Independence Effects of Heat and Ash on Forest Soil Nematode-Trapping Fungi Communities

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Design
- -
- Short-term flaming group (FS): representing a short time burn of 20 min. We used a flame gun to replicate the chemical reaction induced by the flames of a fire and the associated heat directly on the soil but without the residual ashes resulting from the biomass combustion.
- -
- Long-term flaming group (FL): representing a long time burn of 60 min. In this treatment, the flame gun was used as in FS group, but for a duration of 1 h.
- -
- Heating without flames group (Charcoal): In this treatment we placed 200 g of smoldering charcoal in a thin iron box and left on the soil units for 1 h, to expose the soil to heat without being directly affected by the contact with the flames and preventing ash inputs.
- -
- Cool ash group (ash): We spread ash, after cooling (removing heat or flames’ effects), over the soil units, corresponding to burning (in a different location) the same amount of litter layer accumulated on the pine forest stand (approximately 260 g for a 10 × 10 cm soil unit).
- -
- Natural burning group: 260 g of the previously collected litter layer of the pine forest was placed on each soil sample and burned as in a natural surface forest fire.
- -
- Control group: no treatment was performed on the soil units.
2.2. Soil Temperature Measurement and Soil Sampling
2.3. Measurement of Soil Physicochemical Properties
2.4. Isolation, Identification and Count of NTF
2.5. Data Analysis
3. Results
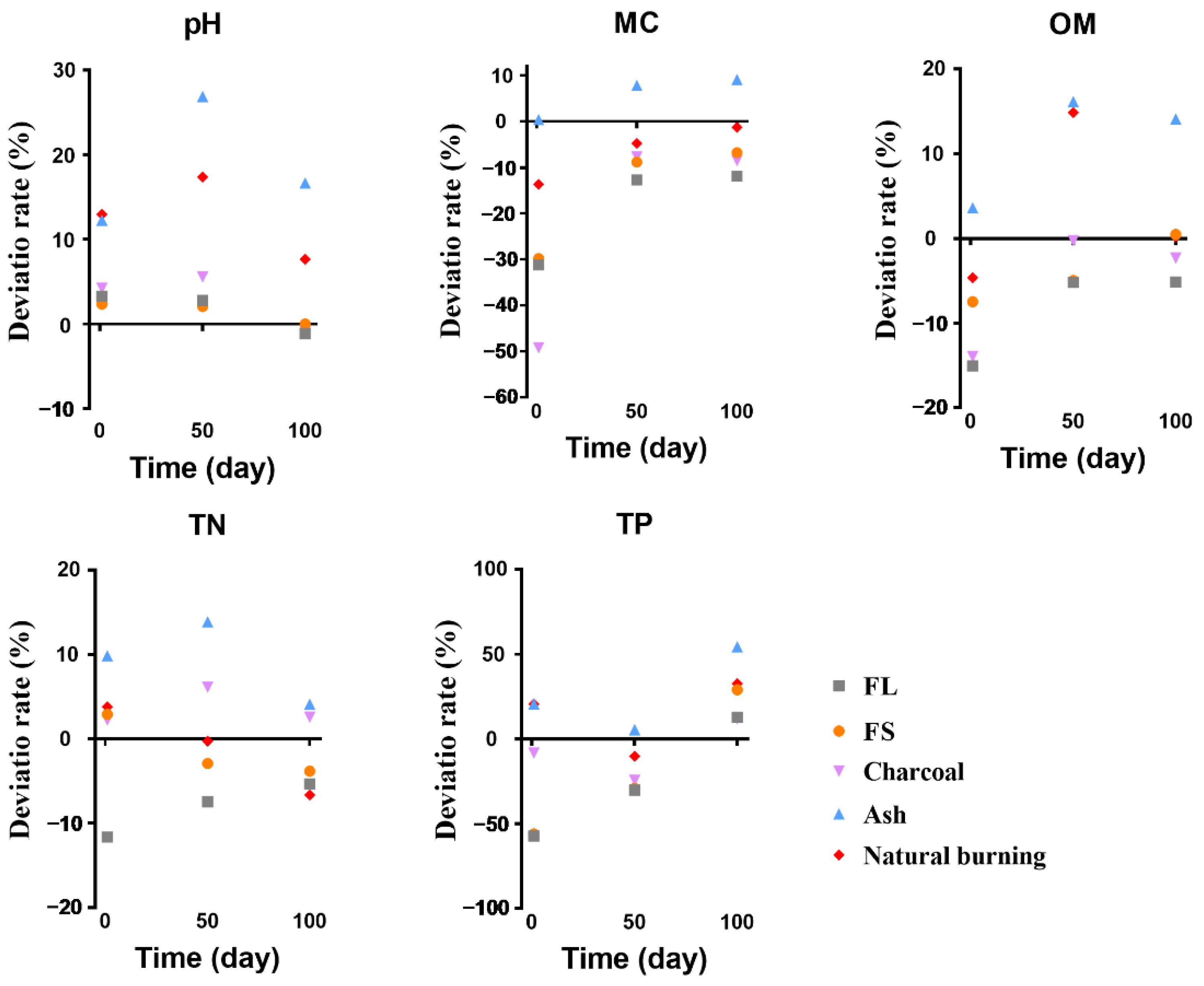
3.1. Soil Physicochemical Properties
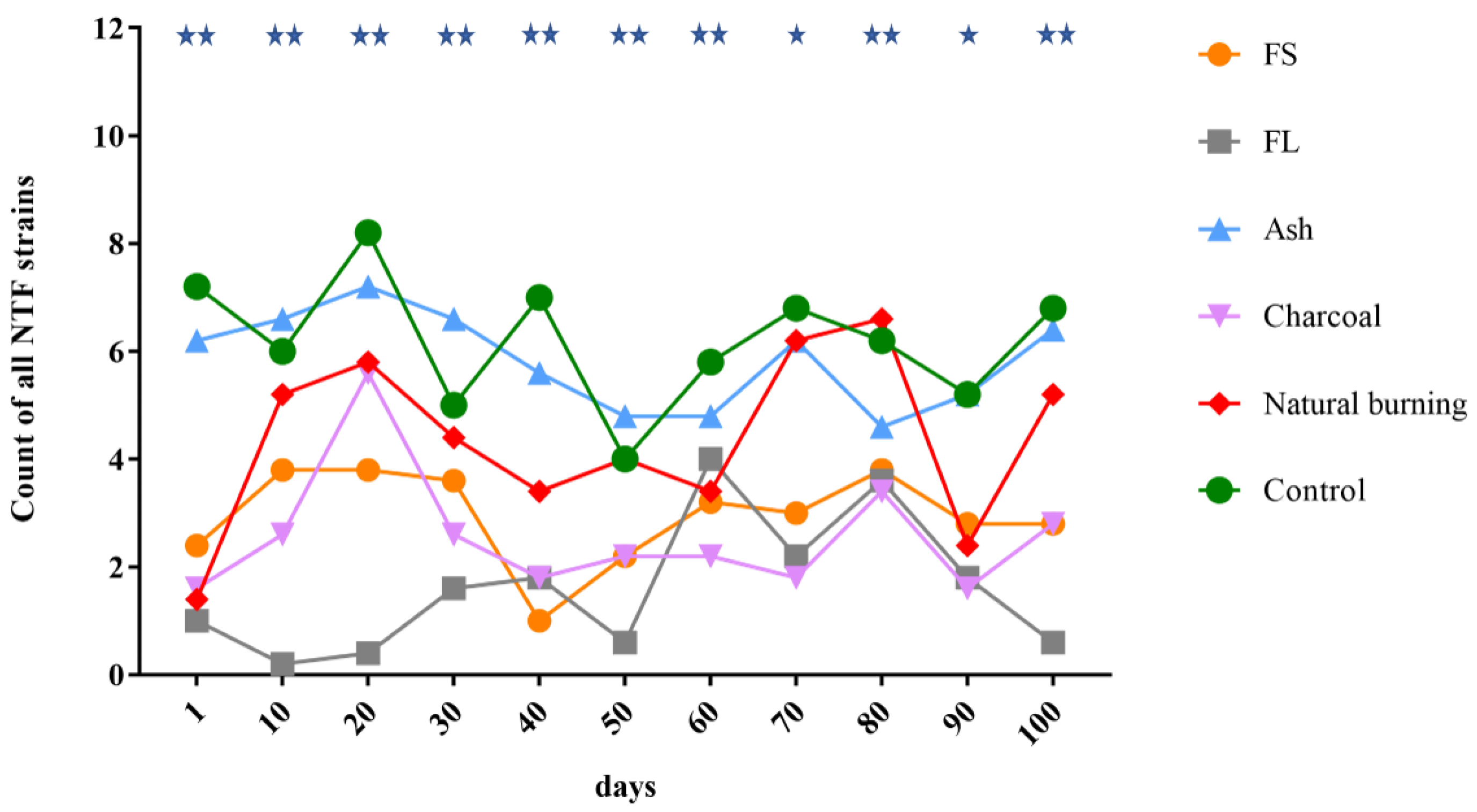
3.2. Differences in the Counts of All NTF Strains
3.3. Relationship between NTF Community and Heat Input
3.4. Time-Varying Changes in the Counts of All NTF Strains
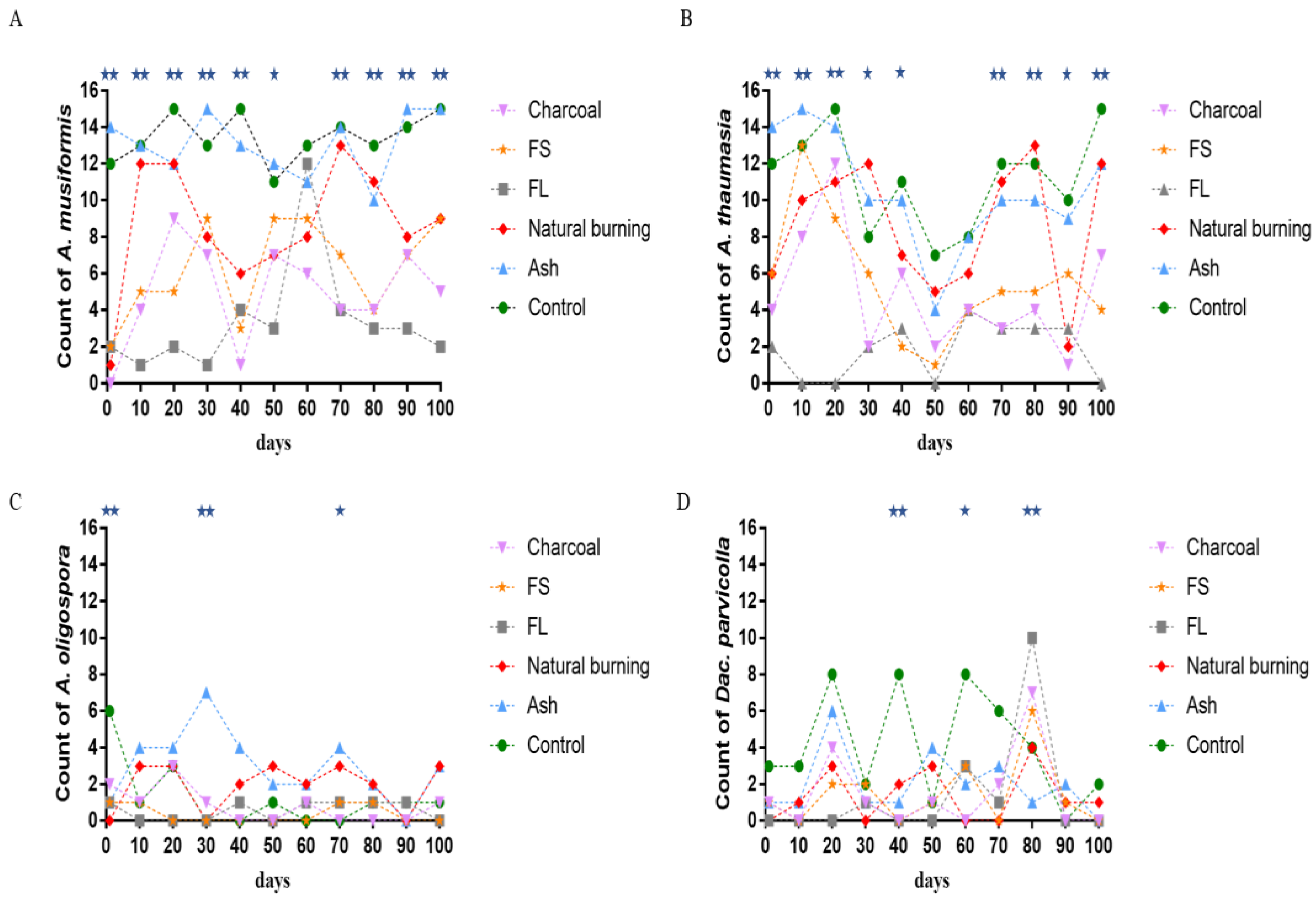
3.5. Time Variation in the Counts of the Widely Distributed NTF Species
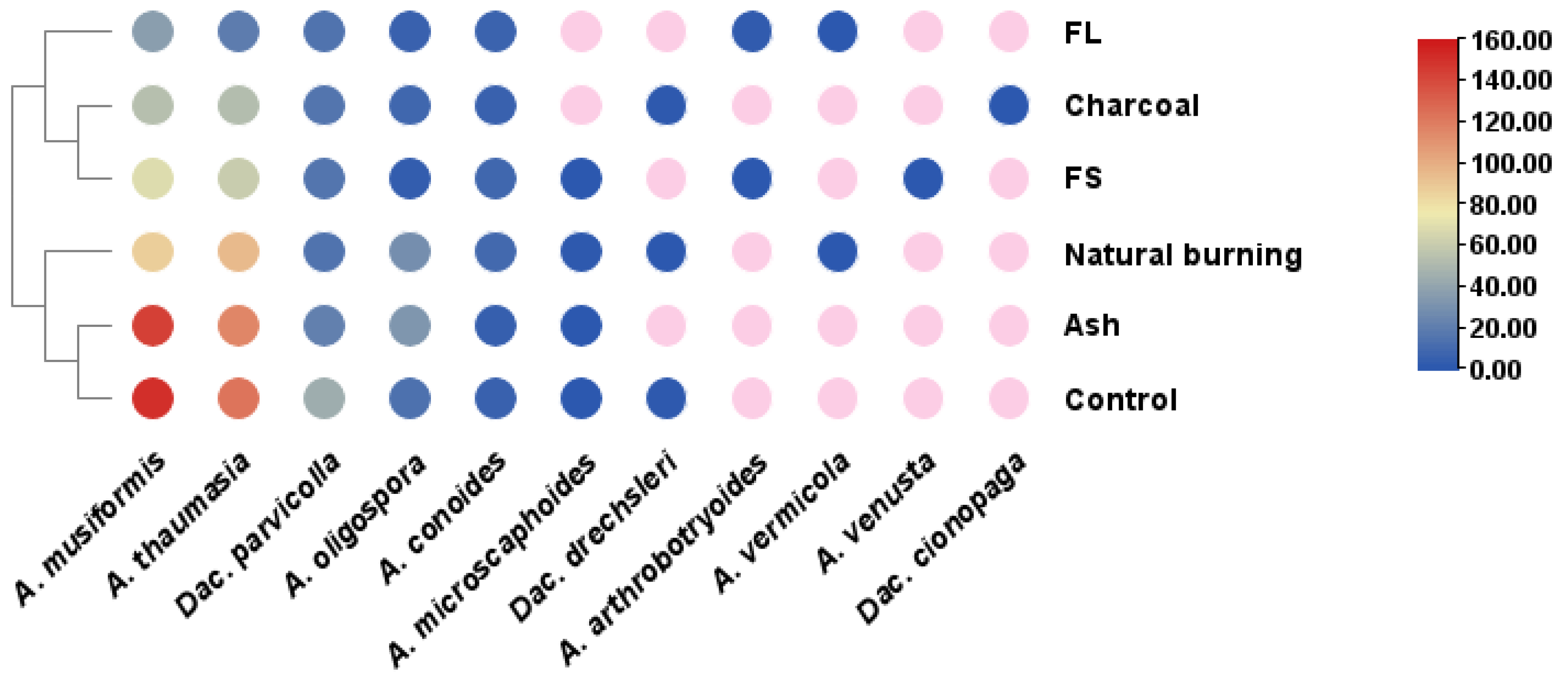
3.6. Differences within the Composition and Structure of the NTF Community
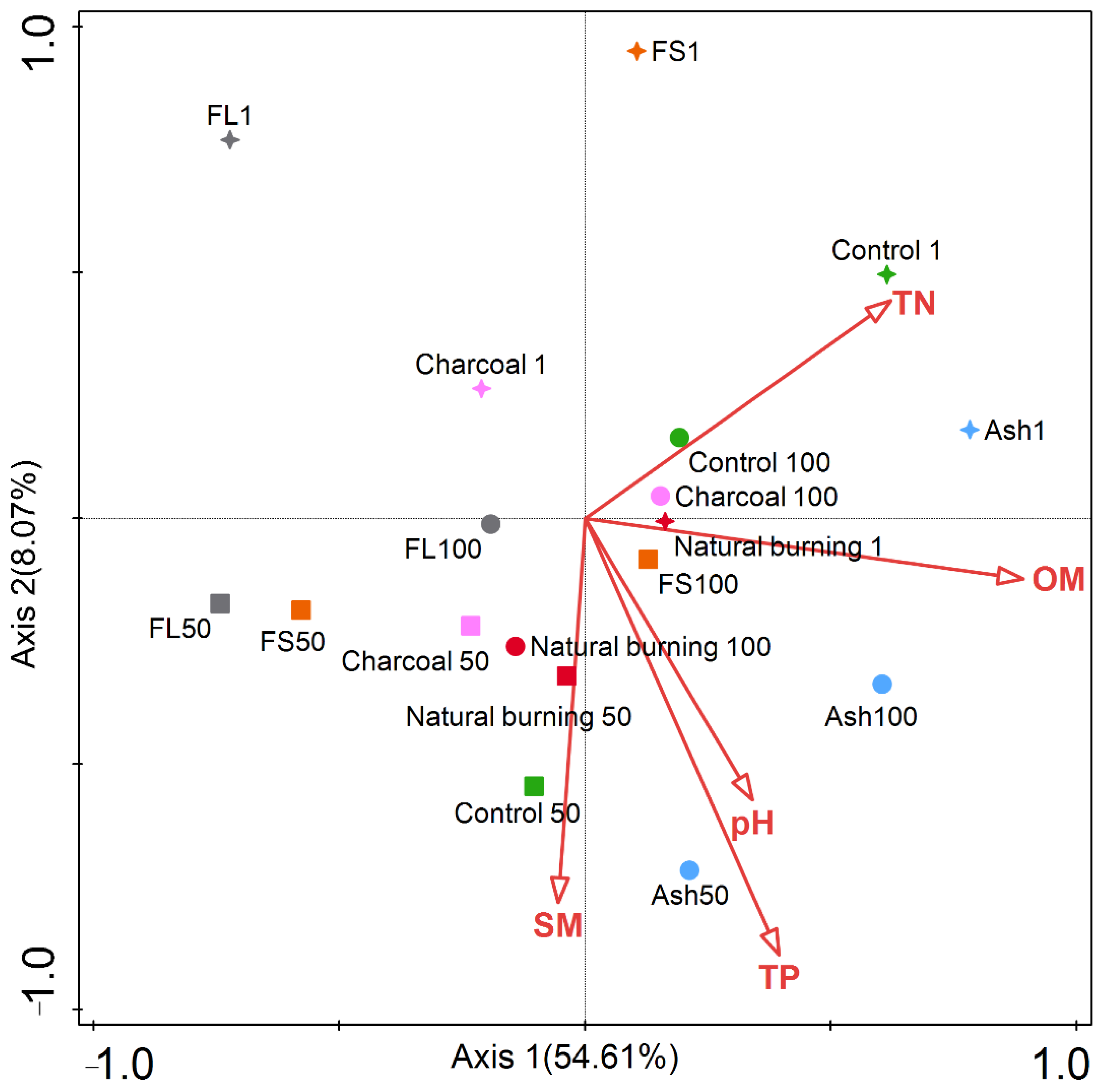
3.7. Interrelationships between Soil Physicochemical Properties and NTF Community
4. Discussion
4.1. Effects of Heat Input on Soil NTF Communities
4.2. Effects of Ash on Soil NTF Communities
4.3. Interference of Fire on Different NTF Species
4.4. Limitations of Designed Fire Experiment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Lamont, B.B.; Pausas, J.G. Fire as a Key Driver of Earth’s Biodiversity. Biol. Rev. 2019, 94, 1983–2010. [Google Scholar] [CrossRef]
- Pausas, J.; Keeley, J.E. A Burning Story: The Role of Fire in the History of Life. Bioscience 2009, 59, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Krawchuk, M.A.; Moritz, M.A.; Parisien, M.-A.; Van Dorn, J.; Hayhoe, K. Global Pyrogeography: The Current and Future Distribution of Wildfire. PLoS ONE 2009, 4, e5102. [Google Scholar] [CrossRef]
- Hantson, S.; Pueyo, S.; Chuvieco, E. Global fire size distribution is driven by human impact and climate. Glob. Ecol. Biogeogr. 2014, 24, 77–86. [Google Scholar] [CrossRef]
- Prichard, S.J.; Stevens-Rumann, C.S.; Hessburg, P.F. Tamm Review: Shifting global fire regimes: Lessons from reburns and research needs. For. Ecol. Manag. 2017, 396, 217–233. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Lombao, A.; Barreiro, A.; Fontúrbel, M.T.; Martín, A.; Carballas, T.; Díaz-Raviña, M. Effect of Repeated Soil Heating at Different Temperatures on Microbial Activity in Two Burned Soils. Sci. Total Environ. 2021, 799, 149440. [Google Scholar] [CrossRef]
- Pereira, P.; Úbeda, X.; Francos, M. Laboratory Fire Simulations: Plant Litter and Soils. In Fire Effects on Soil Properties; Taylor & Francis Group: Leiden, The Netherlands, 2019. [Google Scholar]
- Kumar, S.; Kaushik, G.; Dar, M.A.; Nimesh, S.; López-Chuken, U.J.; Villarreal-Chiu, J.F. Microbial Degradation of Organophosphate Pesticides: A Review. Pedosphere 2018, 28, 190–208. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- de Faria, M.R.; Costa, L.S.A.S.; Chiaramonte, J.B.; Bettiol, W.; Mendes, R. The rhizosphere microbiome: Functions, dynamics, and role in plant protection. Trop. Plant Pathol. 2021, 46, 13–25. [Google Scholar] [CrossRef]
- Kandlikar, G.S.; Johnson, C.; Yan, X.; Kraft, N.J.B.; Levine, J.M. Winning and losing with microbes: How microbially mediated fitness differences influence plant diversity. Ecol. Lett. 2019, 22, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Gao, Q.; Yuan, M.; Wang, G.; Zhou, X.; Feng, J.; Shi, Z.; Hale, L.; Wu, L.; Zhou, A.; et al. Gene-informed decomposition model predicts lower soil carbon loss due to persistent microbial adaptation to warming. Nat. Commun. 2020, 11, 4897. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D.; Shi, Z.; Zhou, X.; Wu, L.; Yang, Y.; et al. Climate warming enhances microbial network complexity and stability. Nat. Clim. Chang. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Xin, W.; Hong, L.G.; Gang, Z.C.; Lai, J.X.; Tong, L.; Ji, Z.P.; Ming, L.L.; Ping, X.J.; Qiang, A.Z.; Zheng, X.; et al. Bacteria Can Mobilize Nematode-Trapping Fungi to Kill Nematodes. Nat. Commun. 2014, 5, 5776. [Google Scholar]
- Rong, S.; Xin-Juan, Z.; Hai-Qing, W.; Fa, Z.; Xiao-Yan, Y.; Wen, X. Succession of soil nematode-trapping fungi following fire disturbance in forest. J. For. Res. 2020, 25, 433–438. [Google Scholar] [CrossRef]
- Pausas, J.G.; Su, W.; Luo, C.; Shen, Z. A shrubby resprouting pine with serotinous cones endemic to southwest China. Ecology 2021, 102, e03282. [Google Scholar] [CrossRef]
- Castellnou, M.; Kraus, D.; Miralles, M. Prescribed Burning and Supression Fire Techniques: From Fuel to Landscape Management. In Best Practices of Fire Use- Prescribed Burning and Suppression Fire Programmes in Selected Case-Study Regions in Europe; Montiel, C., Kraus, D., Eds.; European Forest Institute: Joensuu, Findland, 2010; pp. 4–16. [Google Scholar]
- McCaw, W.L. Managing forest fuels using prescribed fire—A perspective from southern Australia. For. Ecol. Manag. 2013, 294, 217–224. [Google Scholar] [CrossRef]
- Zhang, K.; Hyde, K.D. Nemotode-Trapping Fungi; Springer: Berlin, The Netherlands, 2014; p. 411. [Google Scholar]
- Li, J.; Hyde, K.; Zhang, K.-Q. Methodology for Studying Nematophagous Fungi; Springer: Dordrecht, The Netherlands, 2014; pp. 13–40. [Google Scholar] [CrossRef]
- Deng, W.; Wang, J.-L.; Scott, M.B.; Fang, Y.-H.; Liu, S.-R.; Yang, X.-Y.; Xiao, W. Sampling methods affect Nematode-Trapping Fungi biodiversity patterns across an elevational gradient. BMC Microbiol. 2020, 20, 15. [Google Scholar] [CrossRef]
- Mo, M.-H.; Chen, W.-M.; Yang, H.-R.; Zhang, K.-Q. Diversity and metal tolerance of nematode-trapping fungi in Pb-polluted soils. J. Microbiol. 2008, 46, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.-A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]
- Li, W.; Niu, S.; Liu, X.; Wang, J. Short-term response of the soil bacterial community to differing wildfire severity in Pinus tabulaeformis stands. Sci. Rep. 2019, 9, 1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 143, 200–201. [Google Scholar] [CrossRef]
- Gao, Y.; Dong, S.; Wang, C.; Chen, Y.; Hu, W. Effect of thermal intensity and initial moisture content on heat and moisture transfer in unsaturated soil. Sustain. Cities Soc. 2020, 55, 102069. [Google Scholar] [CrossRef]
- Jiao, S.; Chen, W.; Wang, J.; Du, N.; Li, Q.; Wei, G. Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested ecosystems. Microbiome 2018, 6, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayos, C.B. The roles of texture and structure in the water retention capacity of burnt Mediterranean soils with varying rainfall. Catena 1997, 31, 219–236. [Google Scholar] [CrossRef]
- DeBano, L. The role of fire and soil heating on water repellency in wildland environments: A review. J. Hydrol. Hydromech. 2000, 231–232, 195–206. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, X.-J.; Monkai, J.; Li, F.-T.; Liu, S.-R.; Yang, X.-Y.; Wen, X.; Hyde, K.D. Two new species of nematode-trapping fungi (Dactylellina, Orbiliaceae) from burned forest in Yunnan, China. Phytotaxa 2020, 452, 65–74. [Google Scholar] [CrossRef]
- Miller, A.N.; Raudabaugh, D.B.; Iturriaga, T.; Matheny, P.B.; Petersen, R.H.; Hughes, K.W.; Gube, M.; Powers, R.A.; James, T.Y.; O’Donnell, K. First report of the post-fire morel Morchella exuberans in eastern North America. Mycologia 2017, 109, 710–714. [Google Scholar] [CrossRef]
- Loizides, M.; Bellanger, J.-M.; Clowez, P.; Richard, F.; Moreau, P.-A. Combined phylogenetic and morphological studies of true morels (Pezizales, Ascomycota) in Cyprus reveal significant diversity, including Morchella arbutiphila and M. disparilis spp. nov. Mycol. Prog. 2016, 15, 15–39. [Google Scholar] [CrossRef]
- Sorensen, J.W.; Shade, A. Dormancy dynamics and dispersal contribute to soil microbiome resilience. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicklow, D.T. Fire as an Environmental Cue Initiating Ascomycete Development in a Tallgrass Prairie. Mycologia 2018, 67, 852–862. [Google Scholar] [CrossRef]
- Lennon, J.T.; Jones, S.E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat. Rev. Genet. 2011, 9, 119–130. [Google Scholar] [CrossRef]
- Lamont, B.B.; He, T.; Yan, Z. Fire as a pre-emptive evolutionary trigger among seed plants. Perspect. Plant Ecol. Evol. Syst. 2019, 36, 13–23. [Google Scholar] [CrossRef]
- McLauchlan, K.K.; Higuera, P.E.; Miesel, J.; Rogers, B.M.; Schweitzer, J.; Shuman, J.K.; Tepley, A.J.; Varner, J.M.; Veblen, T.T.; Adalsteinsson, S.A.; et al. Fire as a fundamental ecological process: Research advances and frontiers. J. Ecol. 2020, 108, 2047–2069. [Google Scholar] [CrossRef]
- Fox, S.; Sikes, B.A.; Brown, S.P.; Cripps, C.L.; Glassman, S.I.; Hughes, K.; Semenova-Nelsen, T.; Jumpponen, A. Fire as a driver of fungal diversity—A synthesis of current knowledge. Mycologia 2022, 114, 215–241. [Google Scholar] [CrossRef]
- Bodí, M.B.; Martin, D.A.; Balfour, V.N.; Santín, C.; Doerr, S.H.; Pereira, P.; Cerdà, A.; Mataix-Solera, J. Wildland Fire Ash: Production, Composition and Eco-Hydro-Geomorphic Effects. Earth-Sci. Rev. 2014, 130, 103–127. [Google Scholar] [CrossRef]
- Francos, M.; Úbeda, X.; Pereira, P. Impact of torrential rainfall and salvage logging on post-wildfire soil properties in NE Iberian Peninsula. Catena 2019, 177, 210–218. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, H.; Sietiö, O.-M.; Pumpanen, J.; Heinonsalo, J.; Köster, K.; Berninger, F. Wildfire effects on soil bacterial community and its potential functions in a permafrost region of Canada. Appl. Soil Ecol. 2020, 156, 103713. [Google Scholar] [CrossRef]
- Salo, K.; Domisch, T.; Kouki, J. Forest wildfire and 12 years of post-disturbance succession of saprotrophic macrofungi (Basidiomycota, Ascomycota). For. Ecol. Manag. 2019, 451, 117454. [Google Scholar] [CrossRef]
- Rodríguez, J.; González-Pérez, J.A.; Turmero, A.; Hernández, M.; Ball, A.S.; González-Vila, F.J.; Arias, M.E. Physico-Chemical and Microbial Perturbations of Andalusian Pine Forest Soils Following a Wildfire. Sci. Total Environ. 2018, 634, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Adkins, J.; Docherty, K.M.; Gutknecht, J.L.; Miesel, J.R. How do soil microbial communities respond to fire in the intermediate term? Investigating direct and indirect effects associated with fire occurrence and burn severity. Sci. Total. Environ. 2020, 745, 140957. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, E.; An, Z.; Liu, X. Evolution of Nematode-Trapping Cells of Predatory Fungi of the Orbiliaceae Based on Evidence from Rrna-Encoding DNA and Multiprotein Sequences. Proc. Natl. Acad. Sci. USA 2007, 104, 8379–8384. [Google Scholar] [CrossRef] [Green Version]
- Cooke, R.C. The ecology of nematode-trapping fungi in the soil. Ann. Appl. Biol. 1962, 50, 507–513. [Google Scholar] [CrossRef]
- Fan, X.-J.; Zhang, X.; Zhang, F.; Liu, S.-R.; Su, X.-J.; Yang, X.-Y. Dactylellina Parvicolla Wz27, a Spontaneous Conidial Trap Producing Strain of Nematode-Trapping Fungus. Mycosystema 2018, 37, 305–313, (in Chinese with English abstract). [Google Scholar]
- Kearns, P.J.; Shade, A. Trait-based patterns of microbial dynamics in dormancy potential and heterotrophic strategy: Case studies of resource-based and post-press succession. ISME J. 2018, 12, 2575–2581. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
She, R.; Wang, H.-Q.; Fornacca, D.; Li, F.-T.; Zhang, F.; Yang, Y.-Q.; Zhou, F.-P.; Yang, X.-Y.; Xiao, W. Independence Effects of Heat and Ash on Forest Soil Nematode-Trapping Fungi Communities. Fire 2023, 6, 27. https://doi.org/10.3390/fire6010027
She R, Wang H-Q, Fornacca D, Li F-T, Zhang F, Yang Y-Q, Zhou F-P, Yang X-Y, Xiao W. Independence Effects of Heat and Ash on Forest Soil Nematode-Trapping Fungi Communities. Fire. 2023; 6(1):27. https://doi.org/10.3390/fire6010027
Chicago/Turabian StyleShe, Rong, Hai-Qing Wang, Davide Fornacca, Fei-Teng Li, Fa Zhang, Yao-Quan Yang, Fa-Ping Zhou, Xiao-Yan Yang, and Wen Xiao. 2023. "Independence Effects of Heat and Ash on Forest Soil Nematode-Trapping Fungi Communities" Fire 6, no. 1: 27. https://doi.org/10.3390/fire6010027