
Slow Dynamics and Structure of Supercooled Water in Confinement

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
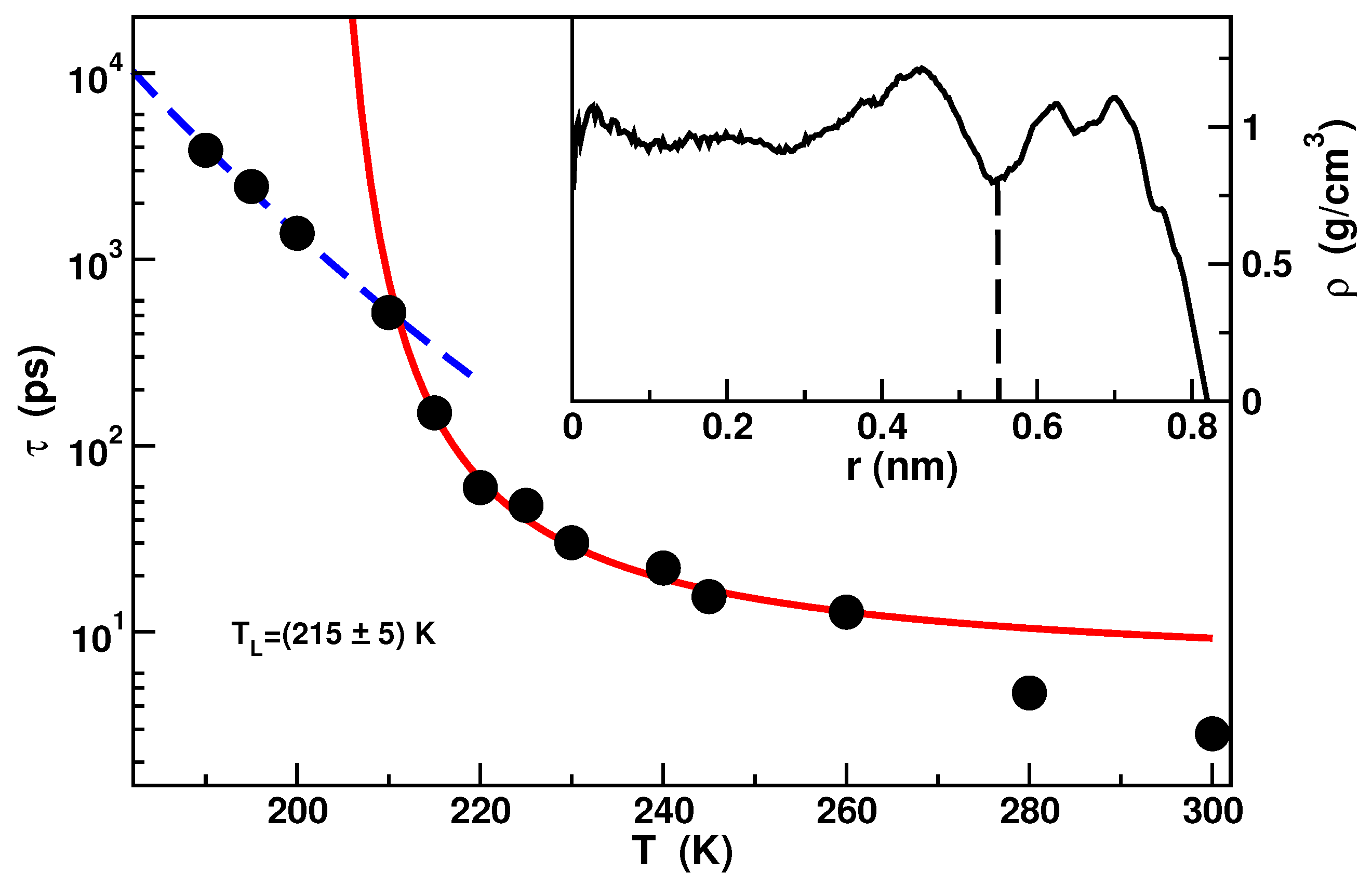
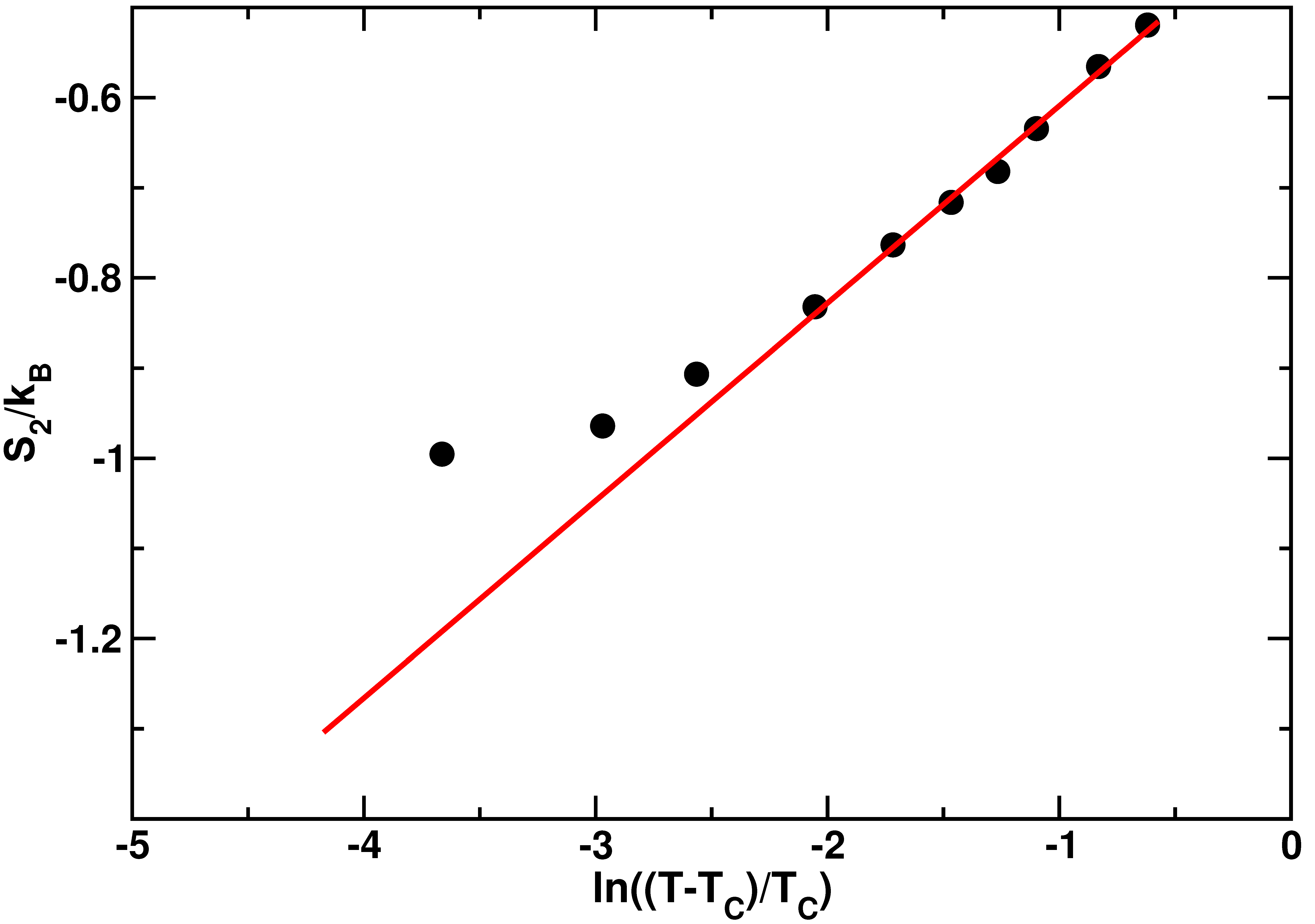
2.1. Confined Water
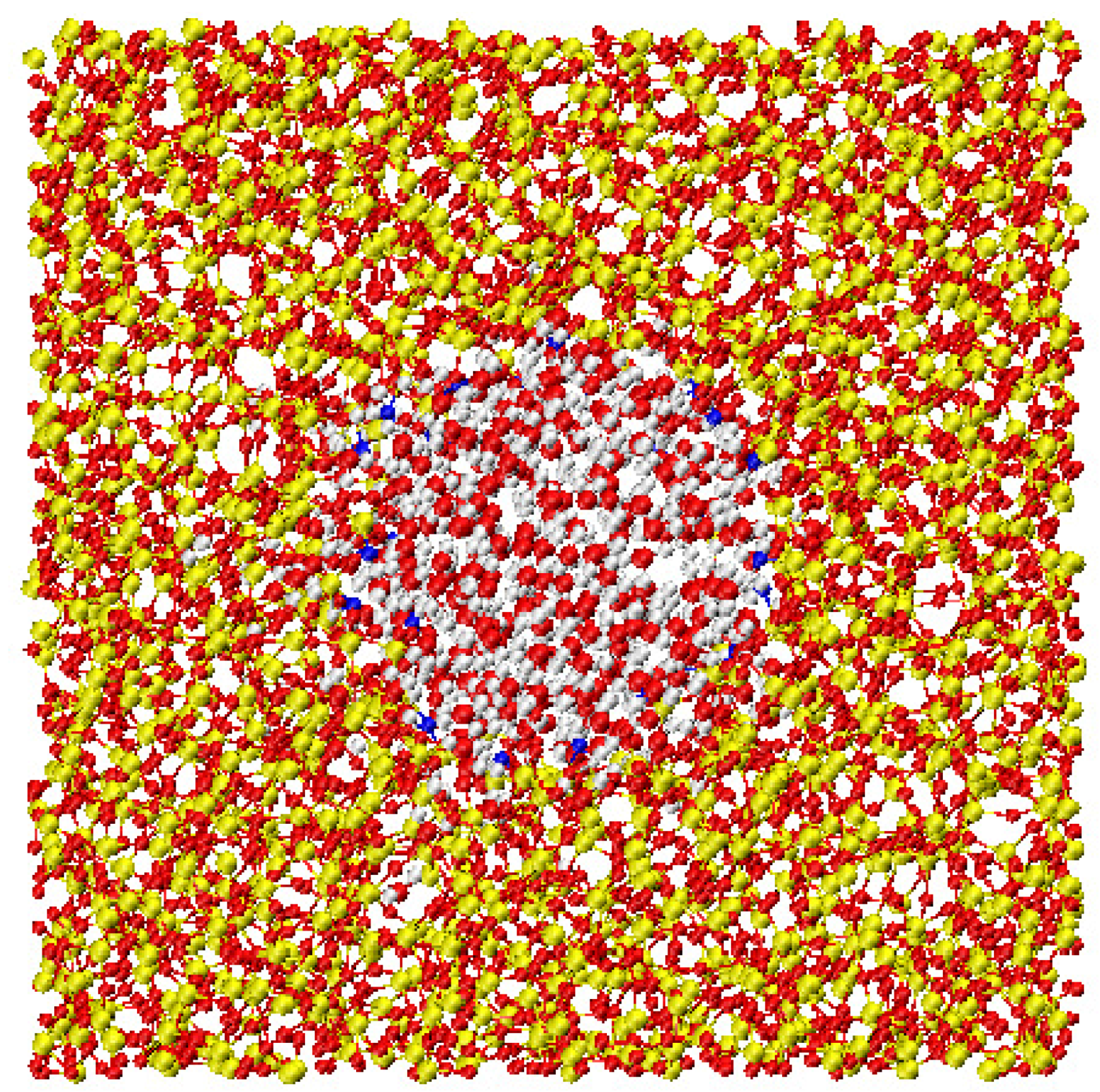
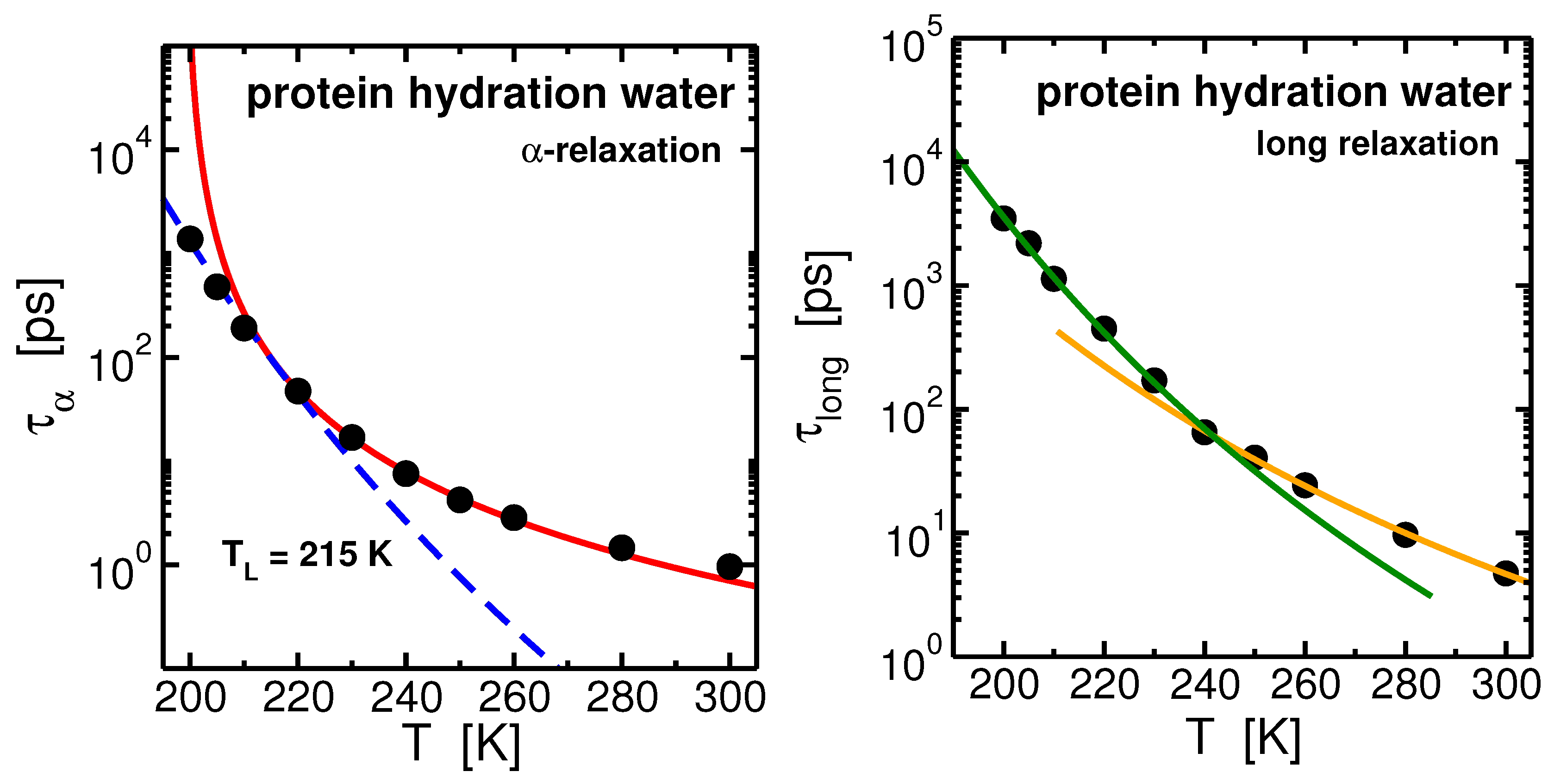
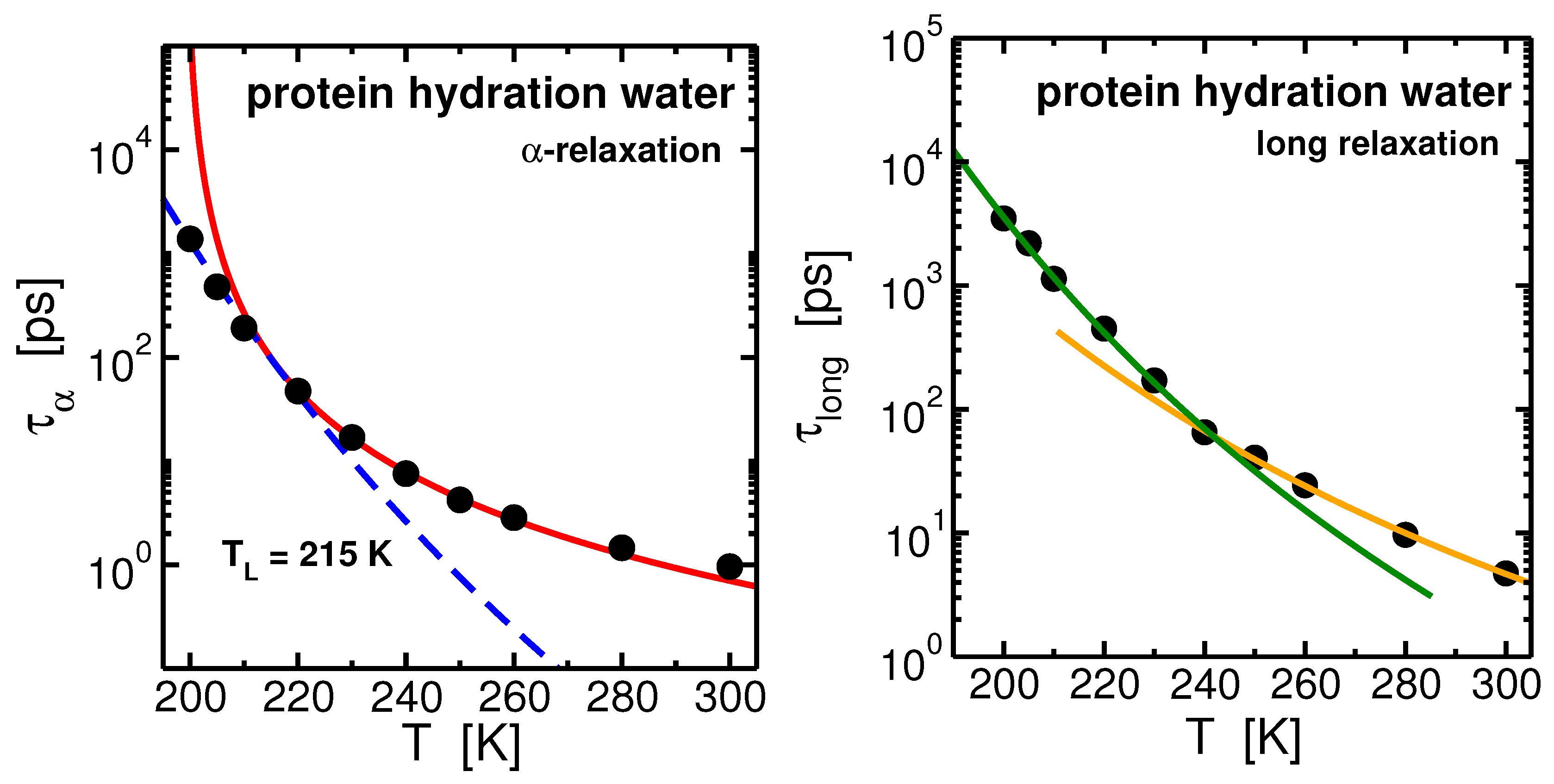
2.2. Protein Hydration Water
3. Methods
4. Conclusions
Conflicts of Interest
Abbreviations
| LLCP | Liquid-Liquid Critical Point |
| LDA | Low Density Amorphous |
| HDA | High Density Amorphous |
| LDL | Low Density Liquid |
| HDL | High Density Liquid |
| MCT | Mode Coupling Theory |
| VFT | Vogel-Fulcher-Tammann |
| FSC | Fragile to Strong Crossover |
| PDT | Protein Dynamical Transition |
| MD | Molecular Dynamics |
| SISF | Self Intermediate Scattering Function |
| RDF | Radial Distribution Functions |
References
- Franks, F. Water: A Matrix of Life, 2nd ed.; RSC Paperbacks, The Royal Society of Chemistry: Cambridge, UK, 2000. [Google Scholar]
- Ball, P. Water: Water-An enduring mystery. Nature 2008, 452, 291–292. [Google Scholar] [CrossRef] [PubMed]
- Rasaiah, J.C.; Garde, S.; Hummer, G. Water in nonpolar confinement: From nanotubes to proteins and beyond. Annu. Rev. Phys. Chem. 2008, 59, 713–740. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Matsumoto, T.; Hirai, D.; Niino, T. Newly developed encapsulation-dehydration protocol for plantcryopreservation. Cryoletters 1999, 21, 53–62. [Google Scholar]
- Kauzmann, W. Some factors in the interpretation of protein denaturation. Adv. Protein Chem. 1959, 14, 1–63. [Google Scholar] [PubMed]
- Gallo, P.; Amann-Winkel, K.; Angell, C.A.; Anisimov, M.A.; Caupin, F.; Chakravarty, C.; Lascaris, E.; Loerting, T.; Panagiotopoulos, A.Z.; Russo, J.; et al. Water: A Tale of Two Liquids. Chem. Rev. 2016, 116, 7463–7500. [Google Scholar] [CrossRef] [PubMed]
- Stanley, H.; Kumar, P.; Xu, L.; Yan, Z.; Mazza, M.; Buldyrev, S.; Chen, S.H.; Mallamace, F. The puzzling unsolved mysteries of liquid water: Some recent progress. Phys. A Stat. Mech. Its Appl. 2007, 386, 729–743. [Google Scholar] [CrossRef]
- Debenedetti, P.G. Supercooled and glassy water. J. Phys. Condens. Matter 2003, 15, R1669–R1726. [Google Scholar] [CrossRef]
- Sellberg, J.A.; Huang, C.; McQueen, T.A.; Loh, N.D.; Laksmono, H.; Schlesinger, D.; Sierra, R.G.; Nordlund, D.; Hampton, C.Y.; Starodub, D.; et al. Ultrafast X-ray probing of water structure below the homogeneous ice nucleation temperature. Nature 2014, 510, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Holten, V.; Bertrand, C.E.; Anisimov, M.A.; Sengers, J.V. Thermodynamics of supercooled water. J. Chem. Phys. 2012, 136, 094507. [Google Scholar] [CrossRef] [PubMed]
- Speedy, R.J.; Angell, C.A. Isothermal compressibility of supercooled water and evidence for a thermodynamic singularity at 45 ∘C. J. Chem. Phys. 1976, 65, 851. [Google Scholar] [CrossRef]
- Mishima, O.; Stanley, H.E. The relationship between liquid, supercooled and glassy water. Nature 1998, 396, 329–335. [Google Scholar]
- Mishima, O.; Calvert, L.D.; Whalley, E. An apparently first-order transition between two amorphous phases of ice induced by pressure. Nature 1985, 314, 76–78. [Google Scholar] [CrossRef]
- Winkel, K.; Elsaesser, M.; Mayer, E.; Loerting, T. Water polyamorphism: Reversibility and (dis)continuity. J. Chem. Phys. 2008, 128, 044510. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.U.; Barstow, B.; Tate, M.V.; Gruner, S.M. Evidence for liquid water during the high-density to low-density amorphous ice transition. Proc. Natl. Acad. Sci. USA 2009, 106, 4596–4600. [Google Scholar] [CrossRef] [PubMed]
- Winkel, K.; Mayer, E.; Loerting, T. Equilibrated High-Density Amorphous Ice and Its First-Order Transition to the Low-Density Form. J. Phys. Chem. B 2011, 115, 14141–14148. [Google Scholar] [CrossRef] [PubMed]
- Klotz, S.; Strässle, T.; Nelmes, R.; Loveday, J.; Hamel, G.; Rousse, G.; Canny, B.; Chervin, J.; Saitta, A. Nature of the polyamorphic transition in ice under pressure. Phys. Rev. Lett. 2005, 94, 025506. [Google Scholar] [CrossRef] [PubMed]
- Stanley, H.E. Unsolved Mysteries of Water in Its Liquid and Glass States. MRS Bull. 1999, 24, 22–30. [Google Scholar] [CrossRef]
- Mishima, O.; Stanley, H.E. Decompression-induced melting of ice IV and the liquid-liquid transition in water. Nature 1998, 392, 164–168. [Google Scholar] [CrossRef]
- Speedy, R.J. Stability-limit conjecture. An interpretation of the properties of water. J. Chem. Phys. 1982, 86, 982–991. [Google Scholar] [CrossRef]
- Poole, P.H.; Sciortino, F.; Essmann, U.; Stanley, H.E. Phase behaviour of metastable water. Nature 1992, 360, 324–328. [Google Scholar] [CrossRef]
- Holten, V.; Anisimov, M. Entropy-driven liquid-liquid separation in supercooled water. Sci. Rep. 2012, 2, 713. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.; Pettersson, L.G.M. Perspective on the structure of liquid water. Chem. Phys. 2011, 389, 1–34. [Google Scholar] [CrossRef]
- Harrington, S.; Poole, P.H.; Sciortino, F.; Stanley, H.E. Equation of state of supercooled water simulated using the extended simple point charge intermolecular potential. J. Chem. Phys. 1997, 107, 7443–7450. [Google Scholar] [CrossRef]
- Yamada, M.; Mossa, S.; Stanley, H.; Sciortino, F. Interplay between Time-Temperature Transformation and the Liquid-Liquid Phase Transition in Water. Phys. Rev. Lett. 2002, 88, 195701. [Google Scholar] [CrossRef] [PubMed]
- Poole, P.H.; Saika-Voivod, I.; Sciortino, F. Density minimum and liquid-liquid phase transition. J. Phys. Condens. Matter 2005, 17, L431. [Google Scholar] [CrossRef]
- Paschek, D. How the liquid-liquid transition affects hydrophobic hydration in deeply supercooled water. Phys. Rev. Lett. 2005, 94, 217802. [Google Scholar] [CrossRef] [PubMed]
- Corradini, D.; Rovere, M.; Gallo, P. A route to explain water anomalies from results on an aqueous solution of salt. J. Chem. Phys. 2010, 132, 134508. [Google Scholar] [CrossRef] [PubMed]
- Abascal, J.L.F.; Vega, C. Widom line and the liquid-liquid critical point for the TIP4P/2005 water model. J. Chem. Phys. 2010, 133, 234502. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.; Car, R.; Debenedetti, P. The liquid-liquid transition in supercooled ST2 water: A comparison between umbrella sampling and well-tempered metadynamics. Faraday Discuss. 2013, 167, 77. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.; Martelli, F.; Liu, Y.; Car, R.; Panagiotopoulos, A.Z.; Debenedetti, P.G. Metastable liquid-liquid transition in a molecular model of water. Nature 2014, 510, 385. [Google Scholar] [CrossRef] [PubMed]
- Poole, P.H.; Bowles, R.K.; Saika-Voivod, I.; Sciortino, F. Free energy surface of ST2 water near the liquid-liquid phase transition. J. Chem. Phys. 2013, 138, 034505. [Google Scholar] [CrossRef] [PubMed]
- Gallo, P.; Sciortino, F. Ising Universality Class for the Liquid-Liquid Critical Point of a One Component Fluid: A Finite-Size Scaling Test. Phys. Rev. Lett. 2012, 109, 177801. [Google Scholar] [CrossRef] [PubMed]
- McMillan, P.F.; Stanley, E.H. Fluid phases: Going supercritical. Nat. Phys. 2010, 6, 479–480. [Google Scholar] [CrossRef]
- Xu, L.; Kumar, P.; Buldyrev, S.V.; Chen, S.H.; Poole, P.H.; Sciortino, F.; Stanley, H.E. Relation between the Widom line and the dynamic crossover in systems with a liquid-liquid phase transition. Proc. Natl. Acad. Sci. USA 2005, 102, 16558–16562. [Google Scholar] [CrossRef] [PubMed]
- Franzese, G.; Stanley, H.E. The Widom line of supercooled water. J. Phys. Condens. Matter 2007, 19, 205126. [Google Scholar] [CrossRef]
- Gallo, P.; Sciortino, F.; Tartaglia, P.; Chen, S.H. Slow Dynamics of Water Molecules in Supercooled States. Phys. Rev. Lett. 1996, 76, 2730–2733. [Google Scholar] [CrossRef] [PubMed]
- Sciortino, F.; Gallo, P.; Tartaglia, P.; Chen, S.H. Supercooled water and the kinetic glass transition. Phys. Rev. E 1996, 54, 6331. [Google Scholar] [CrossRef]
- Torre, R.; Bartolini, P.; Righini, R. Structural relaxation in supercooled water by time-resolved spectroscopy. Nature 2004, 428, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Götze, W.; Sjogren, L. Relaxation processes in supercooled liquids. Rep. Prog. Phys. 1992, 55, 241–376. [Google Scholar] [CrossRef]
- Götze, W. Complex Dynamics of Glass-Forming Liquids; Oxford University Press: New York, NY, USA, 2009; pp. 1–656. [Google Scholar]
- Dehaoui, A.; Issenmann, B.; Caupin, F. Viscosity of deeply supercooled water and its coupling to molecular diffusion. Proc. Natl. Acad. Sci. USA 2015, 112, 12020–12025. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, S.H.; Faraone, A.; Yen, C.W.; Mou, C.Y. Pressure dependence of fragile-to-strong transition and a possible second critical point in supercooled confined water. Phys. Rev. Lett. 2005, 95, 117802. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, S.H.; Faraone, A.; Yen, C.W.; Mou, C.Y.; Kolesnikov, A.I.; Mamontov, E.; Leao, J. Quasielastic and inelastic neutron scattering investigation of fragile-to-strong crossover in deeply supercooled water confined in nanoporous silica matrices. J. Phys. Condens. Matter 2006, 18, S2261. [Google Scholar] [CrossRef]
- Mallamace, F.; Broccio, M.; Corsaro, C.; Faraone, A.; Wanderlingh, U.; Liu, L.; Mou, C.; Chen, S.H. The fragile-to-strong dynamic crossover transition in confined water: Nuclear magnetic resonance results. J. Chem. Phys. 2006, 124, 161102. [Google Scholar] [CrossRef] [PubMed]
- Faraone, A.; Liu, L.; Mou, C.Y.; Yen, C.W.; Chen, S.H. Fragile-to- strong liquid transition in deeply supercooled confined water. J. Chem. Phys. 2004, 121, 10843–10846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lagi, M.; Fratini, E.; Baglioni, P.; Mamontov, E.; Chen, S.H. Dynamic susceptibility of supercooled water and its relation to the dynamic crossover phenomenon. Phys. Rev. E 2009, 79, 040201. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Le, P.; Ito, K.; Leão, J.B.; Tyagi, M.; Chen, S.H. Dynamic crossover in deeply cooled water confined in MCM-41 at 4 kbar and its relation to the liquid-liquid transition hypothesis. J. Chem. Phys. 2015, 143, 114508. [Google Scholar] [CrossRef] [PubMed]
- Starr, F.W.; Sciortino, F.; Stanley, H.E. Dynamics of simulated water under pressure. Phys. Rev. E 1999, 60, 6757–6768. [Google Scholar] [CrossRef]
- Gallo, P.; Rovere, M. Mode coupling and fragile to strong transition in supercooled TIP4P water. J. Chem. Phys. 2012, 137, 164503. [Google Scholar] [CrossRef] [PubMed]
- Gallo, P.; Rovere, M.; Chen, S.H. Dynamic Crossover in Supercooled Confined Water: Understanding Bulk Properties through Confinement. J. Phys. Chem. Lett. 2010, 1, 729–733. [Google Scholar] [CrossRef]
- De Marzio, M.; Camisasca, G.; Rovere, M.; Gallo, P. Mode Coupling Theory And Fragile To Strong Transition in Supercooled TIP4P/2005 Water. J. Chem. Phys. 2016, 144, 074503. [Google Scholar] [CrossRef] [PubMed]
- Franzese, G.; Bianco, V. Water at Biological and Inorganic Interfaces. Food Biophys. 2013, 8, 153–169. [Google Scholar] [CrossRef]
- Chen, S.H.; Liu, L.; Fratini, E.; Baglioni, P.; Faraone, A.; Mamontov, E.; Fomina, M. Observation of fragile-to-strong dynamic crossover in protein hydration water. Proc. Natl. Acad. Sci. USA 2006, 103, 9012–9016. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Yan, Z.; Xu, L.; Mazza, M.G.; Buldyrev, S.V.; Chen, S.H.; Sastry, S.; Stanley, H.E. Glass transition in biomolecules and the liquid-liquid critical point of water. Phys. Rev. Lett. 2006, 97, 177802. [Google Scholar] [CrossRef] [PubMed]
- Amann-Winkel, K.; Boehmer, R.; Fujara, F.; Gainaru, C.; Geil, B.; Loerting, T. Colloquium: Water’s controversial glass transitions. Rev. Mod. Phys. 2016, 88, 011002. [Google Scholar] [CrossRef]
- Laage, D.; Elsaesser, T.; Hynes, J.T. Water Dynamics in the Hydration Shells of Biomolecules. Chem. Rev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Gallo, P.; Rovere, M.; Chen, S.H. Anomalous dynamics of water confined in MCM-41 at different hydrations. J. Phys. Condens. Matter 2010, 22, 284102. [Google Scholar] [CrossRef] [PubMed]
- Gallo, P.; Rovere, M.; Chen, S.H. Water confined in MCM-41: A mode coupling theory analysis. J. Phys. Condens. Matter 2012, 24, 064109. [Google Scholar] [CrossRef] [PubMed]
- De Marzio, M.; Camisasca, G.; Conde, M.M.; Rovere, M.; Gallo, P. Structural properties and fragile to strong transition in confined water. J. Chem. Phys 2016, 146, 084505. [Google Scholar] [CrossRef] [PubMed]
- Gallo, P.; Ricci, M.; Rovere, M. Layer analysis of the structure of water confined in vycor glass. J. Chem. Phys. 2002, 116, 342–346. [Google Scholar] [CrossRef]
- Rosenfeld, Y. A quasi-universal scaling law for atomic transport in simple fluids. J. Phys. Condens. Matter 1999, 11, 5415–5427. [Google Scholar] [CrossRef]
- Jabes, B.S.; Agarwal, M.; Chakravarty, C. Tetrahedral order, pair correlation entropy, and waterlike liquid state anomalies: Comparison of GeO2 with BeF2, SiO2, and H2O. J. Chem. Phys. 2010, 132, 234507. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.; Singh, M.; Sharma, R.; Alam, M.P.; Chakravarty, C. Relationship between Structure, Entropy, and Diffusivity in Water and Water-Like Liquids. J. Phys. Chem. B 2010, 114, 6995–7001. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Agarwal, M.; Chakravarty, C. Estimating the entropy of liquids from atom–atom radial distribution functions: Silica, beryllium fluoride and water. Mol. Phys. 2008, 106, 1925–1938. [Google Scholar] [CrossRef]
- Mittal, J.; Errington, J.R.; Truskett, T.M. Relationship between thermodynamics and dynamics of supercooled liquids. J. Chem. Phys. 2006, 125, 076102. [Google Scholar] [CrossRef] [PubMed]
- Errington, J.R.; Truskett, T.M.; Mittal, J. Excess-entropy-based anomalies for a waterlike fluid. J. Chem. Phys. 2006, 125, 244502. [Google Scholar] [CrossRef] [PubMed]
- Scala, A.; Starr, F.W.; Nave, E.L.; Sciortino, F.; Stanley, H.E. Configurational entropy and diffusivity of supercooled water. Nature 2000, 406, 166–169. [Google Scholar] [PubMed]
- Fomin, Y.D.; Ryzhov, V.N.; Gribova, N.V. Breakdown of excess entropy scaling for systems with thermodynamic anomalies. Phys. Rev. E 2010, 81, 061201. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; Head-Gordon, T. Assessing thermodynamic-dynamic relationships for waterlike liquids. J. Chem. Phys. 2009, 130, 214510. [Google Scholar] [CrossRef] [PubMed]
- Nandi, M.K.; Banerjee, A.; Sengupta, S.; Sastry, S.; Bhattacharyya, S.M. Unraveling the success and failure of mode coupling theory from consideration of entropy. J. Chem. Phys. 2015, 143, 174504. [Google Scholar] [CrossRef] [PubMed]
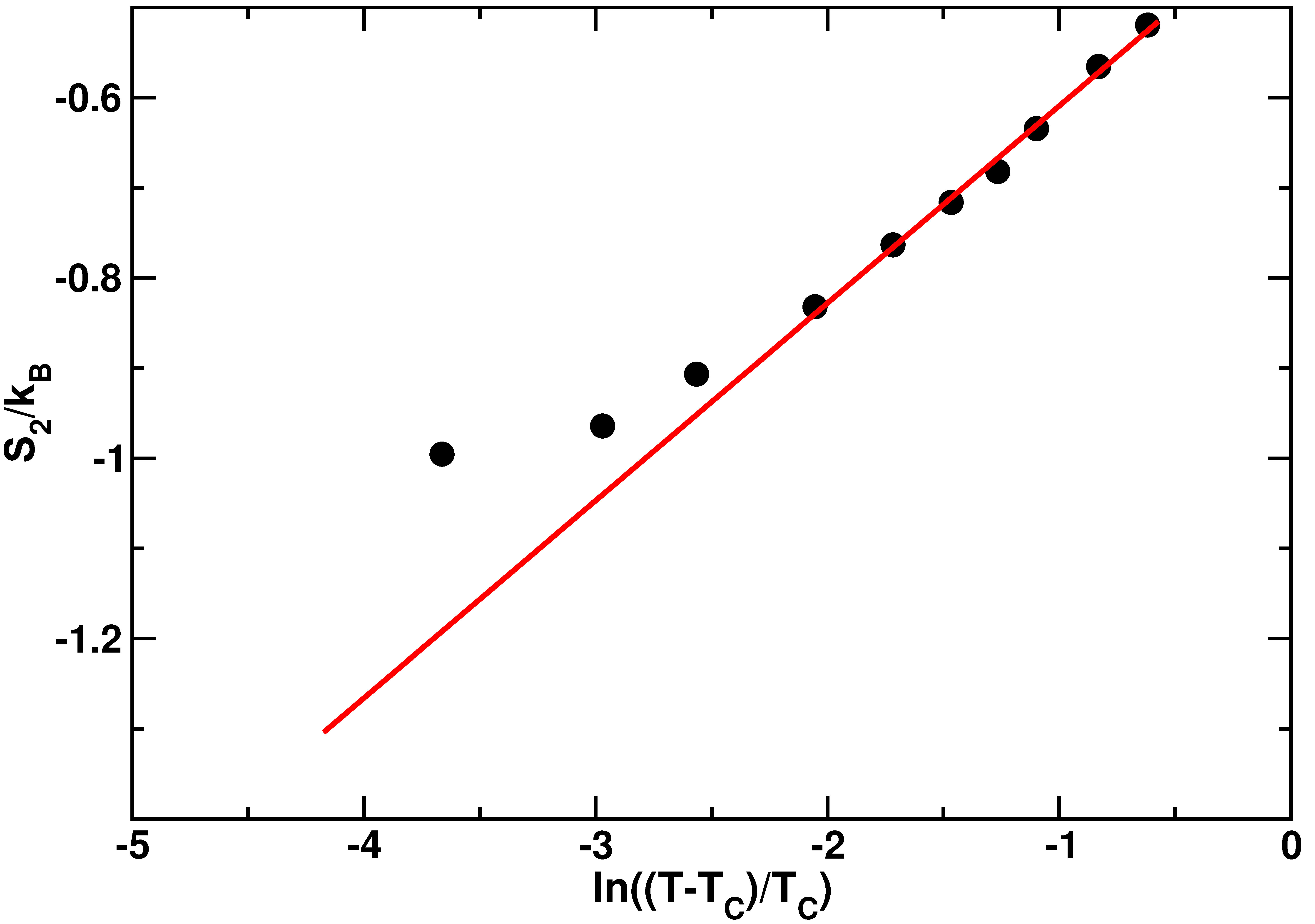
- Gallo, P.; Rovere, M. Relation between the two-body entropy and the relaxation time in supercooled water. Phys. Rev. E 2015, 91, 012107. [Google Scholar] [CrossRef] [PubMed]
- Corradini, D.; Strekalova, E.G.; Stanley, H.E.; Gallo, P. Microscopic mechanism of protein cryopreservation in an aqueous solution with trehalose. Sci. Rep. 2013, 3, 1218. [Google Scholar] [CrossRef] [PubMed]
- Camisasca, G.; De Marzio, M.; Corradini, D.; Gallo, P. Two structural relaxations in protein hydration water and their dynamic crossovers. J. Chem. Phys. 2016, 145, 044503. [Google Scholar] [CrossRef] [PubMed]
- Magno, A.; Gallo, P. Understanding the Mechanisms of Bioprotection: A Comparative Study of Aqueous Solutions of Trehalose and Maltose upon Supercooling. J. Phys. Chem. Lett. 2011, 2, 977–982. [Google Scholar] [CrossRef]
- Comez, L.; Lupi, L.; Morresi, A.; Paolantoni, M.; Sassi, P.; Fioretto, D. More Is Different: Experimental Results on the Effect of Biomolecules on the Dynamics of Hydration Water. J. Phys. Chem. Lett. 2013, 4, 1188–1192. [Google Scholar] [CrossRef] [PubMed]
- Paolantoni, M.; Comez, L.; Gallina, M.E.; Sassi, P.; Scarponi, F.; Fioretto, D.; Morresi, A. Light scattering spectra of water in trehalose aqueous solutions: Evidence for two different solvent relaxation processes. J. Phys. Chem. B 2009, 113, 7874–7878. [Google Scholar] [CrossRef] [PubMed]
- Starr, F.W.; Harrington, S.; Sciortino, F.; Stanley, H.E. Slow Dynamics of Water under Pressure. Phys. Rev. Lett. 1999, 82, 3629–3632. [Google Scholar] [CrossRef]
- Mallamace, F.; Chen, S.H.; Broccio, M.; Corsaro, C.; Crupi, V.; Majolino, D.; Venuti, V.; Baglioni, P.; Fratini, E.; Vannucci, C.; et al. Role of the solvent in the dynamical transitions of proteins: the case of the lysozyme-water system. J. Chem. Phys. 2007, 127, 045104. [Google Scholar] [CrossRef] [PubMed]
- Lagi, M.; Chu, X.; Kim, C.; Mallamace, F.; Baglioni, P.; Chen, S.H. The low-temperature dynamic crossover phenomenon in protein hydration water: Simulations vs. experiments. J. Phys. Chem. B 2008, 112, 1571–1575. [Google Scholar] [CrossRef] [PubMed]
- Spohr, E.; Hartnig, C.; Gallo, P.; Rovere, M. Water in porous glasses. A computer simulation study. J. Mol. Liq. 1999, 80, 165–178. [Google Scholar] [CrossRef]
- Pronk, S.; Páll, S.; Schulz, R.; Larsson, P.; Bjelkmar, P.; Apostolov, R.; Shirts, M.R.; Smith, J.C.; Kasson, P.M.; van der Spoel, D.; et al. GROMACS 4.5: A high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 2013, 29, 845–854. [Google Scholar] [CrossRef] [PubMed]
- MacKerell, A.D.; Banavali, N.K. All-atom empirical force field for nucleic acids: II. Application to molecular dynamics simulations of DNA and RNA in solution. J. Comput. Chem. 2000, 21, 105–120. [Google Scholar] [CrossRef]
- MacKerell, A.D.; Feig, M.; Brooks, C.L. Extending the treatment of backbone energetics in protein force fields: limitations of gas-phase quantum mechanics in reproducing protein conformational distributions in molecular dynamics simulations. J. Comput. Chem. 2004, 25, 1400–1415. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, H.J.C.; Grigera, J.R.; Straatsma, T.P. The missing term in effective pair potentials. J. Phys. Chem. 1987, 91, 6269–6271. [Google Scholar] [CrossRef]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camisasca, G.; De Marzio, M.; Rovere, M.; Gallo, P. Slow Dynamics and Structure of Supercooled Water in Confinement. Entropy 2017, 19, 185. https://doi.org/10.3390/e19040185
Camisasca G, De Marzio M, Rovere M, Gallo P. Slow Dynamics and Structure of Supercooled Water in Confinement. Entropy. 2017; 19(4):185. https://doi.org/10.3390/e19040185
Chicago/Turabian StyleCamisasca, Gaia, Margherita De Marzio, Mauro Rovere, and Paola Gallo. 2017. "Slow Dynamics and Structure of Supercooled Water in Confinement" Entropy 19, no. 4: 185. https://doi.org/10.3390/e19040185
APA StyleCamisasca, G., De Marzio, M., Rovere, M., & Gallo, P. (2017). Slow Dynamics and Structure of Supercooled Water in Confinement. Entropy, 19(4), 185. https://doi.org/10.3390/e19040185