Hong, E.J.; Jung, S.K.; Choi, E.H.; et al. Effects of High Voltage Nanosecond Pulsed Plasma and Micro DBD Plasma on Seed Germination, Growth Development and Physiological Activities in Spinach. Arch. Biochem. Biophys. 2016, 605, 117–128. [Google Scholar] [CrossRef]
- Sera, B.; Sery, M.; Gavril, B.; Gajdova, I. Seed Germination and Early Growth Responses to Seed Pre-Treatment by Non-Thermal Plasma in Hemp Cultivars (Cannabis Sativa L.). Plasma Chem. Plasma Process. 2017, 37, 207–221. [Google Scholar] [CrossRef]
- Zahoranová, A.; Henselová, M.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Medvecká, V.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on the Wheat Seedlings Vigor and on the Inactivation of Microorganisms on the Seeds Surface. Plasma Chem. Plasma Process. 2016, 36, 397–414. [Google Scholar] [CrossRef]
- Mildaziene, V.; Pauzaite, G.; Malakauskiene, A.; Zukiene, R.; Nauciene, Z.; Filatova, I.; Azharonok, V.; Lyushkevich, V. Response of Perennial Woody Plants to Seed Treatment by Electromagnetic Field and Low-Temperature Plasma. Bioelectromagnetics 2016, 37, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Mildažienė, V.; Aleknavičiūtė, V.; Žūkienė, R.; Paužaitė, G.; Naučienė, Z.; Filatova, I.; Lyushkevich, V.; Haimi, P.; Tamošiūnė, I.; Baniulis, D. Treatment of Common Sunflower (Helianthus Annus L.) Seeds with Radio-Frequency Electromagnetic Field and Cold Plasma Induces Changes in Seed Phytohormone Balance, Seedling Development and Leaf Protein Expression. Sci. Rep. 2019, 9, 6437. [Google Scholar] [CrossRef]
- Ghaemi, M.; Majd, A.; Iranbakhsh, A. Transcriptional Responses Following Seed Priming with Cold Plasma and Electromagnetic Field in Salvia Nemorosa L. J. Theor. Appl. Phys. 2020, 14, 323–328. [Google Scholar] [CrossRef]
- Pet’ková, M.; Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, L.; Ševčovičová, A.; Gálová, E. The Effects of Cold Atmospheric Pressure Plasma on Germination Parameters, Enzyme Activities and Induction of Dna Damage in Barley. Int. J. Mol. Sci. 2021, 22, 2833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Jo, J.O.; Huynh, D.L.; Mongre, R.K.; Ghosh, M.; Singh, A.K.; Lee, S.B.; Mok, Y.S.; Hyuk, P.; Jeong, D.K. Growth-Inducing Effects of Argon Plasma on Soybean Sprouts via the Regulation of Demethylation Levels of Energy Metabolism-Related Genes. Sci. Rep. 2017, 7, 41917. [Google Scholar] [CrossRef]
- Holubová, Ľ.; Švubová, R.; Slováková, Ľ.; Bokor, B.; Kročková, V.C.; Renčko, J.; Uhrin, F.; Medvecká, V.; Zahoranová, A.; Gálová, E. Cold Atmospheric Pressure Plasma Treatment of Maize Grains—Induction of Growth, Enzyme Activities and Heat Shock Proteins. Int. J. Mol. Sci. 2021, 22, 8509. [Google Scholar] [CrossRef]
- Attri, P.; Koga, K.; Okumura, T.; Shiratani, M. Impact of Atmospheric Pressure Plasma Treated Seeds on Germination, Morphology, Gene Expression and Biochemical Responses. Jpn. J. Appl. Phys. 2021, 60, 040502. [Google Scholar] [CrossRef]
- Holc, M.; Mozetič, M.; Recek, N.; Primc, G.; Vesel, A.; Zaplotnik, R.; Gselman, P. Wettability increase in plasma-treated agricultural seeds and its relation to germination improvement. Agronomy 2021, 11, 1467. [Google Scholar] [CrossRef]
- Recek, N.; Holc, M.; Vesel, A.; Zaplotnik, R.; Gselman, P.; Mozetič, M.; Primc, G. Germination of Phaseolus Vulgaris l. Seeds after a Short Treatment with a Powerful Rf Plasma. Int. J. Mol. Sci. 2021, 22, 6672. [Google Scholar] [CrossRef]
- Šerá, B.; Šerý, M.; Zahoranová, A.; Tomeková, J. Germination Improvement of Three Pine Species (Pinus) After Diffuse Coplanar Surface Barrier Discharge Plasma Treatment. Plasma Chem. Plasma Process. 2021, 41, 211–226. [Google Scholar] [CrossRef]
- Siddique, S.S.; Hardy, G.E.S.J.; Bayliss, K.L. Cold Plasma: A Potential New Method to Manage Postharvest Diseases Caused by Fungal Plant Pathogens. Plant Pathol. 2018, 67, 1011–1021. [Google Scholar] [CrossRef]
Figure 1.
Some CAP sources used in plasma agriculture. (a) A CAP jet and its discharge photo [9]. (b) A DBD reactor and structure [10]. (c) A radiofrequency (RF) reactor and structure [10]. (d) Two surface DBD sources with honeycomb electrodes and strip electrodes [12].

Figure 2.
Reactive species in gas phase and liquid phase. (a) An emission spectrum of plasma in the gas phase of DBD in the range from 200–400 nm [17]. (b) General picture of CAP-aqueous solution interactions. Reprinted with permission from Ref. [20]. Copyright 2019 John Wiley and Sons.
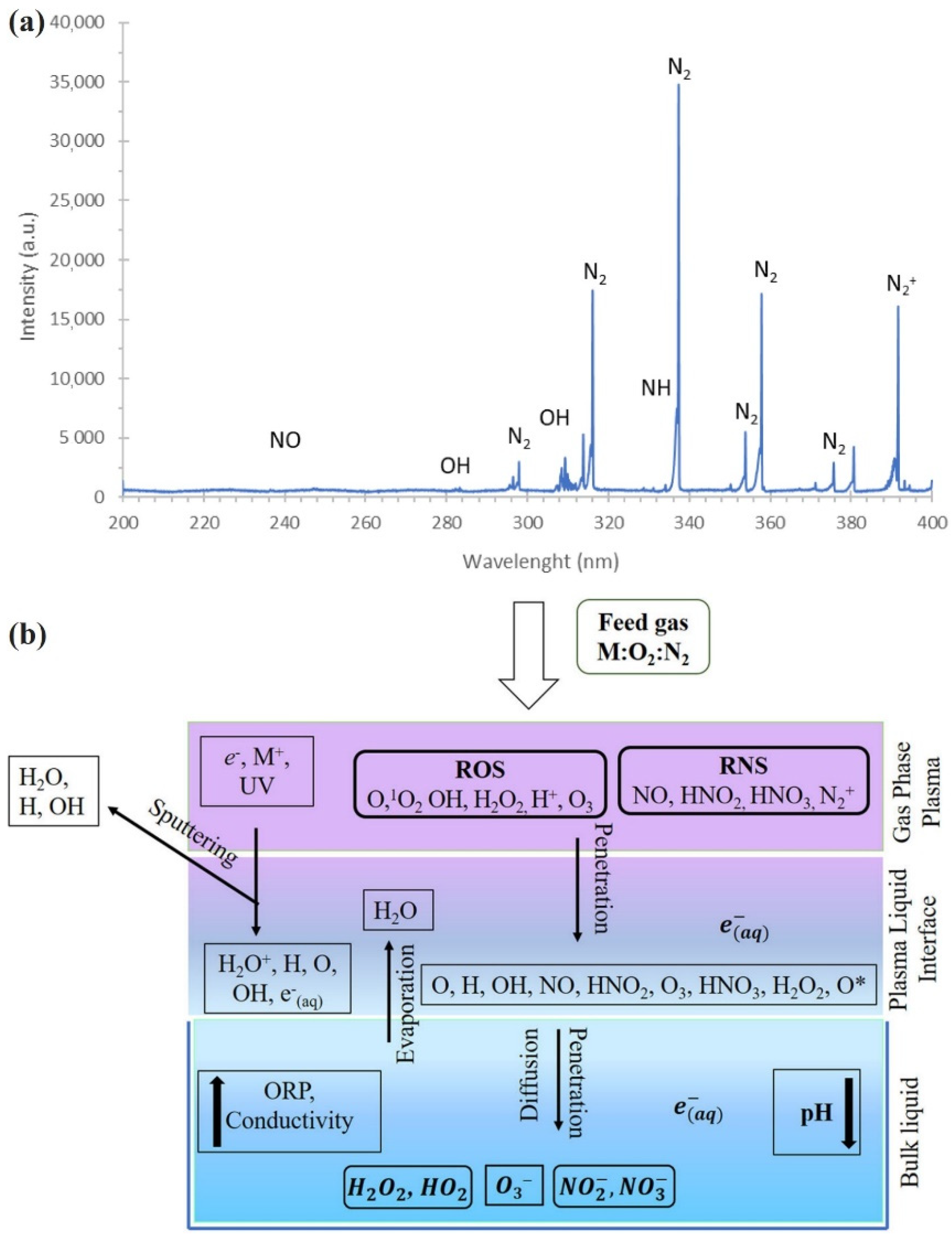
Figure 3.
Plasma treatment drastically eliminated the initial symptoms of a fungus-infected leaf and advanced symptoms after 1–3 weeks plasma treatment. Reprinted with permission from Ref. [35]. Copyright 2014 AIP Publishing.
Figure 3.
Plasma treatment drastically eliminated the initial symptoms of a fungus-infected leaf and advanced symptoms after 1–3 weeks plasma treatment. Reprinted with permission from Ref. [35]. Copyright 2014 AIP Publishing.

Figure 4.
The application of CAP-stimulated solution (PSS) in agriculture. Here, PSS was made by the vertical treatment on solutions. Subsequently, the PSS containing 3-nitro-L-tyrosine can be stably stored in the container at 2 °C to 8 °C for at least a week. PSS can be used to kill the pathogens grown on the plant. In addition, the soak of seeds in PSS will improve the seed germination rate.
Figure 4.
The application of CAP-stimulated solution (PSS) in agriculture. Here, PSS was made by the vertical treatment on solutions. Subsequently, the PSS containing 3-nitro-L-tyrosine can be stably stored in the container at 2 °C to 8 °C for at least a week. PSS can be used to kill the pathogens grown on the plant. In addition, the soak of seeds in PSS will improve the seed germination rate.
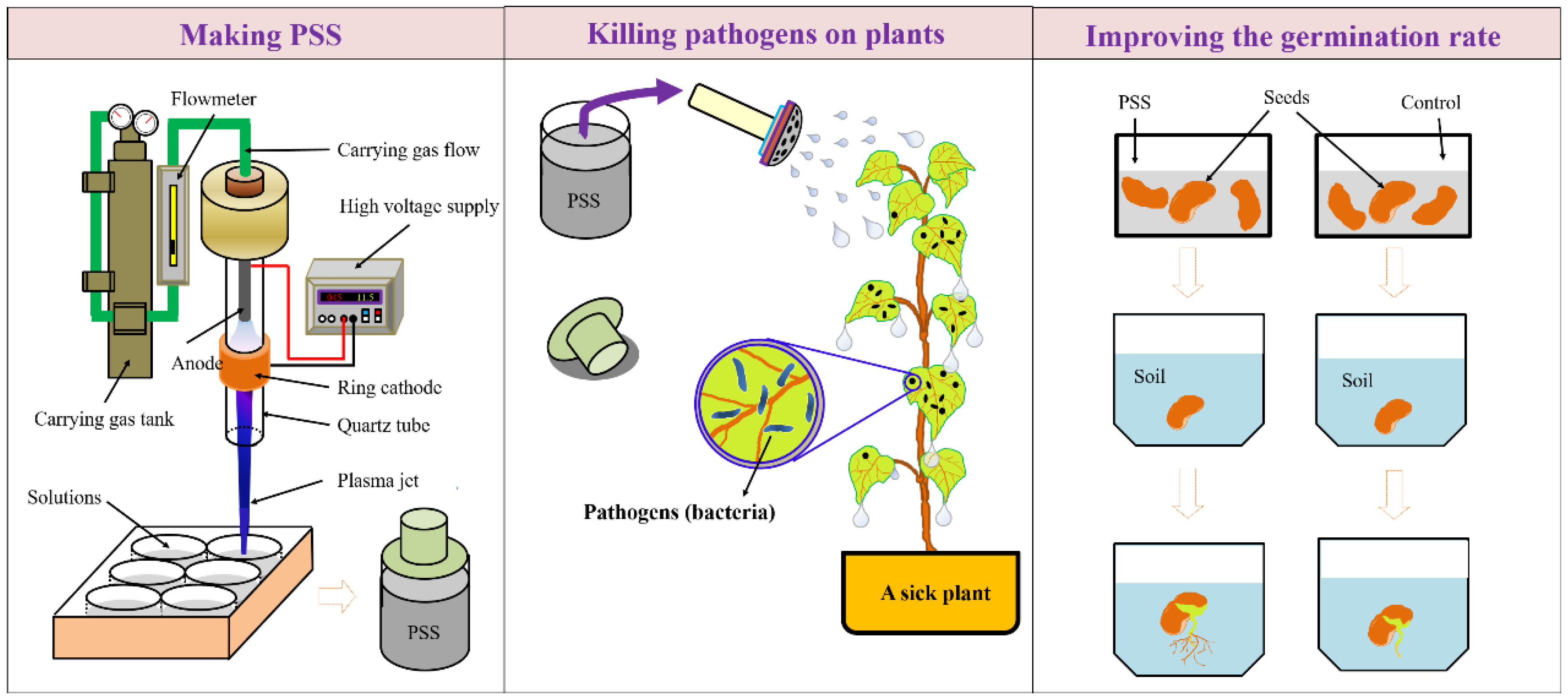
Figure 5.
Air plasma drastically improved the germination rate of mung bean seeds. (a) 0 h, (b) 9 h, (c) 12 h, (d) 24 h, and (e) the germination percentage of mung bean seeds treated with He, N2, air, or O2 plasma as a function of incubation time [93].
Figure 5.
Air plasma drastically improved the germination rate of mung bean seeds. (a) 0 h, (b) 9 h, (c) 12 h, (d) 24 h, and (e) the germination percentage of mung bean seeds treated with He, N2, air, or O2 plasma as a function of incubation time [93].
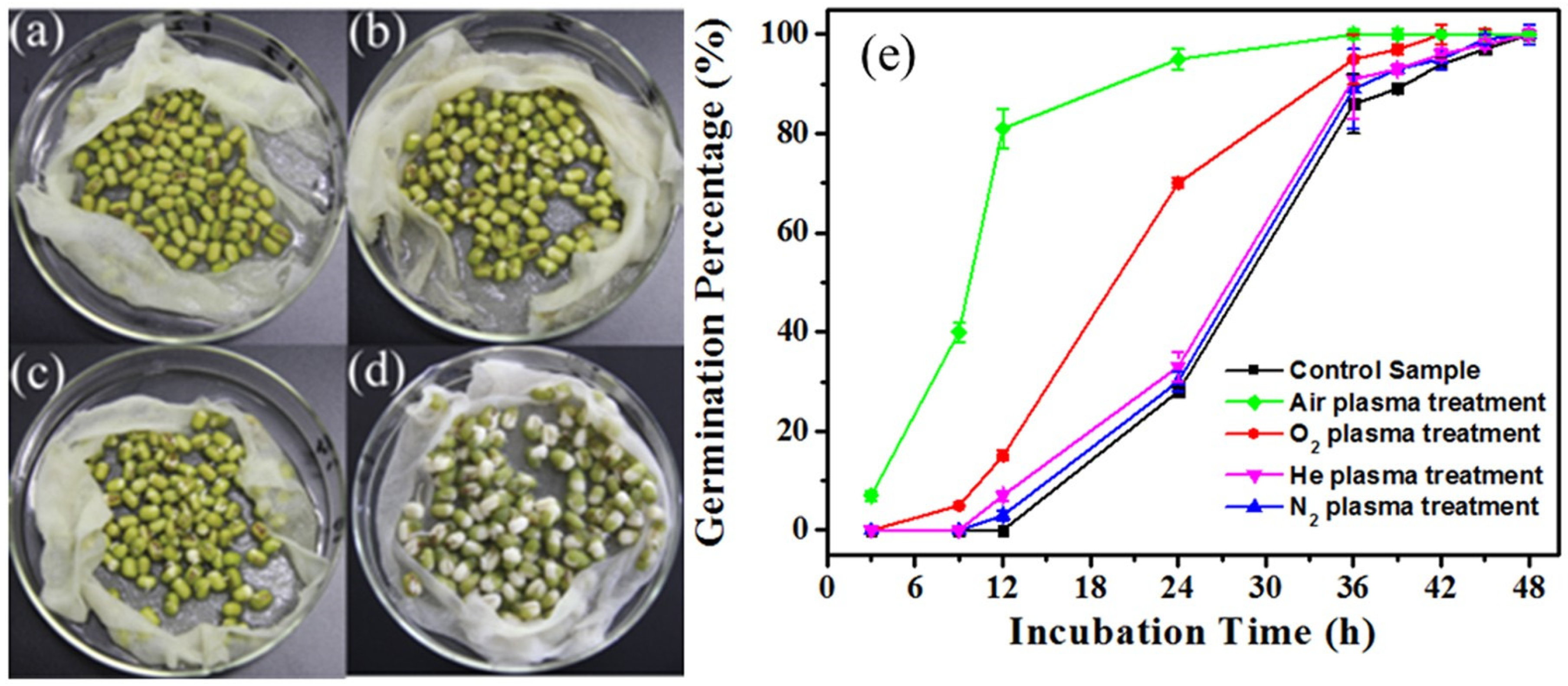
Figure 6.
CAP treatment improved the water uptake of bean seeds. (a) Visual bean seed radicle length after 4 days of incubation. (b) Water droplets on untreated and treated bean seed. SEM images of seeds’ surface were shown below. (c) The effect of plasma treatment on water contact angle on seed’s surfaces. (d) The effect of plasma treatment on water uptake of seeds [109].
Figure 6.
CAP treatment improved the water uptake of bean seeds. (a) Visual bean seed radicle length after 4 days of incubation. (b) Water droplets on untreated and treated bean seed. SEM images of seeds’ surface were shown below. (c) The effect of plasma treatment on water contact angle on seed’s surfaces. (d) The effect of plasma treatment on water uptake of seeds [109].
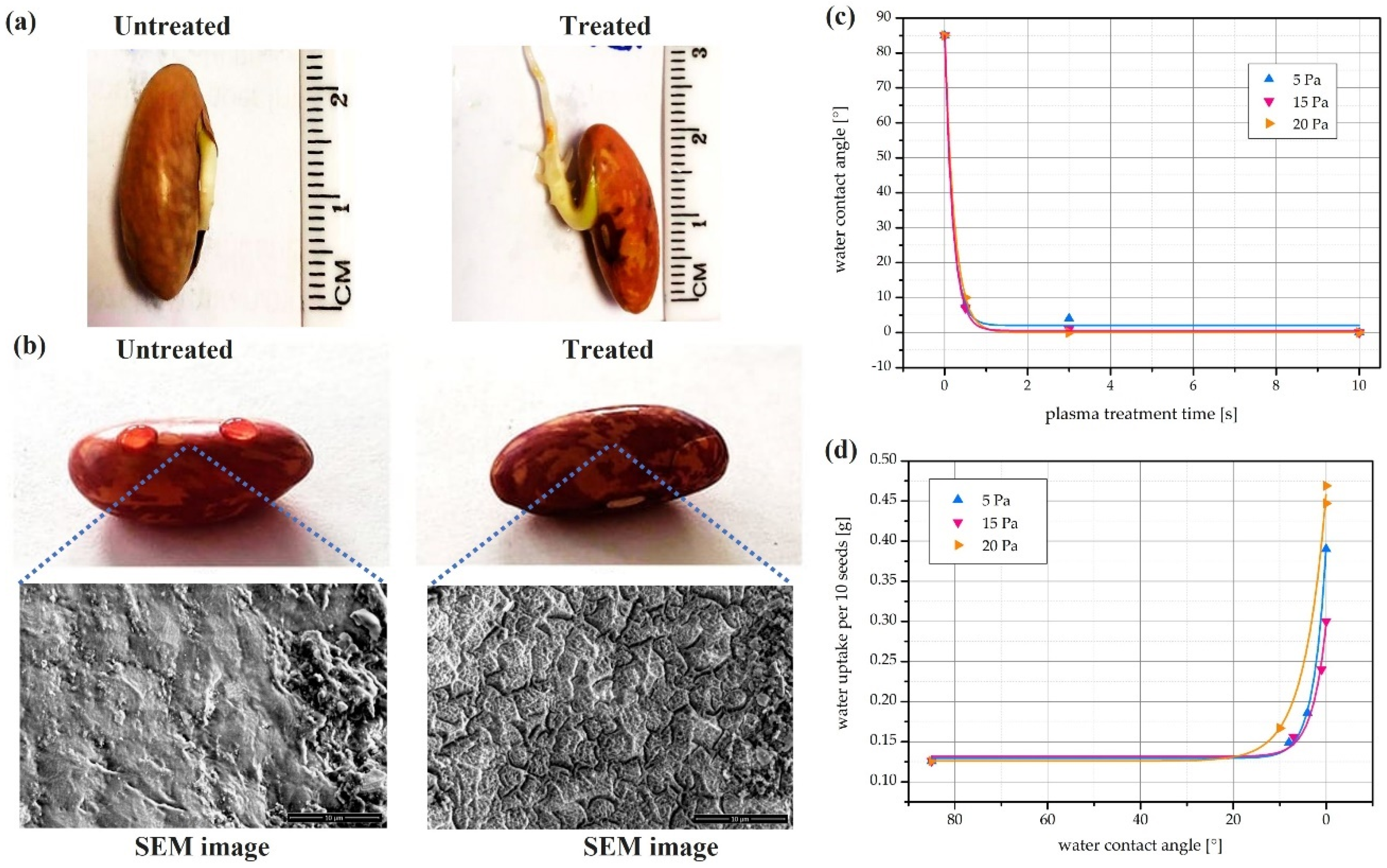
Figure 7.
The effect of CAP-treated tap water on lentil seeds’ seedling. Pictures showing lentil seeds and seedlings on days 6, 12, and 18 after treatment. TAP: tap water, DEM: demineralized water, FTZ: liquid fertilizer, Ctrl: control, PAL: plasma activated liquid [59].
Figure 7.
The effect of CAP-treated tap water on lentil seeds’ seedling. Pictures showing lentil seeds and seedlings on days 6, 12, and 18 after treatment. TAP: tap water, DEM: demineralized water, FTZ: liquid fertilizer, Ctrl: control, PAL: plasma activated liquid [59].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yan, D.; Lin, L.; Zvansky, M.; Kohanzadeh, L.; Taban, S.; Chriqui, S.; Keidar, M. Improving Seed Germination by Cold Atmospheric Plasma. Plasma 2022, 5, 98-110. https://doi.org/10.3390/plasma5010008
AMA Style
Yan D, Lin L, Zvansky M, Kohanzadeh L, Taban S, Chriqui S, Keidar M. Improving Seed Germination by Cold Atmospheric Plasma. Plasma. 2022; 5(1):98-110. https://doi.org/10.3390/plasma5010008
Chicago/Turabian StyleYan, Dayun, Li Lin, Michelle Zvansky, Leat Kohanzadeh, Shannon Taban, Sabrina Chriqui, and Michael Keidar. 2022. "Improving Seed Germination by Cold Atmospheric Plasma" Plasma 5, no. 1: 98-110. https://doi.org/10.3390/plasma5010008