Safety Bubbles: A Review of the Proposed Functions of Froth Nesting among Anuran Amphibians


School of Environmental and Life Sciences, University of Newcastle, Callaghan, NSW 2308, Australia
Ecologies 2021, 2(1), 112-137; https://doi.org/10.3390/ecologies2010006
Submission received: 21 December 2020
/
Revised: 12 January 2021
/
Accepted: 1 February 2021
/
Published: 4 February 2021
(This article belongs to the Special Issue Feature Papers of Ecologies 2021)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The adults of several anuran amphibian species deposit their eggs externally in mucus secretions that are purposely aerated to produce a froth nest. This type of clutch structure has evolved independently several times in this group and has been proposed to serve a variety and often simultaneous adaptive functions associated with protecting offspring from sub-optimal conditions during embryogenesis and later stages after hatching has occurred. These functions range from buffering offspring from sub-optimal temperatures and desiccation, to defending against predation and improving oxygenation. This versatility has likely helped facilitate the reduced reliance of egg development on water and thus the penetration of anurans into environments where permanent aquatic systems are not always available. In this paper, I review the hypothesised functions of the anuran froth nest as a mucus-based solution to the environmental challenges offspring face during development, with consideration of the functions of froth nest breakdown and communal froth nesting, as well.
1. Introduction
An overwhelming majority of anuran amphibians are oviparous, laying their eggs externally and often providing little or no parental care thereafter [1]. Upon oviposition, these eggs are thus exposed to a variety of risk factors, the interaction of which has driven the evolution of a diversity of clutch structures to improve the odds of survival during embryogenesis [2]. One of these includes the purposeful incorporation of air bubbles into mucus to produce a frothy spawn body into which the eggs are deposited and complete development. These frothy masses, commonly referred to as foam nests, have been recorded in species from at least 19 genera across seven families, including hylidae, hyperoliidae, leiuperidae, leptodactylidae, microhylidae, myobatrachidae, and rhacophoridae [3,4,5,6,7,8]. The procedure of aeration and final form of the nest differs widely among families, suggesting that the behaviour has evolved independently several times among the anuran amphibians as a mucus-based solution to the environmental challenges that come with an oviparous life history [9,10,11].
As with other animal groups, mucus secretions produced by anuran amphibians serve a range of functions at various life history stages, including capturing prey [12], defence against pathogens [13], protecting the digestive system [14], lubrication [15], and reproduction [2]. Encapsulating eggs in frothed mucus has been proposed to serve a variety adaptive functions that improve the speed and/or probability of successful embryogenesis and hatchling survival [16], by altering the conditions offspring are exposed to during incubation. As more natural history data are obtained for froth nesting species, there is an increasing need to consolidate this information to provide a foundation for future research in this field. In this study, I explore froth nesting as a reproductive innovation among the anurans. I review evidence gathered for each hypothesised function of the anuran froth nest, along with the function of froth nest breakdown and communal froth nesting. Finally, I present experimental protocols that should be followed in order to streamline future studies in this field.
2. Froth Nest Construction
There is a great diversity of environmental settings in which froth nests are built, including aquatically on water surfaces or margins [5,9], terrestrially on moist forest floors or in burrows [17,18], and arboreally attached to plant structures such as branches and leaves [7,19]. The precursor fluid that constitutes the nest is mucus excreted from glands located on the skin of amplectant adult pairs [20], or as cloacal fluid from the female [21] or male [7], which is released prior to egg extrusions and then mixed with water obtained from the surrounding environment in aquatic species [22], or expelled from the bladder in terrestrial/arboreal species [23]. It has been suggested for several species that the froth is actually a combination of both mucus secretions and the outer layer(s) of jelly that surround each egg [24,25], and thus the eggs are released simultaneously with the mucus. This mixture of mucus, water and possibly jelly is subsequently aerated, which involves highly synchronised movements by the adult pairs while in inguinal or axillary amplexus, though may include assistance from additional non-amplectant males in some species [26]. This results in the production of nests composed of either foam or bubbles (Figure 1) [1,2,8]. Throughout the current study, nest type has been classified as either foam or bubble based for each species. However, both foam and bubble-based nest types have been collectively defined as ‘froth nests’ as per Altig and McDiarmid [2].
Foam-based nests are typically composed of small air bubbles, which are produced primarily through the beating of the adult’s legs. In rhacophorids and leptodactylids it is generally the male that uses his hind legs to physically whip the mucus into foam [6,7,24], while in at least one species of hylid, foam is produced by the female jumping onto the mucus after it has been excreted [9]. In contrast, bubble-based nests typically contain large air bubbles that are transported into the mucus without whipping. In the myobactrachids, females use specialised flanged fingers to scoop air bubbles into the nest as mucus is released from the oviduct [5,27], while adult pairs in at least one microhylid dive below the water to release air bubbles through their nostrils into the mucus above to form the bubble nest [28]. The inclusion of air causes the froth nest of foam and bubble nesting species to become inflated to a much greater size than what would be possible without aeration. This results in the formation of a structured, spherical body, though the nests of some species remain relatively flat due to the lack of incorporated air [28].
Anuran froth nests must be environmentally resilient, often for extended periods even after embryos have hatched, as well as biologically compatible with the survival of the residing offspring that are in close physical contact with the frothed material. This is a biological challenge as detergent-like molecules with surface activity are generally required to overcome the high-surface tension of water to produce stable foams and bubbles, yet these compounds would be damaging to the unprotected cell membranes of sperm and eggs and subsequently embryos in external fertilisers such as frogs [22]. In the absence of these molecules, viscous materials can be whipped by vigorous mechanical agitation (e.g., egg whites), but such a strategy for froth nest construction would come at the cost of high energy expenditure, particularly as foam fluid is not as vicious to allow for effective entrapment of air [29], and with tendency for collapse without additional stabilising processes [30]. Instead, Flemming et al. [22] have reported on a set of proteins (referred to as ranaspumins) present in the froth nest of the South American species Engystomops pustulosus (originally Physalaemus pustulosus), one of which exhibits strong surface activity without causing cell damage associated with conventional small molecule surfactants that are able to penetrate cell membranes, which suggests a remarkable adaptation for froth production among the anurans. Froth nests produced by this species, and perhaps others, have been suggested to be initially formed through the presence of this protein at the air–water interface near the surface of the nest and then further stabilised through the action of other protein components present within the mucus, thereby ensuring the nest persists for sufficient time for embryos to complete development [29].
3. The Evolution of Froth Nesting
It is generally considered that the evolution of froth nesting has helped facilitate the reduced dependence of offspring of some anurans on freestanding water by assisting in the transition of breeding to temporary aquatic systems, as well as terrestrial and arboreal settings [31]. This has thus allowed the penetration of anurans into open formation habitats, as well as arid and seasonal environments, where freestanding water is less available [10,11,16,32]. Evidence for this is based on the protection that froth production has been suggested to offer against several threats encountered when egg deposition occurs away from permanent water, including an increased risk of mortality via anoxia, desiccation or thermal damage as a result of exposure to sub-optimal oxygen, moisture and temperature levels [33,34]. It is thus a component of life history, along with other traits such as site selection, parental care and both egg and clutch size, that has played a role in the colonisation of challenging niches in this group [31], and one of several reproductive modes that has allowed egg deposition to occur outside of water [35].
The various reproductive modes found among froth nesting species today show the potential intermediary steps towards terrestrial egg deposition and this concomitant reduction in the reliance on freestanding water [10,11]: (1) Eggs are laid aquatically but have the capacity to remain viable in the absence of water for short periods of time until subsequent rainfall replenishes water levels [16,36]. (2) Eggs are purposely laid out of water, with hatchlings flushed into waterbodies during rainfall from terrestrial nests [10,17] or drop from arboreal nests to complete development [7,19]. (3) Eggs laid out of water can continue development within the froth nest for an extended period after hatching has occurred [26,37]. (4) Eggs are deposited terrestrially, often in burrows, where they complete development completely independent of water [18,38,39]. While it is apparent that most froth nesting species are still reliant on freestanding water for some or all stages of offspring development, for those species that have made the complete transition to terrestrial development, the froth nest has effectively replaced the need for water by providing all necessary resources (e.g., moisture, oxygen, nutrients) for survival and development.
Two key traits necessary for froth nest construction are common among anurans, including the excretion of mucus alongside eggs and stabilising movements performed during amplexus, the latter of which subsequently evolved into precise hand or legs movements required for incorporating air bubbles into the egg mass [2]. Assuming that most anurans secrete reproductive mucus, this suggests that few changes, mostly behavioural, are required for the shift to this reproductive strategy [10], as evidenced by the presence of both froth and non-froth nesting populations of the Australian bubble-nester, Limnodynastes tasmaniensis [40]. Even so, relatively few species have come to produce froth nests [10], despite most possessing these pre-adaptations needed for their construction, suggesting that this evolutionary change has occurred in response to certain selective pressures that are widespread across families but not across all species.
4. Benefits of Froth Nesting
Currently, five main adaptive functions of froth nesting have been proposed, including increasing egg temperature, buffering eggs against sub-optimal temperatures, protection against desiccation and predation, and improved oxygenation. These are addressed in detail below, along with several other proposed functions that may be supplementary.
4.1. Increasing Egg Temperature
Environmental temperature influences a variety of processes in anuran offspring, which has subsequently driven both behavioural and physiological adaptations key for reproduction in thermally stressful environments [41]. Laying eggs in froth may alter the thermal conditions they are exposed to prior to hatching (and following hatching if they remain within the nest), creating an internal environment that accelerates growth.
Empirical evidence for the heating effect of froth nesting has been shown to be contradictory among studies, even for those on the same species. For example, work conducted by Dobkin and Gettinger [42] showed that the internal temperature of E. pustulosus froth nests warmed more rapidly via solar radiation than the surrounding air, exceeding ambient temperatures by as much as 4 °C during the day. However, Downie [16] found no thermal effect of froth nesting in this species across all but later stages of development, alluding to a possible flaw in Dobkin and Gettinger’s [42] study as they only measured temperatures on late stage nests that had already begun to break down and when tadpoles had already started to hatch. For eggs laid aquatically, the heating benefits of frothing are obtained as the nest floats on the surface of the water where eggs can be exposed to direct solar radiation, which likely increases internal temperatures more rapidly compared to eggs laid below the surface [43]. Here, the low albedo of anuran eggs, which are commonly black, may play a critical role in this process of heat absorption [44]. While the froth nest has been suggested to provide a thermally advantageous environment that allows for rapid development [42], experimental evidence of increased developmental rate as a result of the presence of froth is currently lacking.
Countering this proposed heating benefit of froth nesting, it has also been suggested that the high albedo of aerated secretions, which turn white as a result of the incorporation of air bubbles and become reflective as their surfaces dry (Figure 2), protect eggs from insolation, thereby keeping temperatures down [45,46]. Yet, Dobkin and Gettinger [42] showed that the frothed nests of E. pustulosus did not reflect significant amounts of solar radiation, suggesting that the high albedo of the nest does not serve to protect against overheating. Instead, the process of drying has been suggested to produce a partially water-proof barrier that may remain flexible and resistant to cracking [23], thereby preventing evaporative cooling [42,47].
If the froth nest is able to increase egg temperatures, this may be adaptive by facilitating rapid offspring development [48,49]. For species that exploit temporary aquatic systems, this potential benefit of froth nesting is particularly advantageous, given the time constraints on metamorphosis caused by the periodic drying of these systems. Relatively smaller temperature increases have been reported in species that deposit their eggs in un-frothed spawn (e.g., Rana sylvatica; [43]), while the eggs of temperate amphibians are known to maintain higher than ambient temperatures irrespective of whether or not they are encapsulated in froth [44]. Likewise, species may actively select oviposition sites that are simply warmer [49], highlighting the need for further research to determine the extent to which frothing provides additional heating benefits.
4.2. Insulation against Sub-Optimal Temperatures
Evidence has also been obtained to suggest that the froth nest acts to insulate eggs from sub-optimal or variable environmental temperatures. For example, internal froth nest temperatures have been shown to be lower than that of the nest surface and surrounding air in several species [46,47,50]. Likewise, Méndez-Narváez et al. [33] found internal temperatures of both aquatic and terrestrial froth nests of Leptodactylus species were lower during hotter times of the day and higher at night when temperatures decreased, thereby stabilising internal temperatures in thermally fluctuating environments. As a buffer to the external environment, the froth nests may thus increase the chances of offspring survival by either protecting them from thermal extremes that would cause mortality or damage, or by allowing for faster developmental rates.
This property of the froth nest has been attributed to the presence of trapped air inside the bubbles of the nest that decreases the nest’s overall thermal conductivity, insulating the eggs in a manner similar to air trapped between feathers or fur [51,52]. While the air inside the froth nest is likely to be moist, moist air is not a poorer insulator compared to dry air at biologically relevant temperatures (below 40 °C) [53]. The increase in nest size as a result of the incorporation of air also causes many of the eggs to be located further away from the surface, which likely buffers them more effectively from external temperature fluctuations as heat changes take longer to be transferred to the nest’s core, similar to the insulating effect of placing egg clutches in communal masses [49].
To date, there has been no experimental manipulation of froth nests to examine the extent to which trapped air bubbles provides greater insulation than un-frothed mucus or egg capsules, the latter of which has been shown to act as an insulator in egg masses of the non-frothing species Rana temporaria temporaria [54]. There is currently also no experimental evidence to suggest that egg survival rate is higher in frothed nests compared to un-frothed nests. The role of froth production in protecting eggs from sub-optimal temperatures is also dependent on their innate ability to thermally adjust and/or tolerate short term exposure to such temperatures [55]. This is apparent in E. pustulosus, whose eggs have a wide thermal tolerance that is likely correlated with the wide microhabitat temperatures they have evolved under [56]. Even so, the froth nest may still be of critical value in species that occupy environments with strong fluctuations in temperature, protecting eggs from sub-optimal temperatures both within and beyond thermal tolerances.
4.3. Environmental Control of Froth Nest Temperature
Several of these proposed thermal benefits of froth nesting have opposing effects, suggesting that they are dependent on the conditions of the oviposition site. Indeed, froth nest temperatures are likely to be influenced by their placement in the environment, as this determines the extent of their exposure to external factors such as air and substrate (water or ground) temperatures and solar radiation.
The heating benefits of froth nesting in particular is likely restricted to egg laying environments above ground and in open habitats where direct exposure to solar radiation is possible. For example, increased internal temperatures of E. pustulosus nests were the result of exposure to solar radiation during the day [42], which is in contrast to Lechriodus fletcheri nests which did not show increased daytime temperatures, possibly due to dense canopy cover overhead minimising sunlight exposure (J. Gould, unpubl. data). Nests that are deposited above ground are also more likely to be exposed to greater fluctuations in temperature that may negatively affect embryo development [47,57], particularly in temperate regions where temperatures can decline severely overnight and in temporary aquatic systems which are often poor thermal masses given their smaller size and shallowness [42]. Indeed, the eggs of some Leptodactylus species appear to be more thermally insulated in terrestrially compared to aquatically built nests, which may occur as microhabitat conditions in ground depressions or burrows provide partial or full protection from air temperature changes [33,57]. Finally, the extent of temperature variations may also be attributed to the extent of sunlight exposure. This was demonstrated in nests created by Leptodactylus knudseni and Leptodactylus labyrinthicus, which although of comparative size and placement within the environment, exhibited different temperature fluctuations as a result of differences in oviposition habitat openness [47].
The positioning of eggs within the nest also determines the extent to which each is protected from their thermal environment, with those on the periphery more likely to be exposed to sub-optimal and/or fluctuating temperatures compared to those in the centre [47]. If so, then aspects of the frothed nest, including the amount of mucus that is secreted around the eggs, the amount of air incorporated and the nest’s final viscosity, are critical in determining the extent of exposure of each egg. Nests composed of more mucus and inflated with a large number of air bubbles form larger, more substantive dome-like structures that should provide for better insulation (Figure 2). Indeed, Shepard & Caldwell [47] suggest that the lack of temperature differences seen in E. pustulosus froth nests by Downie [16] is partly due to their small size. Nests composed of less mucus or fewer bubbles form flatter, almost film-like structures where a majority of eggs remain close to the surface (e.g., [28]). While this makes them more vulnerable to external temperature fluctuations as they remain exposed, it could still be advantageous by allowing for more rapid temperature increases via increased sunlight exposure. Most of these aspects of anuran froth nests remain to be fully explored. If the apparent differences in froth nests between species is dependent on the thermal requirements of the eggs under particular environmental conditions, it is imperative for future studies to analyse the correlation between spawn structure and size with factors such as ambient temperature and exposure to solar radiation.
4.4. Anti-Desiccation
Amphibian eggs are highly vulnerable to desiccation if moisture levels begin to drop given that they are surrounded by a highly permeable membrane and lack a protective, non-permeable shell [58]. This is a physiological challenge for species that exploit arid or seasonal environments, as well as temporary aquatic systems if rainfall is intermittent and hydroperiods are frequently below the minimum threshold time required for offspring to complete development [1,16,59]. Laying eggs within a froth nest has been suggested by several authors to offer them protection from the immediate risk of desiccating in the absence of moisture [4,10,42,60]. This may occur in three ways: (1) Drying of the spawn surface via evaporative water loss results in the formation of a reflective and/or non-permeable outer layer that reduces the rate of water loss from inner regions of the nest by deflecting solar radiation and locking in moisture [1,26,42,47,61]. (2) The incorporation of air bubbles results in a relatively larger and structured spawn body that has an insulating effect, reducing the nest’s surface area relative to its volume and placing distance between the residing eggs and external environment [25,51,52,62]. Adults of some species actively insert eggs into the interior of the nest such that few are left near to the surface, adding to this effect [63]. (3) The process of drying may also preserve this three-dimensional structure of the nest, preventing it from expanding and thinning out, which would expose embryos to the environment (Figure 3).
These potential benefits of producing froth maintain moisture around the eggs for a longer period of time than what would be possible if eggs were laid solitarily [16], thereby improving their probability of survival in environments of sub-optimal moisture. However, it remains to be determined the extent to which frothing provides additional benefits to eggs laid in un-frothed mucus. For example, the non-water mass of foam material created by E. pustulosus is composed of a high proportion of carbohydrates [29], which Flemming et al. [22] suggest would be effective in preventing dehydration, suggesting that non-frothed nests would also possess at least this anti-desiccation property.
The importance of froth production in protecting eggs from desiccation should be dependent on the environment in which oviposition occurs, including the degree of exposure to direct solar radiation, humidity and wind. For species that oviposit in temporary waterbodies, frothed mucus creates a protective cocoon that preserves water around the eggs even if standing water has dried up. This sustains the viability of the eggs until subsequent rainfall recharges the system, often breaking apart the nest at the same time to allow for the hatching of tadpoles into water [10]. This has been recorded in L. fletcheri (J. Gould, unpubl. data) and E. pustulosus [16,36], where the eggs of both species can survive undamaged for several days after initially becoming stranded out of water after oviposition, increasing their chances of being ‘rescued’ by follow up rainfall. Méndez-Narváez et al. [33] also showed internal embryos of froth nests created by another aquatic depositor, Physalaemus fischeri, also survived when translocated onto land. Similar benefits have also been recorded in terrestrial nesting species. For example, the formation of a dried outer crust in terrestrial nests produced by L. labyrinthicus has the capacity to protect offspring within a moist environment for days after hatching has occurred, which is evident by the fact that disruption of the crust causes rapid desiccation [47]. This protection is, however, not indefinite, and total egg clutch mortality will occur at some point once the nest desiccates entirely.
For species that oviposit aquatically, the incorporation of air bubbles into the nest causes it to float at the surface of the water where eggs are exposed to the air and seemingly at greater risk of desiccating. While this seems like an evolutionary paradox, these eggs are still less likely to desiccate than those laid in un-frothed spawn or solitarily once the waterbody dries up, given that the froth nest can substitute for the absence of moisture and insulate eggs for extended periods of time [16]. Further, the surfaces of floating froth nests may not begin to dry until there is a lack of freestanding water beneath, which suggests that there is little increased desiccation risk associated with the transition of development to the water’s surface. This has been recorded for L. fletcheri nests in the field (J. Gould, unpubl. data) which do not begin to dry until they become stranded out of water, which may occur as water lost at the surface through evaporation is replenished by water making contact with sections of nest below. While this hypothesis remains to be tested, dried froth nests have been shown to rehydrate and be renewed when exposed to light rain [17].
Similar benefits are provided to species that lay terrestrially and arboreally as eggs are vulnerable to evaporative water loss through wind exposure, though contact with wet soil and/or deposition in excavated basins that are capped may partially reduce contact with the air and protect the eggs from this threat [17,57]. It must be noted that terrestrial oviposition presumably only evolves in regions where desiccation risk is not a limiting factor for offspring survival, namely the tropics [64] but also in dry environments so long as moisture can be maintained around the eggs during incubation [65]. Indeed, the adults of some terrestrial nesters, such as Rhacophorus malabaricus, cover their froth nests in leaf litter, which has been shown to be an anti-desiccation behaviour that further conserves moisture around the developing embryos by reducing their exposed surface area [7]. While likely more advantageous in temporary environments with variable climatic conditions, egg survival may also be improved when placed in froth in other situations where desiccation is not generally considered a primary risk factor. In particular, nests laid on the periphery of permanent waterbodies can still become stranded on dry land if water levels recede [16], exposing them to sub-optimal moisture levels despite their proximity to freestanding water. For species that lay in subterranean burrows, however, there may be little risk of desiccation and, as such, little need for froth production to evolve specifically in response to this threat [10,66].
Another factor that likely influences the selection for froth production for its anti-desiccation properties is the length of time offspring remain within the nest. Silva et al. [17] suggests that this may be of little importance for Physalaemus species whose offspring hatch immediately from froth nests into freestanding water after only a few days [67,68], compared to other froth nesting species (e.g., L. labyrinthicus) whose embryos hatch in the nest and remain there as tadpoles for several weeks [17]. However, this is likely to be context dependent, particularly being influenced by the speed and extent to which offspring are exposed to desiccating conditions upon oviposition. This is clearly demonstrated in species that exploit temporary aquatic systems, who have evolved rapid developmental rates in response to short hydroperiods [44], but may still require the protective properties of froth over the course of this period if freestanding water commonly dries up before egg development is complete.
The capacity of froth to protect eggs from desiccation is also dependent on the location of eggs within the nest itself. Initially, eggs on the periphery of the nest are the first to desiccate given their proximity to the external environment, which has been recorded in both Limnodynastes peronii [69] and L. fletcheri (J. Gould, unpubl. data, Figure 3). Gradually, the desiccation ‘front’ moves inwards to deeper sections of the nest, killing off more eggs until it reaches the centre. Given this disparity in desiccation risk, it would be highly advantageous for adults to purposely bury eggs deeper within the nest [16]. Indeed, some species (e.g., E. pustulosus and Chiromantis xerampelina) have evolved complex nesting sequences whereby eggs are exclusively positioned in the centre where the risk of desiccation is much smaller [23,63,70]. However, many froth producing species scatter eggs throughout the nest, causing eggs to be distributed randomly, including near the surface where the risk of desiccation is much greater [4,71]. It is possible that the benefits of producing complex nest structures that move eggs away from the surface may not outweigh the associated costs on the parents in all species, given that amplexus and nest production are already energetically expensive activities and may increase the risk of predation [72], and especially considering that a large proportion of eggs are still positioned away from the surface even in less organised nest structures.
If egg location within the nest does have an effect on desiccation risk, there should be strong selection for larger spawn structures, either through the production of greater quantities of mucus by females or increased aeration activity by amplecting pairs during nest construction. Indeed, larger L. fletcheri froth nests were found to have a slower rate of evaporative water loss than smaller nests, with more eggs located away from the surface and buffered by a thicker section of mucus (J. Gould, unpubl. data). However, constraints on amplecting adults, such as the size of the female, restrict the size of the nest that can be produced [23]. As such, smaller nests produced by some species, such as Physalaemus riograndensis whose nests are between 3 to 4 cm in diameter [73], are likely to be less effective at protecting against desiccation than those that produce larger nests, such as those of C. xerampelina which may reach 20 cm in diameter [74].
4.5. Predator Defence
Anuran eggs are vulnerable to predation as they are laid externally, immobile and often are not provided parental care [75]. Covering eggs in froth may act as a ‘fixed predator avoidance’ trait [64] that reduces predation risk in a variety of ways: (1) The surface of the froth nest may dry [23,26], creating a hard, shell-like protective barrier that is difficult to penetrate for predators to gain access to the eggs within. (2) If the outer layer of froth has not dried, it may remain highly adhesive (Figure 4), causing predators that make contact with it to become stuck and, therefore, incapacitated before they can reach the eggs [76]. (3) Aeration of the nest causes the eggs to become buried within a comparatively larger and more structured body [63], reducing the number of eggs near the surface. (4) For species that lay aquatically, the ability of froth nests to float causes a large portion of eggs to sit out of the water column, isolating them from aquatic predators [10,16,25]. If the spawn are free-floating and not attached to the edges of the waterbody, they may also form ‘island’ refuges that are difficult for terrestrial predators to reach. Of course, the probability of successful hatching occurring from froth nests is still highly dependent on predation pressure, which is influenced by the density of predators near the nest and the presence of other nests nearby that may relieve this pressure.
Currently, the greatest evidence for the predator defence effect of froth nesting is related to the removal of eggs from the water column. Aquatically deposited anuran eggs are known to be eaten by a variety of aquatic predators, including hetero- and con-specific tadpoles, fish and macro-invertebrates [64,77,78]. As the froth nest floats at the surface of the water, aquatic predators are restricted to consuming eggs at the base and around the perimeter of the structure (Figure 5), which has been observed in L. fletcheri [79] and E. pustulosus [25]. This reduces the total number of eggs at any given time that can be consumed, thereby increasing the likelihood that at least some can complete development. Several species of froth nesters are known to have oophagous tadpoles that are cannibalistic, which may be a strong selecting pressure for the evolution of froth nesting [18,64,79,80]. In contrast, hetero-specific predation is not likely to be a primary threat for those found in waterbodies exploited by few other species, which appears to be a hallmark of several froth nesters that exploit highly temporary waterbodies (e.g., P. diplolistris; [80]).
It is important to realise that the froth nest does not provide absolute protection against any predator, a evidenced by observations of cannibalism in L. fletcheri where more eggs gradually become exposed to the threat of being eaten as peripheral mucus material and eggs are consumed [79]. Froth nesting may also be of little benefit for species such as L. tasmaniensis that deposit their eggs below the froth and remain mostly exposed to the water column throughout development [5]. For all other species, however, having eggs buried within a thick layer of froth slows the rate at which egg consumption occurs, suggesting that the combined effect of being buried within froth that is also free-floating is an effective defensive strategy.
There is currently scarce evidence to suggest that drying of the froth nest surface has a defence benefit. However, the outer layers of mucus produced by some terrestrial froth nesters are known to dry around a moist, inner region where the eggs reside [23,26]. While this may primarily serve to protect the eggs from desiccation, it may have supplementary benefits that include protection against would-be predators, although this requires further investigation. Lingnau and Di-Bernardo [76] suggests that the adhesive quality of froth prior to drying presents an obstacle for some terrestrial predators, restricting them to eggs on the periphery, while records of invertebrates and vertebrates becoming stuck to the top surfaces of froth nests have been made for several species (Figure 6) [81]. However, field observations indicate that some predator types do not get stuck within the froth at all [81,82], while large predators such as snakes simply consume nests whole [76]. Even if the nest does provide some protection against predators, it is also possible that the stark white colouration of the froth nest may actually increase their detectability within the environment, even at great distances, at noted in images taken of nests in the field (Figure 2).
It is clear that froth nesting is not effective against all predator types, which is also apparent in the special case when the predator is oviposited directly into the nest. High offspring mortality rates have been recorded in terrestrial froth nests that have become infested by larvae of ephydrid and phorid flies [67,83,84], and some Loxandrus beetles [85]. Eggs in infested nests can be entirely consumed without the consumption of surrounding froth, indicating that the predatory larvae do not become stuck within the mucus and are able to reach the inner most sections of the nest before hatching occurs. Aquatic froth nests are also known to become infested by invertebrate larvae. For example, the diving beetle Hydaticus parallelus has been recorded ovipositing eggs in L. fletcheri bubble nests, and while the beetle larvae do not prey on the eggs, they hatch synchronously with the tadpoles to prey upon them once they enter the water column [86]. It is possible that such forms of nest parasitism are host species-specific and that the subsequent predation of nests eggs is not simply opportunistic. Indeed, Menin and Giaretta [67] suggest that maggots of an ephydrid fly might be an obligatory predator of exposed foam nests created by leptodactyline species, while Villa [87] more widely suggests there is evidence for close associations between some organisms and amphibian eggs.
If so, it remains to be determined whether froth nesting species have evolved specific physical or biochemical defences against infestation, or the degree to which froth production itself impedes infestation by larvae when compared to un-frothed nests. Nests that are exposed to the air are at higher risk of infestation by flies, and while depositing nests in terrestrial chambers may putatively protect eggs from this particular threat [67], it does not appear to be an effective strategy for protecting them against beetle larvae [88]. The risk of infestation from these invertebrates may only be effectively reduced in species that deposit their eggs in burrows that are subsequently ‘capped’ by mud [57], though predators capable of penetrating such burrows will be able to exploit a considerable food resource in seclusion [89]. Furthermore, infestation levels of nests by some frogflies are known to be temporally variable across the breeding season for some froth nesting species [67], though whether adults adaptively shift their reproduction to avoid periods of high infestation has not been tested. The nests may be composed of proteins that protect against parasitism and egg predation. For example, E. pustulosus nests contain lectin-like proteins (as mentioned earlier), which in plant tissues have been shown to be damaging towards the gut tissue of insects [90,91]. If infestation cannot be avoided, offspring might avoid predation by leaving nests as quickly as possible [92], though at the cost of being exposed to predation threats found outside of the nest.
As previously stated by Downie [16], it is clear that further work is needed to determine whether froth production has multiple predator defence effects and whether it has advantages over other strategies for egg protection, such as plasticity in the timing of egg deposition and hatching or increased egg unpalatability [93,94,95]. However, these alternative devices do not appear to curb egg predation in all species, particularly those with cannibalistic tadpoles which are not easily avoidable [64]. Under these circumstances, placing eggs in froth provides a suitable, alternative strategy that at least partially restricts egg consumption to provide offspring a greater chance of surviving through to hatching whereupon they become mobile and have a greater chance of evading predation.
4.6. Improved Oxygenation
Similar to all other oviparous species, the anuran egg obtains oxygen directly from the surrounding environment [23]. The conundrum here is that each embryo is surrounded by a jelly capsule that acts as a partial barrier to oxygen uptake, which is exacerbated when eggs are deposited together in globular masses [96,97]. Froth nesting may improve oxygen availability in several ways related to the incorporation of air bubbles: (1) For species that oviposit aquatically, it causes the eggs to float at the surface of the water where oxygen levels are generally higher and can approach the eggs from all directions [28], and where they may become mechanically ventilated by rippling of the water’s surface [98]. (2) Each egg becomes encapsulated by a capacious oxygen store that can be exploited throughout development [2,16,23]. (3) The frothed mucus may facilitate oxygen diffusion within the nest structure, which occurs more rapidly in air than in jelly [23]. (4) The froth evenly distributes the eggs and places distance between them, thereby reducing oxygen competition in a manner similar to the function of thick egg capsules in non-froth nesting species [23,97,98].
Given these properties of the froth nest, the incorporation of air bubbles allows for an increase in physical size and number of eggs contained within the mass, allowing eggs to remain viable even when positioned away from the surface and distant from the external environment [23]. This is in contrast to un-frothed egg masses, which must remain thin or contain few embryos given the limit of oxygen diffusion through the jelly, or in masses where eggs are in close contact as the outermost siblings would otherwise intercept a majority of the oxygen [98]. Of course, some froth nesting species deposit their eggs in surface films or below a frothed ‘raft’ [5,28], thereby entirely avoiding the issue of poor oxygen diffusion from the surrounding environment.
While the air incorporated into the froth nest has been considered an additional oxygen supply for embryos [16], it is more likely that the air bubbles act as a primary oxygen source for many froth nesting species and little diffusion from the surrounding environment is required until later embryonic stages or after hatching occurs [23]. This is not surprising, given that anuran froth nests can be composed of up to 77% saturated air [23]. Direct evidence for this has also been obtained by Bastos et al. [9] who showed development to be slower in Scinax rizibilis eggs without froth compared to those surrounded by froth as a result of the reduced oxygen supply. If so, this oxygen store may have facilitated the transition of reproduction to poorly oxygenated environments, such as the capped terrestrial burrows produced by some froth nesting species [57]. The froth nest’s ability to improve oxygenation means that it is likely to be generally beneficial in any terrestrial environment, given that oxygen is not available on the bottom surface of the nest that is resting on substrate [23]. The froth also provides structure to the egg mass that would otherwise collapse as a result of gravity and create anoxic conditions for eggs located away from the surface [23]; a reason why terrestrial eggs masses are typically small and thinly spread [31]. However, the froth nest may not be able to adequately provide for the oxygen demands of hatchlings if they remain within the nest for extended periods. This could explain the swimming behaviour of Leptodactylus bufonius tadpoles that periodically swim to the surface of the nest, which may possibly serve in aeration by distributing oxygen into deeper layers of the nest that become anoxic over time [57].
In aquatic environments, the froth nest also acts as a floating device that suspends the eggs at the water’s surface. Irrespective of the oxygen stored within the nest itself, this improves oxygen availability as surface waters are better oxygenated, particularly in waterbodies that are muddy and/or lacking in significant aquatic vegetation (Seymour 1999). As insufficient oxygen is known to retard development, this is highly beneficial for species that exploit temporary aquatic systems, improving the probability of offspring completing metamorphosis before freestanding water dries up [96]. The displacement of eggs to better oxygenated regions of the water column is likely to be far more crucial for species whose eggs are partially separated from the incorporated air bubbles, as this reduces the capacity of offspring to exploit this stored oxygen supply. For example, L. tasmaniensis, suspend their eggs below a froth raft [5,98], where the eggs are mechanically ventilated by wind agitating the water’s surface. In this species, Seymour and Roberts [98] showed that eggs sank to the bottom of the water column when removed from this raft, causing them to become starved of oxygen. While the benefit of the raft as an oxygen store has not been considered, it could be that the production of froth in this species has evolved only to cause the eggs to become suspended in open water without the need of supporting vegetation and to prevent them from falling to lower levels of water where oxygen levels are lower [98].
5. Additional Adaptive Benefits
Various other functions of froth nesting have been proposed. While they have been given far less attention compared to those stated above, their consideration may provide greater insight into the likely multi-adaptive and flexible role of froth nesting during egg development and post hatching.
5.1. Post-Hatching Refuge
The froth nest creates a ‘safe harbour’ that offspring can exploit for some period after hatching has occurred. This is seen in the terrestrial nester, L. bufonius, whose tadpoles can remain viable within the nest for weeks until they are flushed into nearby water after rainfall [57]. This is highly advantageous given that rainfall across this species’ range can be highly uncertain and intermittent, affording them an opportunity to wait for ideal conditions to arrive. Likewise, C. xerampelina eggs hatch within the centre of froth nests, with the release of perivitelline liquid softening the froth and producing a moist chamber within a shell of dried foam [23]. Downie and Nokhbatolfoghahai [99] suggest that it may be obligatory for the tadpoles of some froth nesting species to remain within the nest until they have reached a stage capable of independent feeding. During this time, the nest provides a protective environment that reduces their vulnerability to future predation by allowing tadpoles to use their yolk reserves to develop until locomotory systems have matured [23,68]. Further, staying within the nest may allow for simultaneous offspring emergence by allowing slower developing siblings to catch up to faster developers, which may be advantageous if mass movement out of the nest prevents desiccation that would otherwise occur during the emergence of smaller, partial cohorts [23].
It is possible that delayed emergence comes with the potential cost of delayed growth if nutrient sources cannot be exploited once yolk stores have been consumed. However, Seymour and Loveridge [23] have shown that C. xerampelina tadpoles can survive without feeding for days after delayed emergence from the froth nest, suggesting that falling yolk reserves is not a limiting factor. If an extended stay within the nest is advantageous or required, the tadpoles of some species are able to release their own mucus secretions to maintain the froth nest around them [100] or aerate the nest to maintain oxygen levels [57], although both activities are energetically expensive and reduce energy that can be invested into growth.
5.2. Insolation
Froth production may shield eggs from sun damage by scattering light, effectively compensating for their sensitivity to ultra-violet (UV) radiation due to a lack of a protective shell [1,101]. This has been proposed by Haddad et al. [4] as a possible benefit of froth nesting in S. rizibilis, and as a reason for the placement of eggs away from the froth nest surface in E. pustulosus [63], which subsequently do not need to be pigmented given this protection. While direct evidence for this property of the froth nest is still to be gathered, other adaptations among anurans have also been suggested to have evolved in response to this environmental sensitivity, such as the common occurrence of darkly pigmented eggs among species that deposit in open habitats [31], adding weight to this hypothesis.
5.3. Inhibition of Development
It has been suggested that the froth functions to inhibit tadpole growth to allow them to persist within the nest for extended periods. Evidence for this comes from the work of Pisano and Del Rio [102] who showed that early elongation was inhibited in L. bufonius and Leptodactylus latinasus tadpoles encased within froth compared to those artificially transferred out of the nest into water. However, Downie [100] argues that this difference in growth rates was not an effect of exposure to froth but simply due to tissue rehydration in individuals exposed to water. Instead, development may be slowed by the tadpoles themselves as a means of preserving yolk reserves if they are required to stay within the nest for extended periods [100].
5.4. Nutrient Source
Laying eggs within froth may provide conditions under which offspring can obtain nutrients rapidly upon hatching. For example, tadpoles that remain within the nest for extended periods after hatching may be able to cannibalise conspecifics that have no means of escape, including non-developing eggs. The tadpoles of several froth nesting species are known to consume trophic eggs that are deliberately left unfertilised by parents to provide nutrients to their offspring upon hatching [17,47,103], though failed eggs may also be exploited with the added benefit of reducing the chance of disease transmission. Downie [100] showed that other forms of cannibalism, such as tadpole-tadpole predation, were a rare occurrence in Leptodactylus fuscus, indicating that it may not be a cost of an extended stay within the froth nest. Hatchlings may also feed on the froth nest itself [104], which is composed of various carbohydrates and proteins [22,105] that should provide for a presumably highly nutritious and easily obtainable meal. Indeed, froth has been shown to be an effective food source in Rhacophorus arboreus [50], but does not appear to be utilised by all froth nesting species, as indicated by the lack of disappearance of froth in some species as tadpoles develop within the nest [57]. It is also been suggested by Downie [16] that froth nests can provide a substrate for microorganisms that can be exploited as a food resource, though the work of Flemming et al. [22] on the microbial resistance of froth nests would dispute this. For other species, froth may simply not be available for consumption if it breaks down and dissipates prior to the tadpoles hatching, which occurs in aquatic species (e.g., [71]).
5.5. Competitive Head-Start
In variable environments, the protective properties of the froth nest allow egg deposition to occur at times when conditions would be unsuitable for most other species. The benefits of this are apparent in terrestrial froth nesting species whose tadpoles at still reliant on freestanding water to complete development, as it allows eggs to be deposited in advance of rainfall filling nearby waterbodies [10,47,106]. This gives offspring a temporal advantage as it allows them to be the first to enter the water and at an advanced developmental stage by the time tadpoles of other species, which are reliant on water to deposit their eggs, have hatched. This may reduce competition for resources and size-limited predation, allow for easy predation of eggs and larvae of hetero-specifics, or result in early metamorphosis that reduces the risk of desiccation in waterbodies with short hydroperiods [1,47,107]. However, this strategy is apparent in both froth and non-froth nesting species that deposit their eggs terrestrially [108], suggesting that terrestrial froth production is one of several routes for reducing reliance on temporary aquatic systems for offspring development.
5.6. Temperature Acclimation
The froth nest may protect eggs from thermal damage while they are adapting to ambient temperatures that are initially above their thermal limit [46]. This may occur as the insulating properties of the nest decline as it begins to break down, thereby gradually exposing the eggs to ambient temperatures and at later embryonic stages that are less susceptible to high temperatures [109,110]. Currently, there have been no studies on the influence of froth production on critical thermal maximum of anuran eggs.
5.7. Microbial Defence
The froth nest may contain specialised proteins that protect against pathogenic microorganisms. For example, many anurans possess skin secretions with bioactive compounds, some of which are antimicrobial and play an important role in innate immune defence [111,112]. These may be transferred onto eggs that are encapsulated in froth produced from skin secretions, which has been recorded in at least one froth nesting species, Polypedates leucomystax [20]. Proteins with anti-microbial activity have also been detected in froth nests produced from oviduct secretions [22]. The transmission of innate defences is highly advantageous for anuran eggs if they are generalised and fast acting against many types of pathogens, creating a protective environment before the tadpole immune system develops [113]. For aquatic species, it may be particularly beneficial for preventing microbial colonisation, considering that the bulk of the nest is formed from surrounding water that contains substantial microbe quantities [22], as well as for species whose offspring remain with the nest for an extended period.
6. The Function of Froth Nest Breakdown
Anuran froth nests are initially stable structures that are mostly resilient to dissociation in water and at high temperatures [10,114], which is due to the activity of various proteins present within the mucus [22,29,115]. However, they do eventually break down at some point prior to or following tadpole hatching. In aquatic nesting species, this is often a gradual process that occurs from the moment oviposition occurs, resulting in the loss of incorporated air bubbles and flattening of the nest over the surface of the water within hours or days before hatching occurs (Figure 7) [71,80,116]. In terrestrial and arboreal nesting species, inner portions of froth may soften but full nest breakdown may not occur until many days or weeks after tadpoles have already hatched into the nest itself [23,117]. While there has been limited investigation into the biochemical processes that catalyse froth breakdown, it is known to occur in terrestrial nests almost immediately upon exposure to a rainfall/flooding event [57], though such environmental degradation due to water exposure does not account for the longevity of aquatic nests. There is some evidence to suggest that the developing embryos play a role in initiating froth breakdown via the release of waste products or deliquescent agents [23,26]. Direct evidence for this comes from records of E. pustulosus and L. fletcheri nests in the field that are resistant to breakdown when eggs are not present [22,71]. Anuran embryos are known to secrete proteolytic enzymes that facilitate hatching via the breakdown of the vitelline membrane [118,119,120], while tadpoles of some froth nesting species produce their own froth to extend the longevity of nesting conditions [100], demonstrating that offspring are capable and often change the conditions of their immediate environment for their advantage.
The breakdown of the froth nest is likely to negatively affect its ability to protect offspring from environmental threats. For aquatic species in particular whose nests break down soon after oviposition, its protective values may only be present during early developmental stages [80]. This was highlighted by Shepard and Caldwell [47] who reported L. labyrinthicus tadpoles desiccating due to the froth disintegrating prior to rainfall and flushing of the nest into nearby water. For aquatic nesting species, the expansion of the nest during breakdown results in an increase in surface area relative to volume that causes eggs, which remain associated with the nest, to come closer to the nest surface-air/water interface where they are more vulnerable to fluctuating conditions and predators [71,121]. This may not be an issue for arboreal nests that maintain a dry outer surface or terrestrial nests that are contained within a burrow where they are not able to dissipate into water [23].
Despite these limitations, froth nest breakdown may be a necessary transformation for two main reasons: (1) Although air bubbles incorporated into the froth nest are a valuable source of oxygen, nest expansion allows for greater diffusion of oxygen from the external environment, thereby compensating for the increased demands by later stage embryos [23,98]. (2) Froth breakdown facilitates easy escape of hatchlings when required, as they would otherwise become trapped within the nest. Additional functions of nest breakdown have also been proposed. Downie [16] reports that collapse of the E. pustulosus froth nest results in the release of water, thereby providing a short-lived source of standing water for tadpoles upon hatching even under dry conditions. Froth breakdown has also been proposed to produce a gliding substrate and path for hatchlings in stranded nests to move to a water source over land [105,122], with direct evidence of this in Physalaemus nattereri tadpoles that suffer increased mortality rates when such mucus trails end short of nearby water [123]. As suggested by Gould [79], a shift in froth nest structure thereby accommodates for the changing needs of the offspring in various ways. It does not appear that females of any froth nesting species supply eggs with additional frothed mucus to compensate for nest breakdown, though C. xerampelina females are known to return to their nests following the night of construction to add an additional layer of foam that enlarges the nest and likely improves its protective properties [70]. However, for most froth nesting species, it remains to be determined whether similar behaviours or forms of parental care are provided to offspring during development while in the nest.
7. Communal Froth Nesting
Communal nesting is a cooperative behavioural strategy in which the eggs or egg clutches of multiple mating partners are placed together in the same vicinity or together in direct physical contact. This is in contrast to grouped spawning, where multiple males may join a mating pair to fertilise a single egg clutch [124,125]. Communal nesting is exhibited by anurans that produce frothed [36,80,126] or un-frothed nests [126], whereby nests are laid relatively synchronously and in physical association to form communal masses. While the formation of these masses has been suggested to be an unintentional result of limited space or mass aggregation of reproducing adults [123], this does not account for their occurrence at sites where space is not a limiting factor or when adult densities are low. Observations have also been made in both L. fletcheri (J. Gould, unpubl. data) and C. xerampelina [70] showing that females do not always choose to lay communally, suggesting that aggregating nests is a deliberate behaviour in at least some froth nesting species (Figure 8).
The communal deposition of froth nests may represent a behavioural strategy for overcoming energy and time constraints related to froth production. If so, it may have two benefits: (1) Mating pairs are unlikely to deliver mucus to nests after initial bouts of nest formation. Joining onto a communal mass allows adult pairs to exploit pre-existing froth to save nest material and energy expenditure, while also ensuring that eggs are adequately deposited into froth even if mating pairs are deficient in mucus production and/or disturb their own froth during nest formation [63,80]. (2) The protective properties of the froth are extended given a reduction in the surface area relative to volume of each nest included within the mass, without compromising oxygen availability to the eggs (Figure 9).
Currently, no evidence has been obtained to suggest that there is a decrease in nesting investment in later adult pairs that utilise communal masses. In contrast, direct evidence has been obtained to suggest communal nesting does indeed provide additional protection to offspring. In particular, Zina [36] showed that communal nesting extended survival times of E. pustulosus eggs when exposed to the absence of freestanding water by increasing the resistance of each froth nest to desiccation. This has also been proposed for communal masses produced by Physalaemus gracilis [127]. Communal nesting could also have thermal benefits similar to those recorded in communal masses of un-frothed nests [126], increasing or maintaining optimal temperatures for eggs in the centre and thereby allowing for faster developmental rates [42]. While communal nesting may simply reduce the risk of predation for each offspring by generating a selfish herd effect [128], such an effect would be generated at localised sites where multiple solitary nests were oviposited. Instead, it has been proposed by Gould et al. [79] that communal nesting may provide additional protection from predation by acting as a larger physical barrier (Figure 9). Currently, direct evidence for the thermal and defence properties of communal nesting are lacking.
The capacity for communal nesting to further buffer eggs from sub-optimal conditions is dependent on the number of nests that are included and their configuration, as this influences the proportion of each nest that remains exposed. For example, aquatic communal masses created by L. fletcheri often contain between two and four nests yet masses of more than 20 nests have been recorded (J. Gould, unpubl. data). Irrespective of the size of the mass, these nests are interconnected side-by-side to form a floating ‘raft’, similar to that produced by Pleurodema diplolistris [80]. This configuration may offer few thermal or desiccation benefits given that much of the top and bottom surfaces of each nest remains exposed, but likely provides greater protection for inner nests from predators such as tadpoles that often feed around the outer edge (Figure 5). Likewise, the arboreal communal masses created by C. xerampelina are built from two or three gravid females, yet masses constructed by more than 40 females has been recorded [70]. These nests are also placed side-by-side, but as the nests are placed out of water, it results in the formation of a much more globular communal mass which may offer greater thermal and moisture benefits. For all communal froth nesting species, there may be an advantage for mating pairs to be the first to oviposit at sites so that their offspring are within the centre of communal masses. However, this may come with associated risks, as there is no guarantee that subsequent mating pairs oviposit at the same site to establish such masses. There may also be associated costs of laying communally, such as making nests easier to locate by predators given their exaggerated size [63] and increased risk of disease transmission, which could explain the presence of plasticity in nesting behaviour in some populations that produce both solitary and communal nests.
8. Testing the Properties of the Froth Nest
It is surprising that more evidence of the adaptive functions of froth nesting has not been obtained, given that it is a reproductive strategy exhibited by species across several continents. Furthermore, froth production has been shown to occur under laboratory conditions [71], thereby making laboratory based studies of the properties of froth production possible for at least some species. Perhaps it is an aspect of anuran reproduction that remains overlooked, and certainly an area of study that continues to be lacking in robust datasets and replication. To ensure future studies do not suffer from the limitations and potential flaws in study design that have met past contributions in this field, I have provided protocols that should be considered when investigating some hypothesised functions of the froth nest.
8.1. Temperature
For studies conducted in the field under natural conditions, nest temperatures should be measured near the top surface, at points around the perimeter, at the base and near the core. Ambient temperatures must also be obtained at various distances from the surface of the nest, including substrate or water temperature below and air temperature above the nest [33]. The influence of solar radiation on nest temperature must also be considered for species that oviposit above ground, which may be done by deliberately and continuously shielding a portion of nests during the day. Temperatures should ideally be repeatedly made for each nest across several points during both day and night, as well as across the developmental period to account for likely changes in the thermal profile of the froth as it ages and breaks down [16]. By closely following and comparing surface and core nest temperatures with ambient temperatures and solar exposure, the capacity for the nest to insulate eggs from external fluctuations in thermal conditions or to heat eggs to an optimal thermal regime can be evaluated. Of course, the effect of other environmental variables such as humidity and exposure to wind should be taken into consideration. For species that oviposit aquatically, the thermal effect of nests becoming stranded on land should also be investigated.
8.2. Moisture
Feld observations of embryos surviving in froth nests under sub-optimal moisture conditions, such as aquatic nests that have become stranded out of water, as well as the successful hatching of tadpoles when moisture levels are restored, are valuable pieces of evidence for the capacity of the froth nest to protect against desiccation-induced mortality. An initial step in testing the anti-desiccation property of the froth nest might include exposing nests to sub-optimal moisture (e.g., out of water) under laboratory conditions that have been controlled for temperature, humidity, wind and solar radiation, and weighing the nests to calculate rates of evaporative water loss over time [36]. Important considerations during this process will be the effect of nest size, volume relative to surface area and possible expansion of the nest over time. Over this period, the proportion of eggs within each nest that have desiccated should be recorded, along with the time it takes for total egg clutch mortality to occur. In order to obtain valid counts, sections of the nest can be excised to compare survival rates between eggs located on the periphery and near the core, with sections subsequently repositioned to preserve the integrity of the nest between measures.
8.3. Predation
The inability for predators to reach eggs in certain regions of the froth nest should be noted as direct evidence of its defence benefit. For species that oviposit aquatically, egg consumption is likely to be restricted to the bottom half of the nest as this is the only region accessible to aquatic predators, though terrestrial and flying predators can access the top surface of the nest floating above the water. Terrestrial nests are more likely to be attacked from the top half, as the bottom half is pressed against the substrate. Across all types of froth nests, it is likely that peripheral eggs are at immediate risk of predation compared to those near the core, as they are exposed and easily accessible. The rate and location of egg consumption should therefore be monitored over time for each nest, as well as whether both egg and nest material are being consumed. The proportion of eggs that successfully make it to the hatching stage should be then recorded, taking note of the effect of predator density on survival.
8.4. Oxygen
For nests that are laid aquatically, oxygen levels at the water’s surface should be compared to those throughout the water column. Fluctuations in oxygen levels throughout the day should also be compared at and below the water’s surface. Higher oxygen levels at the surface would suggest that floating egg clutches would be at an advantage to those laid below the surface in un-frothed nest or solitarily. If this was apparent, froth nests could be collected from the field and exposed to water at varying degrees of oxygenation to test whether environmental oxygen levels influenced development. To test whether the oxygen stored within the froth nest can sustain eggs without diffusion from the surrounding environment, oxygen levels within the nests should be measured [23]. Nests could also be collected from the field and placed under anoxic conditions to monitor embryo development.
8.5. Comparing Frothed and Un-Frothed Nests
What appears to be lacking from most studies is evidence of the relative fitness consequence of being surrounded by froth. Development times and survival rate of eggs in froth nests must be compared to that of eggs in the absence of froth. This may be achieved by physically removing eggs from the froth nest. However, it must be appreciated that such a comparison would not take into consideration the ability of un-frothed mucus to also provide protection against exposure to sub-optimal conditions and, as such, would not indicate the true effect of mucus aeration. This may only be achieved in species that produce both frothed and un-frothed nests, such as L. tasmaniensis [40], as most froth nesting species aerate the mucus at the same time it is being excreted, which would make ascertaining un-frothed mucus relatively difficult. If this cannot be achieved, then differences in developmental times and survival rates of eggs from froth nesting species can be compared to that of eggs of non-frothing species under various environmental conditions. If possible, intraspecific variations in froth nests should also be examined, including the possibility for plasticity in the size and structure of froth nests within populations to accommodate for environmental conditions faced during egg deposition, particularly for species that occupy variable environments.
9. Conclusions
The widespread occurrence of froth nesting among the anuran amphibians and its adaptability to many ecological situations suggests that it is an effective solution to the challenge of depositing eggs into an external environment, offering a degree of protection from an array of suboptimal conditions that may arise at any point during development. By aerating mucus secretions, parents are performing a relatively simple behavioural task but one that leads to the protection of their offspring for an extended period of time after oviposition occurs, even after they have moved away. To this effect, froth nesting should be considered a form of ‘remote’ parental care, which may be more advantageous than other forms of care in which the parents must remain present to impart that protection (e.g., parental brooding; [129]). While further experimental evidence of the benefits of froth nesting are required, it is clear that its functions are many, with strong evidence of its thermal and anti-desiccation and oxygenation benefits in particular. It is also apparent that froth nesting can also be multi-adaptive, serving to protect offspring from various threats at the same time. This versatility means that froth nesting may have been a key reproductive innovation that helped facilitate the partial or complete withdrawal of egg development from water in several lineages, thereby providing an opportunity for previously unused oviposition sites to be exploited.
Funding
This research received no external funding.
Acknowledgments
I would like to thank Michael Mahony for providing feedback.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Duellman, W.; Trueb, L. Biology of Amphibians; McGrawHill: New York, NY, USA, 1986. [Google Scholar]
- Altig, R.; McDiarmid, R.W. Morphological Diversity and Evolution of Egg and Clutch Structure in Amphibians. Herpetol. Monogr. 2007, 21, 1–32. [Google Scholar] [CrossRef]
- Littlejohn, M. The breeding biology of the Baw Baw frog Philoria frosti Spencer. Proc. Linnaean Soc. N. S. Wales 1963, 88, 273–276. [Google Scholar]
- Haddad, C.F.B., Jr.; Pombal, J.P.P.; Gordo, M. Foam Nesting in a Hylid Frog (Amphibia, Anura). J. Herpetol. 1990, 24, 225–226. [Google Scholar] [CrossRef] [Green Version]
- Tyler, M.J.; Davies, M. Foam Nest Construction by Australian Leptodactylid Frogs (Amphibia, Anura, Leptodactylidae). J. Herpetol. 1979, 13, 509. [Google Scholar] [CrossRef]
- Hödl, W. An Analysis of Foam Nest Construction in the Neotropical Frog Physalaemus ephippifer (Leptodactylidae). Copeia 1990, 1990, 547–554. [Google Scholar] [CrossRef]
- Kadadevaru, G.G.; Kanamadi, R.D. Courtship and nesting behaviour of the Malabar gliding frog, Rhacophorus malabaricus (Jerdon, 1870). Curr. Sci. 2000, 79, 377–380. [Google Scholar]
- Frost, D.R. Amphibian Species of the World; Allen Press and Association of Systematics Collections: Lawrence, KS, USA, 1985. [Google Scholar]
- Bastos, R.P.; Haddad, C.F.; Pombal, J.P., Jr. Foam nest in Scinax rizibilis (Amphibia: Anura: Hylidae). Zoologia 2010, 27, 881–886. [Google Scholar] [CrossRef]
- Heyer, W.R. The adaptive ecology of the species groups of the genus Leptodactylus (Amphibia, Leptodactylidae). Evolution 1969, 23, 421–428. [Google Scholar] [CrossRef]
- Martin, A. Parallel evolution in the adaptive ecology of leptodactylid frogs of South America and Australia. Evolution 1970, 24, 643–644. [Google Scholar] [CrossRef]
- Fowler, J.E.; Kleinteich, T.; Franz, J.; Jaye, C.; Fischer, D.A.; Gorb, S.; Weidner, T.; Baio, J.E. Surface chemistry of the frog sticky-tongue mechanism. Biointerphases 2018, 13, 06E408. [Google Scholar] [CrossRef]
- Ramsey, J.P.; Reinert, L.K.; Harper, L.K.; Woodhams, D.C.; Rollins-Smith, L.A. Immune Defenses against Batrachochytrium dendrobatidis, a Fungus Linked to Global Amphibian Declines, in the South African Clawed Frog, Xenopus laevis. Infect. Immun. 2010, 78, 3981–3992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keogh, J.P.; Allen, A.; Garner, A. Relationship between gastric mucus synthesis, secretion and surface gel erosion measured in amphibian stomach In Vitro. Clin. Exp. Pharmacol. Physiol. 1997, 24, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Toledo, R.; Jared, C. Cutaneous adaptations to water balance in amphibians. Comp. Biochem. Physiol. Part A Physiol. 1993, 105, 593–608. [Google Scholar] [CrossRef]
- Downie, J. Functions of the foam in the foam-nesting leptodactylid Physalaemus pustulosus. Herpetol. J. 1988, 1, 302–307. [Google Scholar]
- Silva, W.R.; Giaretta, A.A.; Facure, K.G. On the natural history of the South American pepper frog, Leptodactylus labyrinthicus (Spix, 1824) (Anura: Leptodactylidae). J. Nat. Hist. 2005, 39, 555–566. [Google Scholar] [CrossRef]
- Hero, J.-M.; Galatti, U. Characteristics Distinguishing Leptodactylus pentadactylus and L. knudseni in the Central Amazon Rainforest. J. Herpetol. 1990, 24, 226–228. [Google Scholar] [CrossRef]
- Meegaskumbura, M.; Senevirathne, G.; Biju, S.D.; Garg, S.; Meegaskumbura, S.; Pethiyagoda, R.; Hanken, J.; Schneider, C.J. Patterns of reproductive-mode evolution in Old World tree frogs (Anura, Rhacophoridae). Zoöl. Scr. 2015, 44, 509–522. [Google Scholar] [CrossRef]
- Shahrudin, S.; Ismail, M.N.; Kwan, S.H.; Najimudin, N. Ecology and Protein Composition of Polypedates leucomystax (Gravenhorst, 1829) (Anura: Rhacophoridae) Foam Nests from Peninsular Malaysia. Annu. Res. Rev. Biol. 2017, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Furness, A.I.; McDiarmid, R.W.; Heyer, W.R.; Zug, G.R. Oviduct Modifications in Foam-Nesting Frogs, with Emphasis on the Genus Leptodactylus (Amphibia, Leptodactylidae). S. Am. J. Herpetol. 2010, 5, 13–29. [Google Scholar] [CrossRef]
- Fleming, R.I.; MacKenzie, C.D.; Cooper, A.; Kennedy, M.W. Foam nest components of the túngara frog: A cocktail of proteins conferring physical and biological resilience. Proc. R. Soc. B Biol. Sci. 2009, 276, 1787–1795. [Google Scholar] [CrossRef] [Green Version]
- Seymour, R.S.; Loveridge, J.P. Embryonic and larval respiration in the arboreal foam nests of the African frog Chiromantis xerampelina. J. Exp. Biol. 1994, 197, 31–46. [Google Scholar]
- Heyer, W.R.; Rand, A.S. Foam Nest Construction in the Leptodactylid Frogs Leptodactylus pentadactylus and Physalaemus pustulosus (Amphibia, Anura, Leptodactylidae). J. Herpetol. 1977, 11, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.J. The túngara Frog: A Study in Sexual Selection and Communication; University of Chicago Press: Chicago, IL, USA, 1985. [Google Scholar]
- Coe, M. Observations on the ecology and breeding biology of the genus Chiromantis (Amphibia: Rhacophoridae). J. Zoöl. 1974, 172, 13–34. [Google Scholar] [CrossRef]
- Tyler, M. Australian Frogs; Viking O’Neil: Melbourne, Australia, 1989. [Google Scholar]
- Haddad, F.; Hödl, W. New reproductive mode in anurans: Bubble nest in Chiasmocleis leucosticta (Microhylidae). Copeia 1997, 1997, 585–588. [Google Scholar] [CrossRef]
- Cooper, A.; Kennedy, M.W.; Fleming, R.I.; Wilson, E.H.; Videler, H.; Wokosin, D.L.; Su, T.-J.; Green, R.J.; Lu, J.R. Adsorption of Frog Foam Nest Proteins at the Air-Water Interface. Biophys. J. 2005, 88, 2114–2125. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.; Kennedy, M.W. Biofoams and natural protein surfactants. Biophys. Chem. 2010, 151, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Salthe, S.N.; Duellman, W.E. Quantitative constraints associated with reproductive mode in anurans. In Evolutionary Biology of the Anurans; Vial, J.L., Ed.; University of Missouri Press: Columbia, MO, USA, 1973; pp. 229–249. [Google Scholar]
- Heyer, W.R. A preliminary analysis of the intergeneric relationships of the frog family Leptodactylidae. Smithson. Contrib. Zoöl. 1975, 199, 1–55. [Google Scholar] [CrossRef]
- Méndez-Narváez, J.; Flechas, S.V.; Amézquita, A. Foam Nests Provide Context-Dependent Thermal Insulation to Embryos of Three Leptodactylid Frogs. Physiol. Biochem. Zoöl. 2015, 88, 246–253. [Google Scholar] [CrossRef]
- Pulz, R. Thermal and water relations. In Ecophysiology of Spiders; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 1987; pp. 26–55. [Google Scholar]
- Duellman, W. Reproductive modes in anuran amphibians: Phylogenetic significance of adaptive strategies. S. Afr. J. Sci. 1985, 81, 174–178. [Google Scholar]
- Zina, J. Communal nests in Physalaemus pustulosus (Amphibia: Leptodactylidae): Experimental evidence for female oviposition preferences and protection against desiccation. Amphibia-Reptilia 2006, 27, 148–150. [Google Scholar] [CrossRef] [Green Version]
- Muedeking, M.H.; Heyer, W.R. Descriptions of eggs and reproductive patterns of Leptodactylus pentadactylus (Amphibia: Leptodactylidae). Herpetologica 1976, 32, 137–139. [Google Scholar]
- Davis, S.; Davis, R.; James, A.; Talin, B. Reproductive behavior and larval development of Leptodactylus fallax in Dominica, West Indies. Herpetol. Rev. 2000, 31, 217–220. [Google Scholar]
- Seymour, R.S.; Mahony, M.J.; Knowles, R. Respiration of embryos and larvae of the terrestrially breeding frog Kyarranus loveridgei. Herpetologica 1995, 51, 369–376. [Google Scholar]
- Roberts, J. Call Differentiation in the Limnodynastes Tasmaniensis Complex (Anura: Leptodactylidae); University of Adelaide: Adelaide, Australia, 1976. [Google Scholar]
- Wells, K.D. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicagi, IA, USA, 2007. [Google Scholar]
- Dobkin, D.S.; Gettinger, R.D. Thermal Aspects of Anuran Foam Nests. J. Herpetol. 1985, 19, 271. [Google Scholar] [CrossRef]
- Hassinger, D.D. Notes on the thermal properties of frog eggs. Herpetologica 1970, 26, 49–51. [Google Scholar]
- Salthe, S.; Mecham, J. Reproductive and courtship patterns. In Physiology of the Amphibia; Elsevier: Amsterdam, The Netherlands, 1974; Volume 2, pp. 309–521. [Google Scholar]
- Rand, A. Physalaemus pustulosus (Rana, Sapito Tingara, Foam Toad, Mud-puddle Frog). In Costa Rican Natural History; Janzen, D., Ed.; University of Chicago Press: Chicago, IA, USA, 1983; pp. 412–415. [Google Scholar]
- Gorzula, S. Foam nesting in leptodactylids: A possible function. Br. J. Herpetoloy 1977, 5, 657–659. [Google Scholar]
- Shepard, D.B.; Caldwell, J.P. From Foam to Free-living: Ecology of Larval Leptodactylus Labyrinthicus. Copeia 2005, 2005, 803–811. [Google Scholar] [CrossRef]
- Carroll, E.J., Jr. Thermal tolerance and heat shock protein synthesis during development in the anuran Lepidobatrachus laevis. Dev. Growth Differ. 1996, 38, 9–14. [Google Scholar] [CrossRef]
- Seale, D.B. Physical Factors Influencing Oviposition by the Woodfrog, Rana sylvatica, in Pennsylvania. Copeia 1982, 1982, 627–635. [Google Scholar] [CrossRef]
- Kusano, T.; Sakai, A.; Hatanaka, S. Ecological functions of the foam nests of the Japanese treefrog, Rhacophorus arboreus (Amphibia, Rhacophoridae). Herpetol. J. 2006, 16, 163–169. [Google Scholar]
- Liwanag, H.E.M.; Berta, A.; Dos Santos, D.C.; Abney, M.; Williams, T.M. Morphological and thermal properties of mammalian insulation: The evolution of fur for aquatic living. Biol. J. Linn. Soc. 2012, 106, 926–939. [Google Scholar] [CrossRef] [Green Version]
- Bock, W.J. Explanatory history of the origin of feathers. Am. Zool. 2000, 40, 478–485. [Google Scholar] [CrossRef]
- Boukhriss, M.; Zhani, K.; Ghribi, R. Study of thermophysical properties of a solar desalination system using solar energy. Desalination Water Treat. 2013, 51, 1290–1295. [Google Scholar] [CrossRef]
- Beattie, R.C. A physico-chemical investigation of the jelly capsules surrounding eggs of the Common frog (Rana temporaria temporaria). J. Zoöl. 1980, 190, 1–25. [Google Scholar] [CrossRef]
- Zweifel, R.G. Upper thermal tolerances of anuran embryos in relation to stage of development and breeding habits. Am. Mus. Novit. 1977, 2617, 1–21. [Google Scholar]
- Bernal, M.H.; Lynch, J.D. Thermal Tolerance in Anuran Embryos with Different Reproductive Modes: Relationship to Altitude. Sci. World J. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Philibosian, R.; Ruibal, R.; Shoemaker, V.H.; McClanahan, L.L. Nesting behavior and early larval life of the frog Leptodactylus bufonius. Herpetologica 1974, 30, 381–386. [Google Scholar]
- Touchon, J.C.; Warkentin, K.M. Reproductive mode plasticity: Aquatic and terrestrial oviposition in a treefrog. Proc. Natl. Acad. Sci. USA 2008, 105, 7495–7499. [Google Scholar] [CrossRef] [Green Version]
- Spieler, M.; Linsenmair, K.E. Choice of optimal oviposition sites by Hoplobatrachus occipitalis (Anura: Ranidae) in an unpredictable and patchy environment. Oecologia 1997, 109, 184–199. [Google Scholar] [CrossRef]
- Hödl, W. Foam nest construction in South American leptodactylid frogs. In Studies in Herpetology; Roek, Z., Ed.; Charles University: Prague, Czech Republic, 1986; pp. 565–570. [Google Scholar]
- Romero-Carvajal, A.; Sáenz-Ponce, N.; Venegas-Ferrín, M.; Almeida-Reinoso, D.; Lee, C.; Bond, J.; Ryan, M.J.; Wallingford, J.B.; Del Pino, E.M. Embryogenesis and laboratory maintenance of the foam-nesting túngara frogs, genus Engystomops(= Physalaemus). Dev. Dyn. 2009, 238, 1444–1454. [Google Scholar] [CrossRef] [Green Version]
- Narváez, A.; Santiago, R. Spawning behaviour of Engystomops pustulatus (Anura: Leptodactylidae). J. Nat. Hist. 2017, 5, 267–275. [Google Scholar] [CrossRef]
- Dalgetty, L.; Kennedy, M.W. Building a home from foam-Túngara frog foam nest architecture and three-phase construction process. Biol. Lett. 2010, 6, 293–296. [Google Scholar] [CrossRef]
- Magnusson, W.E.; Hero, J.-M. Predation and the evolution of complex oviposition behaviour in Amazon rainforest frogs. Oecologia 1991, 86, 310–318. [Google Scholar] [CrossRef]
- Roberts, J. Terrestrial Egg Deposition and Direct Development in Arenophyrne rotunda Tyler, a Myobatrachid Frog from Coastal Sand Dunes at Shark Bay, W. A. Wildl. Res. 1984, 11, 191–200. [Google Scholar] [CrossRef]
- Hartmann, P.; Hartmann, M.; Haddad, C. Visual signaling and reproductive biology in a nocturnal treefrog, genus Hyla (Anura: Hylidae). Amphibia-Reptilia 2004, 25, 395–406. [Google Scholar] [CrossRef]
- Menin, M.; Giaretta, A.A. Predation on foam nests of leptodactyline frogs (Anura: Leptodactylidae) by larvae of Beckeriella niger (Diptera: Ephydridae). J. Zoöl. 2003, 261, 239–243. [Google Scholar] [CrossRef]
- Downie, J. Functions of the foam in foam-nesting leptodactylids: The nest as a post-hatching refuge in Physalaemus pustulosus. Herpetol. J. 1993, 3, 35–42. [Google Scholar]
- Moore, J.A. The frogs of eastern New South Wales. Bull. Am. Mus. Nat. Hist. 1961, 121, 149–386. [Google Scholar]
- Jennions, M.D. Breeding Behaviour of the Foam Nest Frog, Chiromantis Xerampelina: Sperm Competition and Polyandry; University of the Witwatersrand: Johannesburg, South Africa, 1992. [Google Scholar]
- Gould, J. Build me up to break me down: Frothed spawn in the sandpaper frog, Lechriodus fletcheri, is formed by female parents and later broken down by their offspring. Aust. J. Zoöl. 2019, 67, 153–161. [Google Scholar] [CrossRef]
- Ryan, M.J.; Bartholomew, G.A.; Rand, A.S. Energetics of Reproduction in a Neotropical Frog, Physalaemus Pustulosus. Ecology 1983, 64, 1456–1462. [Google Scholar] [CrossRef] [Green Version]
- Barrio, A. El género Physalaemus en la Argentina (Anura, Leptodactylidae). Physis 1965, 25, 421–488. [Google Scholar]
- Wager, V. The Frogs of South Africa; Purnell and Sons: Johannesburg, South Africa, 1965. [Google Scholar]
- Crump, M.L. Parental Care among the Amphibia. Adv. Study Behav. 1996, 25, 109–144. [Google Scholar] [CrossRef]
- Lingnau, R.; Di-Bernardo, M. Predation on foam nests of two leptodactylid frogs by Solenopsis sp. (Hymenoptera, Formicidae) and Liophis miliaris (Serpentes, Colubridae). Biociências 2006, 14, 223–224. [Google Scholar]
- Waldman, B. Sibling association among schooling toad tadpoles: Field evidence and implications. Anim. Behav. 1982, 30, 700–713. [Google Scholar] [CrossRef]
- Polis, G.A.; Myers, C.A. A Survey of Intraspecific Predation among Reptiles and Amphibians. J. Herpetol. 1985, 19, 99–107. [Google Scholar] [CrossRef]
- Gould, J.; Clulow, J.; Clulow, S. Food, not friend: Tadpoles of the sandpaper frog (Lechriodus fletcheri) cannibalise conspecific eggs as a food resource in ephemeral pools. Ethology 2019, 126, 486–491. [Google Scholar] [CrossRef]
- Hödl, W. Reproductive behaviour in the neotropical foam-nesting frog Pleurodema diplolistris (Leptodactylidae). Amphibia-Reptilia 1992, 13, 263–274. [Google Scholar] [CrossRef]
- Villa, J.; McDiarmid, R.W.; Gallardo, J.M. Arthropod predators of leptodactylid frog foam nests. Brenesia 1982, 19, 577–589. [Google Scholar]
- Drewes, R.C.; Altig, R. Anuran egg predation and heterocannibalism in a breeding community of East African frogs. Trop. Zoöl. 1996, 9, 333–347. [Google Scholar] [CrossRef]
- Yorke, C.D. Survival of Embryos and Larvae of the Frog Polypedates leucomystax in Malaysia. J. Herpetol. 1983, 17, 235–241. [Google Scholar] [CrossRef]
- Davis, R.A.; Disney, R.H.L. Natural history and description of Aphiura breviceps Schmitz, a scuttle fly (Diptera: Phoridae) whose larvae prey on the eggs of frogs (Anura: Myobatrachidae) in Western Australia. Aust. J. Entomol. 2003, 42, 18–21. [Google Scholar] [CrossRef]
- Carvalho, T.; Facure, K.; Giaretta, A. Predation upon eggs of the terrestrial foam-nesting frog Leptodactylus fuscus (Leptodactylidae) by larvae of the ground beetle Loxandrus oophagus (Carabidae: Loxandrini). Herpetol. Notes 2012, 5, 319–322. [Google Scholar]
- Gould, J.; Valdez, J.W.; Clulow, J.; Clulow, S. Diving beetle offspring oviposited in amphibian spawn prey on the tadpoles upon hatching. Entomol. Sci. 2019, 22, 393–397. [Google Scholar] [CrossRef]
- Villa, J. Frogflies’ from Central and South America with notes on other organisms of the amphibian egg microhabitat. Brenesia 1980, 17, 49–68. [Google Scholar]
- Costa, C.; Vanin, S.A.; De Carvalho, T.R. A new Brazilian species of Loxandrus LeConte, 1852, with description of immatures and notes on natural history (Coleoptera: Carabidae: Loxandrini). Zootaxa 2011, 2745, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Downie, J.; Disney, R.; Collins, L.; Hancock, E. A new species of Megaselia (Diptera, Phoridae) whose larvae prey upon the eggs of Leptodactylus fuscus (Anura, Leptodactylidae). J. Nat. Hist. 1995, 29, 993–1003. [Google Scholar] [CrossRef]
- Chen, M.-S. Inducible direct plant defense against insect herbivores: A review. Insect Sci. 2008, 15, 101–114. [Google Scholar] [CrossRef]
- Dutta, I.; Saha, P.; Majumder, P.; Sarkar, A.; Chakraborti, D.; Banerjee, S.; Das, S. The efficacy of a novel insecticidal protein, Allium sativum leaf lectin (ASAL), against homopteran insects monitored in transgenic tobacco. Plant Biotechnol. J. 2005, 3, 601–611. [Google Scholar] [CrossRef]
- Alcala, A.C.; Brown, W.C. Early life history of two Philippine frogs with notes on deposition of eggs. Herpetologica 1956, 12, 241–246. [Google Scholar]
- Licht, L.E. Unpalatability and toxicity of toad eggs. Herpetologica 1968, 24, 93–98. [Google Scholar]
- Gunzburger, M.S.; Travis, J. Critical Literature Review of the Evidence for Unpalatability of Amphibian Eggs and Larvae. J. Herpetol. 2005, 39, 547–571. [Google Scholar] [CrossRef]
- Vonesh, J.R. Egg predation and predator-induced hatching plasticity in the African reed frog, Hyperolius spinigularis. Oikos 2005, 110, 241–252. [Google Scholar] [CrossRef]
- Seymour, R.S. Respiration of Aquatic and Terrestrial Amphibian Embryos. Am. Zoöl. 1999, 39, 261–270. [Google Scholar] [CrossRef]
- Seymour, R.S. Oxygen diffusion through the jelly capsules of amphibian eggs. Isr. J. Zool. 1994, 40, 493–506. [Google Scholar]
- Seymour, R.S.; Roberts, J.D. Embryonic Respiration and Oxygen Distribution in Foamy and Nonfoamy Egg Masses of the Frog Limnodynastes tasmaniensis. Physiol. Zoöl. 1991, 64, 1322–1340. [Google Scholar] [CrossRef]
- Downie, J.; Nokhbatolfoghahai, M. Presence and absence of the cement gland in foamnesting leptodactylids (Anura: Leptodactylidae): Implications for the transition to terrestrial development. Herpetol. J. 2006, 16, 77–81. [Google Scholar]
- Downie, J. Developmental arrest in Leptodactylus fuscus tadpoles (Anura: Leptodactylidae). I: Descriptive analysis. Herpetol. J. 1994, 4, 29–38. [Google Scholar]
- Blaustein, A.R.; Hoffman, P.D.; Hokit, D.G.; Kiesecker, J.M.; Walls, S.C.; Hays, J.B. UV repair and resistance to solar UV-B in amphibian eggs: A link to population declines? Proc. Natl. Acad. Sci. USA 1994, 91, 1791–1795. [Google Scholar] [CrossRef] [Green Version]
- Pisano, A.; Del Rio, A. New biological properties in the foamy jelly of amphibians. Arch. Zool. Ital. 1968, 53, 189–201. [Google Scholar]
- Gibson, R.C.; Buley, K.R. Maternal Care and Obligatory Oophagy in Leptodactylus fallax: A New Reproductive Mode in Frogs. Copeia 2004, 2004, 128–135. [Google Scholar] [CrossRef]
- Tanaka, S.; Nishihira, M. Foam nest as a potential food source for anuran larvae: A preliminary experiment. J. Ethol. 1987, 5, 86–88. [Google Scholar] [CrossRef]
- Kabisch, K.; Herrmann, H.-J.; Klossek, P.; Luppa, H.; Brauer, K. Foam gland and chemical analysis of the foam of Polypedates leucomystax (Gravenhorst 1829) (Anura: Rhacophoridae). Russ. J. Herpetol. 1998, 5, 10–14. [Google Scholar]
- Lee, A. Studies in Australian amphibia II. Taxonomy, ecology and evolution of the genus Heleioporus Gray (Anura: Leptodactylidae). Aust. J. Zoöl. 1967, 15, 367–439. [Google Scholar] [CrossRef]
- Crump, M.L. Reproductive strategies in a tropical anuran cominunity. Univ. Kans. Mus. Nat. Hist. Misc. Publ. 1974, 61, 1–68. [Google Scholar]
- Savage, R.M. The Ecology and Life History of the Common frog Rana Temporaria Temporaria; Pitman and Sons: London, UK, 1961. [Google Scholar]
- Brown, H.A. High Temperature Tolerance of the Eggs of a Desert Anuran, Scaphiopus hammondii. Copeia 1967, 1967, 365–370. [Google Scholar] [CrossRef]
- Turriago, J.; Parra, C.; Bernal, M. Upper thermal tolerance in anuran embryos and tadpoles at constant and variable peak temperatures. Can. J. Zoöl. 2015, 93, 267–272. [Google Scholar] [CrossRef]
- Kreil, G. Antimicrobial Peptides from Amphibian Skin: An Overview. Novartis Found. Symp. 2007, 186, 77–90. [Google Scholar] [CrossRef]
- Yang, X.; Lee, W.-H.; Zhang, Y. Extremely Abundant Antimicrobial Peptides Existed in the Skins of Nine Kinds of Chinese Odorous Frogs. J. Proteome Res. 2011, 11, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Walke, J.B.; Harris, R.N.; Reinert, L.K.; Rollins-Smith, L.A.; Woodhams, D.C. Social Immunity in Amphibians: Evidence for Vertical Transmission of Innate Defenses. Biotropica 2011, 43, 396–400. [Google Scholar] [CrossRef]
- Hissa, D.C.; Bezerra, W.M.; Freitas, C.D.T.; Ramos, M.V.; Lopes, J.L.S.; Beltramini, L.M.; Roberto, I.J.; Cascon, P.; Melo, V.M.M. Frog Foam Nest Protein Diversity and Synthesis. J. Exp. Zoöl. Part A Ecol. Genet. Physiol. 2016, 325, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Faivovich, J.; Ferraro, D.P.; Basso, N.G.; Haddad, C.F.; Rodrigues, M.T.; Wheeler, W.C.; Lavilla, E.O. A phylogenetic analysis of Pleurodema (Anura: Leptodactylidae: Leiuperinae) based on mitochondrial and nuclear gene sequences, with comments on the evolution of anuran foam nests. Cladistics 2012, 28, 460–482. [Google Scholar] [CrossRef]
- Tyler, M.J.; Crook, G.A. Reproductive biology of the frogs of the Magela Creek system, Northern Territory. Rec. S. Aust. Mus. 1983, 18, 415–440. [Google Scholar]
- Agostinho, C.; Foresti, F.; Lima, S.; Jim, J. Reproduction and population size of Leptodactylus labyrinthicus (Amphibia, Anura, Leptodactylidae). Russ. J. Herpetol. 2002, 9, 15–20. [Google Scholar]
- Salthe, S.N. Increase in Volume of the Perivitelline Chamber during Development of Rana pipiens Schreber. Physiol. Zoöl. 1965, 38, 80–98. [Google Scholar] [CrossRef]
- Urch, U.A.; Hedrick, J.L. The hatching enzyme from Xenopus laevis: Limited proteolysis of the fertilization envelope. J. Supramol. Struct. Cell. Biochem. 1981, 15, 111–117. [Google Scholar] [CrossRef]
- Yoshizaki, N.; Katagiri, C. Cellular basis for the production and secretion of the hatching enzyme by frog embryos. J. Exp. Zoöl. 1975, 192, 203–212. [Google Scholar] [CrossRef]
- Muñoz, M.J.R.; Martínez, T.A.; Acosta, J.C.; Blanco, G.M. Foam nest construction and first report of agonistic behaviour in Pleurodema tucumanum (Anura: Leptodactylidae). Neotrop. Biol. Conserv. 2019, 14, 117–128. [Google Scholar] [CrossRef]
- Breder, C.M. Amphibians and reptiles of the Rio Chucunaque drainage, Darien, Panama, with notes on their life histories and habits. Bull. Am. Mus. Nat. Hist. 1946, 86, 379–435. [Google Scholar]
- Giaretta, A.A.; Facure, K.G. Terrestrial and communal nesting in Eupemphix nattereri (Anura, Leiuperidae): Interactions with predators and pond structure. J. Nat. Hist. 2006, 40, 2577–2587. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, L.; Li, J.; Li, Y.; Wei, S.; Cai, L.; Wu, H. Transcriptomic analysis of differentially expressed genes in the oviduct of Rhacophorus omeimontis provides insights into foam nest construction. BMC Genom. 2019, 20, 1–14. [Google Scholar] [CrossRef]
- Taylor, P. Observations on the breeding habits of Chiromantis xerampelina. J. Herpetol. Assoc. Afr. 1971, 8, 7–8. [Google Scholar]
- Waldman, B.; Ryan, M.J. Thermal Advantages of Communal Egg Mass Deposition in Wood Frogs (Rana sylvatica). J. Herpetol. 1983, 17, 70–72. [Google Scholar] [CrossRef]
- Villamil, J.; Maneyro, R. Communal foam nest in Physalaemus gracilis (Boulenger 1881) (Anura: Leptodactylidae). Herpetol. Notes 2014, 7, 565–566. [Google Scholar]
- Hamilton, W. Geometry for the selfish herd. J. Theor. Biol. 1971, 31, 295–311. [Google Scholar] [CrossRef]
- Furness, A.I.; Capellini, I. The evolution of parental care diversity in amphibians. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Vertical sections of bubble and foam nests produced by anurans. The left panel shows aquatic nests produced by; (A) Chiasmocleis leucosti—eggs laid within a thin bubble nest, (B) Limnodynastes tasmaniensis—eggs laid below a bubble raft, (C) Limnodynastes peronii—eggs laid throughout a bubble nest, and (D) Engystomops pustulosus—eggs laid within the core of a globular foam nest with an egg-free cortex. The middle panel shows terrestrial nests produced by; (E) Leptodactylus labyrinthicusv—eggs dispersed throughout a globular foam nest except for the top surface, and deposited in an uncapped burrow, and (F) Leptodactylus bufonius—eggs dispersed throughout a globular foam nest except for the top surface, and deposited in a capped burrow. The right panel shows arboreal nests produced by; (G) Chiromantis xerampelina—eggs laid in a globular foam nest with an egg-free cortex, and (H) Rhacophorus malabaricus—eggs dispersed throughout a globular foam nest. Eggs with pigment are shown in black while unpigmented eggs are shown in pale yellow. Colourless circles depict the relative size of air bubbles that form the nests of foam and bubble nesting species.
Figure 1.
Vertical sections of bubble and foam nests produced by anurans. The left panel shows aquatic nests produced by; (A) Chiasmocleis leucosti—eggs laid within a thin bubble nest, (B) Limnodynastes tasmaniensis—eggs laid below a bubble raft, (C) Limnodynastes peronii—eggs laid throughout a bubble nest, and (D) Engystomops pustulosus—eggs laid within the core of a globular foam nest with an egg-free cortex. The middle panel shows terrestrial nests produced by; (E) Leptodactylus labyrinthicusv—eggs dispersed throughout a globular foam nest except for the top surface, and deposited in an uncapped burrow, and (F) Leptodactylus bufonius—eggs dispersed throughout a globular foam nest except for the top surface, and deposited in a capped burrow. The right panel shows arboreal nests produced by; (G) Chiromantis xerampelina—eggs laid in a globular foam nest with an egg-free cortex, and (H) Rhacophorus malabaricus—eggs dispersed throughout a globular foam nest. Eggs with pigment are shown in black while unpigmented eggs are shown in pale yellow. Colourless circles depict the relative size of air bubbles that form the nests of foam and bubble nesting species.
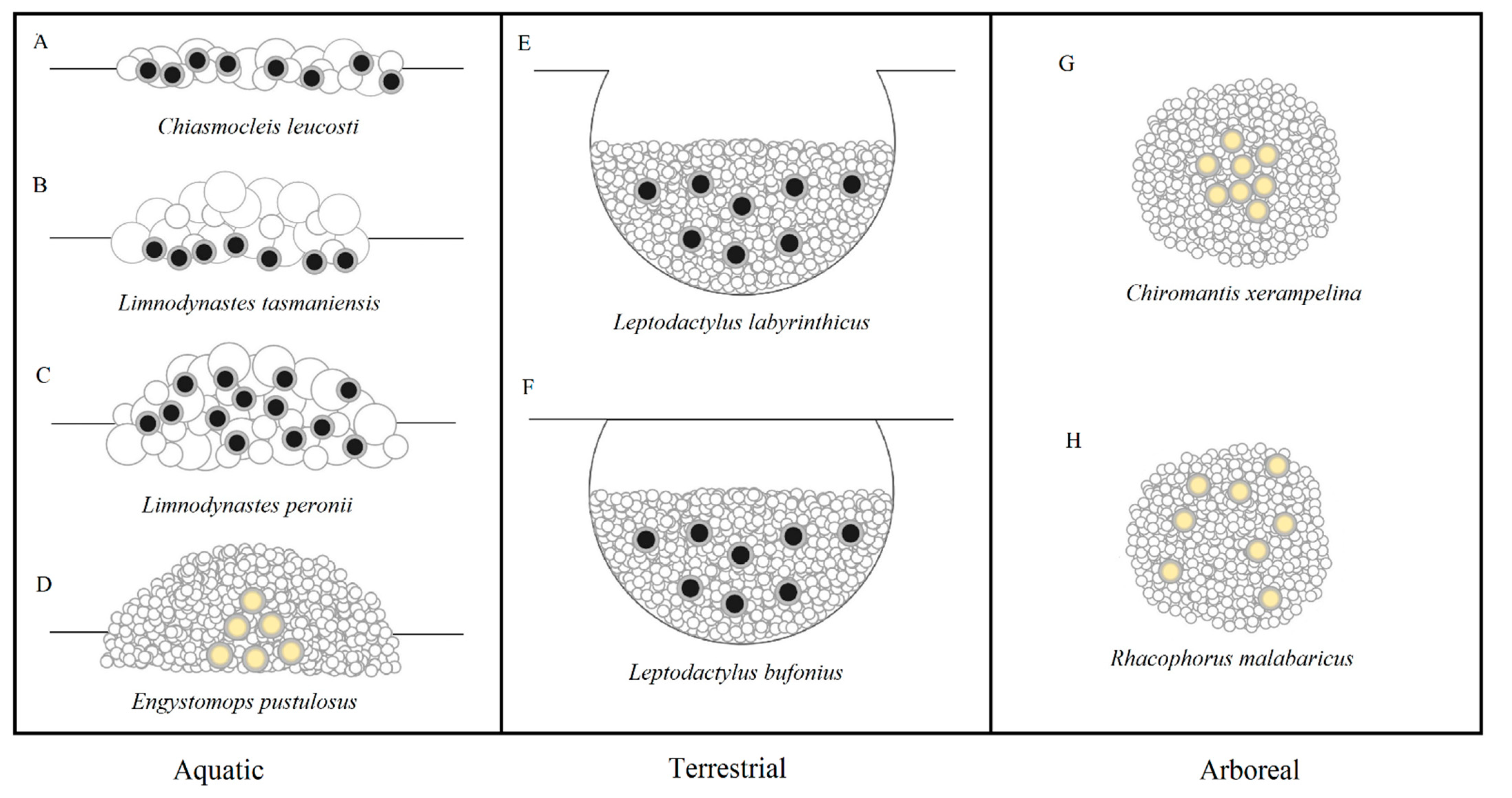
Figure 2.
Freshly oviposited Lechriodus fletcheri froth nest next to the un-frothed egg mass of a sympatric species. Note that the froth nest is floating at the surface of the water, with eggs surrounded by aerated mucus with high albedo, while the un-frothed mass is below the water and surrounding by translucent jelly layers. The nest was observed in the Watagan Mountains, NSW, Australia during the Austral summer of 2018.
Figure 2.
Freshly oviposited Lechriodus fletcheri froth nest next to the un-frothed egg mass of a sympatric species. Note that the froth nest is floating at the surface of the water, with eggs surrounded by aerated mucus with high albedo, while the un-frothed mass is below the water and surrounding by translucent jelly layers. The nest was observed in the Watagan Mountains, NSW, Australia during the Austral summer of 2018.

Figure 3.
Desiccation of the Lechriodus fletcheri froth nest in the absence of freestanding water. Embryos at the top surface have desiccated after several days out of water while those beneath remain alive and developing. Drying of the surface has also preserved the three-dimensional structure of the nest. The nest was obtained from the Watagan Mountains, NSW, Australia during the Austral summer of 2018 and exposed to dry conditions under laboratory conditions.
Figure 3.
Desiccation of the Lechriodus fletcheri froth nest in the absence of freestanding water. Embryos at the top surface have desiccated after several days out of water while those beneath remain alive and developing. Drying of the surface has also preserved the three-dimensional structure of the nest. The nest was obtained from the Watagan Mountains, NSW, Australia during the Austral summer of 2018 and exposed to dry conditions under laboratory conditions.

Figure 4.
Adhesive property of freshly oviposited Lechriodus fletcheri froth nest. The nest was obtained from the Watagan Mountains, NSW, Australia during the Austral summer of 2017 and examined under laboratory conditions.
Figure 4.
Adhesive property of freshly oviposited Lechriodus fletcheri froth nest. The nest was obtained from the Watagan Mountains, NSW, Australia during the Austral summer of 2017 and examined under laboratory conditions.

Figure 5.
Lechriodus fletcheri tadpoles cannibalising eggs around the perimeter of a conspecific froth nest. The nest was obtained from the Watagan Mountains, NSW, Australia during the Austral summer of 2017 and exposed to previously hatched, conspecific tadpoles under laboratory conditions.
Figure 5.
Lechriodus fletcheri tadpoles cannibalising eggs around the perimeter of a conspecific froth nest. The nest was obtained from the Watagan Mountains, NSW, Australia during the Austral summer of 2017 and exposed to previously hatched, conspecific tadpoles under laboratory conditions.

Figure 6.
Potential reptile predator stuck to the top surface of a Lechriodus fletcheri froth nest. The nest was observed in the Watagan Mountains, NSW, Australia during the Austral summer, 2016.
Figure 6.
Potential reptile predator stuck to the top surface of a Lechriodus fletcheri froth nest. The nest was observed in the Watagan Mountains, NSW, Australia during the Austral summer, 2016.

Figure 7.
Lechriodus fletcheri froth nests at various stages of break down. Freshly oviposited nests gradually loose aerated bubbles, spread across the water’s surface and become increasingly translucent prior to embryos hatching. Nests were observed in the Watagan Mountains, NSW, Australia during the Austral summer of 2018.
Figure 7.
Lechriodus fletcheri froth nests at various stages of break down. Freshly oviposited nests gradually loose aerated bubbles, spread across the water’s surface and become increasingly translucent prior to embryos hatching. Nests were observed in the Watagan Mountains, NSW, Australia during the Austral summer of 2018.

Figure 8.
Aquatic communal froth nest masses produced by Lechriodus fletcheri with pigmented eggs distributed throughout each individual bubble nest (A), and arboreal communal mass produced by Chiromantis xerampelina with un-pigmented eggs located in the core of each foam nest (B).
Figure 8.
Aquatic communal froth nest masses produced by Lechriodus fletcheri with pigmented eggs distributed throughout each individual bubble nest (A), and arboreal communal mass produced by Chiromantis xerampelina with un-pigmented eggs located in the core of each foam nest (B).
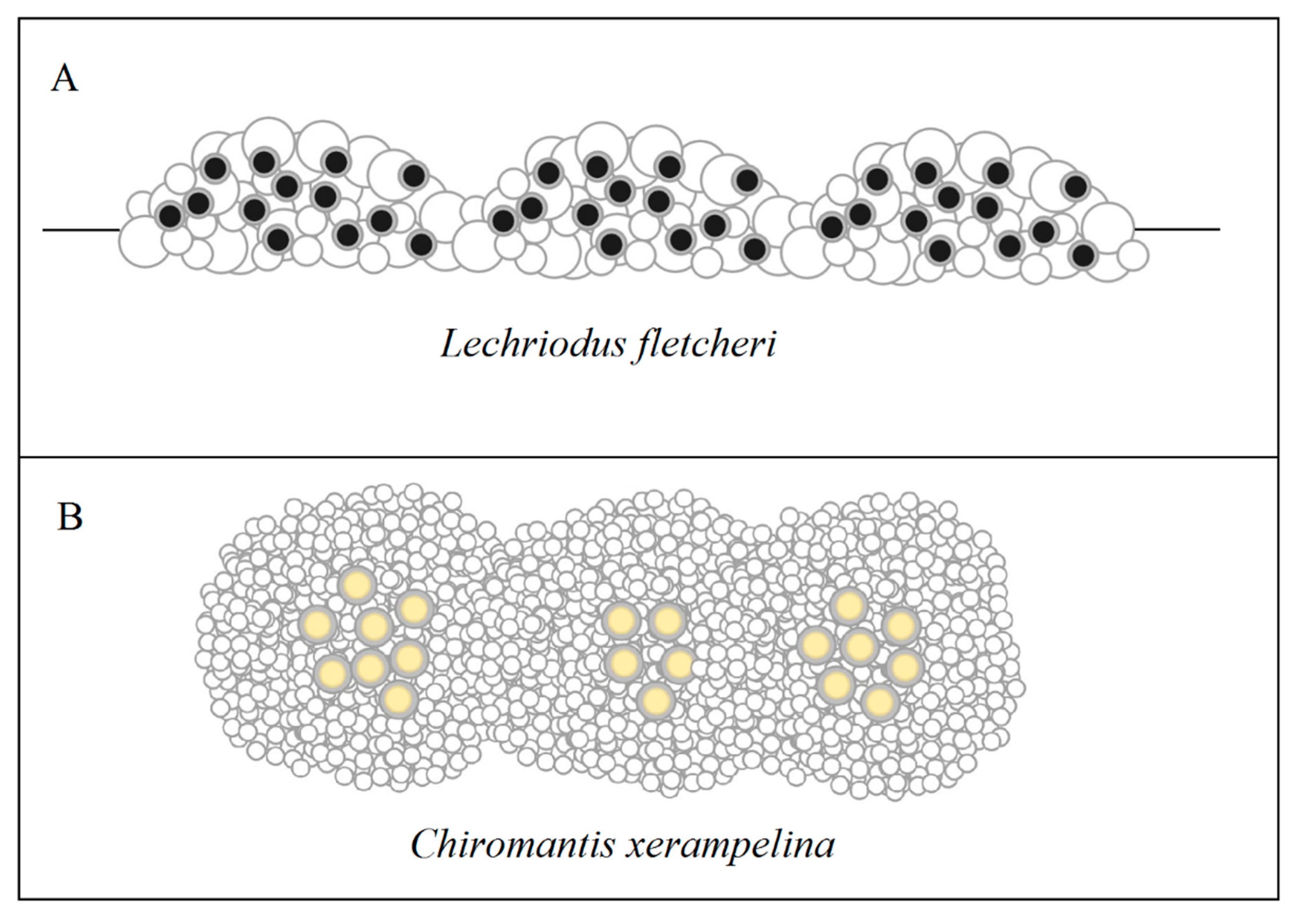
Figure 9.
Communal Lechriodus fletcheri froth nest. At least seven individual froth nests have been laid side-by-side to form this mass. Nests were observed in the Watagan Mountains, NSW, Australia during the Austral summer of 2017.
Figure 9.
Communal Lechriodus fletcheri froth nest. At least seven individual froth nests have been laid side-by-side to form this mass. Nests were observed in the Watagan Mountains, NSW, Australia during the Austral summer of 2017.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gould, J. Safety Bubbles: A Review of the Proposed Functions of Froth Nesting among Anuran Amphibians. Ecologies 2021, 2, 112-137. https://doi.org/10.3390/ecologies2010006
AMA Style
Gould J. Safety Bubbles: A Review of the Proposed Functions of Froth Nesting among Anuran Amphibians. Ecologies. 2021; 2(1):112-137. https://doi.org/10.3390/ecologies2010006
Chicago/Turabian StyleGould, John. 2021. "Safety Bubbles: A Review of the Proposed Functions of Froth Nesting among Anuran Amphibians" Ecologies 2, no. 1: 112-137. https://doi.org/10.3390/ecologies2010006