Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus


1
Marine Biology Department, Texas A&M University at Galveston, Galveston, TX 77554, USA
2
Biology Department, Muhlenberg College, Allentown, PA 18104, USA
*
Author to whom correspondence should be addressed.
Hydrobiology 2024, 3(2), 74-84; https://doi.org/10.3390/hydrobiology3020006
Submission received: 4 March 2024
/
Revised: 12 April 2024
/
Accepted: 17 April 2024
/
Published: 23 April 2024
Abstract
:Marine hermit crabs inhabiting different species of shells in the rocky intertidal may receive varied benefits and costs. Thick shell walls, sculpture, and certain silhouettes may increase protection against crushing predators and waves, but also the energy cost for bearing such a shell. To determine the relative energy demands on Pagurus granosimanus supporting one of three different species of shells commonly inhabited in nature, we measured the consumption of food by free-roaming, size-matched individuals after four days of food deprivation in the laboratory. Surprisingly, while all hermit crabs consumed a significant amount of food, that amount did not differ significantly across the types of shells inhabited, despite the significantly heavier weight of one type of shell (>150% of each of the other two species), and markedly different shell silhouettes. Whether our results were due to the inherent abilities of arthropods to support proportionally immense weights (physiology), reduced movements by animals in heavier shells (plastic behavior), or the way the hermit crabs bear the burden of the shell requires further investigation. Regardless, our results suggest that individuals of P. granosimanus that inhabit heavier shells that likely provide increased protection do not experience corresponding elevated food consumption requirements over the short term, even if those shells are not centered over the body mass of the hermit crab. Pagurus granosimanus crabs can bear the burden.
1. Introduction
Hermit crabs typically inhabit empty gastropod shells to gain protection against predators and the harsh, intertidal environment [1,2,3,4,5], but inappropriate domiciles can exert severe costs on the inhabitant [4,6,7,8]. Hermit crabs, including our study species Pagurus granosimanus (Stimpson, 1859), typically occupy a distinct subset of the available gastropod shells in the wild [9] and predominantly select certain species when given access to a wide array in the laboratory [5]. These choices may be driven, in part, by the comparative weight of the shells. A large, thick-walled shell with a robust apertural lip and calcified shell decorations such as knobs, spines, and ribs may provide ample room to grow and resilient protection against generalized durophagous predators [5,10,11]. Vance [3] experimentally demonstrated that individuals of P. granosimanus that inhabited smaller shells were at higher risk of crab predation, and Spight [12] postulated that hermit crabs rarely used the species Lirabuccinum dirum’s available shells due to its thin walls. Shell weight can also provide hydrodynamic advantages in wave-exposed areas [13,14].
However, a large, thick-walled shell possesses considerable heft, which is likely to increase energy expenditures for a hermit crab that continually bears that shell during daily foraging activities [15,16]. Heavier shells display slow growth [5] and references therein, [17]. In hypoxic conditions, some hermit crabs shift to significantly lighter choices [18], and others shift from heavier, conical shells to smaller, lighter, globose ones when moving to areas of slower-flowing water [14]. Female hermit crabs of some species tend to utilize lighter shells than do the males, perhaps due to the additional weight of eggs [19,20]. These observations suggest there can be a significant, potentially limiting, cost to bearing a heavier shell. Shell inadequacy must constitute a powerful evolutionary force in hermit crab biology [3]), and shell weight has been posited as one of the determining factors of shell value [21]): thicker shells may be more protective, but higher energetic demands incurred by supporting heavier, off-centered shells may elevate caloric intake requirements and increase the need for time spent foraging.
To explore these potential tradeoffs with P. granosimanus, a common hermit crab species in the Pacific Northwest rocky intertidal, we examined whether carrying shells of different species commonly inhabited in nature (Calliostoma ligatum (Gould, 1849), Nucella lamellosa (Gmelin, 1791), or Lirabuccinum dirum (Reeve, 1846); Figure 1) led individuals to consume increased amounts of food as an indication of increased energetic expenditures (similar to the need for increased calorie consumption of many human athletes compared to non-athletes [22]). Because hermit crabs can be removed from their existing shell relatively easily without harm to the animal, and will readily enter almost any shell rather than remaining shell-less, this study system allowed us to create treatments of size-matched hermit crabs bearing shells of different species. We predicted that, after being deprived of food for four days and then allowed to eat ad libitum for 24 h, individuals in species with heavier shells that are not as centered over the crab would consume more food. Thus, we predicted that hermit crabs in N. lamellosa would consume the most food in trials, and those in L. dirum would consume the least.
2. Materials and Methods
2.1. Food Consumption Experiment
To test the relative energy demand that could result in different levels of hunger caused by supporting different species of shells, large individuals of Pagurus granosimanus (0.2 g to 1.3 g) were collected from Colin’s Cove (48°33′4″ N, 123°0′20″ W) on the east side of San Juan Island, Washington state. Each hermit crab was transported to the laboratory and size-matched (±1 mm, as measured using a digital caliper) to a single new snail host shell based on the aperture length of the shell it inhabited when collected in the field. The aperture length of the shell is likely a limiting factor for the acceptability of a potential domicile because a hermit crab’s widest point (across its cephalothorax) will need to fit inside the shell to allow the crab to completely withdraw when threatened [23]. The shells occupied by hermit crabs in these experiments ranged in aperture length from 7.26 to 12.95 mm (average = 9.75; 1SE = 0.20).
Each hermit crab was removed from its shell using a locking adjustable crescent wrench that allowed incremental pressure to gently crack the shell. Once the shell had fractured and large sections fell free, the hind end of the animal was gently prodded with a probe to encourage the animal to voluntarily abandon the shell remnants. From this stage forward, the hermit crabs (and no-crab treatments, explained below) were housed separately in plastic flow-through containers (20 cm × 14 cm × 10 cm with mesh inserts along the side lengths, pore size = 1 mm). The shell-less hermit crab was placed alone in a flow-through tub on a sea table with its single-size-matched experimental shell. A sea table is a large tank in the laboratory with seawater continuously flowing in from the ocean and draining out. The sea table maintained a constant water level that kept the experimental containers fully submerged with a gentle aerating flow through them, but no large food particles entered with the inflow, and no other animals were maintained in the sea table during the experiment. Each new shell provided was one of three species, which represented the experimental treatments: Nucella lamellosa, Calliostoma ligatum, or Lirabuccinum dirum. These shell species differ both in relative weight (Figure 2A) and shell silhouette, with N. lamellosa and L. dirum displaying a greater length:width ratio than C. ligatum that projects to a greater degree along the horizontal axis from the hermit crab’s body (i.e., the last species is a more squat, vertically oriented domicile than the other two species; Figure 1).
Experimental shells (Table 1) were acquired by collecting live snails at Colin’s Cove, freezing them for 24 h, then removing the snail tissue using manual agitation and multiple washes with fresh water for 3 to 4 days until no more snail tissue remained in the shells. Empty shells were then allowed to dry before use in experiments. Thus, all treatments utilized recently vacated, intact shells; empty shells weaken rapidly post-mortem ([24] and the structural integrity of shells is important in hermit crab choice [25]. The deviation of the aperture length of the newly inhabited experimental versus the original shell was not significantly different across treatments (Figure 2B; ANOVA; F2,44 = 2.79, p =0.0721, n = 48).
All experimental animals readily entered, and remained in the new shell. Because hermit crabs are extremely averse to remaining shell-less but empty shells are relatively rare [5,9], our experimental design, where a naked crab is presented with only one choice, likely mimics natural situations. We did not use ovigerous females, but we did not otherwise distinguish between male and female crabs, as Hazlett [5] found that shell preferences were similar between sexes (except as a byproduct of sex-related size differences), even when females are ovigerous. Each treatment was replicated 16 times. All replicates were placed in a single sea table, with treatments alternating positions within the sea table.
When presenting hermit crabs with shells, the hermit crabs were organized into size-matched triplets (aperture length ± 1 mm) to control for any effect body size might have on the food consumption rate, with each of the three experimental shell species inhabited across the triplet. Separate one-way ANOVAs, both using treatment as the independent variable, confirmed that our attempts to size-match hermit crabs across treatments were successful: there were no significant differences among treatments in either the starting weights of shell-less hermit crabs (Figure 2B; F2,45 = 0.375, p = 0.689) or cheliped widths of the crabs (Figure 2C; F2,45 = 0.217, p = 0.806). The species of shell the hermit crab inhabited when originally collected was recorded but had no bearing on the species of shell we assigned to the animal in our experiments. To confirm that the hermit crabs did consume a significant amount of the provided food (i.e., the food was palatable), we also had a treatment (the no-crab treatment) that consisted of replicates without hermit crabs present.
The animals were held for four days without food to allow them to move freely and experience any energetic costs of bearing the shell without consuming anything (i.e., elevating their hunger levels). Previous food acquisition experiments for a congeneric species (Pagurus samuelis) precluded feedings for four to seven days prior to testing [26]. Occasional observations of the animals multiple times each day indicated that all hermit crabs remained in their shells and moved around the containers, including vertically climbing the walls of the container by grasping the mesh of the inserts. After four days without providing food, ~1.0 g of imitation crab meat (TransOcean brand, which is processed fish, typically pollock) was placed in each replicate container for 24 h. All food was procured from the same individual package and all trials were started on the same day to minimize any differences in food across the replicates. In seawater, imitation crab meat neither gained mass, crumbled, or decayed substantially over the length of our experimental time period [27], and personal observations, and this amount of food greatly exceeded what any of the crabs could consume in this time period. The range of the weights of initial food provisions was 0.994 g to 1.078 g (a 0.084 g difference), ensuring equitable starting points and enabling us to compare the amount of food remaining across the treatments as an accurate indication of relative consumption.
At the end of the feeding period, food fragments not eaten by the hermit crabs were collected, gently rinsed with fresh water, then dried for 24 h at 63 °C, and re-weighed (to three decimal places). The food was dried upon the termination of the experiment to avoid any variation in hand-drying that might leave behind water droplets, as these could add significant mass if the hermit crabs shredded the food. The drying process precluded us from merely subtracting the final weight from the initial weight to determine the mass consumed by the crabs. Instead, our experimental dependent response was the dry weight of unconsumed food; animals with larger numbers ate less during the experimental period. The weights of the food portions initially provided in containers did not differ significantly across the treatments (ANOVA; F3,59 = 0.715, p = 0.547).
2.2. Data Analyses
We conducted separate analyses of variance tests (ANOVAs) to compare the effect of the shell species (the independent variable) on the dry weights of the remaining food at the end of the experiment (the dependent variable), as well as on the relative weights of the shell domiciles and of the hermit crabs used in the experiment to assess the consistency of our experimental design across treatments (n = 16). In addition to ANOVAs, we also examined the data on the weights of remaining food by calculating Bayes factors using the oneWayAOV.Fstat function from the BayesFactor package in RStudio [28]. This is a comparative test providing the likelihood of the alternative hypothesis compared to the null hypothesis. Numbers greater than zero show stronger support for the alternative hypothesis; numbers less than zero show stronger support for the null hypothesis. We also used an ANCOVA to determine whether the size of the animals (as indicated by their weight) and the type of shell of the hermit crab significantly affected the amount of food eaten.
All statistical tests were conducted using RStudio version 2023.06.1+524 (Posit Software, PBC) with an alpha-level of p < 0.05 as the threshold of significance, and we calculated Tukey’s pairwise post-hoc comparisons if the overall effect of the independent variable was significant.
3. Results
There was a significant difference among the weights of the species of shells we presented to the hermit crabs (Figure 2A; ANOVA; F2,45 = 55.65, p < 0.0001), with N. lamellosa being significantly heavier than either C. ligatum or L. dirum, but there was no significant difference between the last two species (p = 0.68).
There was a significant effect of treatment on the weight of food remaining (Figure 3; ANOVA; F3,60 = 18.09, p < 0.0001). Subsequent pair-wise comparisons revealed that each of the three treatments with hermit crabs had significantly less food remaining than in the no-crab treatment (p < 0.0001 for each comparison), indicating that the hermit crabs in all types of shells consumed significant amounts of food. However, across the three shell treatments, there was no significant difference in the amount of food remaining (p > 0.4 for each comparison).
The Bayes factor of the entire data set of the no-crab control and all three shell treatments was 32.82, indicating strong support for the hypothesis that there was a significant difference across all treatments in the amount of food remaining. In contrast, among only the three shell treatments the Bayes factor was −1.05, indicating support for the null hypothesis (no effect of type of shell on the amount of food remaining).
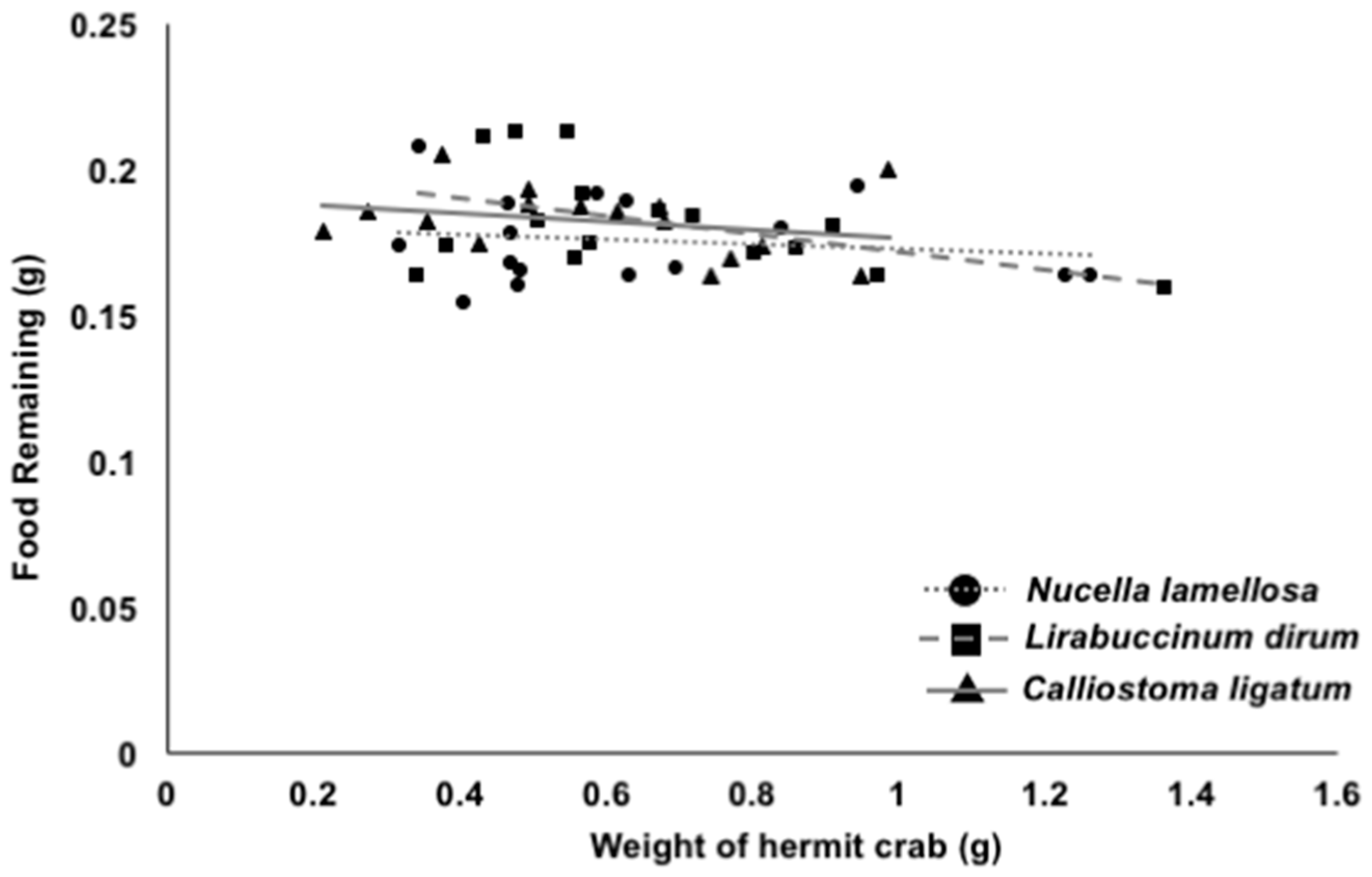
There was a significant negative relationship between hermit crab size and the amount of food remaining (Figure 4, ANCOVA; F1,42 = 4.5203, p = 0.0391), indicating that larger crabs consumed more food. However, there was no significant effect of the type of shell on the amount of food remaining (ANCOVA; F1,42 = 1.15, p = 0.3262).
4. Discussion
Surprisingly, despite the large differences in shell silhouette and weight among our experimental treatments (the average weight of shells of N. lamellosa registered > 3.28 g more, a > 150% increase, than the average weight of either of the other two shell types), hermit crabs supporting the various shells did not consume significantly different amounts of food, suggesting they did not differ in hunger level. Our current findings appear to contradict many previous studies e.g., [29] and others cited in our introduction] that proposed shell weight as an important factor in domicile choice. While Bell [30] noted that hermit crabs will endure an increased energetic cost of carrying epibiont-covered homes, our results suggest that in our study species, the additional energy required may be minimal.
While our results conflict with others’ findings (see introduction), they agree with other studies involving both tropical and temperate hermit crabs. Hazlett [31] noted that the rate of daily movements of a tropical hermit crab was not significantly affected by the weight or species of the shell inhabited. Additionally, a temperate pagurid species showed no effect of shell weight on levels of activity or lactate in the hemolymph, suggesting that heavier shells do not result in fatigue [32], and other hermit crabs carrying shells four times their body weight do not increase their VO2 levels (Herreid and Full, 1996). In retrospect, it is perhaps not surprising the additional mass borne in our experiments did not elevate the rates of food consumption. Some arthropods regularly carry 8.5 times their own body weight, or have joints that can withstand 5000 times their body weight [33,34,35]; these adaptations suggest regular bearing of burdens much greater than our heaviest shells. Additionally, because empty, intact shells are often limited locally for this crab [9], the frequency of acceptable shells may require that hermit crabs sustainably bear a wide range of weights [36].
However, our results may be due, at least in part, to the way the various shells were situated on the hermit crabs. A shell that is heavier when weighed in isolation is not necessarily a heavier burden for a hermit crab to bear. The shape of the shell, combined with its internal configuration, can cause substantial variation in the way it is carried: the angle, how much is centered over the hermit crab’s body, and how much shell is at least semi-supported by the ground (Figure 1). As such, the degree of burden and ensuing stress incurred by the inhabitant may not be correlated with the mere raw weight of the shell but altered (elevated or reduced) by aspects of the shell form that are impacted by growth variables [37].
Longer-term (90-day) studies with a tropical hermit crab have shown that increasing shell weight through the use of shells of different species resulted in a concomitantly decreased growth rate of the hermit crab inhabitant [38]. However, a close examination of Alcaraz et al. [38], see their Figure 1) reveals that hermit crabs in the second-lightest shells (which did not differ significantly in weight from the lightest shells) did not grow significantly more quickly than animals in the two species of shells that were significantly heavier, and even possessed a lower average than the crabs inhabiting one species of the heavier shell. These results suggest that something other than shell weight was affecting the growth rate of the hermit crabs. We recommend repeating experiments similar to Alcaraz et al. [38] by manipulating the weights of a single species of shell.
Our experimental time frame, after only four days of food deprivation and one day for consumption, may have been too short for these aquatic poikilotherms with a relatively low metabolic rate to demonstrate noticeable differences in consumption. However, our length of food deprivation is within the range of that used by other researchers. For instance, Micheli [39] reported declines in prey selectivity in blue crabs after only three days of food deprivation. Working with other species of pagurids, Angel [27], Alcaraz and Garcia-Cabello [16], and Billock and Dunbar [40] conducted trials after (respectively) two, five, or eight days without food. Similar to our results, Angel [27] reported that hermit crabs in tightly fitting shells (assumedly smaller and lighter) did not feed at different rates than individuals in shells of preferred size.
Furthermore, we opted for a shorter experimental food deprivation period to encourage hunger, but not starvation. One crab species revealed a peak of prey searching time and less stationary time in treatments of six and nine days without food [41]. Two other crab species showed a peak in predation rate and time spent foraging after six days without food, but rates lowered in those starved for nine days [42]. We utilized only four days of food deprivation to allow the hermit crabs time to engage in substantial foraging movements, while avoiding exceeding the hunger threshold that might cause the animals to reduce metabolism [16]. We waited only 24 h after placing food in the containers before removing it because past work on spider crabs noted maximum rates of consumption on the first day with a steady decline afterwards [43].
Our experimental arena, which lacked significant wave action, may have skewed results by reducing drag forces, but the lower depths of tide pools, which is where these species are often found [44] working in California; our personal observations], would also have low flow rates. Potentially, our results may merely reflect reduced movements by the animals in heavier shells, as has been shown in some systems e.g., [16]. But, Rittschof et al. [45] showed that hermit crabs in heavy shells are still sufficiently mobile, and in each of our occasional (more than daily) checks of experimental containers, all of the crabs seemed to have engaged in exploratory movements that altered their location in the container. Additionally, we used animals in experiments within a day of field collection, so they had likely not yet resigned themselves to a lack of alternative shells within the arena (if, indeed, hermit crabs make such a decision). In the field, most species of hermit crabs live in areas with a shortage of intact and sturdy empty gastropod shells [5], which is also true for our current study species [9]. Thus, to find a better shell than the one currently possessed would typically necessitate searching behaviors. Therefore, crabs in nature with heavier-than-ideal shells would need to search for better options and thus not reduce their movements to an extreme amount.
On San Juan Island, a high percentage of individuals of P. granosimanus readily switch into larger shells when available [17], and intact larger-sized shells are typically more limited than smaller shells [46,47]. It is possible because we used larger individuals from the population and matched the size of the shells we provided in the experiments to the size of shells the hermit crabs inhabited in the field, that the hermit crabs had been inhabiting substantially smaller-than-ideal shells in the field. If so, the additional weight burden of thicker-shelled N. lamellosa may have been an easy extra burden for those crabs to shoulder.
While our data did show that there was a significant effect of hermit crab size on the amount of food left behind, the slope to the line was very small and there was a poor fit of the line to the data. Hermit crabs across our (intentionally) small range of crab sizes ate almost the same amount, even if they were carrying the heaviest species of shell and were proportionally small for the size of shell they inhabited.
All of these results imply that shell weight alone is not the determining factor of its burden of energetic demand. Hermit crab behavior can be plastic. Hermit crabs alter shell choice in the presence of chemical cues from various predators [48,49], and one species will even choose less-preferred shells if only that shell allows escape from an enclosure [50]. In some species of hermit crab, the cost of walking (measured as oxygen consumption) increases with shell mass, but the amount of exploratory walking is negatively correlated with the heaviness of the shell, suggesting that reduced activity enables the hermit crabs to deal with the high energetic costs of foraging in heavy shells [16]. While we noted that all of the crabs moved during the course of our experiment, we did not attempt to quantify the amount or rate of movement. While Herreid and Full [51] observed that shell-bearing hermit crabs had higher levels of oxygen consumption than conspecifics without shells, as animals increased their speed, there was a marked decrease in the relative cost of transport. The authors surmised this reduction might partially be because crabs carrying shells altered leg positions to drag the shells and thus reduce the burdens supported by the animals. As illustrated above (Figure 1), P. granosimanus appears to support proportionally more of the weight of L. dirum shells while the other two shell species rest partially on the substratum. Thus, the relative weights to support may not be as dramatic as suggested by Figure 2A. Future studies should examine the amount and speeds of movement by the crabs, as well as note the amount of time that shells are dragged as opposed to fully elevated. Additionally, all of these aspects should also be examined in the air as well as while the animals are submerged, as the buoyancy of water may affect the cost. Further investigations should determine whether there are differences among the tropical and temperate hermit crabs, or species-to-species differences, as to whether the options available are close to the maximum weight limit of easily borne shells, or whether previous experiments demonstrating extreme costs overlooked other aspects of different species of shells that were more important than weight differences (such as how the shell is carried).
While the heavy-walled N. lamellosa was the second-most commonly inhabited species of shell for P. granosimanus at our collection site, the least commonly used (but acceptable) species was the lighter L. dirum. The most commonly inhabited shell at this site, Margarites pupillus [9], has a much thinner shell wall than N. lamellosa (more comparable to C. ligatum and L. dirum) and the shell silhouette is more similar to C. ligatum than to N. lamellosa. At this site, empty shells of M. pupillus (66) far outnumbered empty shells of any of the other species, with N. lamellosa a distant second (4) and almost no empty L. dirum (1) or C. ligatum (0) found [9]. Because P. granosimanus apparently has strategies or morphologies that reduce the relative energetic demands of bearing differently shaped and weighted shells, the frequency of available shells may subsume selective pressures due to shell weight or silhouette. From our current results, we conclude that individuals of free-ranging P. granosimanus do not experience short-term energetic demands that then drive different levels of food consumption when inhabiting size-matched shells of the thin-walled, squat C. ligatum, the thin-walled, elongate L. dirum, or the thicker-walled, elongate N. lamellosa.
Author Contributions
Both authors participated in each stage of the research described herein, with the exception that B.Z.T. conducted the laboratory investigation work and wrote the original draft of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
We would like to thank the Crist Family Student Research Award in Biology Fund of Muhlenberg College and the Blinks-NSF REU-BEACON program of Friday Harbor Laboratories (NSF grant number DBI-1262239) for financial support of BZT, and a Muhlenberg College faculty summer research grant to EVI.
Institutional Review Board Statement
Ethical review and approval were not required for this study as it involved only invertebrate animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available upon reasonable request to the corresponding author.
Acknowledgments
We would like to thank the San Juan County Land Bank and Friday Harbor Laboratories for site access and James Russell for statistical advice.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Hazlett, B.A. Social behavior of the Paguridae and Diogenidae of Curacao. Stud. Fauna Curacao Other Caribb. Isl. 1966, 88, 1–143. [Google Scholar]
- Reese, E.R. Behavioral adaptations of intertidal hermit crabs. Am. Zool. 1969, 9, 343–355. [Google Scholar] [CrossRef]
- Vance, R.R. The role of shell adequacy in behavioral interactions involving hermit crabs. Ecology 1972, 53, 1075–1083. [Google Scholar] [CrossRef]
- Fotheringham, N. Population consequences of shell utilization by hermit crabs. Ecology 1976, 57, 570–578. [Google Scholar] [CrossRef]
- Hazlett, B.A. The behavioral ecology of hermit crabs. Annu. Rev. Ecol. Syst. 1981, 12, 1–22. [Google Scholar] [CrossRef]
- Markham, J.C. Notes on growth-patterns and shell-utilization of the hermit crab Pagurus bernhardus. Ophelia 1968, 5, 189–205. [Google Scholar] [CrossRef]
- Nyblade, C.F. Coexistence in Sympatric Hermit Crabs. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 1974. [Google Scholar]
- Abrams, P.; Nyblade, C.; Sheldon, S. Resource partitioning and competition for shells in a subtidal hermit crab species assemblage. Oecologia 1986, 69, 429–445. [Google Scholar] [CrossRef] [PubMed]
- Torjman, B.Z.; Iyengar, E.V. Increased potential for shell competition among intertidal hermit crabs (Pagurus beringanus and Pagurus granosimanus) in the rocky intertidal. J. Mar. Biol. Assoc. UK 2021, 101, 957–967. [Google Scholar] [CrossRef]
- Vermeij, G.J. Marine faunal dominance and molluscan shell form. Evolution 1974, 28, 656–664. [Google Scholar] [CrossRef]
- Bertness, M.D.; Cunningham, C. Crab shell-crushing predation and gastropod architectural defense. J. Exp. Mar. Biol. Ecol. 1981, 50, 213–230. [Google Scholar] [CrossRef]
- Spight, T.M. Availability and use of shells by intertidal hermit crabs. Biol. Bull. 1977, 152, 120–133. [Google Scholar] [CrossRef]
- Arce, E.; Alcaraz, G. Shell preference in a hermit crab: Comparison between a matrix of paired comparisons and a multiple-alternative experiment. Mar. Biol. 2012, 159, 853–862. [Google Scholar] [CrossRef]
- Alcaraz, G.; Toledo, B.; Burciaga, L.M. The energetic costs of living in the surf and impacts on zonation of shells occupied by hermit crabs. J. Exp. Biol. 2020, 223, jeb222703. [Google Scholar] [CrossRef]
- Briffa, M.; Elwood, R.W. Metabolic consequences of shell choice in Pagurus bernhardus: Do hermit crabs prefer cryptic or portable shells? Behav. Ecol. Sociobiol. 2005, 59, 143–148. [Google Scholar] [CrossRef]
- Alcaraz, G.; Garcia-Cabello, K.N. Feeding and metabolic compensations in response to different foraging costs. Hydrobiologia 2017, 787, 217–227. [Google Scholar] [CrossRef]
- Abrams, P.A. Resource partitioning and competition for shells between intertidal hermit crabs on the outer coast of Washington. Oecologia 1987, 72, 248–258. [Google Scholar] [CrossRef]
- Cote, I.M.; Reverdy, B.; Cooke, P.K. Less choosy or different preference? Impact of hypoxia on hermit crab shell assessment and selection. Anim. Behav. 1998, 56, 867–873. [Google Scholar] [CrossRef]
- Elwood, R.W.; Marks, N.; Dick, J.T.A. Consequences of shell-species preferences for female reproductive success in the hermit crab Pagurus bernhardus. Mar. Biol. 1995, 123, 431–434. [Google Scholar] [CrossRef]
- Caruso, T.; Chemello, R. The size and shape of shells used by hermit crabs: A multivariate analysis of Clibanarius erythropus. Acta Oecologica 2009, 35, 349–354. [Google Scholar] [CrossRef]
- Reese, E.S. The behavioral mechanisms underlying shell selection by hermit crabs. Behaviour 1963, 21, 78–126. [Google Scholar] [CrossRef]
- Lopes da Silva, D.J.; Oliveira, T.A.; Oliveira, A. Body composition and food intake in athletes and non athletes portuguese male adolescents. In Stochastic Modeling Data Analysis & Statistical Applications, 2nd ed.; Filus, L., Oliveira, T., Skiadas, C.H., Eds.; ISAST: Groningen, The Netherlands, 2015; pp. 215–224. [Google Scholar]
- Turra, A.; Leite, F.P.P. Shell-size selection by intertidal sympatric hermit crabs. Mar. Biol. 2004, 145, 251–257. [Google Scholar] [CrossRef]
- LaBarbera, M.; Merz, R.A. Postmortem changes in strength of gastropod shells: Evolutionary implications for hermit crabs, snails, and their mutual predators. Paleobiology 1992, 18, 367–377. [Google Scholar] [CrossRef]
- Pechenik, J.A.; Hsieh, J.; Owara, S.; Wong, P.; Marshall, D.; Untersee, S.; Li, W. Factors selecting for avoidance of drilled shells by the hermit crab Pagurus longicarpus. J. Exp. Mar. Biol. Ecol. 2001, 262, 75–89. [Google Scholar] [CrossRef]
- Billock, W.L.; Dunbar, S.G. Shell and food acquisition behaviors: Evidence for contextual decision hierarchies in hermit crabs. J. Exp. Mar. Biol. Ecol. 2011, 398, 26–32. [Google Scholar] [CrossRef]
- Angel, J.E. Effects of shell fit on the biology of the hermit crab Pagurus longicarpus (Say). J. Exp. Mar. Biol. Ecol. 2000, 243, 169–184. [Google Scholar] [CrossRef]
- Renganathan, V. Overview of Bayesian Approach to Statistical Methods. 2022; pp. 10–76. Available online: https://www.amazon.com.au/Overview-Bayesian-Approach-Statistical-Methods/dp/9356201188 (accessed on 16 April 2024).
- Hahn, D.R. Hermit crab shell use patterns: Response to previous shell experience and to water flow. J. Exp. Mar. Biol. Ecol. 1998, 228, 35–51. [Google Scholar] [CrossRef]
- Bell, J.J. Influence of occupant microhabitat on the composition of encrusting communities on gastropod shells. Mar. Biol. 2005, 147, 653–661. [Google Scholar] [CrossRef]
- Hazlett, B.A. Daily movements of the hermit crab Clibanarius vittatus. Bull. Mar. Sci. 1981, 31, 177–183. [Google Scholar]
- Doake, S.; Scantlebury, M.; Elwood, R.W. The costs of bearing arms and armour in the hermit crab Pagurus bernhardus. Anim. Behav. 2010, 80, 637–642. [Google Scholar] [CrossRef]
- Daly-Schveitzer, S.; Beugnon, G.; Lachaud, J.P. Prey weight and overwhelming difficulty impact the choice of retrieval strategy in the Neotropical ant Gnamptogenys sulcata (F. Smith). Insectes Sociaux 2007, 54, 319–328. [Google Scholar] [CrossRef]
- Nguyen, V.; Lilly, B.; Castro, C.E. The exoskeletal structure and tensile loading behavior of an ant neck joint. J. Biomech. 2014, 47, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Segre, P.S.; Taylor, E.D. Large ants do not carry their fair share: Maximal load-carrying performance of leaf-cutter ants (Atta cephalotes). J. Exp. Biol. 2019, 222, jeb199240. [Google Scholar] [CrossRef]
- Young, A.M. Differential utilization of gastropod shells by three hermit crab species in North Inlet, South Carolina, U.S.A. In Crustaceana. Supplement, No. 5, Studies on Decapoda (Biology, Ecology, Morphology, and Systematics); Brill: Leiden, The Netherlands, 1979; pp. 101–104. [Google Scholar]
- Raup, D.M.; Michelson, A. Theoretical morphology of the coiled shell. Science 1965, 147, 1294–1295. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, G.; Chávez-Solís, C.E.; Kruesi, K. Mismatch between body growth and shell preference in hermit crabs is explained by protection from predators. Hydrobiologia 2015, 743, 151–156. [Google Scholar] [CrossRef]
- Micheli, F. Behavioural plasticity in prey-size selectivity of the blue crab Callinectes sapidus feeding on bivalve prey. J. Anim. Ecol. 1995, 64, 63–74. [Google Scholar] [CrossRef]
- Billock, W.L.; Dunbar, S.G. Influence of motivation on behaviour in the hermit crab, Pagurus samuelis. J. Mar. Biol. Assoc. UK 2009, 89, 775–779. [Google Scholar] [CrossRef]
- Long, Q.; Liu, J.; Sun, Y.; Yang, Z.; Tang, B.; Cheng, Y. The effect of food deprivation on foraging behavior and digestive and metabolic capacities of the Chinese mitten crab, Eriocheir sinensis. Fishes 2023, 8, 47. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Dong, S. A comparative study of the effect of starvation regimes on the foraging behavior of Portunus trituberculatus and Charybdis japonica. Physiol. Behav. 2015, 151, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, J.C. Allometric studies on energy relationships in the spider crab Libinia emarginata (Leach). Biol. Bull. 1974, 147, 257–273. [Google Scholar] [CrossRef]
- Orians, G.H.; King, C.E. Shell selection and invasion rates of some Pacific hermit crabs. Pac. Sci. 1964, 18, 297–306. [Google Scholar]
- Rittschof, D.; Sarrica, J.; Rubenstein, D. Shell dynamics and microhabitat selection by striped legged hermit crabs, Clibanarius vittatus (Bosc). J. Exp. Mar. Biol. Ecol. 1995, 192, 157–172. [Google Scholar] [CrossRef]
- Vance, R.R. Competition and mechanism of coexistence in three sympatric species of intertidal hermit crabs. Ecology 1972, 53, 1062–1074. [Google Scholar] [CrossRef]
- Garcia, R.B.; Mantelatto, F.L. Shell selection by the tropical hermit crab Calcinus tibicen (Herbst, 1791) (Anomura, Diogenidae) from Southern Brazil. J. Exp. Mar. Biol. Ecol. 2001, 265, 1–14. [Google Scholar] [CrossRef]
- Alcaraz, G.; Arce, E. Predator discrimination in the hermit crab Calcinus californiensis: Tight for shell breakers, loose for shell peelers. Oikos 2017, 126, 1299–1307. [Google Scholar] [CrossRef]
- Rotjan, R.D.; Blum, J.; Lewis, S.M. Shell choice in Pagurus longicarpus hermit crabs: Does predation threat influence shell selection behavior? Behav. Ecol. Sociobiol. 2004, 56, 171–176. [Google Scholar]
- Krieger, J.; Hörnig, M.K.; Laidre, M.E. Shells as ‘extended architecture’: To escape isolation, social hermit crabs choose shells with the right external architecture. Anim. Cogn. 2020, 23, 1177–1187. [Google Scholar]
- Herreid, C.F.; Full, R.J. Energetics of hermit crabs during locomotion: The costs of carrying a shell. J. Exp. Biol. 1996, 120, 297–308. [Google Scholar] [CrossRef]
Figure 1.
Front and side or posterior views of Pagurus granosimanus in shells of the various experimental treatments: (A,B) Nucella lamellosa (the number on the shell is from a different experiment); (C,D) Calliostoma ligatum; (E,F) Lirabuccinum dirum.
Figure 1.
Front and side or posterior views of Pagurus granosimanus in shells of the various experimental treatments: (A,B) Nucella lamellosa (the number on the shell is from a different experiment); (C,D) Calliostoma ligatum; (E,F) Lirabuccinum dirum.

Figure 2.
Metrics affiliated with the experimental design. (A) The shells in the various treatments did differ significantly in average weight. Individuals of P. granosimanus were placed in appropriately sized shells, as indicated by (B), which is the average aperture lengths of the natural and experimental shells. Hermit crabs were successfully size-matched across treatments as indicated both by (C), the average shell-less body weights of the crabs, and (D), the average widths of chelae. Numbers within the bars indicate sample size; all errors, bars are ±1SE; different lowercase letters above the bars signify statistically significant pair-wise comparisons.
Figure 2.
Metrics affiliated with the experimental design. (A) The shells in the various treatments did differ significantly in average weight. Individuals of P. granosimanus were placed in appropriately sized shells, as indicated by (B), which is the average aperture lengths of the natural and experimental shells. Hermit crabs were successfully size-matched across treatments as indicated both by (C), the average shell-less body weights of the crabs, and (D), the average widths of chelae. Numbers within the bars indicate sample size; all errors, bars are ±1SE; different lowercase letters above the bars signify statistically significant pair-wise comparisons.

Figure 3.
Average weights (±1SE) of food remaining in each container after 24 h. Numbers within the bars indicate sample size. Different letters above the bars indicate statistically significant pair-wise differences.
Figure 3.
Average weights (±1SE) of food remaining in each container after 24 h. Numbers within the bars indicate sample size. Different letters above the bars indicate statistically significant pair-wise differences.

Figure 4.
The weight of each hermit crab compared with the amount of food remaining, grouped by each type of shell treatment. The best-fit line for each shell type was as follows: N. lamellosa: y = −0.008x + 0.1806, R2 = 0.024; L. dirum: y = −0.0308x + 0.2026, R2 = 0.22; and C. ligatum: y = −0.014x + 0.1909, R2 = 0.078. n = 16 for each type of shell.
Figure 4.
The weight of each hermit crab compared with the amount of food remaining, grouped by each type of shell treatment. The best-fit line for each shell type was as follows: N. lamellosa: y = −0.008x + 0.1806, R2 = 0.024; L. dirum: y = −0.0308x + 0.2026, R2 = 0.22; and C. ligatum: y = −0.014x + 0.1909, R2 = 0.078. n = 16 for each type of shell.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Various parameters of the shells used in each experimental treatment. n = 16 for each species. 1SE = standard error.
Table 1.
Various parameters of the shells used in each experimental treatment. n = 16 for each species. 1SE = standard error.
| Species of Shell | Shell Length | Aperture Length | Shell Weight | |||
|---|---|---|---|---|---|---|
| Average | 1SE | Average | 1SE | Average | 1SE | |
| Calliostoma ligatum | 18.57 | 1.28 | 9.30 | 0.34 | 2.13 | 0.21 |
| Nucella lamellosa | 33.62 | 0.78 | 9.93 | 0.30 | 5.42 | 0.35 |
| Lirabuccinum dirum | 29.33 | 1.15 | 10.01 | 0.41 | 1.97 | 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Torjman, B.Z.; Iyengar, E.V. Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus. Hydrobiology 2024, 3, 74-84. https://doi.org/10.3390/hydrobiology3020006
AMA Style
Torjman BZ, Iyengar EV. Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus. Hydrobiology. 2024; 3(2):74-84. https://doi.org/10.3390/hydrobiology3020006
Chicago/Turabian StyleTorjman, Brooke Z., and Erika V. Iyengar. 2024. "Hefting Heavy Shells: Sustenance Demands Caused by Various Abodes of the Hermit Crab Pagurus granosimanus" Hydrobiology 3, no. 2: 74-84. https://doi.org/10.3390/hydrobiology3020006