
The Relevance of Astrocytic Cell Culture Models for Neuroinflammation in Neurodegeneration Research
by
 and
and
André Maciel Preato
1,†,
Ester da Silva Pinheiro
1,†,
Tatiana Rosado Rosenstock
2 and
Isaias Glezer
1,* 1
Department of Biochemistry, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo 04044-020, Brazil
2
Department of Bioscience, In-Vitro Neuroscience, Sygnature Discovery, Nottingham NG1 1GR, UK
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Neuroglia 2024, 5(1), 27-49; https://doi.org/10.3390/neuroglia5010003
Submission received: 31 December 2023
/
Revised: 21 February 2024
/
Accepted: 22 February 2024
/
Published: 29 February 2024
Abstract
:Astrocytes are the predominant glial cells that provide essential support to neurons and promote microenvironment changes in neuropathological states. Astrocyte and astrocytic-like cell culture have substantially contributed to elucidating the molecular pathways involved in key glial roles, including those relevant to neurodevelopment, brain physiology and metabolism, which are not readily accessible with traditional approaches. The in vitro methodology has also been applied to neuroinflammatory and neurodegeneration contexts, revealing cellular changes involved in brain dysfunction. Astrocytes studies in culture started with primary cell approaches using embryonic and postmortem tissue. Further developments included newborn rodent primary cells, cell lines and immortalized astrocytes, which resulted in homogeneous cell-type preparations grown on flat surfaces. To overcome some in vitro shortcomings, tridimensional bioprinted models and organoid culture enabled the mimicking of tissue cellular arrangements and, above these achievements, complex astrocyte cell culture can be generated from induced pluripotent stem cells (iPSCs) to model diseases. These unprecedented breakthroughs allowed the development of platforms to test new therapies in brain cells derived from human material noninvasively obtained from live patients. In this work, we reviewed the most studied astrocytic cell models for discussing limitations, advantages and reliable experimental readouts for neuroinflammation in neurodegeneration research.
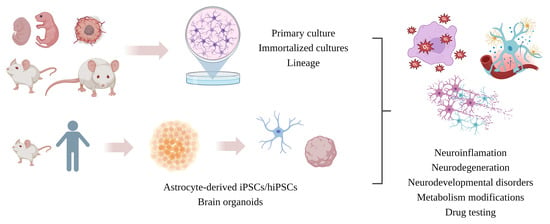
1. Introduction
Glial cells are non-neuronal cells initially described during the 19th century as connective brain elements, which were not thought to be cellular but capable of keeping the brain tissue together [1,2,3] The name derives from their adhesive (glue-like) properties. Initially, the development of knowledge on glial cells was overshadowed by the research on neurons because these are the electrically excitable cells that convey neurotransmitter-based signals in distinct brain circuits [4]. One outcome of this neuronal-centered perspective is that a simplified supporting and protective role has been attributed to the glia for a long period. Astrocytes are well known for providing metabolic and trophic signals to neurons, while forming a blood barrier when associated with neurovascular unity. Oligodendrocyte function is traditionally understood as axon insulation to promote saltatory fast conduction. Microglia are usually defined as immune cells that protect neurons from potential pathogens [5]. All these glial cell types have more elaborate and complex roles in the CNS, and astrocyte research has consistently shown that these cells are versatile, essential to normal brain function and can play important roles in brain diseases by impacting the function of neurons, oligodendrocytes and microglia.
The notion of glia heterogeneity became clear when the star-shaped cells were named astrocytes [6] and, later, cells with other morphologies and characteristics were identified as oligodendrocytes and microglia [7,8,9,10,11,12]. Golgi made important initial descriptions of astrocytes, including the end-feet glia-vascular contacts. Later, Cajal further developed an investigation technique that employed the gold chloride-sublimate staining, which is specific for both protoplasmatic (gray matter) and fibrous (white matter) astrocytes. These and other developments resulted in particular attention to astrocytes during those years [13,14,15,16,17,18,19,20]. Once it was understood that astrocytes represent a distinct cell type, one of the challenges was to isolate these cells to study them apart from the other neural cells (see Figure 1 for a timeline). Here, we review how in vitro experimental approaches with astrocytes provide the means to dissect and test hypotheses on cell-autonomous mechanisms, and we evaluate real-time cellular events in different conditions, including physiological and inflammation models related to neuropathologies. While addressing these aspects, we also present examples of how these models have been applied in studying conditions related to neurodegeneration to illuminate their potential contributions to understanding neuroinflammation and broader neurological disorders.
Microglia and astrocyte reactivity is frequently associated with neurodegenerative diseases, including those that feature protein misfolding and aggregation. Although inflammation is not considered an initiating factor in neurodegenerative diseases, it has been found that neuroinflammation occurs earlier than protein aggregation. In addition, the literature suggests that sustained and deregulated inflammatory responses involving microglia and astrocytes contribute to disease progression. The relationship between inflammation and neurodegenerative diseases have been substantially reviewed elsewhere [21,22,23]. Therapeutic approaches aiming to modulate neuroinflammation in neurodegenerative diseases depend on mechanistic research that dissects precise components of cellular responses, which is currently investigated in astrocytes through several approaches.
At the time of writing this paper, almost 77,000 articles in the national library of medicine (pubmed.gov or www.ncbi.nlm.nih.gov/pubmed/; accessed 21 February 2024) were retrieved with “astrocyte” as the search term, nearly 8000 articles when the term “cell culture” was conditionally added to the search as the obligatory (AND) term and almost 562 articles were retrieved when combining these two terms with “inflammation”. Unfortunately, we are unable to list all potential and relevant work here. When using MeSH terms “((astrocyte[MeSH Terms]) AND (cell culture[MeSH Terms])) AND (inflammation[MeSH Terms])”, the search returned 50 results that were prioritized for selection in this review along with other search strategies. According to official protein/gene symbol notation, we adopted the use of all uppercase nonitalicized letters in abbreviations for proteins (without special characters or hyphen), all uppercase letters italicized for human gene and transcript and only first uppercase and italicized letters for rodent genes/transcript.
2. A Brief Overview of Astrocytes General Features and Physiological Roles
The descriptive approach in the astroglial biology field predominated until the development of purified astrocyte cultures and astrocytic cell lines. However, one of the earliest perceptions from morphological studies led to an important hypothesis that glia drive the connections between themselves and blood vessels, in addition to their participation in cellular interactions that occur during brain development [24,25,26,27,28,29]. Intriguingly, a recent study estimated that 99.8% of all gray matter astrocytes are connected with blood vessels, thereby reinforcing their role in metabolic supply [30].
Revised concepts on the importance of glia in brain functions have been claimed in recent decades, in part due to the popularization that such cells, mainly astrocytes, outnumber human neurons by one order of magnitude. However, detailed literature review and modern cell count techniques, including unbiased stereology and isotropic fractionation, showed that this is not the case. Glia do not extensively outnumber neurons in general, being less than 2:1 in the human cortex and around 3:1 in the spinal cord [31,32,33]. While cell number is not equivalent to cell importance, it is relevant to understanding how neuron-to-glia ratio changes in different species (a higher proportion of astrocytes according to increased brain size), regions (e.g., ~12:1 in the posterior gray matter of the thoracic human spinal cord) and during development and pathological states [34]. Hence, we are tempted to suppose that astrocyte functions in nutrient supply and energetic metabolism should be critical in species with larger brains, which could explain the higher proportion of astrocytes. However, it is not clear what the precise role is for different astrocyte tissue densities in normal physiology.
Astrocytes play critical roles in CNS homeostasis because, in addition to supporting neuronal function, these cells also influence neurotransmission and other glial cells. The physiological activation of the astrocytes involves calcium propagation (calcium waves), a mechanism that allows cellular communication with other astrocytes through gap junctions channels; or pre- and postsynaptic neurons, forming the tripartite synapse by regulating neurotransmitters glutamate and GABA uptake and synthesis [16,35,36,37,38,39,40,41,42,43,44,45]. In addition, astrocytes are implicated in brain tissue waste drainage by fluid exchange from the cerebrospinal fluid (CSF) compartment to the interstitial fluid through astrocytic aquaporin 4 (AQP4), which is essential for normal function of the glymphatic system [46], a newly described waste clearance system. Toxic residues, such as Aβ-amyloid peptides and the Alzheimer’s disease (AD) pathology-associated tau protein, for example, are removed from the brain by the glymphatic system. Local homeostasis at Ranvier nodes, the site of saltatory conduction of action potentials, and oligodendrocyte normal functions, also involves astrocytes that modulate blood flow through vasoactive nitric oxide (NO) and arachidonic acid, and provides energetic metabolites glucose and lactate. Astrocytes projections called end-feets form remarkable associations with the vascular endothelial cells, determining the formation and maintenance of the blood–brain barrier (BBB). These interconnections are included in the structuring of the outermost glial limiting membrane (glia limitans), which insulates the brain parenchyma from the vascular and subarachnoid compartments [47,48,49].
Molecular markers immunostaining successfully allow the identification of astrocytes. Although these cells represent a heterogeneous group, they consistently express glial fibrillary acidic protein (GFAP). This intermediate filament protein is involved in cell motility and structural stability, and is highly expressed during brain damage and other insults. Other markers include another intermediate filament protein, nestin and brain lipid binding protein (BLBP) in the case of immature astrocytes. Mature cells express calcium-binding protein S100β/S100B, excitatory amino acid transporters EAAT1/GLAST-1/SLC1A3 and EAAT2/GLT-1/SLC1A2, glutamine synthetase (GS), aldehyde dehydrogenase ALDH1L1, AQP4 and gap junction components connexin 30 (Cx30/GJB6) and Cx43/GJA1 [50,51,52,53,54,55,56,57,58,59,60]. These molecules are indeed useful for identifying astrocytes, and their functions are well documented in key functional aspects of astrocytic cells. We highlight that these markers may not be used unequivocally for every occasion. GFAP gene expression is usually observed in white matter fibrous astrocytes, and its expression in radial glial progenitor cells makes it less specific [61]. S100B, in contrast, has shown to be predominantly expressed in gray matter protoplasmatic astrocytes, and in differentiating oligodendrocytes [62].
Astrocyte markers should ideally reflect their relevant molecular functions and can be selected and/or used in combination to increase specificity and label most types of astrocytes in different contexts. A recent study profiled astrocyte density and molecular signatures in different murine brain regions using fluorescent labeling driven by Aldh1l1 promoter, which suggested that neuron-to-astrocyte ratios change because of neuronal density, and not because of astrocyte cell numbers. Although astrocyte density is quite homogenous and correlated with endothelial cell numbers, transcriptional signatures point to the existence of molecular features and functions specific to brain regions [63]. In the future, cell markers, alongside neuroanatomical and functional information, will have to be combined for proper evaluation of the varied spectrum of astrocyte physiology. These recent developments on astrocyte heterogeneity will also impact the design, interpretations and perception of limitations for studies conducted in vitro.
3. Reactive Astrocytes and Pathological States
The role of astrocytes in neuropathologies has been intensively studied, including neuroinflammation, neurodegenerative diseases and neurodevelopmental disorders. The mechanisms by which astrocytes intensify or cause detrimental effects to brain elements include: (1) loss of tissue homeostasis control; (2) disease progression by transduction of inflammatory signals into a changed cellular microenvironment; and (3) secretion of molecules that damage or affect other brain cells. Much of the knowledge on astrocytes behavior in pathological states is related to a reactive phenotype, which will be discussed in the following paragraphs.
Escartin et al., in a review article signed by leading research groups, discussed that the terminology for astrocytes in pathological states is not well defined [64]. Indeed, the confusing terminology in the literature, and the lack of validation of in vitro findings using in vivo models, render the comparisons between studies difficult and challenging. According to that review work, special attention should be given to the following terms: “reactive astrocyte”, “astrocytosis”, “reactive gliosis”, “astrogliosis”, “astrocyte activation” and similar. Following the terminology consistency recommendation, in contrast to “astrocyte activation”, which refers to more physiological contexts, we will apply the terms “reactive astrocytes”, “reactive astrogliosis” and “astrocyte reactivity” when referring to a pathological state, or a model that simulates a pathological context.
In vitro studies designed to mimic disease states are also capable of displaying astrogliosis, including increased chondroitin sulfate proteoglycans (CSPGs), which is the hallmark of this process, along with other features of reactive astrocytes [65,66,67,68,69]. In severe pathological states, it is well recognized that intense astrogliosis takes place during brain injury, infection and neurodegenerative processes in general. The increased astrocyte density and other relevant changes in pathological states, including cell shape, size and function, determine profound changes in brain metabolism and reactive defense mechanisms, such as the glial scar discussed below [64,70].
Reactive astrogliosis and certain astrocytes phenotypes can contribute to progressive neuronal damage as a consequence of a proinflammatory state (called A1-, or A1-like astrocytes, in former literature) and from the loss of the protective and prosurvival functional activities typical of A2-like cells (nomenclature also employed here to match common terminology). The bias toward the “reactive” astrocyte can be triggered by varied stimuli such as injury, pathogens, toxic molecules and so on [71,72,73,74]. It is beyond the scope of this review to discuss reductionism of cell state interpretations, and whether astrocytes reactivity leads solely to the secretion of inflammatory mediators and the increase in potentially harmful molecular species. As one may expect, this concept also claims that these cell changes are detrimental to neuronal elements, due to a decreased release of beneficial and neuroprotective astrocyte-derived factors/metabolites (reviewed in [64,75,76,77]). While we refrain from a simplified view of astrocyte states, we observe that the literature frequently interprets astrocyte responses as a dichotomized polarization. This limitation is often present in cell culture studies as well.
The complete understanding of reactive astrocytes and astrogliosis depends on the observation and analysis of other components of the brain environment. In contrast to the experimental setups that aim to approach the entire scenario, cell culture techniques have contributed to understanding the cell-autonomous mechanisms by which astrocytes modify their functions under noxious stimuli. Different paradigms have shown that astrocytes participate in neurotoxic events; for instance, mediating neuronal cell death upon exposure of Aβ-amyloid peptide from AD, or by secreting proteins that reduce neuronal survival in models of neurodegeneration or neurodevelopmental diseases, such as Fragile X syndrome, Rett syndrome and Down syndrome [78,79,80]. For more information on the different neurodegenerative diseases in which astrocytes play critical roles, we suggest dedicated reviews [81,82,83].
4. Experimental Readouts for Reactive Astrocytes in Inflammation Context
As summarized in Figure 2, the analysis of astrocyte reactivity relies greatly on the activity/expression of astrocyte markers, a few metabolic enzymes and transporters, pro-inflammatory molecules and components of the extracellular matrix (ECM).
In experimental settings for neuroinflammation research, pathogen-associated molecular patterns (PAMPs), such as Gram-negative bacteria lipopolysaccharide (LPS), have been widely employed to engage toll-like receptors (TLRs) and promote proinflammatory gene expression in the brain or in cell culture. Shortly after TLRs characterization in the very early 2000s, it was demonstrated that the effects of LPS infusion in the brain depend on its cognate receptor TLR4 [84]. Lenhardt et al. corroborated this when reporting that astrocytes and oligodendrocytes primary cultures do not express significant amounts of TLR4; instead, only microglia cells express it and are a required factor to promote damage induced by LPS [85]. In a dedicated review a few years later, Saura questioned the literature regarding NO production by astrocytes stimulated with LPS, arguing that small microglial contaminations are often present in astrocyte primary culture preparations, which accounts for a dramatic effect in response to LPS [86]. In consequence, NO and other proinflammatory molecules may not be produced by astrocytes.
Even though contaminating microglia is an alarming issue in cell culture, several studies continued to report evidence that astrocytes are fully capable of mounting innate immune responses through the activation of different signaling pathways [87]. These findings were contested by experiments conducted with primary cultures treated with liposomal clodronate, which reduced microglia and blunted proinflammatory cytokines interleukin 1 β (IL-1β; protein symbol: IL1B) and tumor necrosis factor α (TNF-α; protein symbol: TNF) upon IL6 stimulation [88]. In addition, microglia depletion with a CSF1 receptor inhibitor (PLX-5622) successfully reduced microglia content in astrocyte/microglia mixed cultures. These microglia-depleted cultures were incapable of producing TNF when stimulated with LPS, but maintained partial induction of IL6 [89]. Another CSF1R inhibitor (PLX-3397) was also able to deplete microglia from primary astrocyte culture [90], and is a promising tool, along with other CSF1R inhibitors, to accurately evaluate astrocyte responses in inflammatory conditions.
While microglia is naturally in contact with astrocytes, one must note that microglia in standard culture conditions lose their phenotype [91]. Stimuli that trigger contaminating microglia reactivity and proliferation pose a difficult problem regarding the evaluation of genuine astrocytes responses. This contributes to the finding that microglia secreted factors (C1q, TNF and IL1A) cause robust reactivity of astrocytes [92]. Overall, these observations indicate that IL6 is a suitable cytokine to evaluate astrocyte reactivity and has been known to have functions related to astrocytes for quite a long time [93]. Other transcripts have been identified as pan reactive (Gfap and vimentin/Vim), neuroinflammatory-specific (A1/A1-like), or ischemic-specific (A2/A2-like), discriminating phenotypes and highlighting complement C3 as a good marker of A1/A1-like cells in neurodegenerative diseases [92]. Also of note, oncostatin M receptor (Osmr), ceruloplasmin (Cp) and lipocalin 2 (Lcn2) could be sensitive markers of astrocyte reactivity; in fact, these transcripts show major induction in brain tissue infused with LPS (as well as other transcripts classified as A1- or A2-specific), and Osmr induction presents a relative selectivity for astrocytes [94,95]. A well-studied mechanism by which astrocytes respond to inflammatory stimuli, pathological and/or aging conditions is the release of extracellular vesicles (EVs), promoting neuroprotection and neuronal survival in response to anti-inflammatory cytokines, such as IL10 [96,97]. On the other hand, under pro-inflammatory stimuli, such as IL1B or TNF, secreted EVs can promote inflammatory signal transmission, as well as neuronal plasticity and modulation [97,98,99]. The presence of the inflammasome NLRP3 and NLRC4 expression was observed in both human and mouse astrocytes [100] and NLRP2 only in humans [101]. Especially when related to certain disorders, such as demyelination, multiple sclerosis (MS), AD, amyotrophic lateral sclerosis (ALS) and traumatic brain injury [72,100,102,103,104], culture models were extremely important to some of these findings.
Other readouts were studied in the context of brain damage or neurodegenerative diseases, including CSPG production in response to transforming growth factor beta (TGFB1/2/3) signaling, which resulted in glial scar formation [105,106,107]. Loss of AQP4, which impacts cerebrospinal fluid homeostasis, is also a relevant index of pathological changes in astrocytes relevant to barrier function [108,109]. Morphological changes, calcium signaling, and metabolic analytes (ATP levels, reactive oxygen species, synthesis and uptake of neurotransmitters) present changes in models of AD, Parkinson’s disease (PD) and ALS [75,110].
5. Astrocytic In Vitro Models
When selecting an in vitro model, it is critical to consider the advantages and limitations to provide plausible answers to the investigated biological question. The most used cell culture models are cell lines, primary cell culture, immortalized astrocytes (IA), differentiated induced pluripotent stem cells (iPSCs), 3D culture, bioprinting and organoids/mini-brains. We reviewed the most common astrocytic cell models and evaluated their limitations and advantages (see Table 1), along with the reported outcomes (Table 2). In addition, we show some examples of studies conducted with astrocyte primary culture in Table 1. The outcomes from the analysis of different astrocytic cell culture types, especially in the case of convergent results, provide powerful means to establish specific roles of the most abundant glial cell type in neuroinflammation and neurodegeneration research.
5.1. Primary Cell Culture
Brain cell culture procedures started with studies of neuronal development, perhaps because nerve cell biology was one of the most favorable subjects to be investigated under the microscope in the early 20th century. In fact, the hanging drop technique developed by Ross Harrison in 1910 showed for the first time that it was possible to culture neurons outside the body. This breakthrough highly influenced similar applications for numerous cell types (Figure 1), and subsequent improvements like culture flasks, culture chambers and, more recently, microfluidic devices and others refined neuroscience studies in vitro [129]. Human and other mammalian neuroglia observation in cell culture dates from the 1930s, and possibly earlier, as reviewed in a study by Mary Jane Hogue [130]. However, conditions that made it possible to enrich astrocytes were developed later. For instance, dissociated human brain cells from a fetus gave rise to astrocytes and radial glia cells formerly known as spongioblasts [131,132]. This preparation was successfully used to investigate astrocyte transformation by an oncogenic DNA virus, for example [133]. In the 1970s and early 1980s, it was shown that a newborn rodent brain could be used to obtain macroglia [134], and McCarthy and Vellis successfully separated oligodendrocytes from astrocytes by driving sheer forces through shaking to obtain a purified primary cell culture expressing GFAP with typical astrocyte morphology [135,136,137]. These purified preparations represented key methodologies to characterize metabolic and electrophysiological properties of the astrocytes, including the transport of excitatory neurotransmitter glutamate ([82], reviewed in [83]) which is converted to glutamine because mature astrocytes in culture express glutamine synthase (GS) [138,139,140]. There is also in vitro evidence that astrocytes are metabolically involved in ionic and osmotic homeostasis in the brain [141]. Subsequently, it was found that glutamate cotransport with sodium in astrocytes stimulates aerobic glycolysis, consequently with lactate production, which mediates metabolic coupling between neuronal activity and glucose consumption by the brain [142]. With that in mind, at least four relevant biochemical parameters to brain function can be measured from cell culture material due to astrocyte metabolism: glucose consumption, lactate production, glutamate uptake and glutamine synthesis. Primary culture of combined cell types can also help disclose mechanisms of cell–cell interactions, like those involved in the formation of glia limitans, which is important to BBB [143]. In addition, the study of astrocyte function in neurodegenerative diseases has been improved with primary culture techniques. MS, an autoimmune disease mediated by the demyelination of the CNS, is an example of a degenerative disease where astrocytes play both inflammatory and protective roles, secreting chemokines and promoting cell migration through the BBB, but also favoring CNS remyelination by promoting the differentiation of oligodendrocyte precursor cells. These mechanisms have been well studied by different groups utilizing not only in vivo models [144] but also primary cultures of astrocytes and co-cultures with microglia [100,145,146].
Much of the discussion on primary cell culture, microglial contamination and neuroinflammatory analysis has been presented in the Experimental Readouts for Reactive Astrocytes in an Inflammatory Context Section (see above). In fact, primary cell culture of astrocytes is the most used model in vitro to evaluate how these cells respond to inflammation.
Although it is not possible to promptly translate in vitro findings into physiological astrocyte function in brain tissue, primary cell culture data not only enabled exploring new mechanisms and cellular features, but also provided proof of concept that is very difficult to show in vivo. It should be noted that much of the knowledge discussed in this section was reported before the massive development of genetic tools available today to work in vivo (transgenics, gene editing, viral delivery, conditional and cell-specific DNA recombination, etc.). Primary culture from transgenic and knock-out rodent models increased the possibilities to expand our knowledge on gene function in astrocytes.
5.2. Cell Lines
Cell lines are essential for the development of the cell biology field. The advantages of using cell lines are numerous: stable and reproducible preparations within certain passage numbers; cells are handled easily and in a short time they expand to large quantities, sparing numerous animals from being used in experimental research; and the cells are usually available in cell banks so there is a widespread use and it is easier to compare results worldwide. Several studies related to astrocytes have been conducted with cell lines, helping to unveil their cellular functions in the CNS [147,148,149,150,151]. As we will discuss, there are more limitations concerning the use of cell lines in terms of representation of brain astrocyte functions in vivo.
One of the most used cell lines for astrocyte research is C6 rat astroglioma cells. These cells were isolated from rat glioma induced by N-nitrosomethylurea with verified relative preservation of the expression of typical astrocyte markers such as GFAP, glutamate transporter, S100B and GS [152]. The expression of genes typical of brain tumors such as PDGFb, IGF-I and EGFR was also observed in C6 cells [153]. The ambiguous nature of these cells, sharing both astrocytic and tumoral origins, clearly limits the interpretation of data that could be relevant to astrocyte physiology, and that accounts for the fact that C6 are also used in cancer research for glioma and glioblastoma, including new potential drug testing.
The advantages and limitations of the cell lines must be carefully considered while planning the experiment design, because chromosome stability, cell growth, metabolism, oxidative stress resistance and susceptibility to cell death are impacted by the cancerous transformation [152,154]. These limitations may not be present for other types of cell culture listed below.
C6 cells are responsive to LPS and capable of inducing Il6, Tnf, GFAP, ciclo-oxigenase 2 (PTGS2/COX-2) and inducible nitric oxide synthase (NOS2), although at a very high concentration (10 μg/mL) [113]. In addition, the PD-inducing neurotoxin 1-methyl-4-phenyl 1,2,3,6 tetrahydropyridine (MPTP), or ferrous sulfate that induces oxidative stress, also increases proinflammatory molecules [114,155]. These studies were conducted by the same research group, and it will be interesting to verify the capacity of astrocytic cell lines to respond to and promote inflammatory reactivity in comparison to other models. In this direction, it was shown that Aβ25–35-amyloid peptide is able to induce the expression of proinflammatory molecules in C6 cells [127].
5.3. Immortalized Astrocytes (IAs)
To complete studies involving many treatments, replications and validations, in addition to combining results acquired over years, it is important to preserve the cells’ morphologic, proliferative and metabolic patterns between passages. This is usually difficult to achieve with primary cell culture, which presents limited cellular replication and capacity to preserve gene expression and morphology [152]. Immortalization is a suitable solution for this problem.
Cell immortalization can be performed through oncogenes expression, namely hTERT (human telomerase reverse transcriptase) and SV40 large T antigen; one promotes telomere extension during replication and the other interferes with cell-cycle control, respectively. Telomeres shortening during each cell-cycle is a consequence of the linear nature of chromosome endings, and hTERT efficiently replenish the lost DNA fragments, while large tumor antigen interacts and inactivates tumor suppressors p53 and retinoblastoma-protein (pRb) [115,156,157,158,159,160].
Since varied cell lines were generated by immortalization, it is important to observe the different strategies employed in each case, which can impact the choice for a particular astrocytic cell. Immortalization with SV40 large T antigen was achieved in cells from a human fetus brain in 1985. Unlike former experiments with SV40 infection [133] that provided cells with limited passage, the employed strategy consisted in transfecting origin defective SV40 DNA into glia cells, so that the virus would not replicate, which resulted in an increased ratio of replication and survival of the cells called SVG [156]. Similar results of immortalization were observed in other subsequent studies; for instance, HSC2/HSC6 cells using temperature-sensitive mutant of SV40 large T antigen [161], A735 cells also using SV40 large T antigen [162], NHA-E6/E7/hTERT cells using telomerase [163], NHA/TS cells using SV40 early region [164] and C8-D1A [165]. These models have been widely employed, including preclinical studies and BBB modeling [166,167,168]. It is worth noting that IAs from different species can be acquired through commercial platforms.
Another approach to generate immortalized cells involves obtaining a primary cell culture from transgenic animals designed for this purpose. Liu and colleagues were able to isolate immortalized astrocytes from murine optic nerves [169]. The transgenic strain expresses the temperature sensitive SV40 large T antigen mutant tsA58 under the class I MHC H2kb promoter, leading to ubiquitous oncogene expression upon IFNγ exposure [170]. For instance, astrocytes deficient in Abcd1-deficiency (X-linked adrenoleukodystrophy model) have been studied using this technology [171]. As will be discussed in the Section 7, these immortalization techniques can produce cells with similar and dissimilar characteristics compared to primary cell culture and, once again, this can impact interpretations and comparisons of the resulting experimental data.
IAs can replicate inflammatory responses observed with other in vitro models. A proteomic study compared the responses of immortalized human fetal astrocytes to high concentrations of IL1B, or TNF, or LPS, and concluded that several proteins involved in inflammation could be induced by these treatments but, unlike the cytokines, LPS failed to activate the canonical nuclear factor (NF)-κB pathway and induce IL6 [128]. The failure in IL6 induction by LPS is actually surprising considering primary cell culture and cell line data, and deserves further investigation. Cytokine treatment on murine IAs were also able to convert the cells to a neurotoxic phenotype that did not depend on inflammatory molecules released by the astrocytes [116], which reinforces the concept that these cells are sensitive to the proinflammatory milieu typical of neurodegenerative diseases. SV40 large T antigen transformed IAs from the hippocampus of 3xTg-AD and wild-type control mice were shown to display comparable features to primary astrocytes, except for diminished levels of GFAP and AQP4 [117]. Few inflammatory readouts were investigated in this study, but curiously NOS2 induction by LPS and TNF was present in these IAs, and astrocytes as a source of NO have been questioned before (see above). Overall, IAs represent a promising tool to conduct large-scale studies for neuroinflammation in combination with neurodegenerative disease models, although validation of cellular responses must be carefully evaluated.
5.4. iPSC-Derived Astrocytes
Takashi and Yamanaka showed that somatic cells could be reprogrammed to pluripotency by the forced expression of four transcription factors: Pou5f1/Oct-3/4, Sox2, cMyc and Klf4. This was achieved in complete differentiated fibroblasts, and the resulting cells were called induced pluripotent stem cells (iPSCs), which presented morphofunctional features typical of embryonic stem cells [172]. The method was successfully applied to human cells [173].
One of the first astrocyte cell culture obtained from human iPSCs (hiPSCs) was reported in 2011, when iPS(IMR90)-4 cells reprogrammed from an IMR90 fibroblast were subjected to differentiation, resulting in immature astrocytes [174]. More than 90% of these cells showed expression of GFAP and S100B after 24 weeks in culture. In addition, these cells could propagate calcium waves, uptake glutamate and promote synaptogenesis and BBB formation. In conclusion, these cells presented primary astrocyte culture characteristics that could be maintained for 180 days after several passages and expansion. The predifferentiation of hiPSCs into neuroepithelial cells was a key step to achieve astrocytic cells. In contrast, induced astrocytes (iAstrocytes) could also be directly obtained from fibroblasts through the expression of NFIA, NFIB and SOX9 transcription factors [175]. Several methods were reported to generate iAstrocytes through neural progenitor cells (NPC) like the pioneer methods [174,176,177,178,179]. Most methods demanded long culture protocols to generate homogeneous mature astrocytes [174,179], or required sorting to reduce heterogeneity [180,181]. More recently, cell culture media conditions provided the means to reduce the protocol timeline [182].
Due to technical improvements and popularization of hiPSCs generation, this approach has been applied to study neurodegenerative diseases [183] and drug testing [184] through the use of the patients’ cells to discover new treatments. In line with this strategy, patients’ hiPSC-derived astrocytes have received growing attention in the last decade, which is corroborated by many preclinical studies [185].
The possibility to explore astrocytes from human subjects with minimal invasivity (cell collection) allowed the cellular investigation of mechanisms in the same genetic background of the patient, which is very relevant for sporadic cases of neurodegenerative diseases. However, it is important to evaluate if epigenetic marks and gene expression are compatible with differentiated astrocytes and without residual molecular signatures from the parental cell type [186,187,188]. Although hiPSC-derived astrocytes represent several advantages for studying brain diseases, interpretation issues may arise depending on the differences between hiPSC lines that can impact growth properties, susceptibility to cell death, effects of long-term culturing and so on. These problems can be circumvented by the use of several age-matched controls and hiPSCs from different patient donors, or the use of controls generated through correction of the disease-associated mutation (see [189] for a discussion). The high costs and laborious procedures involved in iPSC generation and maintenance make it difficult to obtain representative samples from different patients. The CRISPR-Cas9 gene edition methodology allowed significant advances in terms of introducing or correcting mutation, avoiding genetic alterations caused by viral integration into the genome and other artifacts promoted by previous generation strategies [190]. Several companies and nonprofit organizations provide hiPSCs and hiPSC-derived astrocytes, alleviating research laboratories from the burden involved in hiPSC generation and maintenance.
iAstrocytes development has already produced notable results in the field of neuroinflammation [110]. Santo et al. described a method to generate astrocytes from human iPSCs and ESCs, yielding functional astrocytes that are inflammation responsive to cytokines with increased percentage of IL8- and IL6-positive cells, similarly to primary culture [191]. Stratification of MS patients into benign or progressive MS phenotypes allowed the study of iPSC-derived reactive astrocytes and the discovery of neuroprotective properties in inflammatory conditions related to the benign MS phenotype [192]. Another recent article also made use of co-culture assemblies of neurons and astrocytes derived from iPSCs, modeling tau-pathology that features tau hyperphosphorylation and misfolding, resulting in changed neuroinflammatory patterns [118]. iPSCs of a different source, in this case human peripheral blood mononuclear cells, have also been used to model ALS-astrocytes, which shows increased secretion of IL1B, TNF and IL6 [119]. Other examples involved the investigation of inflammatory disruptors of the BBB modeled with iPSC-derived cells, and drug testing against inflammation using human fibroblasts as source material to generate iAstrocytes [193,194,195].
5.5. 3D Culture and Bioprinting
The traditional culturing in flasks and plates presents an obvious limitation compared to in vivo: cells grown as monolayers do not replicate the microenvironment found in tissues [196]. It is well documented that many cell types present aberrant cell division rates, show flatter shape and lose their phenotypic identity when cultured in 2D [197]. The development of 3D culture methods circumvented this limitation. This involves assembling a natural or a synthetic polymeric scaffold jointly with the cell suspension. In addition, the employed material can simulate the architecture and soft fabric of the brain. The use of hydrogels in combination with ECM components promote important changes in cell behavior, and provide a molecular microenvironment more compatible with the tissue in vivo [198,199,200,201].
The 3D culturing technique can be further combined with bioprinting, and the hydrogel mixture to be mixed with cell suspension, in this case, is called bioink. Bioprinting improves the spatial distribution of biomaterial and cellular components, leading to the construction of a synthetic and complex living tissue. The bioprinting process is controlled and can produce constructs previously determined in size, thickness and shape [199,200,202]. Accordingly, 3D culturing and bioprinting increase the dimension of exploratory evaluations of cell–cell [189,190,203,204], cell–hydrogel [205] and cell–ECM interactions [206] in processes of cell migration [207], shape remodeling [122], differentiation [208,209], neuropathological paradigms [210,211] and so on, with greater chance of compatibility with in vivo conditions.
A 3D collagen hydrogel-based culture was successfully employed with astrocytes, thus generating a culture system that consistently generates less reactive state, resembling undamaged brain phenotypes. Upon TGFB treatment, these cells presented more ramification morphology and expressed molecules associated with gliosis (elevated GFAP, Aquaporin 4, CSPG, Vimentin and IL-6 secretion) [212]. Different materials have been systematically tested to evaluate in vitro conditions that mimic the natural resting state of astrocytes and microglia culture (reviewed in [213]). Although 3D culture is promising for modeling neuroinflammation and neurodegeneration, there is still a lack of systematic studies that compare neuroinflammatory readouts in 3D cultures versus canonical 2D. For instance, SARS-CoV-2 infection and AD has been investigated in 3D cultures [214,215], but neuroinflammatory readouts are not explored in general.
5.6. Neurospheres and Brain Organoids (Mini-Brain)
Neural stem cells (NSCs) have been isolated from the embryonic, neonatal and adult rodent CNS, and respond to the mitogenic signal from epidermal growth factor (EGF) and fibroblast growth factor 2 (FGF-2). These cells are capable of asymmetric division, maintaining the stem cell pool, while retaining the ability to differentiate into neurons and macroglia. NSCs from rodents and humans can be cultured in nonadherent conditions, developing spheroids called neurospheres. Neurospheres in adherent conditions and deprived from growth factors differentiate in neurons and astrocytes [80,216,217]. Neurospheres have been used to access cellular properties from progenitor and stem cells, such as proliferation, self-renewal capacity, and multipotency, while their capacity to generate astrocytes has been less explored. It has been argued that experiments involving neurospheres lack consistency between research groups, due to sensitivity to the culturing method used (for review see [218]). Nevertheless, neurospheres can be considered a good approach to study the development of neural cells.
Further improvements of human NSC culture methods allowed the evolution of a methodology called brain organoids (also known was mini-brains). The hiPSCs grown on feeder cells were cultured in nonadherent conditions, and without 3D scaffolds could generate laminated structures called human cortical spheroids that presented functional astrocytes and only excitatory neurons in a reproducible manner [124]. Brain organoids are self-organized structures that are successfully used to model brain diseases (reviewed in [219,220,221]). The Zika virus (ZIKV) infection is one example of mini-brain modeling for studying fetal virus infection, revealing the dynamics of infectivity in astrocytes [222]. Several methodologies have been developed to include technological upgrades from bioprinting, for instance the inclusion of vascular elements and the use of microfluidic devices [222,223,224,225,226]. Although much effort has been employed to use brain organoids in regenerative medicine, their use as disease models started a new chapter for studying the glia relationship with neuroinflammation. An important implementation is the incorporation of microglia in these organoids, which has been recently achieved [126,227,228,229].
Since the employment of mini-brain–containing microglia cells is quite recent, the reports of neuroinflammation are related to neuronal/macroglial components only. This is in agreement with studying astrocytes disconnected from microglia, which represents most studies conducted so far. Cairns et al. introduced a model of herpes simplex (HSV-1) infection in mini-brains that presented features of AD brains including amyloid plaque–like formations, gliosis, neuroinflammation and decreased functionality. In this study, gliosis and astrocyte reactivity was accessed by increased GFAP, vimentin, LCN2 and serpin family E member 3, while the proinflammatory profile was accessed by the upregulation of TNF, IL1B, IL6 and IFNG [125]. ZIKV infection in mini-brains also induced markers of neuroinflammation [222], which were also shown in a model of methamphetamine exposure that promoted astrogliosis and upregulation of GFAP, inflammasome NRLP1, IL6 and other genes related to inflammation [230]. Altogether, brain organoids offer an advanced platform for testing mechanisms involving astrocytes that are relevant to neuropathologies with inflammatory components.
6. Major Differences between Cell Cultures: Implications for Neuroinflammation Associated with Neurodegeneration
Primary astrocyte cell culture development not only paved the way for studying these cells, but also promoted the use of a widespread technique that is relatively easy to standardize in research laboratories. As mentioned, microglia and other cell-type contamination can lead to variable degrees of astrocyte reactivity that are problematic for results interpretation. In addition, the use of fetal calf serum presents the cells with many soluble molecules usually not present in the brain environment. These culturing techniques yield a reactive phenotype, unlike humans in vivo (see [195] for discussion). New technical advances in cell purification and synthetic medium formulation can overcome these limitations, although the classical culture methodology will still be used for screening analysis and preliminary data acquisition. This also holds true for cell lines, which are advantageous for large-scale analysis such as drug testing and RNAi or CRISPR-Cas9 screenings.
Regarding cell lines and IA, Galland et al. compared the C6 cell line, IA and primary astrocyte culture, suggesting that C6 and IA show different morphology and biochemical activity compared to primary astrocytes [152]. The authors observed a decrease in astrocyte markers such as GFAP, S100B, ALDH1L and AQP4, in addition to a reduced glutamate metabolism in IA cells, indicating an inferior profile when compared to the other cells [115,158,231,232,233,234]. Although these striking differences are present in cell lines, immortalized and C6 cells preserve cell shape plasticity and glucose consumption in response to glutamate [152]. iAstrocytes represent a new chapter in astrocyte in vitro studies, and it is important to compare treatment and disease modeling outcomes with traditional models, as has been done by some groups. So far, the possibilities offered by this technology are more prevalent than the drawbacks.
Both primary astrocytes, cell lines, IA and iAstrocytes may benefit from 3D culturing, which can promote a resting state that is similar to the natural microenvironment in the undisturbed brain. Co-cultures of iPSC-derived neural cells also seem promising for modeling diseases and BBB function. The complexity of varied cell populations in the same system pose a problem similar to studies in vivo, which is the evaluation of individual cell-type responses and contributions. Since neuroinflammation is quite susceptible to the influence of microenvironment modifications promoted by the different neural cells, the gain of reproducing the natural context demands comes with the cost of sophisticated analysis application. Cell sorting, single-cell RNA sequencing and other single-cell approaches are being used to analyze astrocytes responses and generate detailed information of subpopulations and heterogeneity with unprecedented cutting-edge technologies.
7. Conclusions and Future Directions
Cell lines and IAs are important tools in astrocyte research, especially those involving complex molecular/genetic manipulations. One advantage of these cells is their capacity to rapidly expand cellular material and apply large-scale analysis. However, experimental interpretations are quite limited compared to primary cultures, since astrocytes features can be compromised by cell transformation and modify neuroinflammatory responses to a large extent. Also, rodent 2D culturing can be very limited in terms of providing pathological mechanisms, excluding those that are conserved and cell-autonomous, since cells are grown as monolayer and present initial degrees of reactivity, and these are limited to simpler interactions. Co-cultures may circumvent the lack of cell diversity interaction and offer a middle ground compared to more elaborated 3D cell cultures, which recapitulates natural astrocyte behavior. iAstrocytes represent a new frontier to model neurodegenerative diseases, and neuroinflammation research is gathering important information for both protective or detrimental effects from astrocytes reactivity. Organoids combined with iPSC technology and gene-editing tools represent the highest level of neurodegenerative disease modeling. The combination of these different in vitro resources will be essential to validate findings and evaluate cellular responses individually.
Author Contributions
A.M.P.: conception, writing and revising of the work; E.d.S.P.: conception, writing and revising of the work; T.R.R.: conception, writing and revising of the work; I.G.: conception, writing and revising of the work. All authors have read and agreed to the published version of the manuscript.
Funding
I.G. is a member of CEPID Redoxoma (Fundação de Amparo à Pesquisa do Estado de São Paulo; FAPESP 2013/07937-8). A.M.P. and E.d.S.P. are supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that the literature selection, opinion statements and conclusions, were conducted in the absence of any commercial or financial relationships that could be characterized as potential conflicts of interest.
References
- Valentin, G. Über Den Verlauf und Die Letzten Enden Der Nerven; Gedruckt bei Grass, Barth und Comp.: Breslau, Poland, 1836; Volume 18. [Google Scholar]
- Deiters, O.; Schultze, M.J.S. Untersuchungen Über Gehirn und Rückenmark Des Menschen und Der Säugethiere; F. Veiweg: Braunschweig, Germany, 1865. [Google Scholar]
- Gesammelte Abhandlungen zur Wissenschaftlichen Medicin/Von Rudolf Virchow. Available online: https://wellcomecollection.org/works/m3tp5x6w (accessed on 28 December 2023).
- Khakh, B.S.; Deneen, B. The Emerging Nature of Astrocyte Diversity. Annu. Rev. Neurosci. 2019, 42, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Lyons, D.A. Glia as Architects of Central Nervous System Formation and Function. Science 2018, 362, 181–185. [Google Scholar] [CrossRef]
- von Lenhossék, M. Zur Kenntnis der Neuroglia des menschlichen Rückenmarkes. Verh. Anat. Ges. 1891, 5, 193–221. [Google Scholar]
- Robertson, W. On a New Method of Obtaining a Black Reaction in Certain Tissue-Elements of the Central Nervous System. Scott. Med. Surg. J. 1899, 4, 23. [Google Scholar]
- Río-Hortega, P. del Son Homologables La Glía de Escasas Radiaciones y La Célula de Schwann. Boletıín Soc. Esp. Biol. 1922, 10, 25–28. [Google Scholar]
- Penfield, W. Oligodendroglia and Its Relation to Classical Neuroglia. Brain 1924, 47, 430–452. [Google Scholar] [CrossRef]
- Geren, B.B. The Formation from the Schwann Cell Surface of Myelin in the Peripheral Nerves of Chick Embryos. Exp. Cell Res. 1954, 7, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Bunge, M.B.; Bunge, R.P.; Pappas, G.D. Electron Microscopic Demonstration of Connections between Glia and Myelin Sheaths in the Developing Mammalian Central Nervous System. J. Cell Biol. 1962, 12, 448–453. [Google Scholar] [CrossRef]
- Río-Hortega, P.D.; Penfield, W. Cerebral Cicatrix—The Reaction of Neuroglia and Microglia to Brain Wounds. Johns Hopkins Hosp. Bull. 1927, 41, 278–303. [Google Scholar]
- Lugaro, E. Sulle Funzioni Della Nevroglia. Riv. Pat. Nerv. Ment. 1907, 12, 225–233. [Google Scholar]
- Nageotte, J. Pheénomenès de Sécrétion Dans Le Protoplasma Des Cellules Neévrogliques de La Substance Grise. CR Soc. Biol. 1910, 68, 1068–1069. [Google Scholar]
- Parpura, V.; Basarsky, T.A.; Liu, F.; Jeftinija, K.; Jeftinija, S.; Haydon, P.G. Glutamate-Mediated Astrocyte-Neuron Signalling. Nature 1994, 369, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite Synapses: Glia, the Unacknowledged Partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Cajal, S.R. Algunas Conjeturas Sobre El Mecanismo Anatómico de La Ideación, Asociación y Atención. Rev. Med. Cirug. Pract. 1895, 36, 497–508. [Google Scholar]
- Cajal, S.R. Algo Sobre La Significacion Fisiologica de La Consejos Acerca de La Tecnica Del Oro-Sublimado. Trab. Lab. Investig. Biol. Neurogl. Rev. Trim. Microg. 1897, 33–47. [Google Scholar]
- Cajal, S.R. El Encéfalo de Los Reptiles. Trab. Lab. Histol. Fac. Zarag. 1891, 24, 1–31. [Google Scholar]
- Parpura, V.; Verkhratsky, A. Neuroglia at the Crossroads of Homoeostasis, Metabolism and Signalling: Evolution of the Concept. ASN Neuro 2012, 4, e00087. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Kummer, M.P.; Latz, E. Innate Immune Activation in Neurodegenerative Disease. Nat. Rev. Immunol. 2014, 14, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xiao, D.; Mao, Q.; Xia, H. Role of Neuroinflammation in Neurodegeneration Development. Signal Transduct. Target. Ther. 2023, 8, 1–32. [Google Scholar] [CrossRef]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms Underlying Inflammation in Neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef]
- Golgi, C. Sulla Fina Struttura Dei Bulbi Olfattorii; Tipografia di Stefano Calderini: Reggio nell’Emilia, Italy, 1875. [Google Scholar]
- HIs, W. Zur Geschichte des Gehirns Sowie der Centralen und Peripherischen Nervenbahnen Beim Menschlichen Embryo; Königlich Sächsische Gesellschaft der Wissenschaften: Leipzig, Germany, 1888; Volume 24, pp. 341–392. [Google Scholar]
- Cajal, S.R. La Rétine Des Vertébrés. La Cellule 1893, 9, 119–257. [Google Scholar]
- Tessier-Lavigne, M.; Goodman, C.S. The Molecular Biology of Axon Guidance. Science 1996, 274, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Bentivoglio, M.; Mazzarello, P. The History of Radial Glia. Brain Res. Bull. 1999, 49, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, C. The Chemotactic Hypothesis of Cajal: A Century Behind. Prog. Brain Res. 2002, 136, 11–20. [Google Scholar] [CrossRef]
- Hösli, L.; Zuend, M.; Bredell, G.; Zanker, H.S.; Oliveira, C.E.P.D.; Saab, A.S.; Weber, B. Direct Vascular Contact Is a Hallmark of Cerebral Astrocytes. Cell Rep. 2022, 39. [Google Scholar] [CrossRef]
- Bartheld, C.S.V.; Bahney, J.; Herculano-Houzel, S. The Search for True Numbers of Neurons and Glial Cells in the Human Brain: A Review of 150 Years of Cell Counting. J. Comp. Neurol. 2016, 524, 3865–3895. [Google Scholar] [CrossRef]
- Bahney, J.; von Bartheld, C.S. The Cellular Composition and Glia–Neuron Ratio in the Spinal Cord of a Human and a Nonhuman Primate: Comparison With Other Species and Brain Regions. Anat. Rec. 2018, 301, 697–710. [Google Scholar] [CrossRef]
- Sherwood, C.C.; Stimpson, C.D.; Raghanti, M.A.; Wildman, D.E.; Uddin, M.; Grossman, L.I.; Goodman, M.; Redmond, J.C.; Bonar, C.J.; Erwin, J.M.; et al. Evolution of Increased Glia–Neuron Ratios in the Human Frontal Cortex. Proc. Natl. Acad. Sci. USA 2006, 103, 13606–13611. [Google Scholar] [CrossRef]
- Ruiz-Sauri, A.; Orduña-Valls, J.M.; Blasco-Serra, A.; Tornero-Tornero, C.; Cedeño, D.L.; Bejarano-Quisoboni, D.; Valverde-Navarro, A.A.; Benyamin, R.; Vallejo, R. Glia to Neuron Ratio in the Posterior Aspect of the Human Spinal Cord at Thoracic Segments Relevant to Spinal Cord Stimulation. J. Anat. 2019, 235, 997–1006. [Google Scholar] [CrossRef]
- Lovick, T.A.; Brown, L.A.; Key, B.J. Neuronal Activity-Related Coupling in Cortical Arterioles: Involvement of Astrocyte-Derived Factors. Exp. Physiol. 2005, 90, 131–140. [Google Scholar] [CrossRef]
- Shigetomi, E.; Kracun, S.; Sofroniew, M.V.; Khakh, B.S. A Genetically Targeted Optical Sensor to Monitor Calcium Signals in Astrocyte Processes. Nat. Neurosci. 2010, 13, 759–766. [Google Scholar] [CrossRef]
- Di Castro, M.A.; Chuquet, J.; Liaudet, N.; Bhaukaurally, K.; Santello, M.; Bouvier, D.; Tiret, P.; Volterra, A. Local Ca2+ Detection and Modulation of Synaptic Release by Astrocytes. Nat. Neurosci. 2011, 14, 1276–1284. [Google Scholar] [CrossRef]
- Nett, W.J.; Oloff, S.H.; McCarthy, K.D. Hippocampal Astrocytes in Situ Exhibit Calcium Oscillations That Occur Independent of Neuronal Activity. J. Neurophysiol. 2002, 87, 528–537. [Google Scholar] [CrossRef]
- De Bock, M.; Decrock, E.; Wang, N.; Bol, M.; Vinken, M.; Bultynck, G.; Leybaert, L. The Dual Face of Connexin-Based Astroglial Ca2+ Communication: A Key Player in Brain Physiology and a Prime Target in Pathology. Biochim. Biophys. Acta 2014, 1843, 2211–2232. [Google Scholar] [CrossRef]
- Allen, N.J.; Barres, B.A. Glia—More than Just Brain Glue. Nature 2009, 457, 675–677. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and Pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from Brain Glue to Communication Elements: The Revolution Continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Nedergaard, M. Astroglial Cradle in the Life of the Synapse. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130595. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef] [PubMed]
- Kanemaru, K.; Sekiya, H.; Xu, M.; Satoh, K.; Kitajima, N.; Yoshida, K.; Okubo, Y.; Sasaki, T.; Moritoh, S.; Hasuwa, H.; et al. In Vivo Visualization of Subtle, Transient, and Local Activity of Astrocytes Using an Ultrasensitive Ca2+ Indicator. Cell Rep. 2014, 8, 311–318. [Google Scholar] [CrossRef]
- Mogensen, F.L.-H.; Delle, C.; Nedergaard, M. The Glymphatic System (En)during Inflammation. Int. J. Mol. Sci. 2021, 22, 7491. [Google Scholar] [CrossRef] [PubMed]
- Weller, R.O.; Djuanda, E.; Yow, H.-Y.; Carare, R.O. Lymphatic Drainage of the Brain and the Pathophysiology of Neurological Disease. Acta Neuropathol. 2009, 117, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [PubMed]
- Jessen, N.A.; Munk, A.S.F.; Lundgaard, I.; Nedergaard, M. The Glymphatic System: A Beginner’s Guide. Neurochem. Res. 2015, 40, 2583–2599. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Asan, E.; Püschel, B.; Kugler, P. Cellular and Regional Distribution of the Glutamate Transporter GLAST in the CNS of Rats: Nonradioactive In Situ Hybridization and Comparative Immunocytochemistry. J. Neurosci. 1997, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.M.; Sullivan, R.K.P.; Scott, H.L.; Finkelstein, D.I.; Colditz, P.B.; Lingwood, B.E.; Dodd, P.R.; Pow, D.V. Glial Glutamate Transporter Expression Patterns in Brains from Multiple Mammalian Species. Glia 2005, 49, 520–541. [Google Scholar] [CrossRef] [PubMed]
- Anlauf, E.; Derouiche, A. Glutamine Synthetase as an Astrocytic Marker: Its Cell Type and Vesicle Localization. Front. Endocrinol. 2013, 4. [Google Scholar] [CrossRef]
- Ogata, K.; Kosaka, T. Structural and Quantitative Analysis of Astrocytes in the Mouse Hippocampus. Neuroscience 2002, 113, 221–233. [Google Scholar] [CrossRef]
- Hachem, S.; Aguirre, A.; Vives, V.; Marks, A.; Gallo, V.; Legraverend, C. Spatial and temporal expression of S100B in cells of oligodendrocyte lineage. Glia 2005, 51, 81–97. [Google Scholar] [CrossRef]
- Steiner, J.; Bernstein, H.-G.; Bielau, H.; Berndt, A.; Brisch, R.; Mawrin, C.; Keilhoff, G.; Bogerts, B. Evidence for a Wide Extra-Astrocytic Distribution of S100B in Human Brain. BMC Neurosci. 2007, 8, 2. [Google Scholar] [CrossRef]
- Cammer, W. Glutamine Synthetase in the Central Nervous System Is Not Confined to Astrocytes. J. Neuroimmunol. 1990, 26, 173–178. [Google Scholar] [CrossRef] [PubMed]
- D’Amelio, F.; Eng, L.F.; Gibbs, M.A. Glutamine Synthetase Immunoreactivity Is Present in Oligodendroglia of Various Regions of the Central Nervous System. Glia 1990, 3, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Asan, E.; Lesch, K.-P.; Kugler, P. A Splice Variant of Glutamate Transporter GLT1/EAAT2 Expressed in Neurons: Cloning and Localization in Rat Nervous System. Neuroscience 2002, 109, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Nagy, J.I.; Patel, D.; Ochalski, P.A.Y.; Stelmack, G.L. Connexin30 in Rodent, Cat and Human Brain: Selective Expression in Gray Matter Astrocytes, Co-Localization with Connexin43 at Gap Junctions and Late Developmental Appearance. Neuroscience 1999, 88, 447–468. [Google Scholar] [CrossRef] [PubMed]
- Hol, E.M.; Pekny, M. Glial Fibrillary Acidic Protein (GFAP) and the Astrocyte Intermediate Filament System in Diseases of the Central Nervous System. Curr. Opin. Cell Biol. 2015, 32, 121–130. [Google Scholar] [CrossRef]
- Eng, L.F.; Ghirnikar, R.S.; Lee, Y.L. Glial Fibrillary Acidic Protein: GFAP-Thirty-One Years (1969–2000). Neurochem. Res. 2000, 25, 1439–1451. [Google Scholar] [CrossRef]
- Du, J.; Yi, M.; Zhou, F.; He, W.; Yang, A.; Qiu, M.; Huang, H. S100B Is Selectively Expressed by Gray Matter Protoplasmic Astrocytes and Myelinating Oligodendrocytes in the Developing CNS. Mol. Brain 2021, 14, 154. [Google Scholar] [CrossRef]
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.; Qu, Z.; Hashimoto, H.; Gradinaru, V.; Kawaguchi, R.; Khakh, B.S. Molecular Basis of Astrocyte Diversity and Morphology across the CNS in Health and Disease. Science 2022, 378, eadc9020. [Google Scholar] [CrossRef]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive Astrocyte Nomenclature, Definitions, and Future Directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Wilson, C.L.; Hayward, S.L.; Kidambi, S. Astrogliosis in a Dish: Substrate Stiffness Induces Astrogliosis in Primary Rat Astrocytes. RSC Adv. 2016, 6, 34447–34457. [Google Scholar] [CrossRef]
- Yu, P.; Wang, H.; Katagiri, Y.; Geller, H.M. An In Vitro Model of Reactive Astrogliosis and Its Effect on Neuronal Growth. In Astrocytes: Methods and Protocols; Milner, R., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; pp. 327–340. ISBN 978-1-61779-452-0. [Google Scholar]
- Badia-Soteras, A.; de Vries, J.; Dykstra, W.; Broersen, L.M.; Verkuyl, J.M.; Smit, A.B.; Verheijen, M.H.G. High-Throughput Analysis of Astrocyte Cultures Shows Prevention of Reactive Astrogliosis by the Multi-Nutrient Combination Fortasyn Connect. Cells 2022, 11, 1428. [Google Scholar] [CrossRef]
- Cullen, D.K.; Simon, C.M.; LaPlaca, M.C. Strain Rate-Dependent Induction of Reactive Astrogliosis and Cell Death in Three-Dimensional Neuronal-Astrocytic Co-Cultures. Brain Res. 2007, 1158, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Pogoda, K.; Chin, L.; Georges, P.C.; Byfield, F.J.; Bucki, R.; Kim, R.; Weaver, M.; Wells, R.G.; Marcinkiewicz, C.; Janmey, P.A. Compression Stiffening of Brain and Its Effect on Mechanosensing by Glioma Cells. New J. Phys. 2014, 16, 075002. [Google Scholar] [CrossRef]
- Pekny, M.; Nilsson, M. Astrocyte Activation and Reactive Gliosis. Glia 2005, 50, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Bellaver, B.; Rocha, A.S.; Souza, D.G.; Leffa, D.T.; De Bastiani, M.A.; Schu, G.; Lukasewicz Ferreira, P.C.; Venturin, G.T.; Greggio, S.; Ribeiro, C.T.; et al. Activated Peripheral Blood Mononuclear Cell Mediators Trigger Astrocyte Reactivity. Brain. Behav. Immun. 2019, 80, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, J.; Chen, Y.; Collier, J.M.; Capuk, O.; Jin, S.; Sun, M.; Mondal, S.K.; Whiteside, T.L.; Stolz, D.B.; et al. NOX Activation in Reactive Astrocytes Regulates Astrocytic LCN2 Expression and Neurodegeneration. Cell Death Dis. 2022, 13, 371. [Google Scholar] [CrossRef]
- Fan, Y.-Y.; Huo, J. A1/A2 Astrocytes in Central Nervous System Injuries and Diseases: Angels or Devils? Neurochem. Int. 2021, 148, 105080. [Google Scholar] [CrossRef]
- Bretheau, F.; Castellanos-Molina, A.; Bélanger, D.; Kusik, M.; Mailhot, B.; Boisvert, A.; Vallières, N.; Lessard, M.; Gunzer, M.; Liu, X.; et al. The Alarmin Interleukin-1α Triggers Secondary Degeneration through Reactive Astrocytes and Endothelium after Spinal Cord Injury. Nat. Commun. 2022, 13, 5786. [Google Scholar] [CrossRef]
- Lawrence, J.M.; Schardien, K.; Wigdahl, B.; Nonnemacher, M.R. Roles of Neuropathology-Associated Reactive Astrocytes: A Systematic Review. Acta Neuropathol. Commun. 2023, 11, 42. [Google Scholar] [CrossRef]
- Sekar, S.; McDonald, J.; Cuyugan, L.; Aldrich, J.; Kurdoglu, A.; Adkins, J.; Serrano, G.; Beach, T.G.; Craig, D.W.; Valla, J.; et al. Alzheimer’s Disease Is Associated with Altered Expression of Genes Involved in Immune Response and Mitochondrial Processes in Astrocytes. Neurobiol. Aging 2015, 36, 583–591. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in Neurodegenerative Disorders: The Roles of Microglia and Astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Garwood, C.J.; Pooler, A.M.; Atherton, J.; Hanger, D.P.; Noble, W. Astrocytes Are Important Mediators of Aβ-Induced Neurotoxicity and Tau Phosphorylation in Primary Culture. Cell Death Dis. 2011, 2, e167. [Google Scholar] [CrossRef]
- Bi, F.; Huang, C.; Tong, J.; Qiu, G.; Huang, B.; Wu, Q.; Li, F.; Xu, Z.; Bowser, R.; Xia, X.-G.; et al. Reactive Astrocytes Secrete Lcn2 to Promote Neuron Death. Proc. Natl. Acad. Sci. USA 2013, 110, 4069–4074. [Google Scholar] [CrossRef]
- Caldwell, A.L.M.; Sancho, L.; Deng, J.; Bosworth, A.; Miglietta, A.; Diedrich, J.K.; Shokhirev, M.N.; Allen, N.J. Aberrant Astrocyte Protein Secretion Contributes to Altered Neuronal Development in Multiple Models of Neurodevelopmental Disorders. Nat. Neurosci. 2022, 25, 1163–1178. [Google Scholar] [CrossRef]
- Ding, Z.-B.; Song, L.-J.; Wang, Q.; Kumar, G.; Yan, Y.-Q.; Ma, C.-G. Astrocytes: A Double-Edged Sword in Neurodegenerative Diseases. Neural Regen. Res. 2021, 16, 1702–1710. [Google Scholar] [CrossRef]
- Brandebura, A.N.; Paumier, A.; Onur, T.S.; Allen, N.J. Astrocyte Contribution to Dysfunction, Risk and Progression in Neurodegenerative Disorders. Nat. Rev. Neurosci. 2023, 24, 23–39. [Google Scholar] [CrossRef]
- Edison, P. Astroglial Activation: Current Concepts and Future Directions. Alzheimers Dement. J. Alzheimers Assoc. 2024. [Google Scholar] [CrossRef] [PubMed]
- Glezer, I.; Zekki, H.; Scavone, C.; Rivest, S. Modulation of the Innate Immune Response by NMDA Receptors Has Neuropathological Consequences. J. Neurosci. 2003, 23, 11094–11103. [Google Scholar] [CrossRef] [PubMed]
- Lehnardt, S.; Lachance, C.; Patrizi, S.; Lefebvre, S.; Follett, P.L.; Jensen, F.E.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. The Toll-Like Receptor TLR4 Is Necessary for Lipopolysaccharide-Induced Oligodendrocyte Injury in the CNS. J. Neurosci. 2002, 22, 2478–2486. [Google Scholar] [CrossRef] [PubMed]
- Saura, J. Microglial Cells in Astroglial Cultures: A Cautionary Note. J. Neuroinflamm. 2007, 4, 26. [Google Scholar] [CrossRef]
- Gorina, R.; Font-Nieves, M.; Márquez-Kisinousky, L.; Santalucia, T.; Planas, A.M. Astrocyte TLR4 Activation Induces a Proinflammatory Environment through the Interplay between MyD88-Dependent NFκB Signaling, MAPK, and Jak1/Stat1 Pathways. Glia 2011, 59, 242–255. [Google Scholar] [CrossRef]
- Kumamaru, H.; Saiwai, H.; Kobayakawa, K.; Kubota, K.; van Rooijen, N.; Inoue, K.; Iwamoto, Y.; Okada, S. Liposomal Clodronate Selectively Eliminates Microglia from Primary Astrocyte Cultures. J. Neuroinflamm. 2012, 9, 116. [Google Scholar] [CrossRef]
- Hupp, S.; Iliev, A.I. CSF-1 Receptor Inhibition as a Highly Effective Tool for Depletion of Microglia in Mixed Glial Cultures. J. Neurosci. Methods 2020, 332, 108537. [Google Scholar] [CrossRef]
- Van Zeller, M.; Sebastião, A.M.; Valente, C.A. Microglia Depletion from Primary Glial Cultures Enables to Accurately Address the Immune Response of Astrocytes. Biomolecules 2022, 12, 666. [Google Scholar] [CrossRef] [PubMed]
- Bohlen, C.J.; Bennett, F.C.; Tucker, A.F.; Collins, H.Y.; Mulinyawe, S.B.; Barres, B.A. Diverse Requirements for Microglial Survival, Specification, and Function Revealed by Defined-Medium Cultures. Neuron 2017, 94, 759–773.e8. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes Are Induced by Activated Microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, N.J.; Benveniste, E.N. Interleukin-6 Expression and Regulation in Astrocytes. J. Neuroimmunol. 1999, 100, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Glezer, I.; Chernomoretz, A.; David, S.; Plante, M.-M.; Rivest, S. Genes Involved in the Balance between Neuronal Survival and Death during Inflammation. PLoS ONE 2007, 2, e310. [Google Scholar] [CrossRef]
- Glezer, I.; Rivest, S. Oncostatin M Is a Novel Glucocorticoid-Dependent Neuroinflammatory Factor That Enhances Oligodendrocyte Precursor Cell Activity in Demyelinated Sites. Brain Behav. Immun. 2010, 24, 695–704. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 Paradigm of Macrophage Activation: Time for Reassessment. F1000prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef]
- Datta Chaudhuri, A.; Dasgheyb, R.M.; DeVine, L.R.; Bi, H.; Cole, R.N.; Haughey, N.J. Stimulus-Dependent Modifications in Astrocyte-Derived Extracellular Vesicle Cargo Regulate Neuronal Excitability. Glia 2020, 68, 128–144. [Google Scholar] [CrossRef]
- Chaudhuri, A.D.; Dastgheyb, R.M.; Yoo, S.-W.; Trout, A.; Talbot, C.C.; Hao, H.; Witwer, K.W.; Haughey, N.J. TNFα and IL-1β Modify the miRNA Cargo of Astrocyte Shed Extracellular Vesicles to Regulate Neurotrophic Signaling in Neurons. Cell Death Dis. 2018, 9, 363. [Google Scholar] [CrossRef]
- Yu, T.; Wang, X.; Zhi, T.; Zhang, J.; Wang, Y.; Nie, E.; Zhou, F.; You, Y.; Liu, N. Delivery of MGMT mRNA to Glioma Cells by Reactive Astrocyte-Derived Exosomes Confers a Temozolomide Resistance Phenotype. Cancer Lett. 2018, 433, 210–220. [Google Scholar] [CrossRef]
- Freeman, L.; Guo, H.; David, C.N.; Brickey, W.J.; Jha, S.; Ting, J.P.-Y. NLR Members NLRC4 and NLRP3 Mediate Sterile Inflammasome Activation in Microglia and Astrocytes. J. Exp. Med. 2017, 214, 1351–1370. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, J.; de Rivero Vaccari, J.P.; Keane, R.W. Human Astrocytes Express a Novel NLRP2 Inflammasome. Glia 2013, 61, 1113–1121. [Google Scholar] [CrossRef]
- Liu, H.-D.; Li, W.; Chen, Z.-R.; Hu, Y.-C.; Zhang, D.-D.; Shen, W.; Zhou, M.-L.; Zhu, L.; Hang, C.-H. Expression of the NLRP3 Inflammasome in Cerebral Cortex After Traumatic Brain Injury in a Rat Model. Neurochem. Res. 2013, 38, 2072–2083. [Google Scholar] [CrossRef] [PubMed]
- Johann, S.; Heitzer, M.; Kanagaratnam, M.; Goswami, A.; Rizo, T.; Weis, J.; Troost, D.; Beyer, C. NLRP3 Inflammasome Is Expressed by Astrocytes in the SOD1 Mouse Model of ALS and in Human Sporadic ALS Patients. Glia 2015, 63, 2260–2273. [Google Scholar] [CrossRef]
- Couturier, J.; Stancu, I.-C.; Schakman, O.; Pierrot, N.; Huaux, F.; Kienlen-Campard, P.; Dewachter, I.; Octave, J.-N. Activation of Phagocytic Activity in Astrocytes by Reduced Expression of the Inflammasome Component ASC and Its Implication in a Mouse Model of Alzheimer Disease. J. Neuroinflamm. 2016, 13, 20. [Google Scholar] [CrossRef]
- Choi, J.-K.; Park, S.-Y.; Kim, K.H.; Park, S.R.; Lee, S.-G.; Choi, B.H. GM-CSF Reduces Expression of Chondroitin Sulfate Proteoglycan (CSPG) Core Proteins in TGF-β-Treated Primary Astrocytes. BMB Rep. 2014, 47, 679–684. [Google Scholar] [CrossRef]
- Schachtrup, C.; Ryu, J.K.; Helmrick, M.J.; Vagena, E.; Galanakis, D.K.; Degen, J.L.; Margolis, R.U.; Akassoglou, K. Fibrinogen Triggers Astrocyte Scar Formation by Promoting the Availability of Active TGF-β after Vascular Damage. J. Neurosci. 2010, 30, 5843–5854. [Google Scholar] [CrossRef] [PubMed]
- Gris, P.; Tighe, A.; Levin, D.; Sharma, R.; Brown, A. Transcriptional Regulation of Scar Gene Expression in Primary Astrocytes. Glia 2007, 55, 1145–1155. [Google Scholar] [CrossRef]
- Klemens, J.; Ciurkiewicz, M.; Chludzinski, E.; Iseringhausen, M.; Klotz, D.; Pfankuche, V.M.; Ulrich, R.; Herder, V.; Puff, C.; Baumgärtner, W.; et al. Neurotoxic Potential of Reactive Astrocytes in Canine Distemper Demyelinating Leukoencephalitis. Sci. Rep. 2019, 9, 11689. [Google Scholar] [CrossRef]
- Ren, Z.; Iliff, J.J.; Yang, L.; Yang, J.; Chen, X.; Chen, M.J.; Giese, R.N.; Wang, B.; Shi, X.; Nedergaard, M. ‘Hit & Run’ Model of Closed-Skull Traumatic Brain Injury (TBI) Reveals Complex Patterns of Post-Traumatic AQP4 Dysregulation. J. Cereb. Blood Flow Metab. 2013, 33, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, M.; Lehtonen, S.; Jaronen, M.; Goldsteins, G.; Hämäläinen, R.H.; Koistinaho, J. Astrocyte Alterations in Neurodegenerative Pathologies and Their Modeling in Human Induced Pluripotent Stem Cell Platforms. Cell. Mol. Life Sci. 2019, 76, 2739–2760. [Google Scholar] [CrossRef] [PubMed]
- Prah, J.; Winters, A.; Chaudhari, K.; Hersh, J.; Liu, R.; Yang, S.-H. A Novel Serum Free Primary Astrocyte Culture Method That Mimic Quiescent Astrocyte Phenotype. J. Neurosci. Methods 2019, 320, 50–63. [Google Scholar] [CrossRef]
- Taylor, X.; Cisternas, P.; You, Y.; You, Y.; Xiang, S.; Marambio, Y.; Zhang, J.; Vidal, R.; Lasagna-Reeves, C.A. A1 Reactive Astrocytes and a Loss of TREM2 Are Associated with an Early Stage of Pathology in a Mouse Model of Cerebral Amyloid Angiopathy. J. Neuroinflamm. 2020, 17, 223. [Google Scholar] [CrossRef] [PubMed]
- Niranjan, R.; Nath, C.; Shukla, R. Melatonin Attenuated Mediators of Neuroinflammation and Alpha-7 Nicotinic Acetylcholine Receptor mRNA Expression in Lipopolysaccharide (LPS) Stimulated Rat Astrocytoma Cells, C6. Free Radic. Res. 2012, 46, 1167–1177. [Google Scholar] [CrossRef]
- Niranjan, R.; Nath, C.; Shukla, R. The Mechanism of Action of MPTP-Induced Neuroinflammation and Its Modulation by Melatonin in Rat Astrocytoma Cells, C6. Free Radic. Res. 2010, 44, 1304–1316. [Google Scholar] [CrossRef]
- Furihata, T.; Ito, R.; Kamiichi, A.; Saito, K.; Chiba, K. Establishment and Characterization of a New Conditionally Immortalized Human Astrocyte Cell Line. J. Neurochem. 2016, 136, 92–105. [Google Scholar] [CrossRef]
- Efremova, L.; Chovancova, P.; Adam, M.; Gutbier, S.; Schildknecht, S.; Leist, M. Switching from Astrocytic Neuroprotection to Neurodegeneration by Cytokine Stimulation. Arch. Toxicol. 2017, 91, 231–246. [Google Scholar] [CrossRef]
- Rocchio, F.; Tapella, L.; Manfredi, M.; Chisari, M.; Ronco, F.; Ruffinatti, F.A.; Conte, E.; Canonico, P.L.; Sortino, M.A.; Grilli, M.; et al. Correction: Gene Expression, Proteome and Calcium Signaling Alterations in Immortalized Hippocampal Astrocytes from an Alzheimer’s Disease Mouse Model. Cell Death Dis. 2020, 11, 1–2. [Google Scholar] [CrossRef]
- Rickner, H.D.; Jiang, L.; Hong, R.; O’Neill, N.K.; Mojica, C.A.; Snyder, B.J.; Zhang, L.; Shaw, D.; Medalla, M.; Wolozin, B.; et al. Single Cell Transcriptomic Profiling of a Neuron-Astrocyte Assembloid Tauopathy Model. Nat. Commun. 2022, 13, 6275. [Google Scholar] [CrossRef] [PubMed]
- BaofengFeng, U.; Amponsah, A.E.; Guo, R.; Liu, X.; Zhang, J.; Du, X.; Zhou, Z.; He, J.; Ma, J.; Cui, H. Autophagy-Mediated Inflammatory Cytokine Secretion in Sporadic ALS Patient iPSC-Derived Astrocytes. Oxid. Med. Cell. Longev. 2022, 2022, e6483582. [Google Scholar] [CrossRef] [PubMed]
- Kawatani, K.; Nambara, T.; Nawa, N.; Yoshimatsu, H.; Kusakabe, H.; Hirata, K.; Tanave, A.; Sumiyama, K.; Banno, K.; Taniguchi, H.; et al. A Human Isogenic iPSC-Derived Cell Line Panel Identifies Major Regulators of Aberrant Astrocyte Proliferation in Down Syndrome. Commun. Biol. 2021, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Coleman, H.A.; Meagher, L.; Forsythe, J.S.; Parkington, H.C. 3D Functional Neuronal Networks in Free-Standing Bioprinted Hydrogel Constructs. Adv. Healthc. Mater. 2023, 12, 2300801. [Google Scholar] [CrossRef] [PubMed]
- Placone, A.L.; McGuiggan, P.M.; Bergles, D.E.; Guerrero-Cazares, H.; Quiñones-Hinojosa, A.; Searson, P.C. Human Astrocytes Develop Physiological Morphology and Remain Quiescent in a Novel 3D Matrix. Biomaterials 2015, 42, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Gensini, L.; Serna, J.A.; Rubio, D.; Orozco, J.C.; Bolaños, N.I.; Cruz, J.C.; Muñoz-Camargo, C. Three-Dimensional Neuroimmune Co-Culture System for Modeling Parkinson’s Disease Microenvironmentsin Vitro. Biofabrication 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Paşca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.-Y.; O’Rourke, N.A.; Nguyen, K.D.; et al. Functional Cortical Neurons and Astrocytes from Human Pluripotent Stem Cells in 3D Culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Cairns, D.M.; Rouleau, N.; Parker, R.N.; Walsh, K.G.; Gehrke, L.; Kaplan, D.L. A 3D Human Brain-like Tissue Model of Herpes-Induced Alzheimer’s Disease. Sci. Adv. 2020, 6, eaay8828. [Google Scholar] [CrossRef]
- Schafer, S.T.; Mansour, A.A.; Schlachetzki, J.C.M.; Pena, M.; Ghassemzadeh, S.; Mitchell, L.; Mar, A.; Quang, D.; Stumpf, S.; Ortiz, I.S.; et al. An in Vivo Neuroimmune Organoid Model to Study Human Microglia Phenotypes. Cell 2023, 186, 2111–2126.e20. [Google Scholar] [CrossRef]
- Calvillo, M.; Diaz, A.; Limon, D.I.; Mayoral, M.A.; Chánez-Cárdenas, M.E.; Zenteno, E.; Montaño, L.F.; Guevara, J.; Espinosa, B. Amyloid-Β25–35 Induces a Permanent Phosphorylation of HSF-1, but a Transitory and Inflammation-Independent Overexpression of Hsp-70 in C6 Astrocytoma Cells. Neuropeptides 2013, 47, 339–346. [Google Scholar] [CrossRef]
- Dozio, V.; Sanchez, J.-C. Profiling the Proteomic Inflammatory State of Human Astrocytes Using DIA Mass Spectrometry. J. Neuroinflamm. 2018, 15, 331. [Google Scholar] [CrossRef]
- Millet, L.J.; Gillette, M.U. Over a Century of Neuron Culture: From the Hanging Drop to Microfluidic Devices. Yale J. Biol. Med. 2012, 85, 501–521. [Google Scholar] [PubMed]
- Hogue, M.J. Human Fetal Brain Cells in Tissue Cultures; Their Identification and Motility. J. Exp. Zool. 1947, 106, 85–107. [Google Scholar] [CrossRef] [PubMed]
- Kriegstein, A.; Alvarez-Buylla, A. The Glial Nature of Embryonic and Adult Neural Stem Cells. Annu. Rev. Neurosci. 2009, 32, 149–184. [Google Scholar] [CrossRef] [PubMed]
- Shein, H.M. Propagation of Human Fetal Spongioblasts and Astrocytes in Dispersed Cell Cultures. Exp. Cell Res. 1965, 40, 554–569. [Google Scholar] [CrossRef] [PubMed]
- Shein, H.M. Neoplastic Transformation of Hamster Astrocytes in Vitro by Simian Virus 40 and Polyoma Virus. Science 1968, 159, 1476–1477. [Google Scholar] [CrossRef] [PubMed]
- Raff, M.C.; Miller, R.H.; Noble, M. A Glial Progenitor Cell That Develops in Vitro into an Astrocyte or an Oligodendrocyte Depending on Culture Medium. Nature 1983, 303, 390–396. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.D.; de Vellis, J. Preparation of Separate Astroglial and Oligodendroglial Cell Cultures from Rat Cerebral Tissue. J. Cell Biol. 1980, 85, 890–902. [Google Scholar] [CrossRef]
- Booher, J.; Sensenbrenner, M. Growth and Cultivation of Dissociated Neurons and Glial Cells from Embryonic Chick, Rat and Human Brain in Flask Cultures. Neurobiology 1972, 2, 97–105. [Google Scholar]
- Vellis, J.D.; Cole, R. Preparation of Mixed Glial Cultures from Postnatal Rat Brain. Methods Mol. Biol. Clifton NJ 2012, 814, 49–59. [Google Scholar] [CrossRef]
- Bouvier, M.; Szatkowski, M.; Amato, A.; Attwell, D. The Glial Cell Glutamate Uptake Carrier Countertransports pH-Changing Anions. Nature 1992, 360, 471–474. [Google Scholar] [CrossRef]
- Kimelberg, H.K. Primary Astrocyte Cultures—A Key to Astrocyte Function. Cell. Mol. Neurobiol. 1983, 3, 1–16. [Google Scholar] [CrossRef]
- Schousboe, A.; Svenneby, G.; Hertz, L. Uptake and Metabolism of Glutamate in Astrocytes Cultured from Dissociated Mouse Brain Hemispheres. J. Neurochem. 1977, 29, 999–1005. [Google Scholar] [CrossRef]
- Friede, R.L. The Enzymatic Response of Astrocytes to Various Ions In Vitro. J. Cell Biol. 1964, 20, 5–15. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Glutamate Uptake into Astrocytes Stimulates Aerobic Glycolysis: A Mechanism Coupling Neuronal Activity to Glucose Utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef]
- Struckhoff, G. Cocultures of Meningeal and Astrocytic Cells—A Model for the Formation of the Glial-Limiting Membrane. Int. J. Dev. Neurosci. 1995, 13, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Yamasaki, R.; Takase, E.O.; Iida, K.; Watanabe, M.; Masaki, K.; Wijering, M.H.C.; Yamaguchi, H.; Kira, J.-I.; Isobe, N. Iguratimod Ameliorates the Severity of Secondary Progressive Multiple Sclerosis in Model Mice by Directly Inhibiting IL-6 Production and Th17 Cell Migration via Mitigation of Glial Inflammation. Biology 2023, 12, 1217. [Google Scholar] [CrossRef] [PubMed]
- Szpakowski, P.; Ksiazek-Winiarek, D.; Czpakowska, J.; Kaluza, M.; Milewska-Jedrzejczak, M.; Glabinski, A. Astrocyte-Derived Exosomes Differentially Shape T Cells’ Immune Response in MS Patients. Int. J. Mol. Sci. 2023, 24, 7470. [Google Scholar] [CrossRef] [PubMed]
- Corvace, F.; Faustmann, T.J.; Heckers, S.; Faustmann, P.M.; Ismail, F.S. Experimental Investigations of Monomethyl and Dimethyl Fumarate in an Astrocyte-Microglia Co-Culture Model of Inflammation. Pharmacology 2023, 108, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Quincozes-Santos, A.; Bobermin, L.D.; Souza, D.G.; Bellaver, B.; Gonçalves, C.-A.; Souza, D.O. Guanosine Protects C6 Astroglial Cells against Azide-Induced Oxidative Damage: A Putative Role of Heme Oxygenase 1. J. Neurochem. 2014, 130, 61–74. [Google Scholar] [CrossRef]
- Arús, B.A.; Souza, D.G.; Bellaver, B.; Souza, D.O.; Gonçalves, C.-A.; Quincozes-Santos, A.; Bobermin, L.D. Resveratrol Modulates GSH System in C6 Astroglial Cells through Heme Oxygenase 1 Pathway. Mol. Cell. Biochem. 2017, 428, 67–77. [Google Scholar] [CrossRef]
- Dall’Igna, O.P.; Bobermin, L.D.; Souza, D.O.; Quincozes-Santos, A. Riluzole Increases Glutamate Uptake by Cultured C6 Astroglial Cells. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2013, 31, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Badisa, R.B.; Wiley, C.; Randell, K.; Darling-Reed, S.F.; Latinwo, L.M.; Agharahimi, M.; Soliman, K.F.A.; Goodman, C.B. Identification of Cytotoxic Markers in Methamphetamine Treated Rat C6 Astroglia-like Cells. Sci. Rep. 2019, 9, 9412. [Google Scholar] [CrossRef] [PubMed]
- Mazzio, E.A.; Harris, N.; Soliman, K.F. Food Constituents Attenuate Monoamine Oxidase Activity and Peroxide Levels in C6 Astrocyte Cells. Planta Med. 1998, 64, 603–606. [Google Scholar] [CrossRef]
- Galland, F.; Seady, M.; Taday, J.; Smaili, S.S.; Gonçalves, C.A.; Leite, M.C. Astrocyte Culture Models: Molecular and Function Characterization of Primary Culture, Immortalized Astrocytes and C6 Glioma Cells. Neurochem. Int. 2019, 131, 104538. [Google Scholar] [CrossRef]
- Sibenaller, Z.A.; Etame, A.B.; Ali, M.M.; Barua, M.; Braun, T.A.; Casavant, T.L.; Ryken, T.C. Genetic Characterization of Commonly Used Glioma Cell Lines in the Rat Animal Model System. Neurosurg. Focus 2005, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vries, G.H.D.; Boullerne, A.I. Glial Cell Lines: An Overview. Neurochem. Res. 2010, 35, 1978–2000. [Google Scholar] [CrossRef]
- Urati, A.; Dey, M.; Gautam, A.S.; Singh, R.K. Iron-Induced Cellular in Vitro Neurotoxic Responses in Rat C6 Cell Line. Environ. Toxicol. 2022, 37, 1968–1978. [Google Scholar] [CrossRef]
- Major, E.O.; Miller, A.E.; Mourrain, P.; Traub, R.G.; Widt, E.D.; Sever, J. Establishment of a Line of Human Fetal Glial Cells That Supports JC Virus Multiplication. Proc. Natl. Acad. Sci. USA 1985, 82, 1257–1261. [Google Scholar] [CrossRef]
- Prowse, K.R.; Greider, C.W. Developmental and Tissue-Specific Regulation of Mouse Telomerase and Telomere Length. Proc. Natl. Acad. Sci. USA 1995, 92, 4818–4822. [Google Scholar] [CrossRef]
- Morikawa, M.; Asai, K.; Kokubo, M.; Fujita, K.; Yoneda, K.; Yamamoto, N.; Inoue, Y.; Iida, J.; Kishimoto, T.; Kato, T. Isolation and Characterization of a New Immortal Rat Astrocyte with a High Expression of NGF mRNA. Neurosci. Res. 2001, 39, 205–212. [Google Scholar] [CrossRef]
- Frisa, P.S.; Jacobberger, J.W. Cell Density Related Gene Expression: SV40 Large T Antigen Levels in Immortalized Astrocyte Lines. BMC Cell Biol. 2002, 3, 10. [Google Scholar] [CrossRef]
- Kamiichi, A.; Furihata, T.; Kishida, S.; Ohta, Y.; Saito, K.; Kawamatsu, S.; Chiba, K. Establishment of a New Conditionally Immortalized Cell Line from Human Brain Microvascular Endothelial Cells: A Promising Tool for Human Blood–Brain Barrier Studies. Brain Res. 2012, 1488, 113–122. [Google Scholar] [CrossRef]
- Whittemore, S.R.; Neary, J.T.; Kleitman, N.; Sanon, H.R.; Benigno, A.; Donahue, R.P.; Norenberg, M.D. Isolation and Characterization of Conditionally Immortalized Astrocyte Cell Lines Derived from Adult Human Spinal Cord. Glia 1994, 10, 211–226. [Google Scholar] [CrossRef]
- Price, T.N.C.; Burke, J.F.; Mayne, L.V. A Novel Human Astrocyte Cell Line (A735) with Astrocyte-Specific Neurotransmitter Function. Vitro Cell. Dev. Biol.-Anim. 1999, 35, 279–288. [Google Scholar] [CrossRef]
- Sonoda, Y.; Ozawa, T.; Hirose, Y.; Aldape, K.D.; McMahon, M.; Berger, M.S.; Pieper, R.O. Formation of Intracranial Tumors by Genetically Modified Human Astrocytes Defines Four Pathways Critical in the Development of Human Anaplastic Astrocytoma. Cancer Res. 2001, 61, 4956–4960. [Google Scholar]
- Sasai, K.; Akagi, T.; Aoyanagi, E.; Tabu, K.; Kaneko, S.; Tanaka, S. O6-Methylguanine-DNA Methyltransferase Is Downregulated in Transformed Astrocyte Cells: Implications for Anti-Glioma Therapies. Mol. Cancer 2007, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Alliot, F.; Pessac, B. Astrocytic Cell Clones Derived from Established Cultures of 8-Day Postnatal Mouse Cerebella. Brain Res. 1984, 306, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Booth, R.; Kim, H. Characterization of a Microfluidic in Vitro Model of the Blood-Brain Barrier (μBBB). Lab. Chip 2012, 12, 1784–1792. [Google Scholar] [CrossRef]
- Ito, R.; Umehara, K.; Suzuki, S.; Kitamura, K.; Nunoya, K.-I.; Yamaura, Y.; Imawaka, H.; Izumi, S.; Wakayama, N.; Komori, T.; et al. A Human Immortalized Cell-Based Blood-Brain Barrier Triculture Model: Development and Characterization as a Promising Tool for Drug-Brain Permeability Studies. Mol. Pharm. 2019, 16, 4461–4471. [Google Scholar] [CrossRef] [PubMed]
- Isogai, R.; Morio, H.; Okamoto, A.; Kitamura, K.; Furihata, T. Generation of a Human Conditionally Immortalized Cell-Based Multicellular Spheroidal Blood-Brain Barrier Model for Permeability Evaluation of Macromolecules. Bio-Protocol 2022, 12, e4465. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Patel, G.C.; Mao, W.; Clark, A.F. Establishment of a Conditionally Immortalized Mouse Optic Nerve Astrocyte Line. Exp. Eye Res. 2018, 176, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Jat, P.S.; Noble, M.D.; Ataliotis, P.; Tanaka, Y.; Yannoutsos, N.; Larsen, L.; Kioussis, D. Direct Derivation of Conditionally Immortal Cell Lines from an H-2Kb-tsA58 Transgenic Mouse. Proc. Natl. Acad. Sci. USA 1991, 88, 5096–5100. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Toida, A.; Horiuchi, Y.; Watanabe, S.; Sasahara, M.; Kawaguchi, K.; So, T.; Imanaka, T. Generation of an Immortalized Astrocytic Cell Line from Abcd1-Deficient H-2KbtsA58 Mice to Facilitate the Study of the Role of Astrocytes in X-Linked Adrenoleukodystrophy. Heliyon 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Krencik, R.; Weick, J.P.; Liu, Y.; Zhang, Z.-J.; Zhang, S.-C. Specification of Transplantable Astroglial Subtypes from Human Pluripotent Stem Cells. Nat. Biotechnol. 2011, 29, 528–534. [Google Scholar] [CrossRef]
- Caiazzo, M.; Giannelli, S.; Valente, P.; Lignani, G.; Carissimo, A.; Sessa, A.; Colasante, G.; Bartolomeo, R.; Massimino, L.; Ferroni, S.; et al. Direct Conversion of Fibroblasts into Functional Astrocytes by Defined Transcription Factors. Stem Cell Rep. 2015, 4, 25–36. [Google Scholar] [CrossRef]
- Haidet-Phillips, A.M.; Roybon, L.; Gross, S.K.; Tuteja, A.; Donnelly, C.J.; Richard, J.-P.; Ko, M.; Sherman, A.; Eggan, K.; Henderson, C.E.; et al. Gene Profiling of Human Induced Pluripotent Stem Cell-Derived Astrocyte Progenitors Following Spinal Cord Engraftment. Stem Cells Transl. Med. 2014, 3, 575–585. [Google Scholar] [CrossRef] [PubMed]
- McGivern, J.V.; Patitucci, T.N.; Nord, J.A.; Barabas, M.-E.A.; Stucky, C.L.; Ebert, A.D. Spinal Muscular Atrophy Astrocytes Exhibit Abnormal Calcium Regulation and Reduced Growth Factor Production. Glia 2013, 61, 1418–1428. [Google Scholar] [CrossRef] [PubMed]
- Serio, A.; Bilican, B.; Barmada, S.J.; Ando, D.M.; Zhao, C.; Siller, R.; Burr, K.; Haghi, G.; Story, D.; Nishimura, A.L.; et al. Astrocyte Pathology and the Absence of Non-Cell Autonomy in an Induced Pluripotent Stem Cell Model of TDP-43 Proteinopathy. Proc. Natl. Acad. Sci. USA 2013, 110, 4697–4702. [Google Scholar] [CrossRef] [PubMed]
- Shaltouki, A.; Peng, J.; Liu, Q.; Rao, M.S.; Zeng, X. Efficient Generation of Astrocytes from Human Pluripotent Stem Cells in Defined Conditions. Stem Cells 2013, 31, 941–952. [Google Scholar] [CrossRef]
- Chaboub, L.S.; Deneen, B. Astrocyte Form and Function in the Developing Central Nervous System. Semin. Pediatr. Neurol. 2013, 20, 230–235. [Google Scholar] [CrossRef]
- Yuan, S.H.; Martin, J.; Elia, J.; Flippin, J.; Paramban, R.I.; Hefferan, M.P.; Vidal, J.G.; Mu, Y.; Killian, R.L.; Israel, M.A.; et al. Cell-Surface Marker Signatures for the Isolation of Neural Stem Cells, Glia and Neurons Derived from Human Pluripotent Stem Cells. PLOS ONE 2011, 6, e17540. [Google Scholar] [CrossRef]
- Tcw, J.; Wang, M.; Pimenova, A.A.; Bowles, K.R.; Hartley, B.J.; Lacin, E.; Machlovi, S.I.; Abdelaal, R.; Karch, C.M.; Phatnani, H.; et al. An Efficient Platform for Astrocyte Differentiation from Human Induced Pluripotent Stem Cells. Stem Cell Rep. 2017, 9, 600–614. [Google Scholar] [CrossRef]
- Xu, X.; Lei, Y.; Luo, J.; Wang, J.; Zhang, S.; Yang, X.-J.; Sun, M.; Nuwaysir, E.; Fan, G.; Zhao, J.; et al. Prevention of β-Amyloid Induced Toxicity in Human iPS Cell-Derived Neurons by Inhibition of Cyclin-Dependent Kinases and Associated Cell Cycle Events. Stem Cell Res. 2013, 10, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Höing, S.; Rudhard, Y.; Reinhardt, P.; Glatza, M.; Stehling, M.; Wu, G.; Peiker, C.; Böcker, A.; Parga, J.A.; Bunk, E.; et al. Discovery of Inhibitors of Microglial Neurotoxicity Acting Through Multiple Mechanisms Using a Stem-Cell-Based Phenotypic Assay. Cell Stem Cell 2012, 11, 620–632. [Google Scholar] [CrossRef]
- Burkhardt, M.F.; Martinez, F.J.; Wright, S.; Ramos, C.; Volfson, D.; Mason, M.; Garnes, J.; Dang, V.; Lievers, J.; Shoukat-Mumtaz, U.; et al. A Cellular Model for Sporadic ALS Using Patient-Derived Induced Pluripotent Stem Cells. Mol. Cell. Neurosci. 2013, 56, 355–364. [Google Scholar] [CrossRef]
- Leventoux, N.; Morimoto, S.; Imaizumi, K.; Sato, Y.; Takahashi, S.; Mashima, K.; Ishikawa, M.; Sonn, I.; Kondo, T.; Watanabe, H.; et al. Human Astrocytes Model Derived from Induced Pluripotent Stem Cells. Cells 2020, 9, 2680. [Google Scholar] [CrossRef]
- Namihira, M.; Kohyama, J.; Abematsu, M.; Nakashima, K. Epigenetic Mechanisms Regulating Fate Specification of Neural Stem Cells. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2099–2109. [Google Scholar] [CrossRef]
- Yasui, T.; Uezono, N.; Nakashima, H.; Noguchi, H.; Matsuda, T.; Noda-Andoh, T.; Okano, H.; Nakashima, K. Hypoxia Epigenetically Confers Astrocytic Differentiation Potential on Human Pluripotent Cell-Derived Neural Precursor Cells. Stem Cell Rep. 2017, 8, 1743–1756. [Google Scholar] [CrossRef]
- Jiménez-Moreno, N.; Stathakos, P.; Caldwell, M.A.; Lane, J.D. Induced Pluripotent Stem Cell Neuronal Models for the Study of Autophagy Pathways in Human Neurodegenerative Disease. Cells 2017, 6, 24. [Google Scholar] [CrossRef]
- Moon, S.B.; Kim, D.Y.; Ko, J.-H.; Kim, Y.-S. Recent Advances in the CRISPR Genome Editing Tool Set. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Mei, A.; Marchetto, M.C. Generation of Inflammation-Responsive Astrocytes from Glial Progenitors Derived from Human Pluripotent Stem Cells. STAR Protoc. 2022, 3, 101261. [Google Scholar] [CrossRef] [PubMed]
- Kerkering, J.; Muinjonov, B.; Rosiewicz, K.S.; Diecke, S.; Biese, C.; Schiweck, J.; Chien, C.; Zocholl, D.; Conrad, T.; Paul, F.; et al. iPSC-Derived Reactive Astrocytes from Patients with Multiple Sclerosis Protect Cocultured Neurons in Inflammatory Conditions. J. Clin. Investig. 2023, 133, e164637. [Google Scholar] [CrossRef] [PubMed]
- Colombo, E.; Bassani, C.; De Angelis, A.; Ruffini, F.; Ottoboni, L.; Comi, G.; Martino, G.; Farina, C. Siponimod (BAF312) Activates Nrf2 While Hampering NFκB in Human Astrocytes, and Protects From Astrocyte-Induced Neurodegeneration. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Colombo, E.; Pascente, R.; Triolo, D.; Bassani, C.; De Angelis, A.; Ruffini, F.; Ottoboni, L.; Comi, G.; Martino, G.; Farina, C. Laquinimod Modulates Human Astrocyte Function and Dampens Astrocyte-Induced Neurotoxicity during Inflammation. Molecules 2020, 25, 5403. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Leng, K.; Park, J.; Sorets, A.G.; Kim, S.; Shostak, A.; Embalabala, R.J.; Mlouk, K.; Katdare, K.A.; Rose, I.V.L.; et al. Reactive Astrocytes Transduce Inflammation in a Blood-Brain Barrier Model through a TNF-STAT3 Signaling Axis and Secretion of Alpha 1-Antichymotrypsin. Nat. Commun. 2022, 13, 6581. [Google Scholar] [CrossRef] [PubMed]
- Maclean, F.L.; Williams, R.J.; Horne, M.K.; Nisbet, D.R. A Commentary on the Need for 3D-Biologically Relevant In Vitro Environments to Investigate Astrocytes and Their Role in Central Nervous System Inflammation. Neurochem. Res. 2016, 41, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.M.; Chen, C.S. Deconstructing the Third Dimension – How 3D Culture Microenvironments Alter Cellular Cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef]
- Xu, T.; Gregory, C.A.; Molnar, P.; Cui, X.; Jalota, S.; Bhaduri, S.B.; Boland, T. Viability and Electrophysiology of Neural Cell Structures Generated by the Inkjet Printing Method. Biomaterials 2006, 27, 3580–3588. [Google Scholar] [CrossRef]
- Lovett, M.L.; Nieland, T.J.F.; Dingle, Y.-T.L.; Kaplan, D.L. Innovations in 3D Tissue Models of Human Brain Physiology and Diseases. Adv. Funct. Mater. 2020, 30, 1909146. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, S.; Anand, S.; Shah, T.; Tasoglu, S. Bioprinting for Neural Tissue Engineering. Trends Neurosci. 2018, 41, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Czell, D. Genetics of Amyotrophic Lateral Sclerosis. Praxis 2019, 108, 37–44. [Google Scholar] [CrossRef]
- Warren, D.; Tomaskovic-Crook, E.; Wallace, G.G.; Crook, J.M. Engineering in Vitro Human Neural Tissue Analogs by 3D Bioprinting and Electrostimulation. APL Bioeng. 2021, 5, 020901. [Google Scholar] [CrossRef]
- Führmann, T.; Hillen, L.M.; Montzka, K.; Wöltje, M.; Brook, G.A. Cell-Cell Interactions of Human Neural Progenitor-Derived Astrocytes within a Microstructured 3D-Scaffold. Biomaterials 2010, 31, 7705–7715. [Google Scholar] [CrossRef]
- Fang, A.; Li, D.; Hao, Z.; Wang, L.; Pan, B.; Gao, L.; Qu, X.; He, J. Effects of Astrocyte on Neuronal Outgrowth in a Layered 3D Structure. Biomed. Eng. OnLine 2019, 18, 74. [Google Scholar] [CrossRef]
- Frampton, J.P.; Hynd, M.R.; Shuler, M.L.; Shain, W. Fabrication and Optimization of Alginate Hydrogel Constructs for Use in 3D Neural Cell Culture. Biomed. Mater. 2011, 6, 15002. [Google Scholar] [CrossRef]
- Matthiesen, I.; Jury, M.; Boroojeni, F.R.; Ludwig, S.L.; Holzreuter, M.; Buchmann, S.; Träger, A.Å.; Selegård, R.; Winkler, T.E.; Aili, D.; et al. Astrocyte 3D Culture and Bioprinting Using Peptide Functionalized Hyaluronan Hydrogels. Sci. Technol. Adv. Mater. 2023, 24, 2165871. [Google Scholar] [CrossRef]
- O’Donnell, J.C.; Katiyar, K.S.; Panzer, K.V.; Cullen, D.K. A Tissue-Engineered Rostral Migratory Stream for Directed Neuronal Replacement. Neural Regen. Res. 2018, 13, 1327–1331. [Google Scholar] [CrossRef]
- Seidlits, S.K.; Khaing, Z.Z.; Petersen, R.R.; Nickels, J.D.; Vanscoy, J.E.; Shear, J.B.; Schmidt, C.E. The Effects of Hyaluronic Acid Hydrogels with Tunable Mechanical Properties on Neural Progenitor Cell Differentiation. Biomaterials 2010, 31, 3930–3940. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Fitzgerald, W.; Liu, Q.-Y.; O’Shaughnessy, T.J.; Maric, D.; Lin, H.J.; Alkon, D.L.; Barker, J.L. CNS Stem and Progenitor Cell Differentiation into Functional Neuronal Circuits in Three-Dimensional Collagen Gels. Exp. Neurol. 2004, 190, 276–288. [Google Scholar] [CrossRef]
- Batenburg, K.L.; Sestito, C.; Cornelissen-Steijger, P.; Weering, J.R.T.V.; Price, L.S.; Heine, V.M.; Scheper, W. A 3D Human Co-Culture to Model Neuron-Astrocyte Interactions in Tauopathies. Biol. Proced. Online 2023, 25, 4. [Google Scholar] [CrossRef]
- Prasannan, P.; Siney, E.; Chatterjee, S.; Johnston, D.; Shah, M.; Mudher, A.; Willaime-Morawek, S. A 3D-Induced Pluripotent Stem Cell-Derived Human Neural Culture Model to Study Certain Molecular and Biochemical Aspects of Alzheimer’s Disease. Vitro Models 2022, 1, 447–462. [Google Scholar] [CrossRef]
- East, E.; Golding, J.P.; Phillips, J.B. A Versatile 3D Culture Model Facilitates Monitoring of Astrocytes Undergoing Reactive Gliosis. J. Tissue Eng. Regen. Med. 2009, 3, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.M.D.; Kavanagh, E.; Allenby, G.; Vassey, M. Bioengineered 3D Glial Cell Culture Systems and Applications for Neurodegeneration and Neuroinflammation. SLAS Discov. 2017, 22, 583–601. [Google Scholar] [CrossRef]
- de Melo, B.A.G.; Mundim, M.V.; Lemes, R.M.R.; Cruz, E.M.; Ribeiro, T.N.; Santiago, C.F.; da Fonsêca, J.H.L.; Benincasa, J.C.; Stilhano, R.S.; Mantovani, N.; et al. 3D Bioprinted Neural-Like Tissue as a Platform to Study Neurotropism of Mouse-Adapted SARS-CoV-2. Adv. Biol. 2022, 6, 2200002. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.; Long, X.; Xu, T. Three-Dimensional-Engineered Bioprinted in Vitro Human Neural Stem Cell Self-Assembling Culture Model Constructs of Alzheimer’s Disease. Bioact. Mater. 2022, 11, 192–205. [Google Scholar] [CrossRef]
- Brewer, G.J.; Torricelli, J.R. Isolation and Culture of Adult Neurons and Neurospheres. Nat. Protoc. 2007, 2, 1490–1498. [Google Scholar] [CrossRef]
- Gage, F.H.; Coates, P.W.; Palmer, T.D.; Kuhn, H.G.; Fisher, L.J.; Suhonen, J.O.; Peterson, D.A.; Suhr, S.T.; Ray, J. Survival and Differentiation of Adult Neuronal Progenitor Cells Transplanted to the Adult Brain. Proc. Natl. Acad. Sci. USA 1995, 92, 11879–11883. [Google Scholar] [CrossRef]
- Jensen, J.B.; Parmar, M. Strengths and Limitations of the Neurosphere Culture System. Mol. Neurobiol. 2006, 34, 153–161. [Google Scholar] [CrossRef]
- Shou, Y.; Liang, F.; Xu, S.; Li, X. The Application of Brain Organoids: From Neuronal Development to Neurological Diseases. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Sun, N.; Meng, X.; Liu, Y.; Song, D.; Jiang, C.; Cai, J. Applications of Brain Organoids in Neurodevelopment and Neurological Diseases. J. Biomed. Sci. 2021, 28, 30. [Google Scholar] [CrossRef]
- Mayhew, C.N.; Singhania, R. A Review of Protocols for Brain Organoids and Applications for Disease Modeling. STAR Protoc. 2023, 4, 101860. [Google Scholar] [CrossRef]
- Qian, X.; Nguyen, H.N.; Song, M.M.; Hadiono, C.; Ogden, S.C.; Hammack, C.; Yao, B.; Hamersky, G.R.; Jacob, F.; Zhong, C.; et al. Brain-Region-Specific Organoids Using Mini-Bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef]
- Nikolakopoulou, P.; Rauti, R.; Voulgaris, D.; Shlomy, I.; Maoz, B.M.; Herland, A. Recent Progress in Translational Engineered in Vitro Models of the Central Nervous System. Brain 2020, 143, 3181–3213. [Google Scholar] [CrossRef] [PubMed]
- Han, H.-W.; Hsu, S. Using 3D Bioprinting to Produce Mini-Brain. Neural Regen. Res. 2017, 12, 1595. [Google Scholar] [CrossRef] [PubMed]
- Cakir, B.; Xiang, Y.; Tanaka, Y.; Kural, M.H.; Parent, M.; Kang, Y.-J.; Chapeton, K.; Patterson, B.; Yuan, Y.; He, C.-S.; et al. Engineering of Human Brain Organoids with a Functional Vascular-like System. Nat. Methods 2019, 16, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.-N.; Jin, Y.; An, Y.; Kim, J.; Choi, Y.S.; Lee, J.S.; Kim, J.; Choi, W.-Y.; Koo, D.-J.; Yu, W.; et al. Microfluidic Device with Brain Extracellular Matrix Promotes Structural and Functional Maturation of Human Brain Organoids. Nat. Commun. 2021, 12, 4730. [Google Scholar] [CrossRef]
- Ormel, P.R.; Vieira de Sá, R.; van Bodegraven, E.J.; Karst, H.; Harschnitz, O.; Sneeboer, M.A.M.; Johansen, L.E.; van Dijk, R.E.; Scheefhals, N.; Berdenis van Berlekom, A.; et al. Microglia Innately Develop within Cerebral Organoids. Nat. Commun. 2018, 9, 4167. [Google Scholar] [CrossRef]
- Hong, Y.; Dong, X.; Chang, L.; Xie, C.; Chang, M.; Aguilar, J.S.; Lin, J.; Lin, J.; Li, Q.Q. Microglia-Containing Cerebral Organoids Derived from Induced Pluripotent Stem Cells for the Study of Neurological Diseases. iScience 2023, 26, 106267. [Google Scholar] [CrossRef]
- Sabate-Soler, S.; Nickels, S.L.; Saraiva, C.; Berger, E.; Dubonyte, U.; Barmpa, K.; Lan, Y.J.; Kouno, T.; Jarazo, J.; Robertson, G.; et al. Microglia Integration into Human Midbrain Organoids Leads to Increased Neuronal Maturation and Functionality. Glia 2022, 70, 1267–1288. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.; Tiwari, S.K.; Agrawal, K.; Hui, H.; Qin, Y.; Rana, T.M. Glial Cell Diversity and Methamphetamine-Induced Neuroinflammation in Human Cerebral Organoids. Mol. Psychiatry 2021, 26, 1194–1207. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Raju, T.R.; Bignami, A.; Dahl, D. Glial Fibrillary Acidic Protein in Monolayer Cultures of C-6 Glioma Cells: Effect of Aging and Dibutyryl Cyclic AMP. Brain Res. 1980, 200, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, K.; Moriya, Y.; Furuta, T.; Ohnishi, R.; Nishimoto, A. S-100 Protein in Human Glial Tumours. Acta Neurochir. 1982, 65, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Groves, A.K.; Entwistle, A.; Jat, P.S.; Noble, M. The Characterization of Astrocyte Cell Lines That Display Properties of Glial Scar Tissue. Dev. Biol. 1993, 159, 87–104. [Google Scholar] [CrossRef]
Figure 1.
Timeline of major developments in astrocytic cell culture. Major breakthroughs and milestones related to astrocyte cell culture are given in chronological order.
Figure 1.
Timeline of major developments in astrocytic cell culture. Major breakthroughs and milestones related to astrocyte cell culture are given in chronological order.
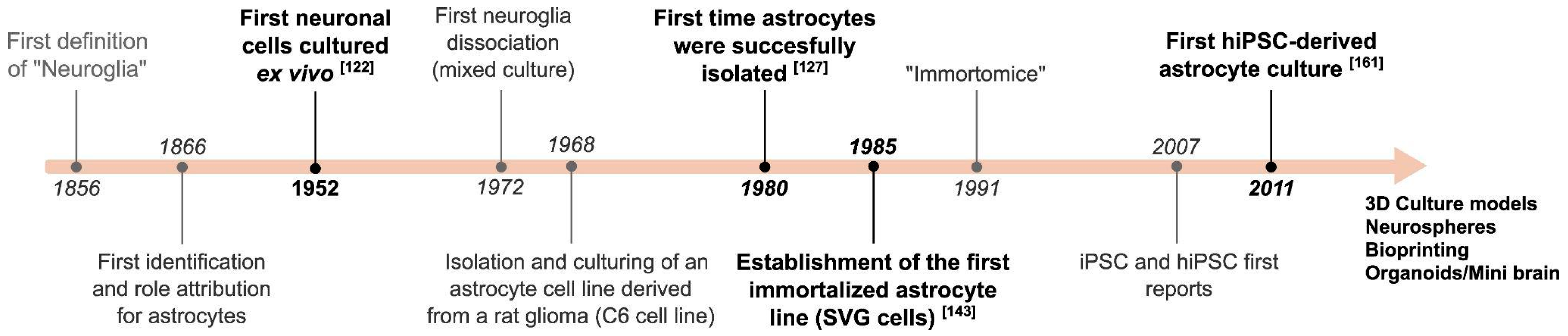
Figure 2.
Meaningful metabolic, gene expression and morphological changes in astrocytes. (A) Typical readouts related to astrocyte identification and those associated with reactive astrocytes. (B) Physiological astrocyte functions in barrier formation and tripartite synapse.
Figure 2.
Meaningful metabolic, gene expression and morphological changes in astrocytes. (A) Typical readouts related to astrocyte identification and those associated with reactive astrocytes. (B) Physiological astrocyte functions in barrier formation and tripartite synapse.
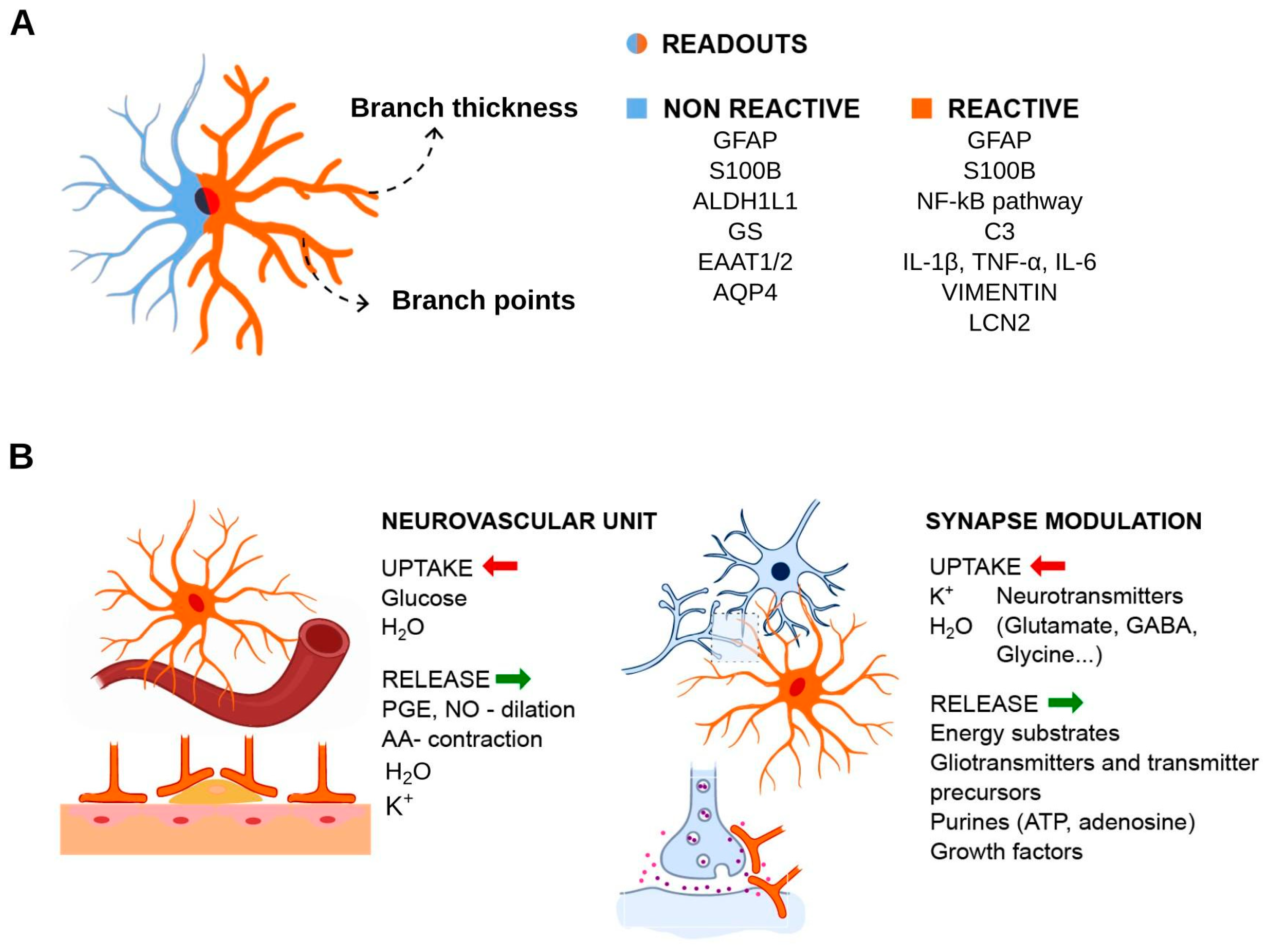

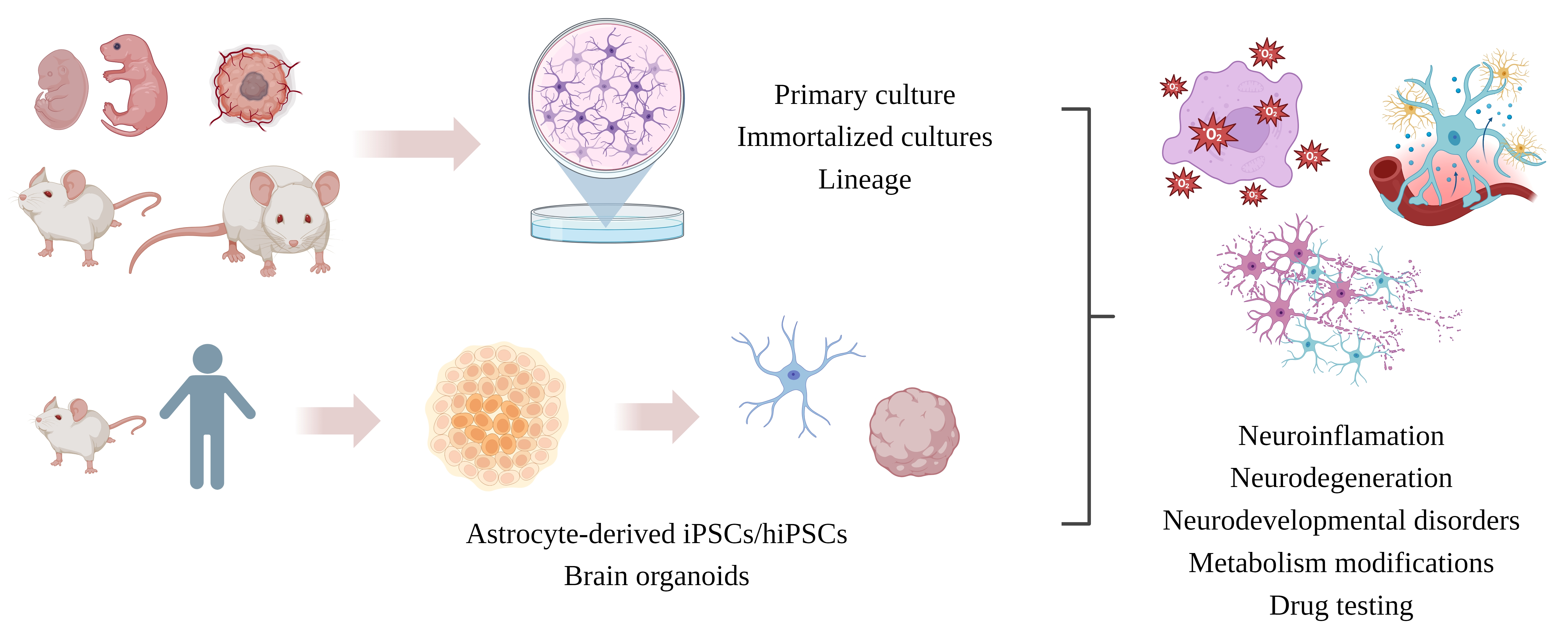{kind=link}
{kind=link}
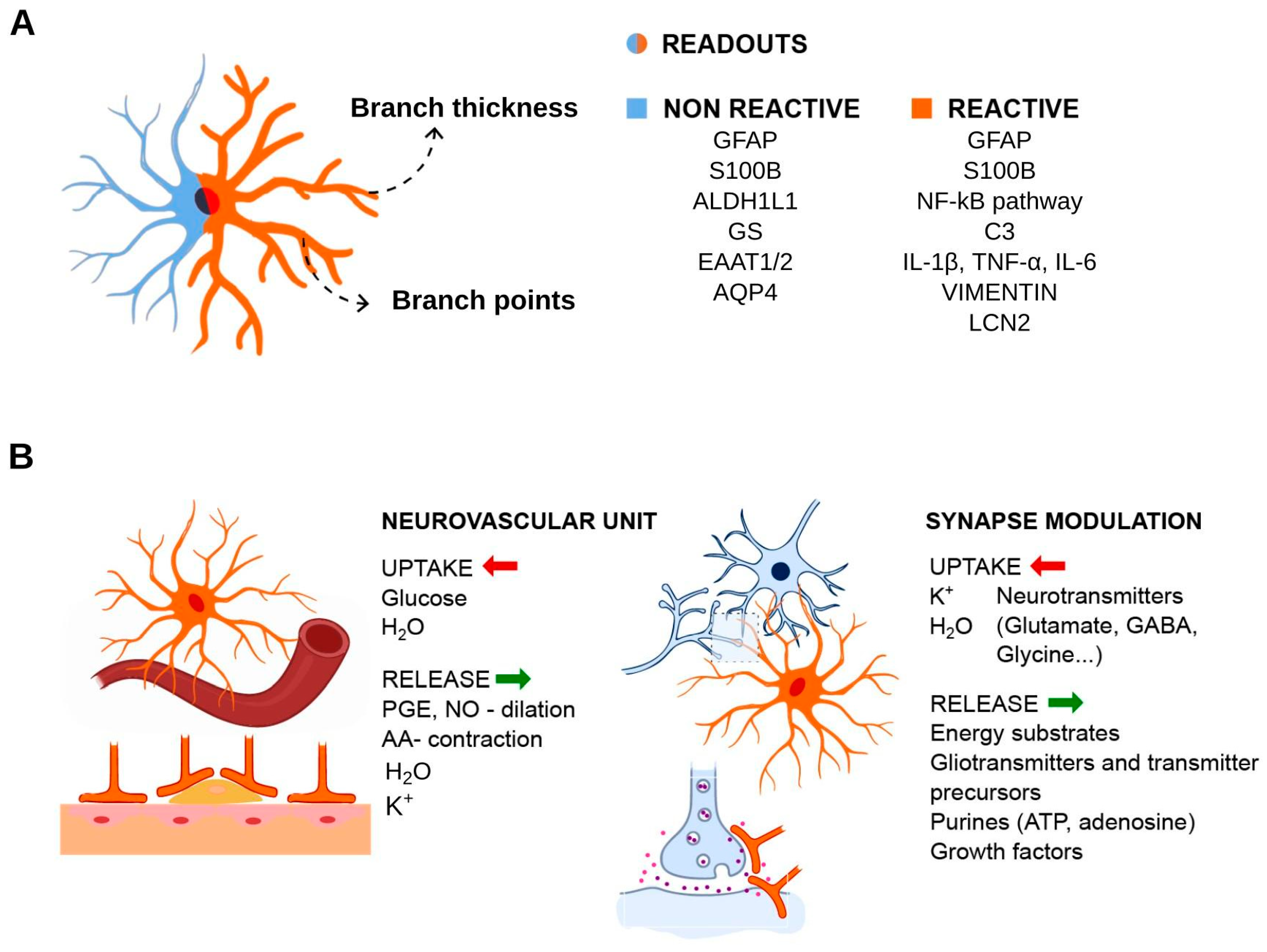{kind=link}
Table 1.
Comparison between the different cell culture techniques for astrocyte research.
| Culture Type Cell Line | Main Advantage | Limitations |
|---|---|---|
| Primary culture | Low cost and simple methodology. Disseminated methodology that is comparable to literature. | Usually employs serum (not obligatory) and is susceptible to induction of reactive phenotypes. High percentage of cellular senescence with passages; low proliferation; difficult to scale up; prone to contamination. |
| Cell lines | Easy to obtain (many are commercial); expandable; allow several passages; easy to manipulate; reduces animal usage | Cancer cell phenotype; expression profile differs from in vivo astrocytes. |
| Immortalized astrocytes | Intended to keep primary cell characteristics; less cell death and increased resilience to passages, freezing/thaw; easier to manipulate | Similar to primary cell culture; genomic integration that promotes cellular changes |
| iPSC-derived astrocyte | Relevant to disease studies (derived from patient); can be grafted; more similar to in vivo; reduces animal usage; good proliferation; allows differentiation into astrocyte types; serum-free | High cost; long-term culture; can present mutations |
| Bioprinting | Robotized; standardized dimensions; improves ECM | Expensive; delicate constructs; demands bioink characterization |
| 3D culture/organoids | Allows the analysis of complex interactions between different cell types; mimics in vivo conditions; cell heterogeneity | High cost; difficult to reproduce results, cell death in the organoid core; difficult nutrient diffusion |
Table 2.
Readouts in different astrocyte culture models (neuroinflammation studies).
| Astrocyte-Related Readouts | Primary Culture | Cell Lines | Immortalized Astrocytes | iPSC-Derived Astrocyte | Bioprinting | 3D Culture and Organoids |
|---|---|---|---|---|---|---|
| GFAP, S100B, Vimentin | [88,89,90,91,92,111,112] | [113,114] | [115,116,117] | [118,119,120] | [121,122,123] | [124] (GFAP only), [125] |
| C3, AQP4, ALDH1L1, CD44 | [92,111,112] | - | [115,117] | [118,119,120] | - | [126] |
| GS, GLT-1, GLAST | [111] | - | [115,117] | [118] | - | [126] |
| Metabolic assays (e.g., glutamate clearance/uptake, lactate and glucose uptake) | [111] | - | [115] | - | - | - |
| Pro-inflammatory (e.g., NFkB, TNF-α, IL-1 α, IL-1 β, IL-6) | [87,88,91,93] | [113,114,127] | [116,128] | [119] | [123] | [126] |
| Anti-inflammatory (e.g., IL-10) | - | - | - | - | - | [126] |
| Oxidative stress (e.g., ROS production, iNOS, DDIT3/CHOP *) | - | [113,114,127] | [116] | - | - | - |
| Intracellular calcium | - | [113,114] | - | - | - | - |
* DDIT3/CHOP: C/EBP homologous protein 10.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Preato, A.M.; Pinheiro, E.d.S.; Rosenstock, T.R.; Glezer, I. The Relevance of Astrocytic Cell Culture Models for Neuroinflammation in Neurodegeneration Research. Neuroglia 2024, 5, 27-49. https://doi.org/10.3390/neuroglia5010003
AMA Style
Preato AM, Pinheiro EdS, Rosenstock TR, Glezer I. The Relevance of Astrocytic Cell Culture Models for Neuroinflammation in Neurodegeneration Research. Neuroglia. 2024; 5(1):27-49. https://doi.org/10.3390/neuroglia5010003
Chicago/Turabian StylePreato, André Maciel, Ester da Silva Pinheiro, Tatiana Rosado Rosenstock, and Isaias Glezer. 2024. "The Relevance of Astrocytic Cell Culture Models for Neuroinflammation in Neurodegeneration Research" Neuroglia 5, no. 1: 27-49. https://doi.org/10.3390/neuroglia5010003