
Effects of Species of Leaves and Conditioning Time on Vernal Colonization by Temperate Lotic Isopods (Lirceus sp.)

1
Division of Environmental Medicine, New York University Grossman School of Medicine, New York, NY 10010, USA
2
Biology Department, Muhlenberg College, Allentown, PA 18104, USA
3
Delaware Department of Natural Resources and Environmental Control, Division of Watershed Stewardship, Dover, DE 19904, USA
*
Author to whom correspondence should be addressed.
Hydrobiology 2024, 3(2), 63-73; https://doi.org/10.3390/hydrobiology3020005
Submission received: 29 February 2024
/
Revised: 12 April 2024
/
Accepted: 15 April 2024
/
Published: 19 April 2024
(This article belongs to the Special Issue Ecosystem Disturbance in Small Streams)
Abstract
:While some streams have dense populations of aquatic detritivorous isopods, research on the colonization of leaf packs typically focuses on aquatic insects. To determine whether shifts in dominant local forest species might impact isopod populations, we placed leaf packs of red/sugar maple, American beech, and red oak on the substratum of riffles and pools in Cedar Creek (Allentown, PA, USA) in April 2019. We retrieved the packs after one week, re-deployed them, and re-collected them after two weeks of submersion, enumerating the number of isopods (Lirceus sp.) upon each retrieval. Surprisingly, neither the species of leaf nor the stream microhabitat significantly affected the number of isopods. However, the duration of leaf conditioning was important; significantly more isopods inhabited leaves after two weeks of submersion than after only one week. Maple and oak leaves displayed significantly more skeletonization after two weeks than the beech leaves, which remained intact. However, the similar numbers of isopods across leaf species suggest either the presence of acceptable, consumable microbial communities on all three species of leaves or that a tradeoff exists between the value of food and the importance of refuge provided by intact leaves.
1. Introduction
Within first- and second-order streams, floating senescent leaves often snag and accumulate in certain locations, creating a biomass of habitat—a leaf pack—that subsequently provides both food and shelter for benthic invertebrates [1,2]. These allochthonous inputs frequently dominate the carbon resources within streams [3,4]. Detritivores often use various leaves differently [5,6,7,8], disproportionately shredding leaves with weaker structural integrity or varied nutrient content and secondary compounds [9,10,11,12,13]. The use of detritus is also heavily influenced over time by leaching and the colonizing species of microbes, especially fungi [10,14,15,16,17,18,19,20,21,22,23,24], and the leaf species can impact the microbial community e.g., [4,25,26]. It is not yet well understood how these traits collectively affect the use of various leaves by macroinvertebrates, as past studies have yielded mixed results [4,27]. In addition to direct nutritional benefits, animals sheltered within leaf packs may gain refuge from predators and drag forces [7,8,12,28]. Therefore, the consumption of leaf material that results in “skeletonization” can cause a conflict between the values of food and shelter.
While numerous past studies have examined the colonization of leaf packs by larval aquatic insects, detritivorous aquatic isopods are rarely mentioned in these studies (although see [29,30,31,32,33]). However, the rate of nutrient cycling, local levels of primary productivity, and fish populations can depend on the local biomass of isopods [16,34,35,36,37], which can act as a keystone species [38]. Some marine isopods prioritize shelter over food resources [39]. Stream isopods may avoid habitats with predators [40] or retreat to refuges when detecting fish chemical cues [32]. While some stream isopods may be abundant in riffles but virtually absent from pools due to the higher risk of predation, incidental observations in our experimental stream suggested high isopod densities in bankside pools. If predation pressures vary across microhabitats, the relative priority of shelter versus nutrition may also change for detritivores.
Our current study examined colonization by the lotic isopod Lirceus sp. Rafinesque, 1820 within leaf packs in a temperate urban stream over time to determine (1) whether isopods disproportionately colonized leaves of different common deciduous broad-leaved species, and whether that preference was impacted by either (2) microhabitat or (3) leaf-conditioning time. We utilized red oak (Quercus rubra L.), maple (Acer saccharum Marsh. and Acer rubrum L.), and American beech (Fagus grandifolia Ehrh.) leaves, which differ in nutrient concentrations [27,41]. Maple leaves have lower levels of lignin and tannins [10,42] and degrade more rapidly than beech or oak [2,10,14,43], Iyengar, pers. obs. Laboratory studies suggested that Lirceus sp. may prefer to consume red maple leaves more than other species [44]. Therefore, we predicted that after one week of submersion, leaf packs containing red maple would have the highest numbers of isopods. In contrast, red oak leaves, due to their higher toughness and lower levels of phosphorus and nitrogen [27], would likely contain the lowest numbers of isopods. However, after two weeks of submersion, high degrees of skeletonization in the most palatable leaves due to their consumption would reduce the attraction of maple leaves. Therefore, we predicted that more isopods would reside in red oak leaves after two weeks of submersion. Lastly, we predicted that more isopods would utilize leaf packs in the bankside pools compared with the riffles, due to less turbulence.
2. Materials and Methods
2.1. Experimental Design
To investigate the relative importance of different species of leaves to stream isopods, we constructed and deployed leaf packs from the first to third week in April 2019 on the benthos of Cedar Creek (Cedar Beach Parkway, Allentown, PA, USA; 40°35′29.03″ N, 75°30′57.37″ W). The experimental animals were members of Lirceus sp. [45,46,47], but conclusive species-level identification was not possible due to taxonomic uncertainty within the genus ([48] and personal communication with Frank Kuserk; Moravian College and Mike Broomall; Stroud Water Resource Center). The lack of a mandibular palp excluded L. fontinalis as a possibility. The presence of a carina at the base of the antennules and the lack of median and distal processes on the gnathopod suggested that the animals under consideration may be Lirceus brachyurus (Harger, 1876).
Cedar Creek is a low-order, low-gradient permanent stream that meanders through a well-groomed public park on the edge of the third largest city (by population) in Pennsylvania. Despite multiple direct and indirect impacts by human activities, the waters maintain a large population of mayflies and caddisflies and are stocked with trout a short distance downstream. Within this stream reach, the density of individuals of Lirceus sp. can be extremely high, with a 10 min sampling effort yielding more than 500 isopods of various sizes (pers. observ.) at any time of the year. Incidental observations suggest that, while certain locations in the stream may have higher densities of isopods, there is an appreciable number of isopods anywhere on the surface of the benthos. Therefore, even a few days is likely sufficient for a large number of isopods to have encountered deployed leaf packs. This study was conducted in springtime, when this site contains little submerged leaf litter, to prevent floating leaves from contaminating the mono-species leaf packs that we provided. Typically, there is little autochthonous material in this area of the stream in April, other than a covering of filamentous algae on the submerged metal mesh that is used to stabilize the bankside.
We used the leaves of four deciduous tree species in three treatments: American beech (Fagus grandifolia), American red oak (Quercus rubra), and maple (a combination of Acer saccharum and Acer rubrum). All are native species common in the state of Pennsylvania and occur near this stream, though not always streamside. Leaves were collected from the floors of nearby forests in March 2019, dried at room temperature (20 °C), and then stored loosely inside black plastic bags at room temperature until use. Only intact, minimally worn leaves were used.
Each leaf pack (the unit of experimental replication; 15.24 cm × 15.24 cm) was composed of leaves contained within a plastic mesh pouch (Vexar hardware netting; pore size 1.27 cm × 1.27 cm). The mesh was large enough to enable access and egress for all local invertebrates except crayfish, while simultaneously reducing leaf attrition through factors other than consumption. Each leaf pack contained three handfuls of dried leaves to mirror the thickness of local natural leaf packs. We focused on ensuring that the entire surface area of the leaf pack was covered and that the numbers of leaf layers within the packs were consistent, even across species, rather than focusing on the dry weight, especially as stems contribute disproportionately to weight. However, the later weighing of comparable amounts of dried leaves indicated that the weights of the packs were likely around 0.17 g for the red maple, 0.16 g for the beech, and 0.24 g for the red oak leaves.
Nine leaf packs were attached in a Latin Square design arrangement, with each pack connected at the corners using plastic cable ties (this unit is hereafter referred to as a “mat;” Figure 1). Thus, each mat contained three replicates of each of the three treatments, and the various leaf species treatments were interspersed, contiguous with each other, yet separate, to allow roaming isopods to likely encounter multiple treatments. Each mat was secured flush to the benthos using a plastic tent stake at each of the four corners. Adjacent mats were at least one meter apart.
Leaf packs were deployed in two microhabitats: on a submerged gravel bed in the center of the stream and in a shallow pool/run adjacent to the bank (N = 9 per each leaf species × microhabitat treatment combination; for a total of 54 leaf packs initially deployed). The total stream width in the stream reach used for our experiment was approximately 4.75 m, with the submerged gravel area 3.5 m of that width. The depth of the gravel bed was 0.25 m, while the depth bankside was 0.45 m. The submerged gravel bed experienced higher stream velocities and therefore was composed of larger benthic rocks with little mud, while the bankside location corresponded with slower water velocities and a thin layer of muddy sediment overlying the gravel benthos. Widely spaced (10 m to 15 m apart) single individuals of various deciduous tree species, including red maple, river birch (Betula nigra), and weeping willow (Salix babylonica, which is not native to this area), were present along either side of the stream but did not cast much shade over either microhabitat, especially as they were just beginning to leaf out. Both microhabitats were almost fully exposed to the sun. Unfortunately, we did not measure water temperature during the experiment, but water temperatures of 12 °C to 14 °C have been recorded in this stream reach in April; April 2019 likely displayed water temperatures close to that range.
A week post-deployment, mats were retrieved and deconstructed streamside, with each pack immediately being placed individually into a bag of stream water to minimize the loss of isopods. In the laboratory, leaves were gently removed from the pack and isopods were collected, enumerated, and preserved in 70% ethyl alcohol. Other aufwuchs and sediment were not intentionally removed to minimize our impact on the natural leaf conditioning process.
During transport (less than 30 min) and the two days of processing, the leaves remained in stream water to promote the continuity of the established microbial and invertebrate community. After the removal of the isopods, leaves from the bankside location were replaced within their original packs and then re-deployed at the same spot as before. Mats from the gravel bed were not re-deployed due to time constraints. Seven days after re-deployment (16 days since initial in-stream placement), the bankside mats were retrieved and processed as before, with the additional step that the leaves of each pack received a single qualitative aggregate score according to their degree of skeletonization: minor, moderate, or extreme.
2.2. Data Analysis
We calculated a three-way ANOVA to examine the effects of the independent variables of leaf species, stream microhabitat, and time of submersion on the number of isopods within the leaf pack and included the leaf mat (comprised of nine leaf packs) as a random blocking variable. We used the computer program Data Desk (version 6.0; Data Description, Ithaca, NY, USA) for ANOVA calculations. We assessed compliance to the assumptions inherent for an ANOVA by calculating Kolmogorov tests for normality and Levene’s tests for homogeneity of variance and visually inspected the plots of residuals. To remove violations of equal variance in the three-way ANOVA, the dependent variable was transformed before calculations using ln(y + 1). After the determination of the significance of any factor, we utilized Bonferroni-adjusted pairwise post hoc comparisons.
Differences among the qualitative assessments of skeletonization upon retrieval after two weeks of submersion were analyzed using a Fisher Exact Test for Count Data [49]. This nonparametric analysis was calculated using RStudio Software (R-4.0.2; [50]). We utilized p < 0.05 as the threshold for significance in all analyses.
3. Results
Within the three-way ANOVA, there was no significant effect of leaf species or microhabitat (bankside versus gravel bed) on the number of isopods (Figure 2; Table 1; F2,68 = 0.342, p = 0.712 and F1,68 = 2.44, p = 0.123, respectively) and neither interaction term was significant (leaf species × microhabitat: F2,68 = 0.455, p = 0.637; leaf species × week: F2,68 = 0.030, p = 0.971). There were significantly more isopods within the leaf packs after two weeks of submersion than after only one week of submersion (Figure 3, Table 1; F1,68 = 66.786, p < 0.0001), and the mat, a random factor, had a significant effect on the number of isopods therein (F4,68 = 3.137, p = 0.020). There was an outlier value within the maple leaf bankside pack replicates that contained an unusually large number of isopods, perhaps due to projecting branches from a riparian bush that retained additional allochthonous material near that leaf pack. However, excluding this replicate did not alter statistical conclusions.
Skeletonization was minimal, or non-existent, for all leaves after one week of submersion in the stream, so no statistics were calculated. However, after two weeks of submersion, the results of the Fisher Exact Test indicated there was a significant difference in the relative degree of skeletonization across the various types of leaves (Table 2; p = 0.009). Beech leaves displayed a significantly lower degree of skeletonization than either maple leaves (adjusted post hoc p = 0.0068) or red oak leaves (adjusted post hoc p = 0.044), while maple and oak were not significantly different from each other (adjusted post hoc p = 0.835). Additionally, a qualitative fingertip assessment (conducted by rolling intact sections of the leaves between the thumb and forefinger) suggested that the individual maple leaves had thinned the most compared to their original thickness before initial deployment, while the oak leaves had thinned the least.
4. Discussion
4.1. Leaves as Nutritional Resources versus Shelters
We predicted that the fewest isopods would colonize packs of red oak leaves and that the largest numbers of isopods would be found within maple leaves one-week post-deployment. However, the isopods did not discriminate among the three taxa of locally common deciduous leaves, inhabiting the leaf packs in surprisingly similar numbers. Similarly, Fuell et al. [51] noted that the biomass and relative abundance of Lirceus isopods did not differ by leaf species when examining abscised willow oak and green ash leaves.
Our findings of equal colonization may in part be explained by the trade-off between leaf toughness and carbon transfer. Recent studies have changed the longstanding view that slowly decomposing leaves are of “poor quality”. Instead, slower leaf decomposition can elevate carbon transfer to consumers, while rapidly decomposing leaf litter (previously deemed “high-quality”) supports microbial productivity [4,52]. In fact, some studies have found that long-lived shredders grow more rapidly by feeding on oak leaves rather than maple leaves [53,54]. Similar to our current results, Friberg and Jacobsen [55] found that the feeding preference of amphipod and trichopteran shredders was unrelated to the toughness, fiber content, total phosphorous, carbon to nitrogen ratio, or total nitrogen of the food. While previous laboratory observations [2,14,44] suggested that isopods preferred red maple leaves, those studies assessed feeding preference, not colonization, as in our current study. The presence of lotic isopods within leaf packs is the result of the leaf’s value as both a food resource and protective shelter, so it is important to consider whether these two criteria are congruent across leaf species.
The presence of isopods across all leaf packs during our first collection, despite almost no evidence of herbivory, suggests that isopods either purposefully or incidentally used leaf packs as refugia. While previous researchers posited that nourishment was the key role of leaves [28,56], the presence of invertebrates within artificial leaves in a previous study [28] suggests an additional function of shelter and protection from visually oriented predators. Hiding within a pack of non-conditioned leaves during the day may provide comparatively similar protection across species of leaves. However, as leaves condition over time and various species either degrade or become palatable at different rates—as evidenced by the elevated degree of skeletonization of the maple leaves in our study after two weeks—the trade-off between acceptable food and acceptable cover from predators may continue to promote equal numbers of isopods across species of leaves.
As climate change, anthropogenic impacts, and other factors alter the composition of local forests, the proportional representation of the species of leaves entering streams is also changing. Red maple and red oak have increased in prevalence within forests in the northeastern United States from their pre-European settlement distribution [57,58]. Our results suggest that plantings of any of these three tree types (maple, red oak, and American beech) in near-stream environments will equally promote isopod colonization, but slowly decaying leaves (such as beech) may be important in providing longer-lasting protective shelters.
4.2. Leaf Packs in Different Microhabitats
We were surprised to find no significant difference in the density of isopods between microhabitats, as our own previous autumn field collections seemed to yield more isopods in the stream-side shallows than on the gravel beds. However, while dense accumulations of leaves typically existed in the bankside pools in autumn collections, the gravel beds contained only patchily distributed leaves. Therefore, the leaf packs may have introduced a three-dimensional environment with boundary layers therein [59] that provided reduced-flow refugia [28] and locally elevated isopod densities in areas typically lacking amalgamations of leaf litter, especially in springtime (as also proposed in [28]). If so, the low-technology and relatively low-effort step of anthropogenic leaf pack augmentation in the springtime may help isopods and other stream invertebrates that struggle with flooding, which would be especially important in areas of increasing extreme weather events driven by climate change.
4.3. Colonization of Leaf Packs under Different Durations of Conditioning
The majority of water-soluble compounds are typically leached from leaves in streams within 24 h of submersion [15], while microbial and fungal colonization takes one or two weeks [3,4,16], but whether the microbial community would have stabilized in terms of its relative attraction to detritivorous isopods after only one week is unknown. Therefore, we examined the isopod colonization of the same leaves over a two-week period. Most studies of leaf pack colonization according to conditioning deploy replicate packs that are retrieved after varying lengths of time, but re-deploying the same leaves after rapid processing in the laboratory, as we did in our study, better controls for the impacts of variability among leaves of the same species. By removing the isopods before re-deployment, both collections examined isopods that found and entered the packs within one week. Thus, our methodology specifically measures the number of isopods that colonized the packs according to leaf conditioning time, not a gradual build-up over two weeks of a population of colonists.
While the isopods did not discriminate among leaf species, the significant increase in isopods over time across all leaf species suggests that microbial growth attracts isopods. Previous research has shown that detritivores prefer rapidly conditioned leaves over freshly fallen or sterile leaves [14,60], because they feed on colonizing fungi [57,58,61,62]. Leaf conditioning may also ameliorate the aquatic environment inside a leaf pack (for instance, buffering leachate or tannins of oaks). Similarly, Ligeiro et al. [63] found that exposure time, rather than leaf-detritus species, impacted colonization by aquatic invertebrates, while Graça et al. [24] noted that fungal biomass, rather than durability or nitrogen content, determined preference in other freshwater isopod species.
Fungi may even represent a complete food source rather than merely improving the food quality [64,65,66]. Differences in the suites of fungal species hosted by various leaves may drive the selectivity of detritivores more than parameters specific to the leaves themselves [38,67,68]. Therefore, the similar numbers of isopods suggest comparable amounts of isopod-acceptable fungi across all species of leaves, which we did not expect. Exploiting multiple leaf species, each supporting various acceptable fungal communities, could have provided a more complete diet to an individual isopod. Or the conditions in early spring may have restricted growth to a small subset of fungal species which could colonize all three species of leaves used in our experiments. In fact, the isopods may not have targeted the leaf material itself, as another species of isopods scrape fungal hyphae and only incidentally ingest leaf material [24]. The intact nature of the experimental beech leaves initially suggested that isopods did not use these leaves for nourishment, but grazing the fungi may have left no trace on these tough leaves. It would be of great interest to examine the specific fungal communities encrusting on leaves and whether warmer weather would promote a more diverse fungal community across various leaf species. However, while there is controversy in the literature as to whether macroinvertebrate shredders target the leaves themselves or the biofilm community that forms on the leaves e.g., [69,70], this differentiation is largely irrelevant to our current questions of interest. We investigated the overall attraction of various species of leaves to colonization by isopods during the process of leaf conditioning, irrespective of whether the animals were using the packs for food (leaf material or biofilm), shelter, or other purposes.
4.4. The Possible Impact of Experimental Season
Previous leaf pack studies have predominantly examined stream detritus processing in autumn e.g., [21]. However, while many local deciduous trees release the bulk of their leaves in autumn, some trees (e.g., American beech) retain senescent leaves, losing them incrementally. Additionally, thick layers of detritus on the forest floor can preserve leaves in largely pristine condition through much of the winter, with snowmelt or storms bringing springtime pulses of leaf litter into the streams of northern deciduous forests [71]. Consequently, while peaking in late autumn, there is a relatively continuous input of allochthonous leaves to many streams in the northeastern United States throughout the autumn, winter, and spring [72,73]. For animals such as isopods, whose populations persist year-round, nutrition and dietary preferences may shift seasonally with resource availability or temperatures [74]. Thus, examining leaf preference throughout the year is imperative for the complete understanding of isopod colonization.
However, isopods may not recognize preferred leaf species that were seasonally unusual. By the late winter/early spring in our current study area, local streams typically lack rapidly decomposing maple leaves. Consumers often experience a time-lag until they recognize the acceptability of previously rare resources [75]. Most new allochthonous material entering the streams at this time are pollen, flower parts, and insect frass, rather than leaf litter [21]. Slowly decaying leaves may be the only litter resources remaining this late in the season [2,15,19]. Isopods are present year-round and therefore likely have adaptations to utilize the remaining late-season tougher leaves. Thus, seasonally influenced frequency-dependent search images [76,77,78] may prime isopods to only recognize and colonize the most-abundant leaves in springtime: slower-decomposing, less-palatable species. The two-week leaf pack deployment may not have been enough time to allow for the recognition of an atypically timed pulse of a higher-value food. Whether the presence of maple leaf packs over a longer period of time would produce a shift in gustatory search images is of interest for future studies.
5. Conclusions
While colonization by isopods within leaf packs did greatly increase over time (likely concurrent with fungal colonization), the type of leaves within the pack did not significantly predict the number of isopods within. Additionally, there was no evidence that isopods preferentially colonized a particular microhabitat in this stream (as submerged gravel beds and bankside pools contained similar densities). If the function of leaves includes both food and shelter, our results suggest that near-stream plantings of any of these three tree types (maple, red oak, and American beech) will equally promote isopod colonization. However, durable, slowly decaying leaves (such as beech) may be important in providing long-term protective shelters for stream isopods, especially in the springtime when there are few alternative shelters because other leaves have disappeared and autochthonous primary productivity is nascent.
Author Contributions
Conceptualization, methodology, formal analysis, and writing—review and editing R.E.H., A.N.S. and E.V.I.; investigation and writing—original draft preparation, R.E.H. and A.N.S.; resources, E.V.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were not required for this study as it involved only invertebrate animals. Appropriate educator and fishing licenses were obtained by EVI from the Pennsylvania Fish and Boat Commission.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting the reported results will be provided upon reasonable request to the corresponding author.
Acknowledgments
We would like to thank Allentown Parks and Recreation for site access, Peter Saenger and Thomas Shotzbarger for assistance with supplies, and Zachary Delecce for assistance with processing samples.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Minshall, G.W. Role of allochthonous detritus in the trophic structure of a woodland springbrook community. Ecology 1967, 48, 139–149. [Google Scholar] [CrossRef]
- Kaushik, N.K.; Hynes, H.B.N. The fate of dead leaves that fall into streams. Hydrobiologia 1971, 68, 465–516. [Google Scholar]
- Cross, W.F.; Benstead, J.P.; Frost, P.C.; Thomas, S.A. Ecological stoichiometry in freshwater benthic systems: Recent progress and perspectives. Freshw. Biol. 2005, 50, 1895–1912. [Google Scholar] [CrossRef]
- Marks, J.C. Revisiting the fates of dead leaves that fall into streams. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 547–568. [Google Scholar] [CrossRef]
- Smock, L.A.; Metzler, G.M.; Gladden, J.E. Role of debris dams in the structure and functioning of low-gradient headwater streams. Ecology 1989, 70, 764–775. [Google Scholar] [CrossRef]
- Richardson, J.S. Seasonal food limitation of detritivores in a montane stream: An experimental test. Ecology 1991, 72, 873–887. [Google Scholar] [CrossRef]
- Hildrew, A.G.; Dobson, M.K.; Groom, A.; Ibbotson, A.; Lancaster, J.; Rundle, S.D. Flow and retention in the ecology of stream invertebrates. Int. Ver. Theor. Angew. Limnol. Verhandlungen 1991, 24, 1742–1747. [Google Scholar] [CrossRef]
- Dobson, M.; Hildrew, A.G. A test of resource limitation among shredding detritivores in low order streams in southern England. J. Anim. Ecol. 1992, 61, 69–78. [Google Scholar] [CrossRef]
- Rosenthal, G.A.; Janzen, D. (Eds.) Herbivores: Their Interaction with Secondary Plant Metabolites; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Gessner, M.O.; Chauvet, E. Importance of stream microfungi in controlling breakdown rates of leaf litter. Ecology 1994, 75, 1807–1817. [Google Scholar] [CrossRef]
- Canhoto, C.; Graça, M.A.S. Food value of introduced eucalypt leaves for a Mediterranean stream detritivore: Tipula lateralis. Freshw. Biol. 1995, 34, 209–214. [Google Scholar] [CrossRef]
- Graça, M.A.S. The role of invertebrates on leaf litter decomposition in streams—A review. Int. Rev. Hydrobiol. 2001, 86, 383–393. [Google Scholar] [CrossRef]
- Stoler, A.B.; Relyea, R.A. Leaf litter quality induces morphological and developmental changes in larval amphibians. Ecology 2013, 94, 1594–1603. [Google Scholar] [CrossRef]
- Kaushik, N.K.; Hynes, H.B.N. Experimental study on the role of autumn-shed leaves in aquatic environments. J. Ecol. 1968, 56, 229–243. [Google Scholar] [CrossRef]
- Petersen, R.C.; Cummins, K.W. Leaf processing in a woodland stream. Freshw. Biol. 1974, 4, 343–368. [Google Scholar] [CrossRef]
- Cummins, K.W. Structure and function of stream ecosystems. BioScience 1974, 24, 631–641. [Google Scholar] [CrossRef]
- Bärlocher, F.; Kendrick, B. Assimilation efficiency of Gammarus pseudolimnaeus (Amphipoda) feeding on fungal mycelium or autumn-shed leaves. Oikos 1975, 26, 55–59. [Google Scholar] [CrossRef]
- Bärlocher, F.; Kendrick, B. Leaf-conditioning by microorganisms. Oecologia 1975, 20, 359–362. [Google Scholar] [CrossRef]
- Boling, R.H.; Goodman, E.D.; Van Sickle, J.A.; Zimmer, J.O.; Cummings, K.W.; Petersen, R.C.; Reice, S.R. Toward a model of detritus processing in a woodland stream. Ecology 1975, 56, 141–151. [Google Scholar] [CrossRef]
- Suberkropp, K.; Godshalk, G.L.; Klug, M.J. Changes in the chemical composition of leaves during processing in a woodland Stream. Ecology 1976, 57, 720–727. [Google Scholar] [CrossRef]
- Anderson, N.H.; Sedell, J.R. Detritus processing by macroinvertebrates in stream ecosystems. Annu. Rev. Entomol. 1979, 24, 351–377. [Google Scholar] [CrossRef]
- Golladay, S.W.; Webster, J.R.; Benfield, E.F. Factors affecting food utilization by a leaf shredding aquatic insect: Leaf species and conditioning time. Ecography 1983, 6, 157–162. [Google Scholar] [CrossRef]
- Chamier, A.-C. Cellulose digestion and metabolism in the freshwater amphipod Gammarus pseudolimnaeus Bousfield. Freshw. Biol. 1991, 25, 33–40. [Google Scholar] [CrossRef]
- Graça, M.A.S.; Maltby, L.; Calow, P. Importance of fungi in the diet of Gammarus pulex and Asellus aquaticus I: Feeding strategies. Oecologia 1993, 93, 139–144. [Google Scholar] [CrossRef]
- Gulis, V.; Suberkropp, K. Leaf litter decomposition and microbial activity in nutrient-enriched and unaltered reaches of a headwater stream. Freshw. Biol. 2003, 48, 123–134. [Google Scholar] [CrossRef]
- Besemer, K. Biodiversity, community structure and function of biofilms in stream ecosystems. Res. Microbiol. 2015, 166, 774–781. [Google Scholar] [CrossRef]
- Ostrofsky, M.L. Relationship between chemical characteristics of autumn-shed leaves and aquatic processing rates. J. N. Am. Benthol. Soc. 1997, 16, 750–759. [Google Scholar] [CrossRef]
- Dobson, M.; Hildrew, A.G.; Ibbotson, A.; Garthwaite, J. Enhancing litter retention in streams. Freshw. Biol. 1992, 28, 71–79. [Google Scholar] [CrossRef]
- Styron, C.E.; Burbanck, W.D. Ecology of an aquatic isopod, Lirceus fontinalis Raf., emphasizing radiation effects. Am. Midl. Nat. 1967, 78, 389–415. [Google Scholar] [CrossRef]
- Styron, C.E. Ecology of two populations of an aquatic isopod, Lirceus fontinalis Raf. Ecology 1968, 49, 629–636. [Google Scholar] [CrossRef]
- Murphy, P.M.; Learner, M.A. The life history and production of Asellus aquaticus (Crustacea: Isopoda) in the River Ely, South Wales. Freshw. Biol. 1982, 12, 435–444. [Google Scholar] [CrossRef]
- Holomuzki, J.R.; Short, T.M. Habitat use and fish avoidance behaviors by the stream-dwelling isopod Lirceus fontinalis. Oikos 1988, 52, 79–86. [Google Scholar] [CrossRef]
- Sparkes, T.C. The effects of size-dependent predation risk on the interaction between behavioral and life history traits in a stream-dwelling isopod. Behav. Ecol. Sociobiol. 1996, 39, 411–417. [Google Scholar] [CrossRef]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef] [PubMed]
- Covich, A.P.; Palmer, M.A.; Crowl, T.A. The role of benthic invertebrate species in freshwater ecosystems: Zoobenthic species influence energy flows and nutrient cycling. BioScience 1999, 49, 119–127. [Google Scholar] [CrossRef]
- Sparkes, T.C.; Mills, C.M.; Volesky, L.A.; Talkington, J.A. Leaf degradation, macroinvertebrate shredders and energy flow in streams: A laboratory-based exercise examining ecosystem processes. Am. Biol. Teach. 2008, 70, 90–94. [Google Scholar] [CrossRef]
- Griffiths, N.A.; Tank, J.L.; Roley, S.S.; Stephen, M.L. Decomposition of maize leaves and grasses in restored agricultural streams. Freshw. Sci. 2012, 31, 848–864. [Google Scholar] [CrossRef]
- Lafuente, E.; Lürig, M.D.; Rövekamp, M.; Matthews, B.; Buser, C.; Vorburger, C.; Räsänen, K. Building on 150 years of knowledge: The freshwater isopod Asellus aquaticus as an integrative eco-evolutionary model system. Front. Ecol. Evol. 2021, 9, 748212. [Google Scholar] [CrossRef]
- Vesakoski, O.; Merilaita, S.; Jormalainen, V. Reckless males, rational females: Dynamic trade-off between food and shelter in the marine isopod Idotea balthica. Behav. Process. 2008, 79, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, K.L.; Gall, B.G. Complex predator–prey interactions between the rusty crayfish (Orconectes rusticus) and invertebrate and vertebrate prey within their native range. J. Freshw. Ecol. 2014, 29, 267–277. [Google Scholar] [CrossRef]
- Frost, P.C.; Elser, J.J. Growth responses of littoral mayflies to the phosphorus content of their food. Ecol. Lett. 2002, 5, 232–240. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Linkins, A.E. Factors controlling mass loss and nitrogen dynamics of plant litter decaying in northern streams. Bull. Mar. Sci. 1984, 35, 341–356. [Google Scholar]
- Webster, J.R.; Benfield, E.F. Vascular plant breakdown in freshwater ecosystems. Annu. Rev. Ecol. Syst. 1986, 17, 567–594. [Google Scholar] [CrossRef]
- Iyengar, E.V.; Schwartz, C.I.; Davidson, A.T. Long-term maintenance requirements of the riparian isopod, Lirceus sp. Hydrobiologia 2017, 802, 53–69. [Google Scholar] [CrossRef]
- Hubricht, L.; Mackin, J.G. The freshwater isopods of the genus Lirceus (Asellota, Asellidae). Am. Midl. Nat. 1949, 42, 334–349. [Google Scholar] [CrossRef]
- Williams, W.D. Freshwater Isopods (Asellidae) of North America; U.S Environmental Protection Agency: Cincinnati, OH, USA, 1976.
- Pennak, R.W. Freshwater Invertebrates of the United States, 2nd ed.; Wiley: New York, NY, USA, 1978. [Google Scholar]
- Styron, C.E. Taxonomy of two populations of an aquatic isopod, Lirceus fontinalis Raf. Am. Midl. Nat. 1969, 82, 402–416. [Google Scholar] [CrossRef]
- McDonald, J.H. Handbook of Biological Statistics, 3rd ed.; Sparky House Publishing: Baltimore, MA, USA, 2014; pp. 75–85. [Google Scholar]
- R Studio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 16 June 2020).
- Fuell, A.K.; Entrekin, S.A.; Owen, G.S.; Owen, S.K. Drivers of leaf decomposition in two wetland types in the Arkansas River Valley, U.S.A. Wetlands 2013, 33, 1127–1137. [Google Scholar] [CrossRef]
- Siders, A.C.; Compson, Z.G.; Hungate, B.A.; Dijkstra, P.; Koch, G.W.; Wymore, A.S.; Grandy, A.S.; Marks, J.C. Litter identity affects assimilation of carbon and nitrogen by a shredding caddisfly. Ecosphere 2018, 9, e02340. [Google Scholar] [CrossRef]
- Hutchens, J.J.; Benfield, E.F.; Webster, J.R. Diet and growth of a leaf-shredding caddisfly in southern Appalachian streams of contrasting disturbance history. Hydrobiologia 1997, 396, 193–201. [Google Scholar] [CrossRef]
- Fuller, C.L.; Evans-White, M.A.; Entrekin, S.A. Growth and stoichiometry of a common aquatic detritivore respond to changes in resource stoichiometry. Oecologia 2015, 177, 837–848. [Google Scholar] [CrossRef]
- Friberg, N.; Jacobsen, D. Feeding plasticity of two detritivore-shredders. Freshw. Biol. 1994, 32, 133–142. [Google Scholar] [CrossRef]
- Dudgeon, D.; Wu, K.K.Y. Leaf litter in a tropical stream: Food or substrate for macroinvertebrates? Arch. Hydrobiol. 1999, 146, 65–82. [Google Scholar] [CrossRef]
- Abrams, M.D. The red maple paradox. BioScience 1998, 48, 355–364. [Google Scholar] [CrossRef]
- Abrams, M.D. Where has all the white oak gone? BioScience 2003, 53, 927–939. [Google Scholar] [CrossRef]
- Statzner, B.; Gore, J.A.; Resh, V.H. Hydraulic stream ecology: Observed patterns and potential applications. J. N. Am. Benthol. Soc. 1988, 7, 307–360. [Google Scholar] [CrossRef]
- Bärlocher, F.; Kendrick, B. Fungi in the diet of Gammarus pseudolimnaeus (Amphipoda). Oikos 1973, 24, 295–300. [Google Scholar] [CrossRef]
- Bärlocher, F.; Kendrick, B. Dynamics of fungal populations on leaves in a stream. J. Ecol. 1974, 62, 761–791. [Google Scholar] [CrossRef]
- Suberkropp, K.; Arsuffi, T.L.; Anderson, J.P. Comparison of degradative ability, enzymatic activity, and palatability of aquatic hyphomycetes grown on leaf litter. Appl. Environ. Microbiol. 1983, 46, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Ligeiro, R.; Moretti, M.S.; Gonçalves, J.F.; Callisto, M. What is more important for invertebrate colonization in a stream with low-quality litter inputs: Exposure time or leaf species? Hydrobiologia 2010, 654, 125–136. [Google Scholar] [CrossRef]
- Cummins, K.W.; Klug, M.J. Feeding ecology of stream invertebrates. Annu. Rev. Ecol. Syst. 1979, 10, 147–172. [Google Scholar] [CrossRef]
- Rossi, L.; Fano, A.E. Role of fungi in the trophic niche of the congeneric detritivorous Asellus aquaticus and A. coxalis (Isopoda). Oikos 1979, 32, 380–385. [Google Scholar] [CrossRef]
- Willoughby, L.G.; Marcus, J.H. Feeding and growth in the isopod Asellus aquaticus on actinomycetes, considered as model filamentous bacteria. Freshw. Biol. 1979, 9, 441–449. [Google Scholar] [CrossRef]
- Suberkropp, K.; Arsuffi, T.L. Degradation, growth, and changes in palatability of leaves colonized by six aquatic hyphomycete species. Mycologia 1984, 76, 398–407. [Google Scholar] [CrossRef]
- Arsuffi, T.L.; Suberkropp, K. Selective feeding by shredders on leaf-colonizing stream fungi: Comparison of macroinvertebrate taxa. Oecologia 1989, 79, 30–37. [Google Scholar] [CrossRef] [PubMed]
- France, R.L. Leaves as “crackers”, biofilm as “peanut butter”: Exploratory use of stable isotopes as evidence for microbial pathways in detrital food webs. Oceanol. Hydrobiol. Stud. 2011, 40, 110–115. [Google Scholar] [CrossRef]
- Pereira, A.; Trabulo, J.; Fernandes, I.; Pascoal, C.; Cássio, F.; Duarte, S. Spring stimulates leaf decomposition in moderately eutrophic streams. Aquat. Sci. 2017, 79, 197–207. [Google Scholar] [CrossRef]
- Fisher, S.G.; Likens, G.E. Energy flow in Bear Brook, New Hampshire: An integrative approach to stream ecosystem metabolism. Ecol. Monogr. 1973, 43, 421–439. [Google Scholar] [CrossRef]
- Giller, P.; Twomey, H. Benthic macroinvertebrate community organisation in two contrasting rivers: Between-site differences and seasonal patterns. Biol. Environ. Proc. R. Ir. Acad. 1993, 93B, 115–126. [Google Scholar]
- Haapala, A.; Muotka, T. Seasonal dynamics of detritus and associated macroinvertebrates in a channelized boreal stream. Fundam. Appl. Limnol. 1998, 142, 171–189. [Google Scholar] [CrossRef]
- Gordon, T.A.C.; Neto-Cerejeira, J.; Furey, P.C.; O’Gorman, E.J. Changes in feeding selectivity of freshwater invertebrates across a natural thermal gradient. Curr. Zool. 2018, 64, 231–242. [Google Scholar] [CrossRef]
- Tinbergen, L. The natural control of insects in pinewoods. Arch. Néerl. Zool. 1960, 13, 265–343. [Google Scholar] [CrossRef]
- Dukas, R.; Kamil, A. Limited attention: The constraint underlying search image. Behav. Ecol. 2001, 12, 192–199. [Google Scholar] [CrossRef]
- Clark, C.W.; Dukas, R. The behavioral ecology of a cognitive constraint: Limited attention. Behav. Ecol. 2003, 14, 151–156. [Google Scholar] [CrossRef]
- Cross, F.R.; Jackson, R.R. Olfactory search-image use by a mosquito-eating predator. Proc. R. Soc. B Biol. Sci. 2010, 277, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Design of the field experiment. (A) Relative location of the leaf mats within Cedar Creek. Each mat is represented by a square and was at least 1 m from each of the nearest mats. G = on gravel benthos within a central riffle; S = on thin silt layer overlaying gravel within a bankside run/pool. The arrow indicates the direction of stream flow. (B) Depiction of individual leaf mat. Each square represents a leaf pack. B = American beech, M = maple (red maple and sugar maple), O = red oak. Lines between leaf packs indicate that packs were connected to each other at the corners via plastic zip ties, and black lines at corners represent plastic tent stakes that affixed the mat to the benthos.
Figure 1.
Design of the field experiment. (A) Relative location of the leaf mats within Cedar Creek. Each mat is represented by a square and was at least 1 m from each of the nearest mats. G = on gravel benthos within a central riffle; S = on thin silt layer overlaying gravel within a bankside run/pool. The arrow indicates the direction of stream flow. (B) Depiction of individual leaf mat. Each square represents a leaf pack. B = American beech, M = maple (red maple and sugar maple), O = red oak. Lines between leaf packs indicate that packs were connected to each other at the corners via plastic zip ties, and black lines at corners represent plastic tent stakes that affixed the mat to the benthos.

Figure 2.
Comparison of microhabitat and leaf species on isopod abundance. One week after submersion, there was no significant effect of leaf type nor of microhabitat on the average (+/−1SE) number of isopods in leaf packs. N = 9 for each bar. The large variance in the bankside maple treatment was largely due to one replicate. Without that replicate, the average (+/−1SE) for this treatment was 3.375 (+/−0.843), and the statistical conclusions remained unchanged.
Figure 2.
Comparison of microhabitat and leaf species on isopod abundance. One week after submersion, there was no significant effect of leaf type nor of microhabitat on the average (+/−1SE) number of isopods in leaf packs. N = 9 for each bar. The large variance in the bankside maple treatment was largely due to one replicate. Without that replicate, the average (+/−1SE) for this treatment was 3.375 (+/−0.843), and the statistical conclusions remained unchanged.

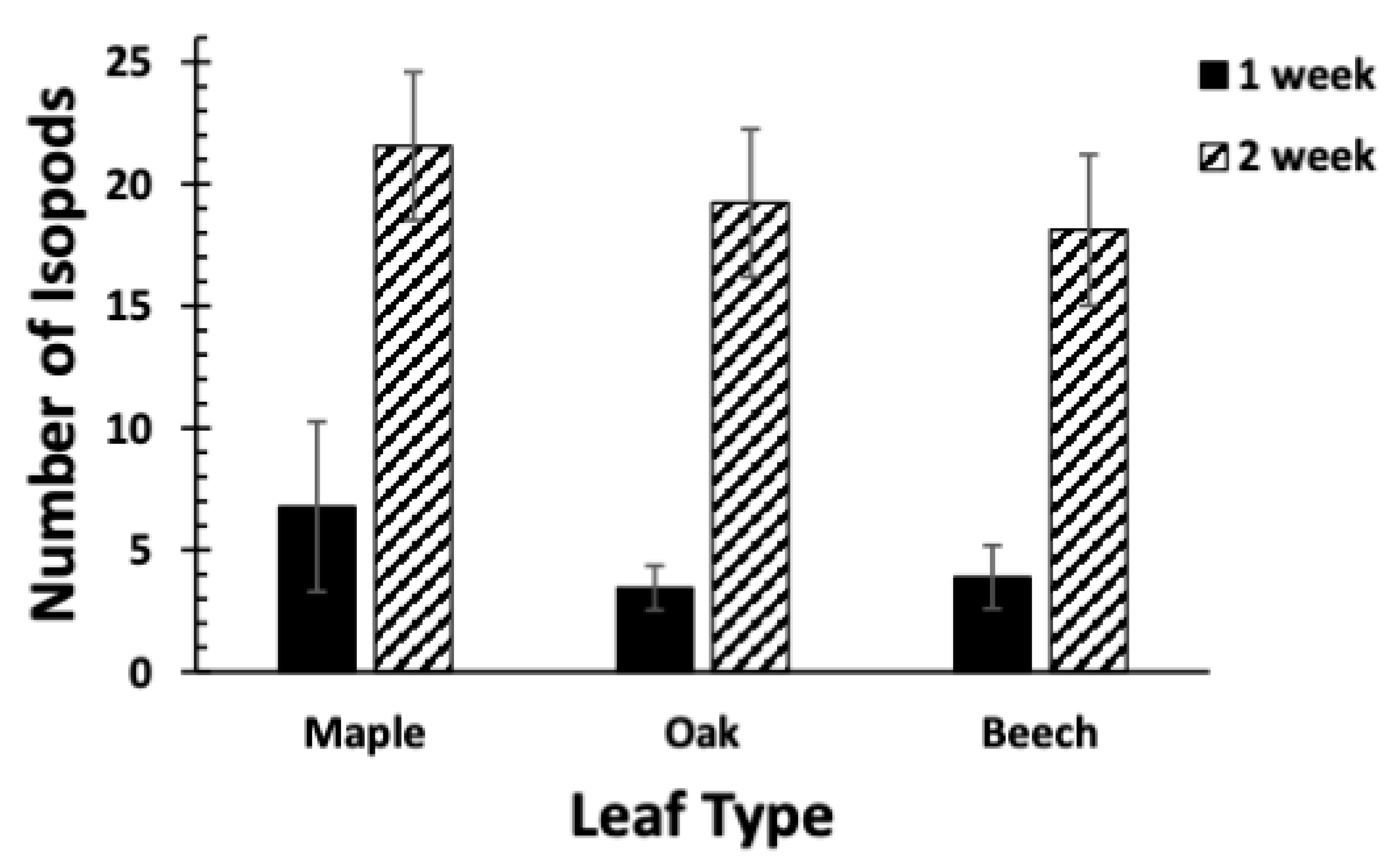
Figure 3.
Comparison of submersion time and leaf species on isopod abundance. There was a significant effect of duration of submersion, but no significant effect of leaf species, on the average (+/−1SE) number of isopods within each pack positioned within the bankside pool. N = 9 for each bar.
Figure 3.
Comparison of submersion time and leaf species on isopod abundance. There was a significant effect of duration of submersion, but no significant effect of leaf species, on the average (+/−1SE) number of isopods within each pack positioned within the bankside pool. N = 9 for each bar.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Statistical output for the three-way ANOVA analysis conducted using the isopod colonization data displayed in Figure 2 and Figure 3. The response variable (number of isopods within each leaf pack) was transformed using ln (y + 1) before conducting analyses to meet the assumptions inherent in an ANOVA.
Table 1.
Statistical output for the three-way ANOVA analysis conducted using the isopod colonization data displayed in Figure 2 and Figure 3. The response variable (number of isopods within each leaf pack) was transformed using ln (y + 1) before conducting analyses to meet the assumptions inherent in an ANOVA.
| Source | df | Sum of Squares | Mean Square | F-Ratio | p-Value |
|---|---|---|---|---|---|
| Intercept | 1 | 256.80 | 256.80 | 163.11 | 0.0002 |
| Leaf species | 2 | 0.34 | 0.17 | 0.34 | 0.72 |
| Week | 1 | 33.52 | 33.52 | 66.79 | <0.0001 |
| Microhabitat | 1 | 1.23 | 1.23 | 2.44 | 0.12 |
| Mat | 4 | 6.30 | 1.57 | 3.14 | 0.02 |
| Leaf species × Week | 2 | 0.03 | 0.02 | 0.03 | 0.97 |
| Leaf species × Microhabitat | 2 | 0.46 | 0.23 | 0.45 | 0.64 |
| Error | 68 | 34.13 | 0.50 | ||
| Total | 80 | 94.18 |
Table 2.
Number of leaf packs exhibiting designated degrees of leaf skeletonization after submersion in the stream for two weeks.
Table 2.
Number of leaf packs exhibiting designated degrees of leaf skeletonization after submersion in the stream for two weeks.
| Species of Leaf | Degree of Skeletonization | ||
|---|---|---|---|
| Minor | Moderate | Extreme | |
| American beech | 9 | 0 | 0 |
| Maple (red and sugar) | 2 | 3 | 4 |
| Red oak | 4 | 3 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Heller, R.E.; Stouffer, A.N.; Iyengar, E.V. Effects of Species of Leaves and Conditioning Time on Vernal Colonization by Temperate Lotic Isopods (Lirceus sp.). Hydrobiology 2024, 3, 63-73. https://doi.org/10.3390/hydrobiology3020005
AMA Style
Heller RE, Stouffer AN, Iyengar EV. Effects of Species of Leaves and Conditioning Time on Vernal Colonization by Temperate Lotic Isopods (Lirceus sp.). Hydrobiology. 2024; 3(2):63-73. https://doi.org/10.3390/hydrobiology3020005
Chicago/Turabian StyleHeller, Renee E., Alison N. Stouffer, and Erika V. Iyengar. 2024. "Effects of Species of Leaves and Conditioning Time on Vernal Colonization by Temperate Lotic Isopods (Lirceus sp.)" Hydrobiology 3, no. 2: 63-73. https://doi.org/10.3390/hydrobiology3020005