The Role of Vegetation on the Ecosystem Radiative Entropy Budget and Trends Along Ecological Succession

Abstract
:1. Introduction
2. Theory and Methods
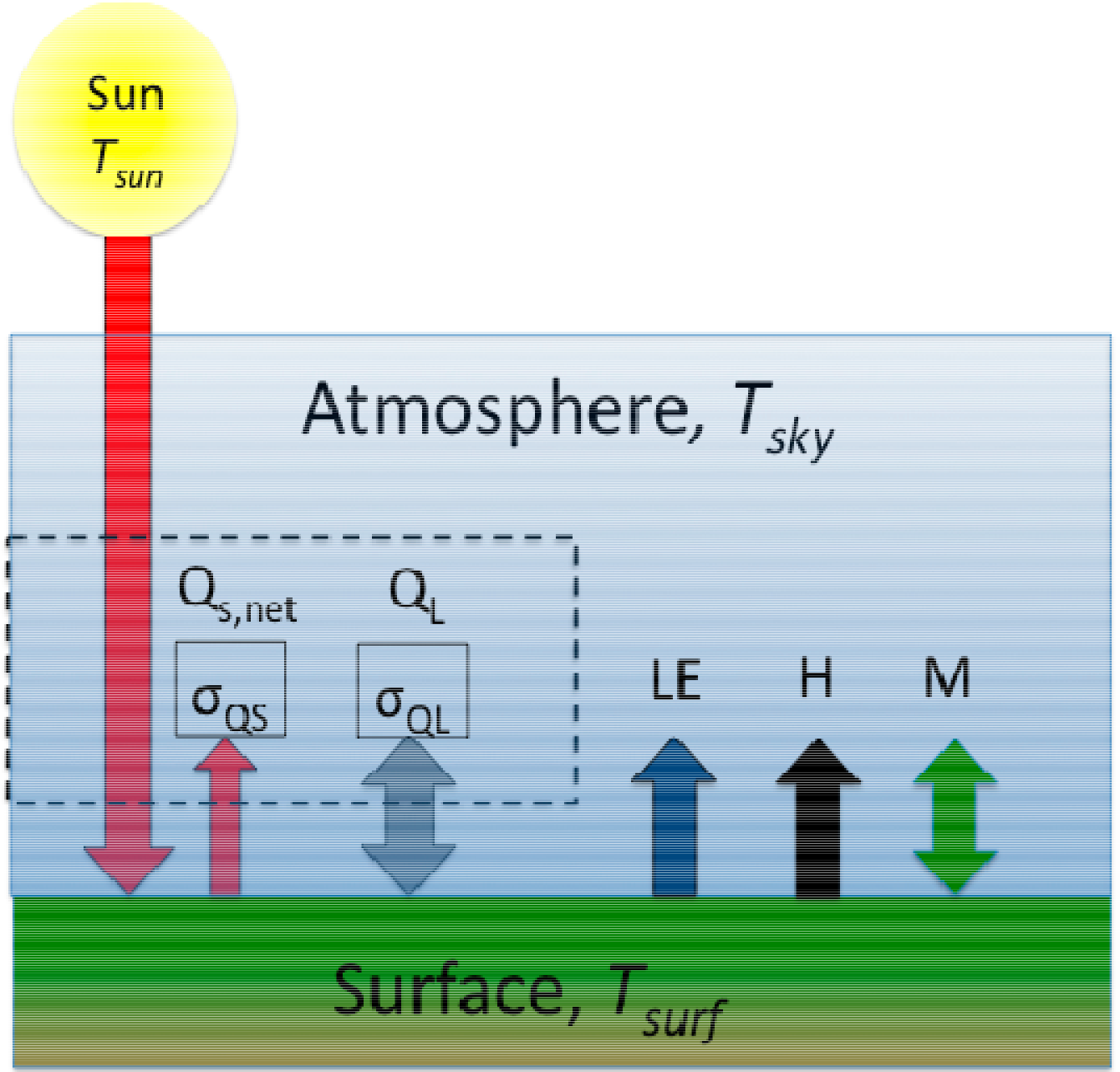
2.1. Ecosystem Energy Balance
2.2. Ecosystem Entropy Production (σ)
2.3. Empirical Maximum Entropy Production (EMEP)
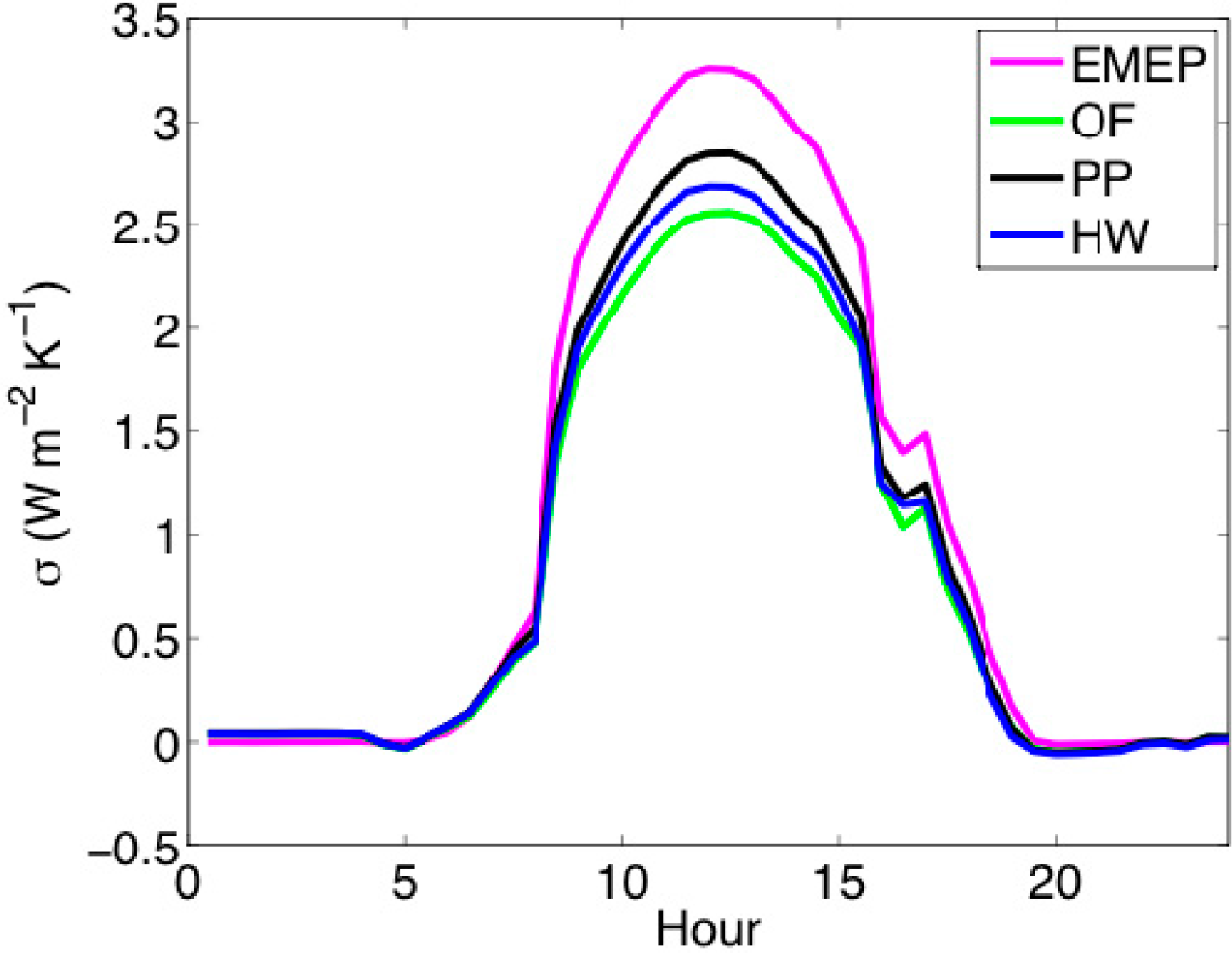
2.4. Ecosystem Observations: Duke Forest
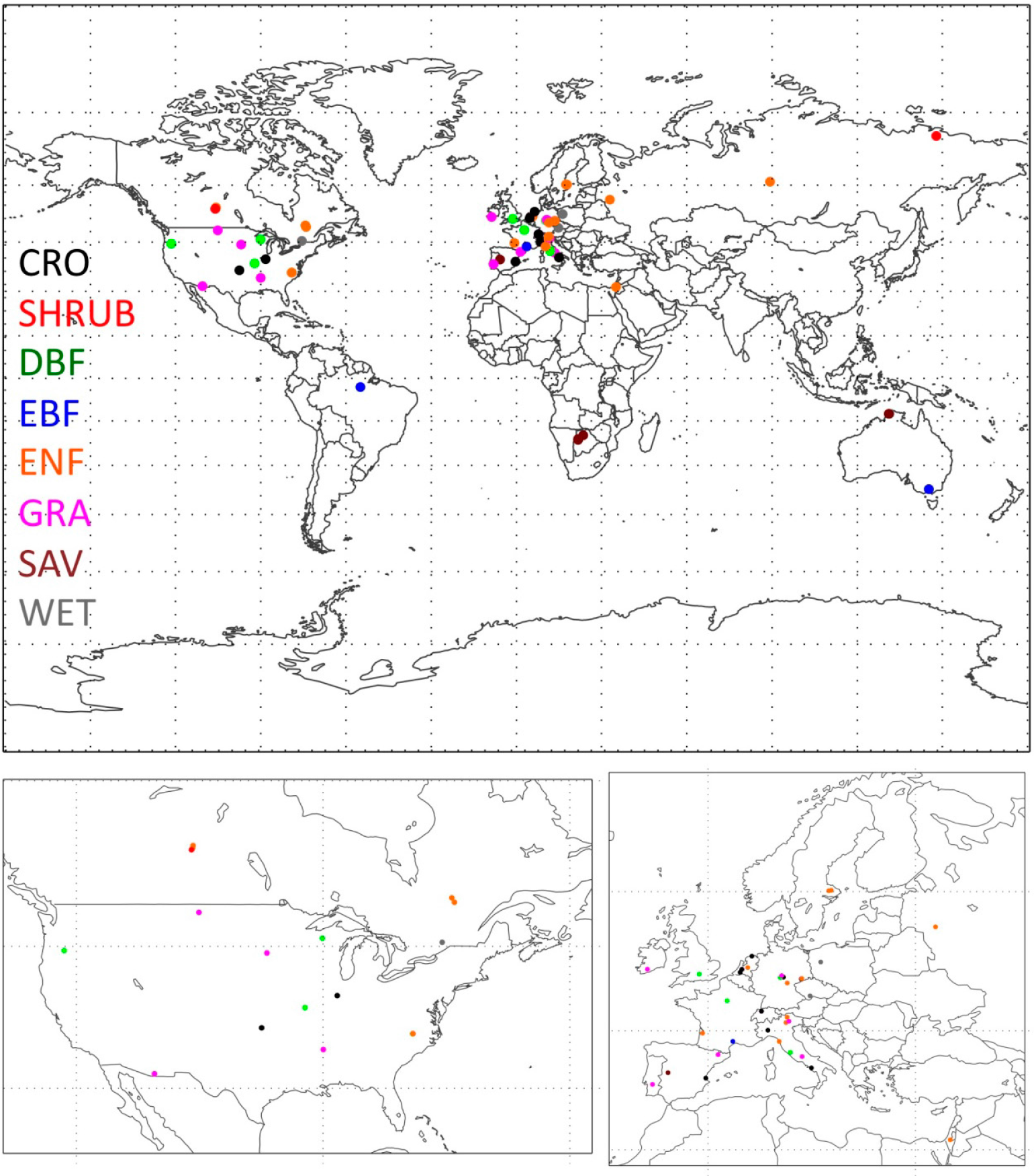
2.5. Ecosystem Observations: FLUXNET
2.6. Data Quality and Gapfilling
2.7. Statistical Analyses
3. Results
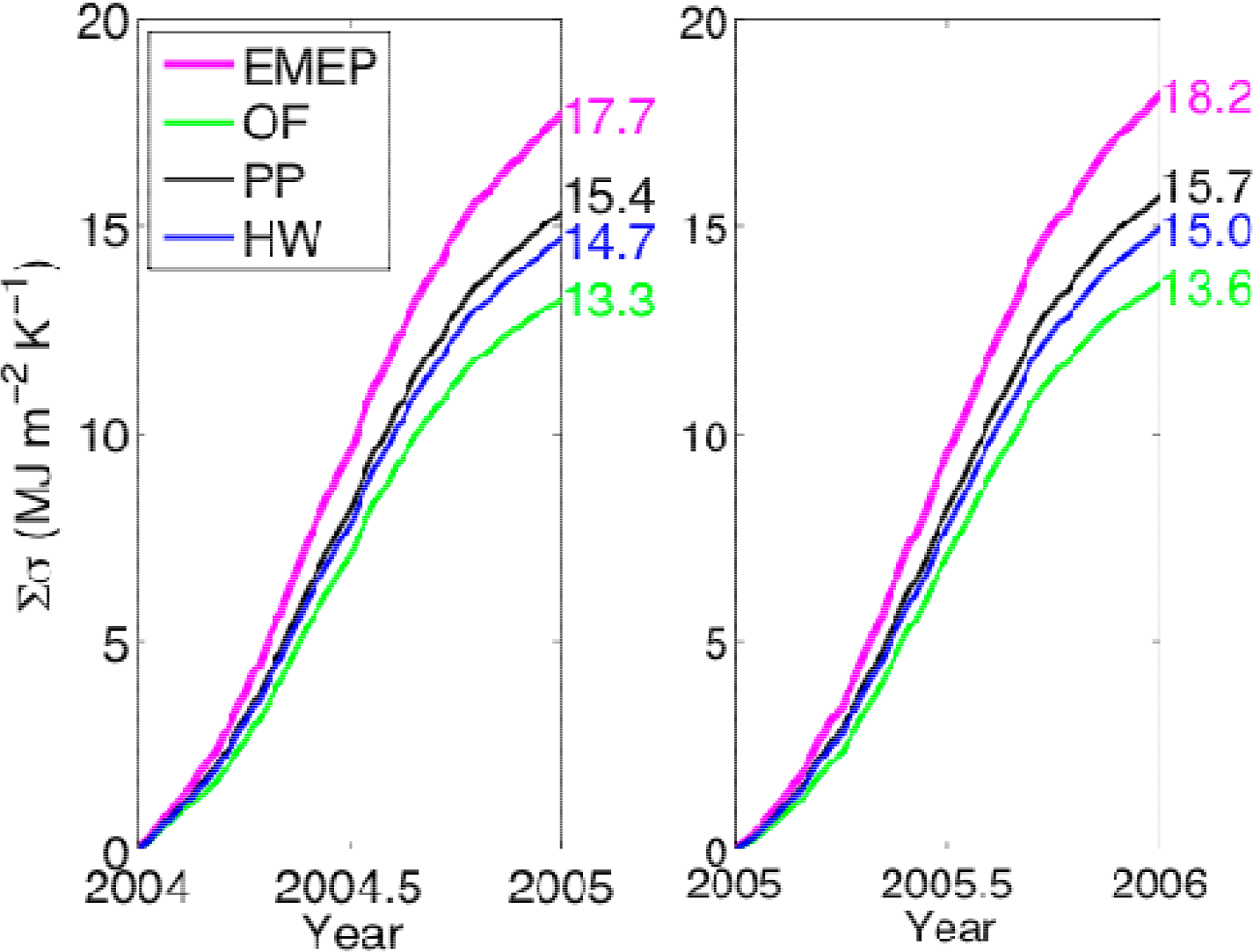
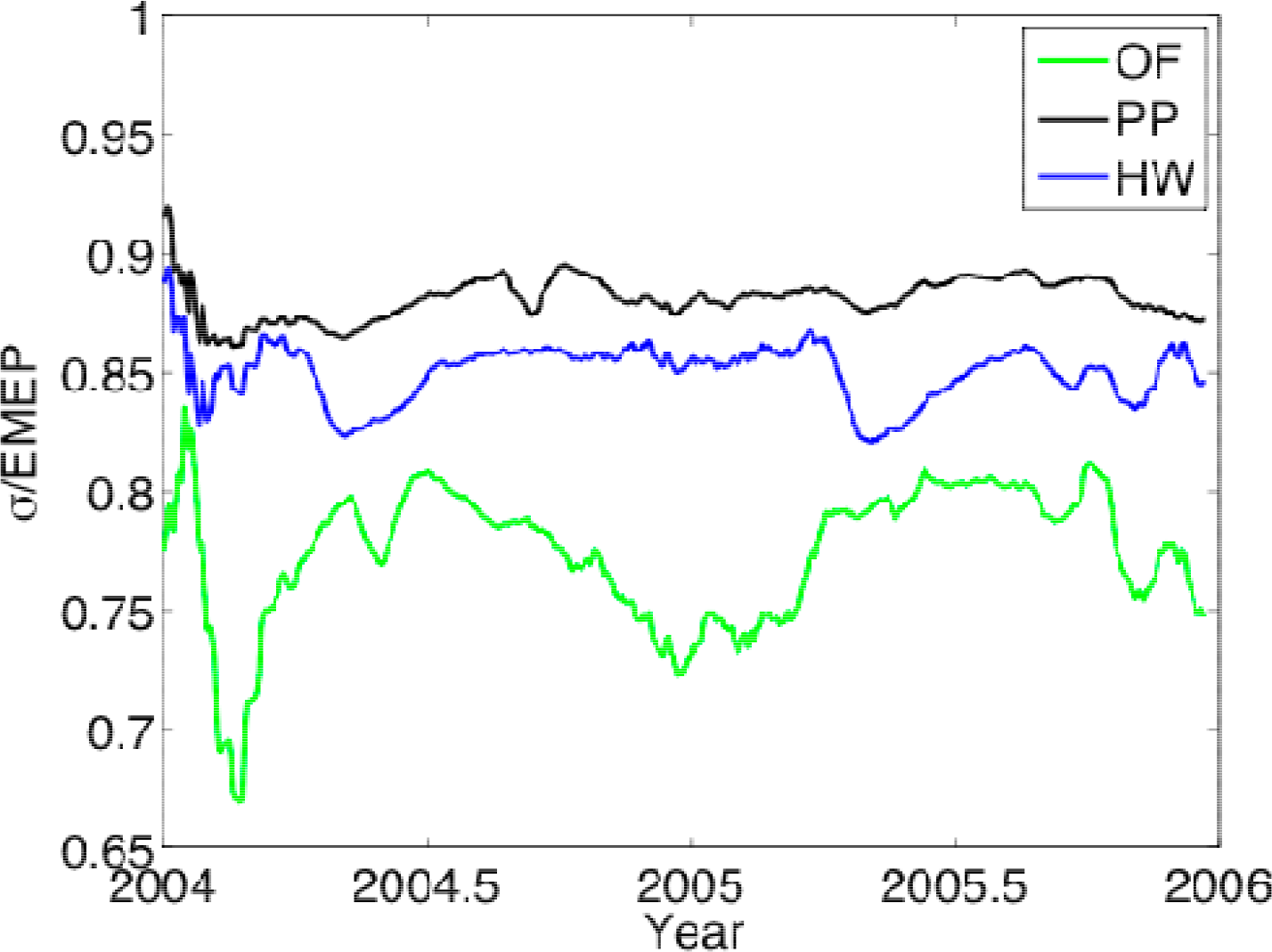
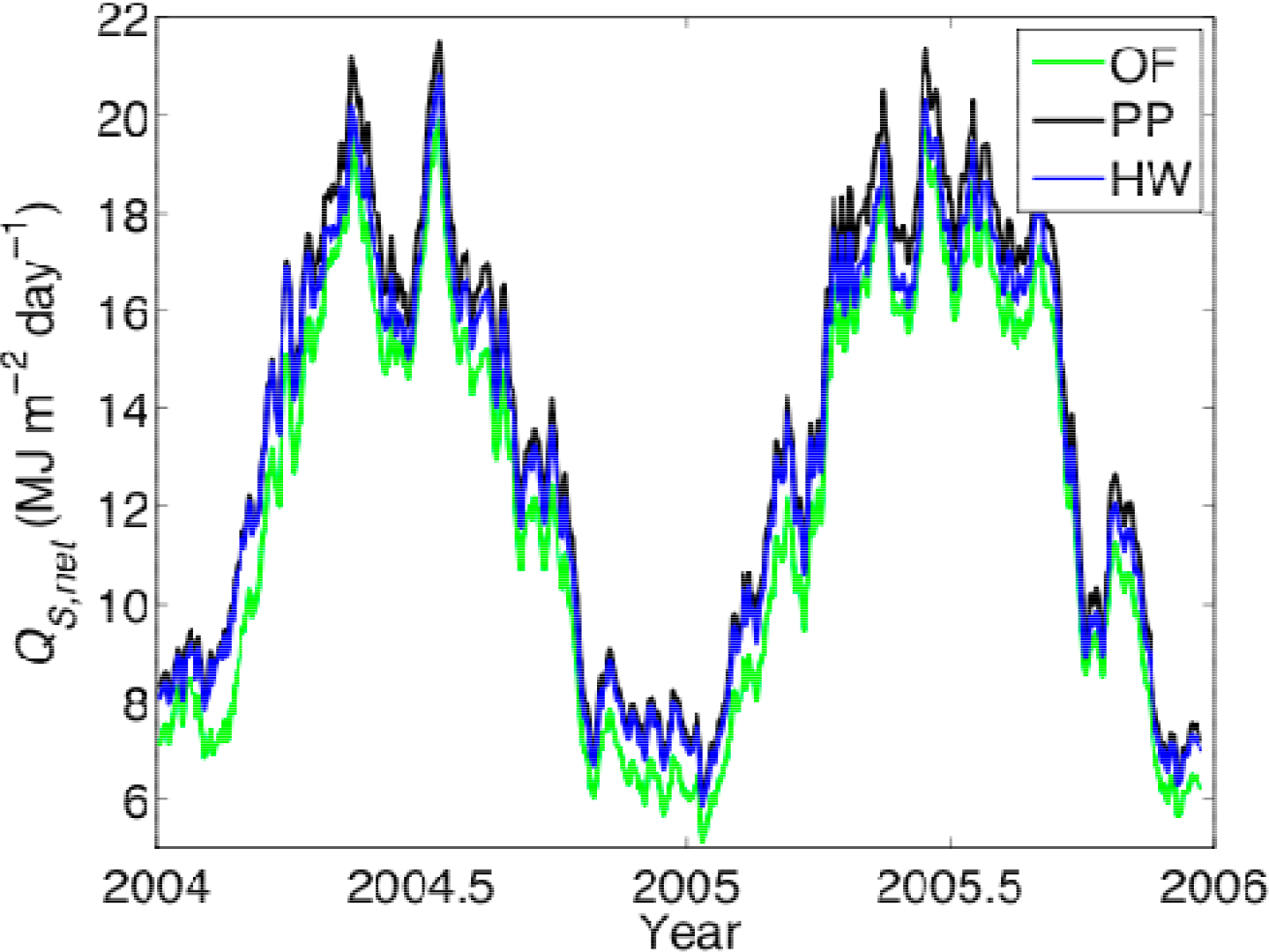
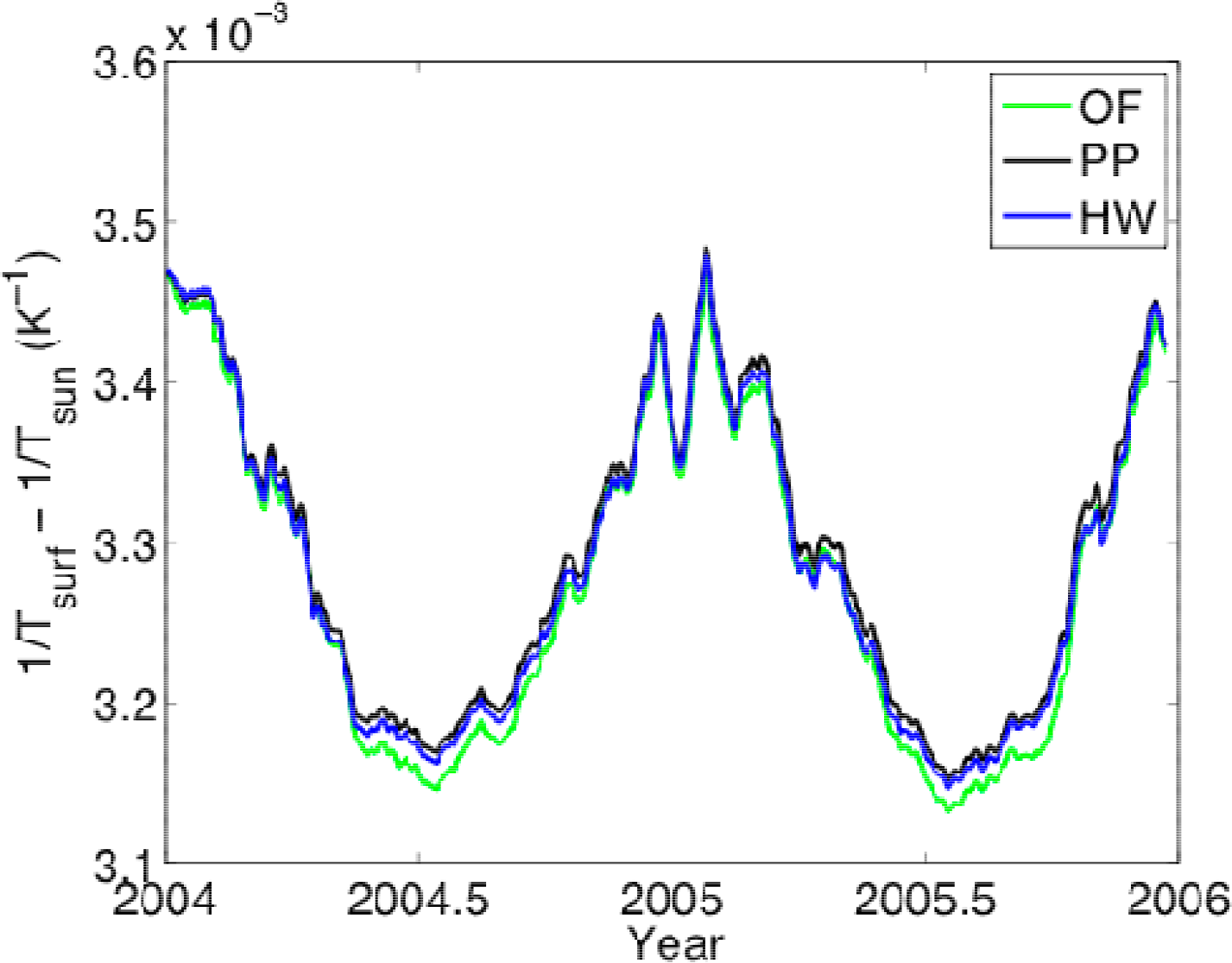
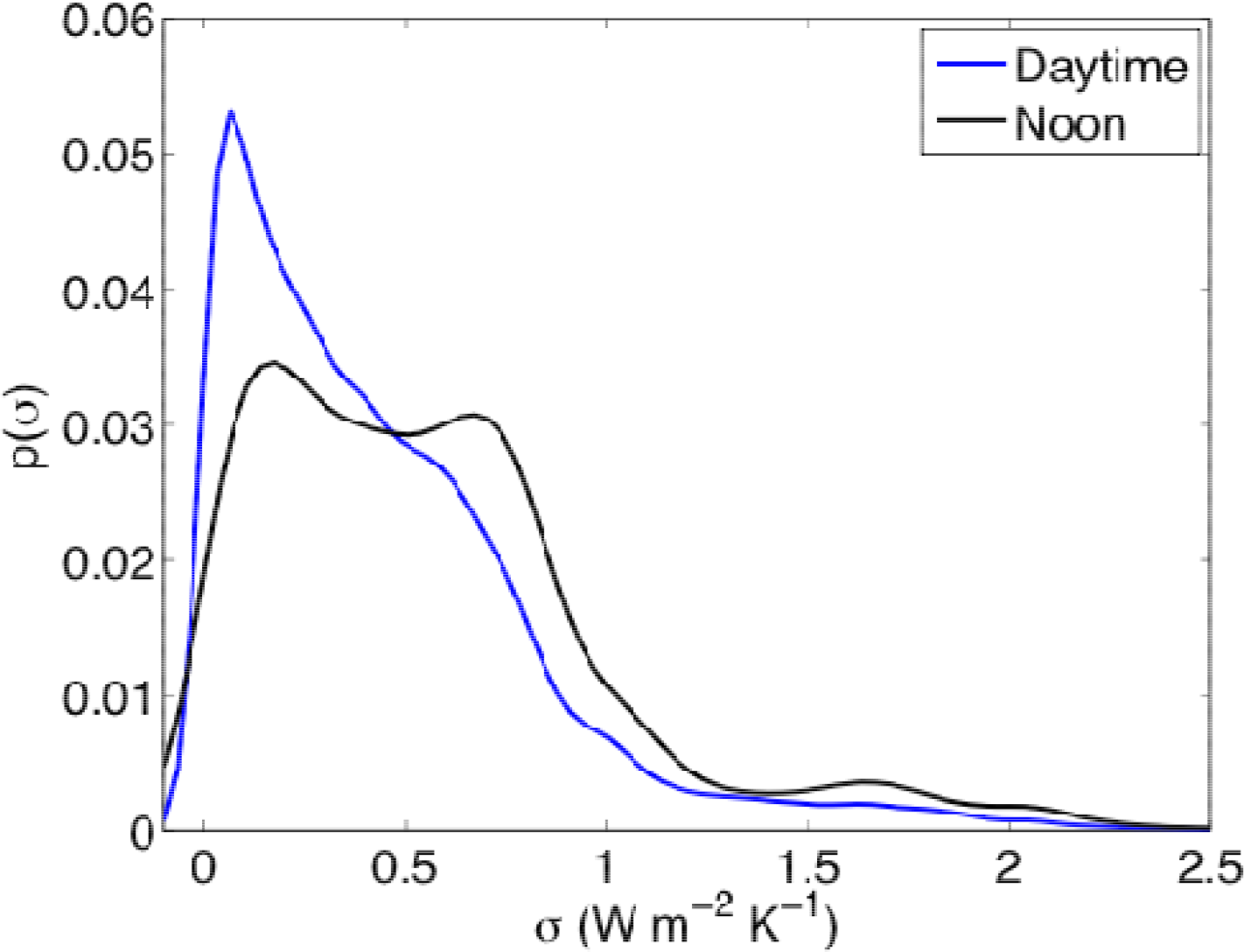
3.1. Entropy Production along Ecological Succession: Duke Forest
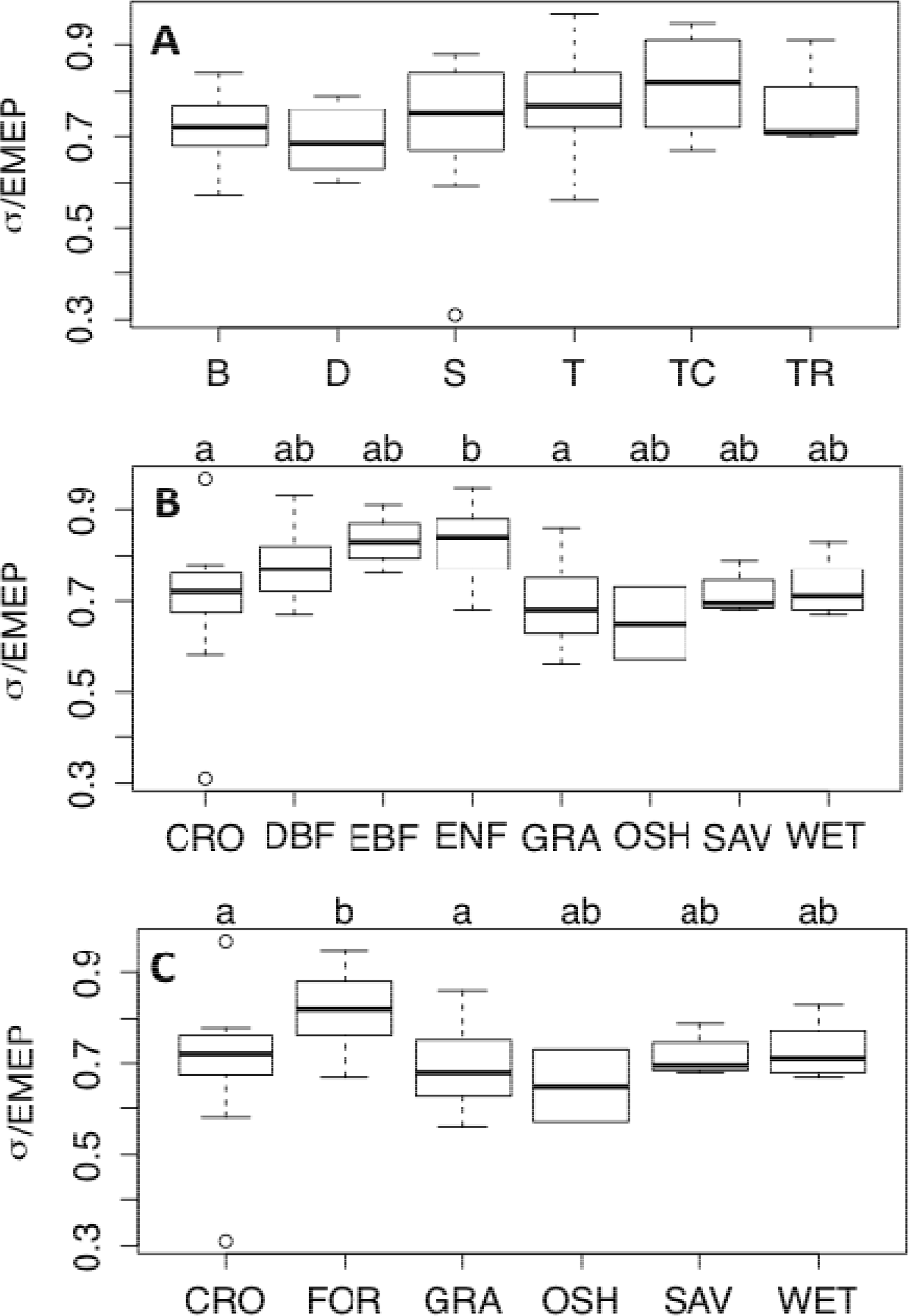
3.2. Entropy Production as a Function of Climate and Vegetation Type: FLUXNET
4. Discussion
5. Conclusions
- Ecosystem energy gain via (lower) shortwave albedo is the most relevant component for forcing ecosystem entropy production closer to its empirical maximum value;
- Entropy production was higher at a pine plantation in the Duke Forest, representative of an intermediate successional stage, than a grass field, representing early succession, and hardwood vegetation, meant to approximate a later successional stage. These results lend support to the notion that ecosystem entropy production may increase then decrease along succession [7], but FLUXNET observations suggest that older-successional ecosystems often have the highest entropy production with respect to an estimated maximum, lending support to the model of Skene [5].
- Further results from the FLUXNET analysis suggest that the relationship between succession and entropy production depends on vegetation characteristics, and late successional ecosystems frequently exhibited high values of σ/EMEP.
- Empirical and modeling studies of ecosystem entropy production may provide relevant insights on how ecosystems might maintain a cool surface as a buffer against the impacts of climate change, and how human management may improve or impede the important ecosystem service of microclimate regulation.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schrödinger, E. What Is Life? With Mind and Matter and Autobiographical Sketches; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Schneider, E.D.; Kay, J.J. Life as a manifestation of the second law of thermodynamics. Math. Comput. Model 1994, 19, 25–48. [Google Scholar]
- Lotka, A.J. Contribution to the energetics of evolution. Proc. Natl. Acad. Sci. USA 1922, 8, 147–151. [Google Scholar]
- Prigogine, I. Introduction to Thermodynamics of Irreversible Processes, 2nd ed; Interscience Publishers: New York, NY, USA, 1961; p. 119. [Google Scholar]
- Skene, K.R. The energetics of ecological succession: A logistic model of entropic output. Ecol. Model 2013, 250, 287–293. [Google Scholar]
- Kleidon, A.; Malhi, Y.; Cox, P.M. Maximum entropy production in environmental and ecological systems. Philos. Trans. R. Soc. B Biol. Sci 2010, 365, 1297–1302. [Google Scholar]
- Holdaway, R.J.; Sparrow, A.D.; Coomes, D.A. Trends in entropy production during ecosystem development in the Amazon Basin. Philos. Trans. R. Soc. B Biol. Sci 2010, 365, 1437–1447. [Google Scholar]
- Celeste, L.; Pignatti, S. Analysis of the chorological diversity in some South-European vegetational series. Annali di Bot 1988, 46, 25–34. [Google Scholar]
- Lin, H.; Cao, M.; Zhang, Y. Self-organization of tropical seasonal rain forest in southwest China. Ecol. Model 2011, 222, 2812–2816. [Google Scholar]
- Ludovisi, A. Biotic and abiotic entropy production in lake ecosystems. Ecol. Model 2004, 179, 145–147. [Google Scholar]
- Ludovisi, A.; Poletti, A. Use of thermodynamic indices as ecological indicators of the development state of lake ecosystems. 1. Entropy production indices. Ecol. Model 2003, 159, 203–222. [Google Scholar]
- Meysman, F.J.R.; Bruers, S. A thermodynamic perspective on food webs: Quantifying entropy production within detrital-based ecosystems. J. Theor. Biol 2007, 249, 124–139. [Google Scholar]
- Aoki, I. Entropy production in living systems: From organisms to ecosystems. Thermochim. Acta 1995, 250, 359–370. [Google Scholar]
- Aoki, I. Holological study of lakes from an entropy viewpoint—Lake Mendota. Ecol. Model 1989, 45, 81–93. [Google Scholar]
- Unrean, P.; Srienc, F. Metabolic networks evolve towards states of maximum entropy production. Metab. Eng 2011, 13, 666–673. [Google Scholar]
- Jaynes, E.T. Information theory and statistical mechanics. Phys. Rev 1957, 106, 620–630. [Google Scholar]
- Ulanowicz, R.E. An hypothesis on the development of natural communities. J. Theor. Biol 1980, 85, 223–245. [Google Scholar]
- Ulanowicz, R.E.; Hannon, B.M. Life and the production of entropy. Proc. R. Soc. London Ser. B 1987, 232, 181–192. [Google Scholar]
- Jørgensen, S.E.; Marques, J.C.; Müller, F.; Nielsen, S.N.; Patten, P.C.; Tiezzi, E.; Ulanowicz, R.E. A New Ecology: Systems Perspective; Elsevier: New York, NY, USA, 2007; p. 275. [Google Scholar]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar]
- Brunsell, N.A.; Schymanski, S.J.; Kleidon, A. Quantifying the thermodynamic entropy budget of the land surface: Is this useful? Earth Syst. Dyn 2011, 2, 87–103. [Google Scholar]
- Juang, J.-Y.; Katul, G.G.; Siqueira, M.B.S.; Stoy, P.C.; Novick, K.A. Separating the effects of albedo from eco-physiological changes on surface temperature along a successional chronosequence in the southeastern US. Geophys. Res. Lett 2007, 34, L21408. [Google Scholar]
- Chen, Y.; Sun-Mack, S. Surface Spectral Emissivity Derived from MODIS Data. Proceedings of the SPIE 3rd International Asia-Pacific Environmental Remote Sensing Symposium 2002: Remote Sensing of the Atmosphere, Ocean, Environment, and Space, Hangzhou, China, 23–27 October 2002.
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics, 2nd ed; Springer: New York, NY, USA, 1998. [Google Scholar]
- KleIdon, A. A model of surface heat fluxes based on the theory of maximum entropy production. Soil. Water Res 2008, 3, S89–S94. [Google Scholar]
- Stoy, P.C.; Mauder, M.; Foken, T.; Marcolla, B.; Boegh, E.; Ibrom, A.; Arain, M.A.; Arneth, A.; Aurela, M.; Bernhofer, C.; et al. A data-driven analysis of energy balance closure across FLUXNET research sites: The role of landscape scale heterogeneity. Agric. For. Meteorol 2013, 171, 137–152. [Google Scholar]
- Foken, T. The energy balance closure problem: An overview. Ecol. Appl 2008, 18, 1351–1367. [Google Scholar]
- Leuning, R.; van Gorsel, E.; Massman, W.J.; Isaac, P.R. Reflections on the surface energy imbalance problem. Agric. For. Meteorol 2012, 156, 65–74. [Google Scholar]
- Hollinger, D.Y.; Ollinger, S.V.; Richardson, A.D.; Meyers, T.; Dail, D.B.; Martin, M.E.; Scott, N.A.; Arkebauer, T.J.; Baldocchi, D.D.; Clark, K.L.; et al. Albedo estimates for land surface models and support for a new paradigm based on foliage nitrogen concentration. Glob. Chang. Biol 2009, 16, 696–710. [Google Scholar]
- Stoy, P.C.; Katul, G.G.; Siqueira, M.B.S.; Juang, J.-Y.; McCarthy, H.R.; Oishi, A.C.; Uebelherr, J.M.; Kim, H.-S.; Oren, R. Separating the effects of climate and vegetation on evapotranspiration along a successional chronosequence in the southeastern U.S. Glob. Chang. Biol 2006, 12, 2115–2135. [Google Scholar]
- Stoy, P.C.; Katul, G.G.; Siqueira, M.B.S.; Juang, J.-Y.; Novick, K.; McCarthy, H.R.; Oishi, A.C.; Oren, R. Role of vegetation in determining carbon sequestration along ecological succession in the southeastern United States. Glob. Chang. Biol 2008, 14, 1409–1427. [Google Scholar]
- Oosting, H.J. An ecological analysis of the plant communities of Piedmont, North Carolina. Am. Midl. Nat 1942, 28, 1–126. [Google Scholar]
- Novick, K.A.; Stoy, P.C.; Katul, G.G.; Ellsworth, D.S.; Siqueira, M.B.S.; Juang, J.; Oren, R. Carbon dioxide and water vapor exchange in a warm temperate grassland. Oecologia 2004, 138, 259–274. [Google Scholar]
- Pataki, D.E.; Oren, R. Species differences in stomatal control of water loss at the canopy scale in a mature bottomland deciduous forest. Adv. Water Resour 2003, 26, 1267–1278. [Google Scholar]
- Stoy, P.C.; Katul, G.G.; Siqueira, M.B.S.; Juang, J.-Y.; Novick, K.A.; Uebelherr, J.M.; Oren, R. An evaluation of models for partitioning eddy covariance-measured net ecosystem exchange into photosynthesis and respiration. Agric. For. Meteorol 2006, 141, 2–18. [Google Scholar]
- Guerschman, J.P.; van Dijk, A.I.J.M.; Mattersdorf, G.; Beringer, J.; Hutley, L.B.; Leuning, R.; Pipunic, R.C.; Sherman, B.S. Scaling of potential evapotranspiration with MODIS data reproduces flux observations and catchment water balance observations across Australia. J. Hydrol 2009, 369, 107–119. [Google Scholar]
- Hutley, L.B.; O’Grady, A.P.; Eamus, D. Evapotranspiration from Eucalypt open-forest savanna of Northern Australia. Funct. Ecol 2000, 14, 183–194. [Google Scholar]
- Martin, D.; Beringer, J.; Hutley, L.B.; McHugh, I. Carbon cycling in a mountain ash forest: Analysis of below ground respiration. Agric. For. Meteorol 2007, 147, 58–70. [Google Scholar]
- Saleska, S.R.; Miller, S.D.; Matross, D.M.; Goulden, M.; Wofsy, S.; da Rocha, H.R.; de Camargo, P.B.; Crill, P.; Daube, B.C.; de Freitas, H.C.; et al. Carbon in Amazon forests: Unexpected seasonal fluxes and disturbance-induced losses. Science 2003, 302, 1554–1557. [Google Scholar]
- Williams, C.A.; Albertson, J.D. Soil moisture controls on canopy-scale water and carbon fluxes in an African savanna. Water Resour. Res 2004, 40, W09302. [Google Scholar]
- Veenendaal, E.M.; Kolle, O.; Lloyd, J. Seasonal variation in energy fluxes and carbon dioxide exchange for a broad-leaved semi-arid savanna (Mopane woodland) in Southern Africa. Glob. Chang. Biol 2004, 10, 318–328. [Google Scholar]
- Lafleur, P.M.; Roulet, N.T.; Bubier, J.L.; Frolking, S.; Moore, T.R. Interannual variability in the peatland-atmosphere carbon dioxide exchange at an ombrotrophic bog. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Giasson, M.-A.; Coursolle, C.; Margolis, H.A. Ecosystem-level CO2 fluxes from a boreal cutover in eastern Canada before and after scarification. Agric. For. Meteorol 2006, 140, 23–40. [Google Scholar]
- Bergeron, O.; Margolis, H.A.; Black, T.A.; Coursolle, C.; Dunn, A.L.; Barr, A.G.; Wofsy, S.C. Comparison of carbon dioxide fluxes over three boreal black spruce forests in Canada. Glob. Chang. Biol 2007, 13, 89–107. [Google Scholar]
- Amiro, B.D.; Orchansky, A.L.; Barr, A.G.; Black, T.A.; Chambers, S.D.; Chapin, F.S., III; Goulden, M.L.; Litvak, M.; Liu, H.P.; McCaughey, J.H. The effect of post-fire stand age on the boreal forest energy balance. Agric. For. Meteorol 2006, 140, 41–50. [Google Scholar]
- Amiro, B.D. Paired-tower measurements of carbon and energy fluxes following disturbance in the boreal forest. Glob. Chang. Biol 2001, 7, 253–268. [Google Scholar]
- Rayment, M.B.; Jarvis, P.G. Seasonal gas exchange of black spruce using an automatic branch bag system. Can. J. For. Res 1999, 29, 1528–1538. [Google Scholar]
- Ammann, C.; Flechard, C.R.; Leifeld, J.; Neftel, A.; Fuhrer, J. The carbon budget of newly established temperate grassland depends on management intensity. Agric. Ecosyst. Environ 2007, 121, 5–20. [Google Scholar]
- Ammann, C.; Spirig, C.; Leifeld, J.; Neftel, A. Assessment of the nitrogen and carbon budget of two managed temperate grassland fields. Agric. Ecosyst. Environ 2009, 133, 150–162. [Google Scholar]
- Broquet, G.; Chevallier, F.; Bréon, F.-M.; Kadygrov, N.; Alemanno, M.; Apadula, F.; Hammer, S.; Haszpra, L.; Meinhardt, F.; Morguí, J.A. Regional inversion of CO2 ecosystem fluxes from atmospheric measurements: Reliability of the uncertainty estimates. Atmos. Chem. Phys 2013, 13, 9039–9056. [Google Scholar]
- Anthoni, P.M.; Freibauer, A.; Kolle, O.; Schulze, E.-D. Winter wheat carbon exchange in Thuringia, Germany. Agric. For. Meteorol 2004, 121, 55–67. [Google Scholar]
- Owen, K.E.; Tenhunen, J.; Reichstein, M.; Wang, Q.; Falge, E.; Geyer, R.; Xiao, X.; Stoy, P.C.; Amman, C.; Arain, A.; et al. Linking flux network measurements to continental scale simulations: Ecosystem carbon dioxide exchange capacity under non-water-stressed conditions. Glob. Chang. Biol 2007, 13, 734–760. [Google Scholar]
- Knohl, A.; Schulze, E.-D.; Kolle, O.; Buchmann, N. Large carbon uptake by an unmanaged 250-year-old deciduous forest in Central Germany. Agric. For. Meteorol 2003, 118, 151–167. [Google Scholar]
- Don, A.; Rebmann, C.; Kolle, O.; Scherer-Lorenzen, M.; Schulze, E.-D. Impact of afforestation-associated management changes on the carbon balance of grassland. Glob. Chang. Biol 2009, 15, 1990–2002. [Google Scholar]
- Bernhofer, C.; Aubinet, M.; Clement, R.; Grelle, A.; Grünwald, T.; Ibrom, A.; Jarvis, P.; Rebmann, C.; Schulze, E.-D.; Tenhunen, J.D. Spruce forests (Norway and Sitka spruce, including Douglas fir): Carbon and water fluxes and balances, ecological and ecophysiological determinants. In Fluxes of Carbon, Water and Energy of European Forests; Springer: New York, NY, USA, 2003; pp. 99–123. [Google Scholar]
- Anthoni, P.M.; Knohl, A.; Rebmann, C.; Freibauer, A.; Mund, M.; Ziegler, W.; Kolle, O.; Schulze, E.-D. Forest and agricultural land-use-dependent CO2 exchange in Thuringia, Germany. Glob. Chang. Biol 2004, 10, 2005–2019. [Google Scholar]
- Kutsch, W.L.; Aubinet, M.; Buchmann, N.; Smith, P.; Osborne, B.; Eugster, W.; Wattenbach, M.; Schrumpf, M.; Schulze, E.D.; Tomelleri, E.; et al. The net biome production of full crop rotations in Europe. Agric. Ecosyst. Environ 2010, 139, 336–345. [Google Scholar]
- Casals, P.; Gimeno, C.; Carrara, A.; Lopez-Sangil, L.; Sanz, M. Soil CO2 efflux and extractable organic carbon fractions under simulated precipitation events in a Mediterranean Dehesa. Soil. Biol. Biochem 2009, 41, 1915–1922. [Google Scholar]
- Gilmanov, T.G.; Soussana, J.F.; Aires, L.; Allard, V.; Ammann, C.; Balzarolo, M.; Barcza, Z.; Bernhofer, C.; Campbell, C.L.; Cernusca, A. Partitioning European grassland net ecosystem CO2 exchange into gross primary productivity and ecosystem respiration using light response function analysis. Agric. Ecosyst. Environ 2007, 121, 93–120. [Google Scholar]
- Davi, H.; Dufrêne, E.; Francois, C.; Le Maire, G.; Loustau, D.; Bosc, A.; Rambal, S.; Granier, A.; Moors, E. Sensitivity of water and carbon fluxes to climate changes from 1960 to 2100 in European forest ecosystems. Agric. For. Meteorol 2006, 141, 35–56. [Google Scholar]
- Berbigier, P.; Bonnefond, J.-M.; Mellmann, P. CO2 and water vapour fluxes for 2 years above Euroflux forest site. Agric. For. Meteorol 2001, 108, 183–197. [Google Scholar]
- Rambal, S.; Joffre, R.; Ourcival, J.-M. The growth respiration component in eddy CO2 flux from a Quercus ilex Mediterranean forest. Glob. Chang. Biol 2004, 10, 1460–1469. [Google Scholar]
- Peichl, M.; Leahy, P.; Kiely, G. Six-year stable annual uptake of carbon dioxide in intensively managed humid temperate grassland. Ecosystems 2010, 14, 112–126. [Google Scholar]
- Grünzweig, J.M.; Lin, T.; Rotenberg, E.; Schwartz, A.; Yakir, D. Carbon sequestration in arid-land forest. Glob. Chang. Biol 2003, 9, 791–799. [Google Scholar]
- Wohlfahrt, G.; Anderson-Dunn, M.; Bahn, M.; Balzarolo, M.; Berninger, F.; Campbell, C.; Carrara, A.; Cescatti, A.; Christensen, T.; Dore, S. Biotic, abiotic, and management controls on the net ecosystem CO2 exchange of European mountain grassland ecosystems. Ecosystems 2008, 11, 1338–1351. [Google Scholar]
- Reichstein, M.; Rey, A.; Freibauer, A.; Tenhunen, J.; Valentini, R.; Banza, J.; Casals, P.; Cheng, Y.; Grünzweig, J.M.; Irvine, J.; et al. Modeling temporal and large-Scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Skiba, U.; Drewer, J.; Tang, Y.S.; van Dijk, N.; Helfter, C.; Nemitz, E.; Famulari, D.; Cape, J.N.; Jones, S.K.; Twigg, M. Biosphere-atmosphere exchange of reactive nitrogen and greenhouse gases at the NitroEurope core flux measurement sites: Measurement strategy and first data sets. Agric. Ecosyst. Environ 2009, 133, 139–149. [Google Scholar]
- Cescatti, A.; Marcolla, B. Drag coefficient and turbulence intensity in conifer canopies. Agric. For. Meteorol 2004, 121, 197–206. [Google Scholar]
- Marcolla, B.; Cescatti, A.; Manca, G.; Zorer, R.; Cavagna, M.; Fiora, A.; Gianelle, D.; Rodeghiero, M.; Sottocornola, M.; Zampedri, R. Climatic controls and ecosystem responses drive the inter-annual variability of the net ecosystem exchange of an alpine meadow. Agric. For. Meteorol 2011, 151, 1233–1243. [Google Scholar]
- Marcolla, B.; Cescatti, A.; Montagnani, L.; Manca, G.; Kerschbaumer, G.; Minerbi, S. Importance of advection in the atmospheric CO2 exchanges of an alpine forest. Agric. For. Meteorol 2005, 130, 193–206. [Google Scholar]
- Rey, A.; Pegoraro, E.; Tedeschi, V. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Glob. Chang. Biol 2002, 8, 851–866. [Google Scholar]
- Chiesi, M.; Maselli, F.; Bindi, M.; Fibbi, L.; Cherubini, P.; Arlotta, E.; Tirone, G.; Matteucci, G.; Seufert, G. Modelling carbon budget of Mediterranean forests using ground and remote sensing measurements. Agric. For. Meteorol 2005, 135, 22–34. [Google Scholar]
- Jacobs, C.M.J.; Jacobs, A.F.G.; Bosveld, F.C.; Hendriks, D.M.D.; Hensen, A.; Kroon, P.S.; Moors, E.J.; Nol, L.; Schrier-Uijl, A.; Veenendaal, E.M. Variability of annual CO2 exchange from Dutch grasslands. Biogeosciences 2007, 4, 803–816. [Google Scholar]
- Moors, E.J.; Jacobs, C.; Jans, W.; Supit, I.; Kutsch, W.L.; Bernhofer, C.; Béziat, P.; Buchmann, N.; Carrara, A.; Ceschia, E. Variability in carbon exchange of European croplands. Agric. Ecosyst. Environ 2010, 139, 325–335. [Google Scholar]
- Dolman, A.J.; Moors, E.J.; Elbers, J.A. The carbon uptake of a mid latitude pine forest growing on sandy soil. Agric. For. Meteorol 2002, 111, 157–170. [Google Scholar]
- Chojnicki, B.H.; Urbaniak, M.; Józefczyk, D.; Augustin, J.; Olejnik, J. Measurements of gas and heat fluxes at Rzecin wetland. In Wetlands: Monitoring, Modelling and Managemen; Okrus, T., Maltby, E., Szatylowicz, J., Miroslaw-Swiatek, D., Kotowski, W., Eds.; Taylor & Francis: London, UK, 2007; pp. 125–131. [Google Scholar]
- Pereira, J.S.; Mateus, J.A.; Aires, L.M.; Pita, G.; Pio, C.; David, J.S.; Andrade, V.; Banza, J.; David, T.S.; Paço, T.A. Net ecosystem carbon exchange in three contrasting Mediterranean ecosystems-the effect of drought. Biogeosciences 2007, 4, 791–802. [Google Scholar]
- Van der Molen, M.K.; van Huissteden, J.; Parmentier, F.J.W.; Petrescu, A.M.R.; Dolman, A.J.; Maximov, T.C.; Kononov, A.V; Karsanaev, S.V; Suzdalov, D.A. The growing season greenhouse gas balance of a continental tundra site in the Indigirka lowlands, NE Siberia. Biogeosciences 2007, 4, 985–1003. [Google Scholar]
- Kurbatova, J.; Li, C.; Varlagin, A.; Xiao, X.; Vygodskaya, N. Modeling carbon dynamics in two adjacent spruce forests with different soil conditions in Russia. Biogeosciences 2008, 5, 969–980. [Google Scholar]
- Kurbatova, J.; Arneth, A.; Vygodskaya, N.N.; Kolle, O.; Varlargin, A.V; Milyukova, I.M.; Tchebakova, N.M.; Schulze, E.; Lloyd, J. Comparative ecosystem-atmosphere exchange of energy and mass in a European Russian and a central Siberian bog I. Interseasonal and interannual variability of energy and latent heat fluxes during the snowfree period. Tellus B 2002, 54, 497–513. [Google Scholar]
- Lagergren, F.; Eklundh, L.; Grelle, A.; Lundblad, M.; Mölder, M.; Lankreijer, H.; Lindroth, A. Net primary production and light use efficiency in a mixed coniferous forest in Sweden. Plant Cell Environ 2005, 28, 412–423. [Google Scholar]
- Gioli, B.; Miglietta, F.; de Martino, B.; Hutjes, R.W.A.; Dolman, A.J.; Lindroth, A.; Schumacher, M.; Sanz, M.J.; Manca, G.; Peressotti, A.; et al. Comparison between tower and aircraft-based eddy covariance fluxes in five European regions. Agric. For. Meteorol 2004, 127, 1–16. [Google Scholar]
- Herbst, M.; Rosier, P.T.W.; McNeil, D.D.; Harding, R.J.; Gowing, D.J. Seasonal variability of interception evaporation from the canopy of a mixed deciduous forest. Agric. For. Meteorol 2008, 148, 1655–1667. [Google Scholar]
- Fischer, M.L.; Billesbach, D.P.; Berry, J.A.; Riley, W.J.; Torn, M.S. Spatiotemporal variations in growing season exchanges of CO2, H2O, and sensible heat in agricultural fields of the southern Great Plains. Earth Interact 2007, 11, 1–21. [Google Scholar]
- Saito, M.; Maksyutov, S.; Hirata, R.; Richardson, A.D. An empirical model simulating diurnal and seasonal CO 2 flux for diverse vegetation types and climate conditions. Biogeosciences 2009, 6, 585–599. [Google Scholar]
- Gilmanov, T.G.; Aires, L.; Barcza, Z.; Baron, V.S.; Belelli, L.; Beringer, J.; Billesbach, D.; Bonal, D.; Bradford, J.; Ceschia, E. Productivity, respiration, and light-response parameters of world grassland and agroecosystems derived from flux-tower measurements. Rangel. Ecol. Manag 2010, 63, 16–39. [Google Scholar]
- Meyers, T.P.; Hollinger, S.E. An assessment of storage terms in the surface energy balance of maize and soybean. Agric. For. Meteorol 2004, 125, 105–115. [Google Scholar]
- Oren, R.; Ellsworth, D.E.; Johnsen, K.H.; Phillips, N.; Ewers, B.E.; Maier, C.; Schäfer, K.V.R.; McCarthy, H.; Hendrey, G.; et al. Soil fertility limits carbon sequestration by forest-ecosystems in CO2 enriched atmosphere. Nature 2001, 411, 469–472. [Google Scholar]
- Gilmanov, T.G.; Tieszen, L.L.; Wylie, B.K.; Flanagan, L.B.; Frank, A.B.; Haferkamp, M.R.; Meyers, T.P.; Morgan, J.A. Integration of CO2 flux and remotely-sensed data for primary production and ecosystem respiration analyses in the Northern Great Plains: Potential for quantitative spatial extrapolation. Glob. Ecol. Biogeogr 2005, 14, 271–292. [Google Scholar]
- Wilson, T.B.; Meyers, T.P. Determining vegetation indices from solar and photosynthetically active radiation fluxes. Agric. For. Meteorol 2007, 144, 160–179. [Google Scholar]
- Schmid, H.P.; Grimmond, C.S.B.; Cropley, F.; Offerle, B.; Su, H.B. Measurements of CO2 and energy fluxes over a mixed hardwood forest in the mid-western United States. Agric. For. Meteorol. 2000, 103, 357–374. [Google Scholar]
- Gu, L.; Meyers, T.; Pallardy, S.G.; Hanson, P.J.; Yang, B.; Heuer, M.; Hosman, K.P.; Riggs, J.S.; Sluss, D.; Wullschleger, S.D. Direct and indirect effects of atmospheric conditions and soil moisture on surface energy partitioning revealed by a prolonged drought at a temperate forest site. J. Geophys. Res. Atmos 2006, 111. [Google Scholar] [CrossRef]
- Cook, B.D.; Davis, K.J.; Wang, W.; Desai, A.; Berger, B.W.; Teclaw, R.M.; Martin, J.G.; Bolstad, P.V.; Bakwin, P.S.; Yi, C. Carbon exchange and venting anomalies in an upland deciduous forest in northern Wisconsin, USA. Agric. For. Meteorol 2004, 126, 271–295. [Google Scholar]
- Agarwal, D.A.; Humphrey, M.; Beekwilder, N.F.; Jackson, K.R.; Goode, M.M.; van Ingen, C. A data-centered collaboration portal to support global carbon-flux analysis. Concurr. Comput. Pract. Exp 2010, 22, 2323–2334. [Google Scholar]
- Juang, J.-Y.; Katul, G.G.; Porporato, A.; Stoy, P.C.; Siqueira, M.S.; Detto, M.; Kim, H.-S.; Oren, R. Eco-hydrological controls on summertime convective rainfall triggers. Glob. Chang. Biol 2007, 13, 887–896. [Google Scholar]
- Novick, K.; Brantley, S.; Miniat, C.F.; Walker, J.; Vose, J.M. Inferring the contribution of advection to total ecosystem scalar fluxes over a tall forest in complex terrain. Agric. For. Meteorol 2014, 185, 1–13. [Google Scholar]
- Kleidon, A. Nonequilibrium thermodynamics and maximum entropy production in the Earth system. Naturwissenschaften 2009, 96, 653–677. [Google Scholar]
- Kleidon, A. Energy balance. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Elsevier: Oxford, UK, 2008; Volume 2, pp. 1276–1289. [Google Scholar]
- Luyssaert, S.; Jammet, M.; Stoy, P.; Estel, S.; Pongratz, J.; Ceschia, E.; Churkina, G.; Don, A.; Erb, K.; Ferlicoq, M.; et al. Land management and land-cover change have impacts of similar magnitude on surface temperature. Nat. Clim. Chang 2014, 4, 389–393. [Google Scholar]
- Luyssaert, S.; Schulze, E.D.; Borner, A.; Knohl, A.; Hessenmoller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar]
- De Frenne, P.; Rodríguez-Sánchez, F.; Coomes, D.A.; Baeten, L.; Verstraeten, G.; Vellend, M.; Bernhardt-Römermann, M.; Brown, C.D.; Brunet, J.; Cornelis, J. Microclimate moderates plant responses to macroclimate warming. Proc. Natl. Acad. Sci. USA 2013, 110, 18561–18565. [Google Scholar]
- Norris, C.; Hobson, P.; Ibisch, P. Microclimate and vegetation function as indicators of forest thermodynamic efficiency. J. Appl. Ecol 2011, 49, 562–570. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | σ/EMEP | Veg.a | Clim.b | Year(s) | Lat. | Long. | Ref. |
|---|---|---|---|---|---|---|---|
| AUFog | 0.71 | WET | TR | 2006–2007 | −12.5425 | 131.3070 | [36] |
| AUHow | 0.70 | SAV | TR | 2001–2006 | −12.4943 | 131.1520 | [37] |
| AUWac | 0.83 | EBF | T | 2005–2007 | −37.4290 | 145.1870 | [38] |
| BRSa3 | 0.91 | EBF | TR | 2000–2003 | −3.01803 | −54.9714 | [39] |
| BWGhg | 0.69 | SAV | D | 2003 | −21.51 | 21.74 | [40] |
| BWGhm | 0.68 | SAV | D | 2003 | −21.20 | 21.75 | [40] |
| BWMa1 | 0.79 | SAV | D | 1999–2001 | −19.9155 | 23.5605 | [41] |
| CAMer | 0.67 | WET | TC | 2003–2004 | 45.4094 | −75.5186 | [42] |
| CAQcu | 0.70 | ENF | B | 2004–2006 | 49.2671 | −74.0365 | [43] |
| CAQfo | 0.84 | ENF | B | 2003–2005 | 49.6925 | −74.3421 | [44] |
| CASF1 | 0.77 | ENF | B | 2003–2005 | 54.4850 | −105.8180 | [45] |
| CASF2 | 0.68 | ENF | B | 2003–2005 | 54.2539 | −105.8780 | [46,47] |
| CASF3 | 0.57 | OSH | B | 2003–2005 | 54.0916 | −106.0050 | [46,47] |
| CHOe1 | 0.68 | GRA | T | 2003–2006 | 47.2856 | 7.7321 | [48] |
| CHOe2 | 0.58 | CRO | T | 2005 | 47.2860 | 7.7340 | [49] |
| CZwet | 0.83 | WET | T | 2006 | 49.0250 | 14.7720 | [50] |
| EGeb | 0.74 | CRO | T | 2004–2006 | 51.1001 | 10.9143 | [51] |
| DEGri | 0.84 | GRA | T | 2006 | 50.9495 | 13.5125 | [52] |
| DEHai | 0.79 | DBF | T | 2004–2006 | 51.0793 | 10.4520 | [53] |
| DEKli | 0.66 | CRO | T | 2004–2006 | 50.8929 | 13.5225 | [52] |
| DEMeh | 0.72 | GRA | T | 2003–2006 | 51.275 | 10.6555 | [54] |
| DETha | 0.93 | ENF | T | 2004–2006 | 50.9636 | 13.5669 | [55] |
| DEWet | 0.89 | ENF | T | 2002–2006 | 50.4535 | 11.4575 | [56] |
| ESES2 | 0.78 | CRO | S | 2005–2006 | 39.2755 | −0.3152 | [57] |
| ESLMa | 0.72 | ENF | B | 2005–2006 | 39.9415 | −5.7734 | [58] |
| ESVDA | 0.68 | WET | B | 2005–2006 | 42.1522 | 1.4485 | [59] |
| FRFon | 0.77 | DBF | T | 2005–2006 | 48.4763 | 2.7801 | [60] |
| FRLBr | 0.80 | ENF | T | 2003–2006 | 44.7171 | −0.7693 | [61] |
| FRPue | 0.76 | EBF | S | 2005–2006 | 43.7414 | 3.5958 | [62] |
| IEDri | 0.70 | GRA | T | 2003–2005 | 51.9867 | −8.7518 | [63] |
| ILYat | 0.76 | ENF | D | 2004–2005 | 31.3450 | 35.0515 | [64] |
| ITAmp | 0.65 | GRA | S | 2005–2006 | 41.9041 | 13.6052 | [65] |
| ITBCi | 0.71 | CRO | S | 2006 | 40.5238 | 14.9574 | [66] |
| ITCas | 0.69 | CRO | S | 2006 | 45.0628 | 8.6685 | [67] |
| ITLav | 0.85 | ENF | T | 2004, 2006 | 45.9553 | 11.2812 | [68] |
| ITMBo | 0.56 | GRA | T | 2004–2006 | 46.0156 | 11.0467 | [69] |
| ITRen | 0.85 | ENF | T | 2004–2006 | 46.5878 | 11.4347 | [70] |
| ITRo1 | 0.67 | DBF | S | 2005–2006 | 42.4081 | 11.9300 | [71] |
| ITSRo | 0.88 | ENF | S | 2004, 2006 | 43.7279 | 10.2844 | [72] |
| NLCa1 | 0.67 | GRA | T | 2003–2006 | 51.9710 | 4.9270 | [73] |
| NLLan | 0.73 | CRO | T | 2005–2006 | 51.9536 | 4.9029 | [74] |
| NLLoo | 0.82 | ENF | T | 1999–2000, 2002–2006 | 52.1679 | 5.7440 | [75] |
| NLLut | 0.72 | CRO | T | 2006 | 53.3989 | 6.3560 | [74] |
| NLMol | 0.86 | CRO | T | 2005 | 51.650 | 4.6390 | [74] |
| PLwet | 0.77 | WET | T | 2004–2005 | 52.7622 | 16.3094 | [76] |
| PTMi2 | 0.59 | GRA | S | 2004–2006 | 38.4765 | −8.0246 | [77] |
| RUCok | 0.73 | OSH | B | 2003–2005 | 70.6167 | 147.8830 | [78] |
| RUFyo | 0.91 | ENF | TC | 1998–2004 | 56.4617 | 32.9240 | [79] |
| RUZot | 0.79 | ENF | B | 2002–2004 | 60.8008 | 89.3508 | [80] |
| SENor | 0.95 | ENF | TC | 2005 | 60.0865 | 17.4795 | [81] |
| SESk2 | 0.84 | ENF | T | 2004–2005 | 60.1297 | 17.8401 | [82] |
| UKPL3 | 0.72 | DBF | T | 2005–2006 | 51.4500 | −1.2667 | [83] |
| USARM | 0.31 | CRO | S | 2003–2006 | 36.6058 | −97.4888 | [84] |
| USAud | 0.60 | GRA | D | 2002–2006 | 31.5907 | −110.51 | [85] |
| USBkg | 0.86 | GRA | TC | 2004–2006 | 44.3453 | −96.8362 | [86] |
| USBo1 | 0.78 | CRO | TC | 2001–2007 | 40.0062 | −88.2924 | [87] |
| USDk1 | 0.78 | GRA | S | 2004–2005 | 35.9712 | −79.0934 | [33] |
| USDk2 | 0.85 | DBF | S | 2004–2005 | 35.9736 | −79.1004 | [34] |
| USDk3 | 0.88 | ENF | S | 2004–2005 | 35.9782 | −79.0942 | [88] |
| USFPe | 0.63 | GRA | D | 2001–2006 | 48.3077 | −105.1019 | [89] |
| USGoo | 0.84 | GRA | S | 2002–2006 | 34.2547 | −89.8735 | [90] |
| USMMS | 0.77 | DBF | T | 2002–2004 | 39.3231 | −86.4131 | [91] |
| USMOz | 0.93 | DBF | T | 2004–2006 | 38.7441 | −92.2000 | [92] |
| USWCr | 0.72 | DBF | TC | 1999–2006 | 45.8059 | −90.0799 | [93] |
| T | TC | TR | D | B | A | S | Sum | |
|---|---|---|---|---|---|---|---|---|
| CRO | 6 | 1 | 0 | 0 | 0 | 0 | 4 | 11 |
| OSH | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 2 |
| DBF | 3 | 1 | 0 | 0 | 0 | 0 | 4 | 8 |
| EBF | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 3 |
| ENF | 7 | 2 | 0 | 1 | 5 | 0 | 2 | 17 |
| GRA | 7 | 1 | 0 | 2 | 0 | 0 | 4 | 14 |
| MF | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| SAV | 0 | 0 | 1 | 3 | 0 | 0 | 1 | 5 |
| WET | 2 | 1 | 1 | 0 | 0 | 0 | 0 | 4 |
| Sum | 26 | 6 | 3 | 6 | 7 | 0 | 16 | 64 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stoy, P.C.; Lin, H.; Novick, K.A.; Siqueira, M.B.S.; Juang, J.-Y. The Role of Vegetation on the Ecosystem Radiative Entropy Budget and Trends Along Ecological Succession. Entropy 2014, 16, 3710-3731. https://doi.org/10.3390/e16073710
Stoy PC, Lin H, Novick KA, Siqueira MBS, Juang J-Y. The Role of Vegetation on the Ecosystem Radiative Entropy Budget and Trends Along Ecological Succession. Entropy. 2014; 16(7):3710-3731. https://doi.org/10.3390/e16073710
Chicago/Turabian StyleStoy, Paul C., Hua Lin, Kimberly A. Novick, Mario B. S. Siqueira, and Jehn-Yih Juang. 2014. "The Role of Vegetation on the Ecosystem Radiative Entropy Budget and Trends Along Ecological Succession" Entropy 16, no. 7: 3710-3731. https://doi.org/10.3390/e16073710
APA StyleStoy, P. C., Lin, H., Novick, K. A., Siqueira, M. B. S., & Juang, J.-Y. (2014). The Role of Vegetation on the Ecosystem Radiative Entropy Budget and Trends Along Ecological Succession. Entropy, 16(7), 3710-3731. https://doi.org/10.3390/e16073710