Information-Domain Analysis of Cardiovascular Complexity: Night and Day Modulations of Entropy and the Effects of Hypertension



Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Data Collection
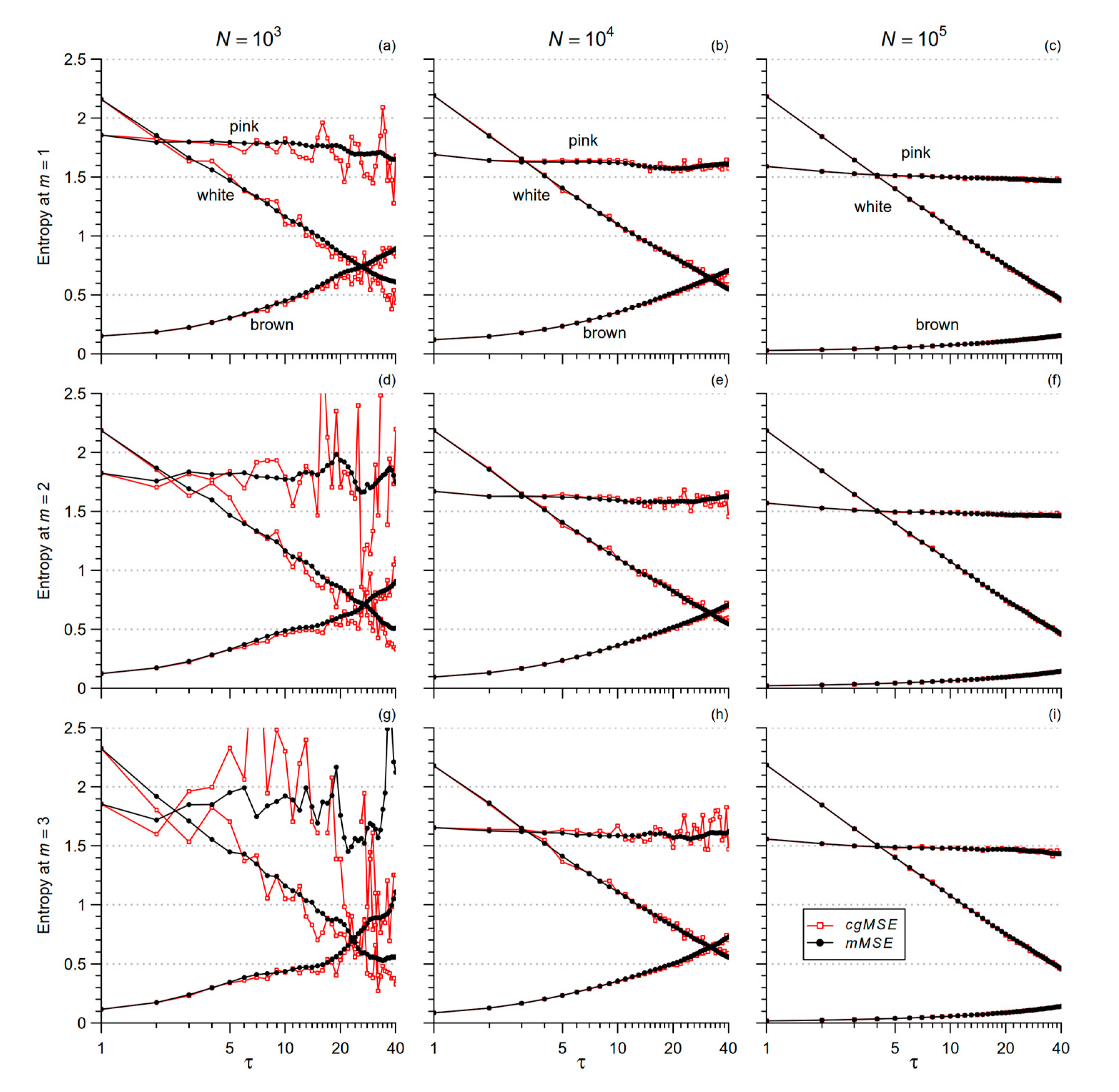
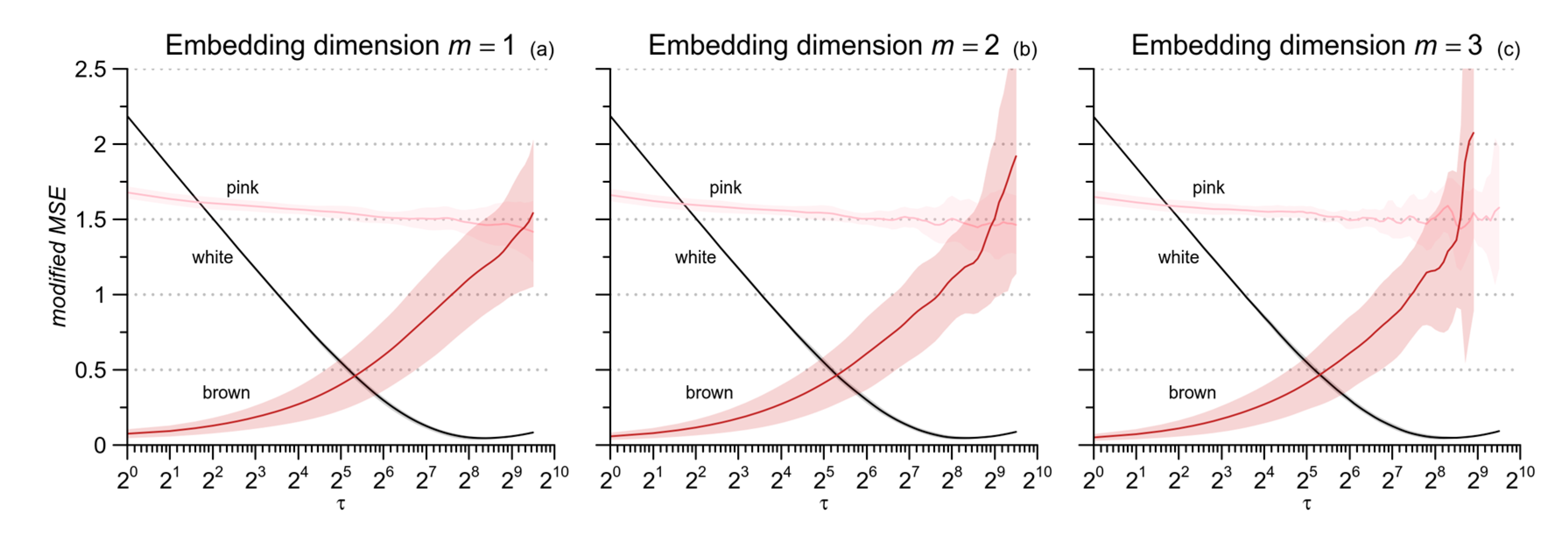
2.2. Coarse-Grained MSE and Modified MSE
2.3. MSE of Cardiovascular Series: From Scales in Beats to Temporal Scales in Seconds
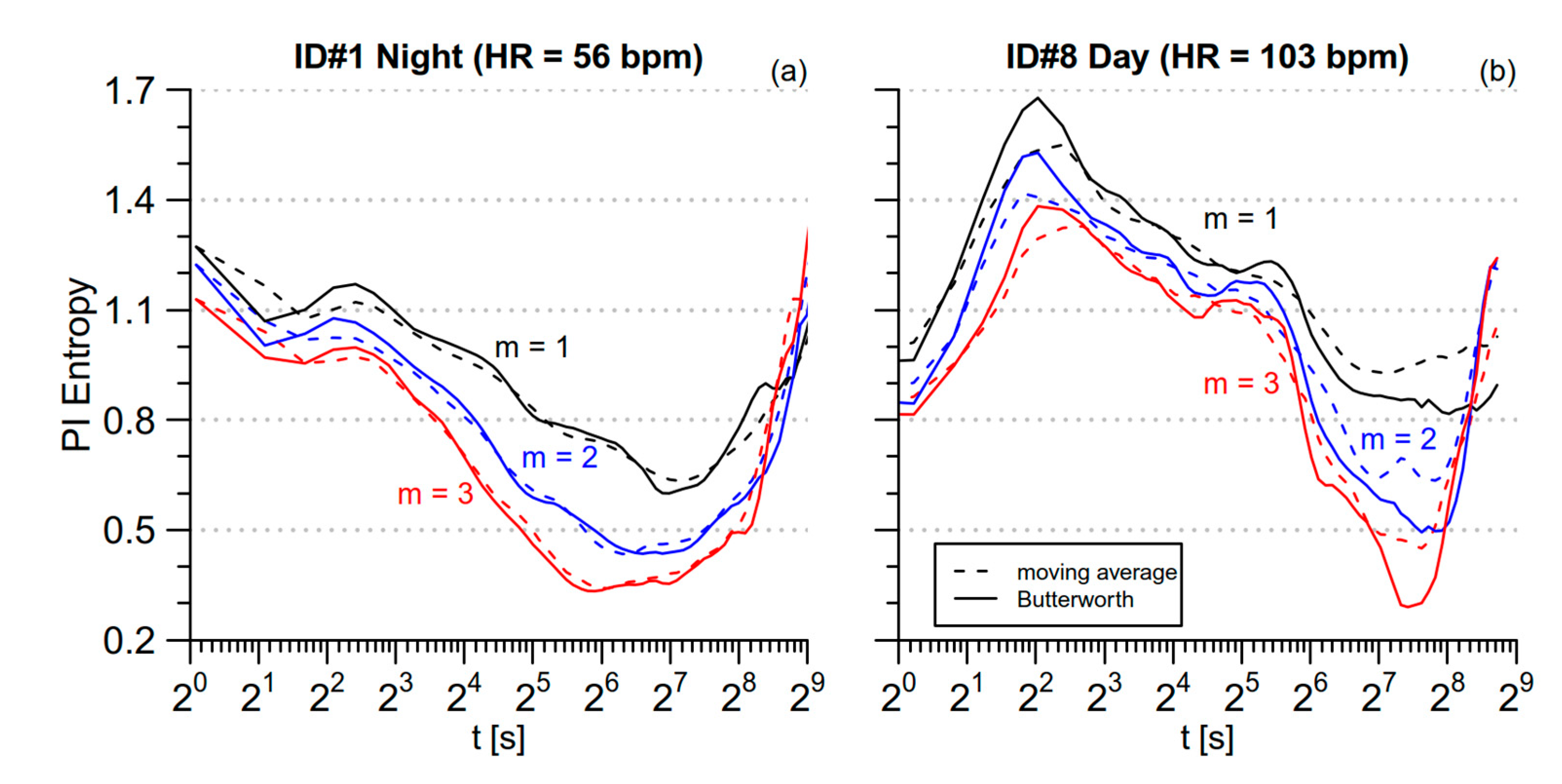
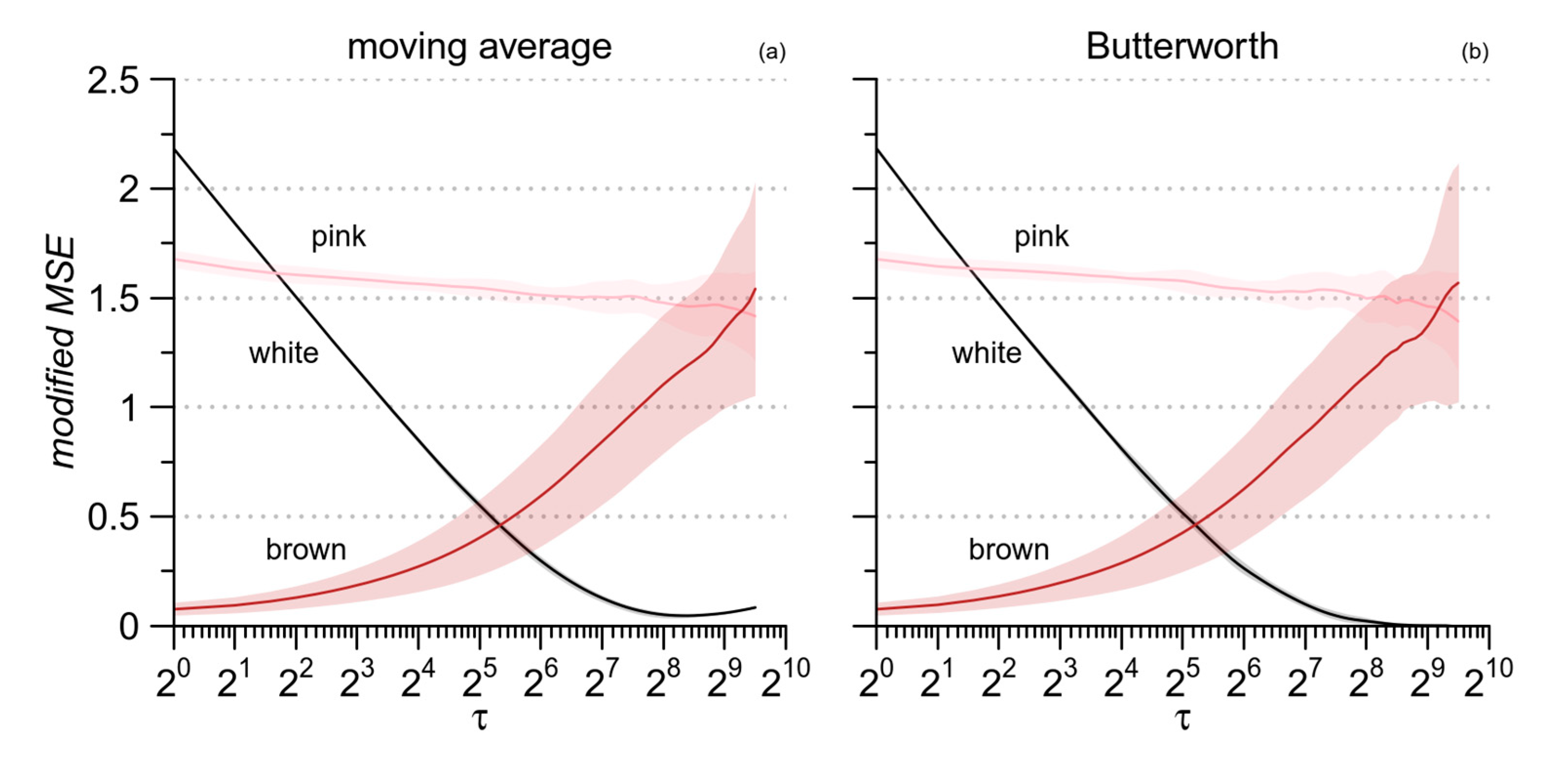
2.4. Low-Pass Filtering for mMSE Estimates
2.5. Multiscale Cross-Entropy between SBP and PI
2.6. Statistical Analysis
3. Results
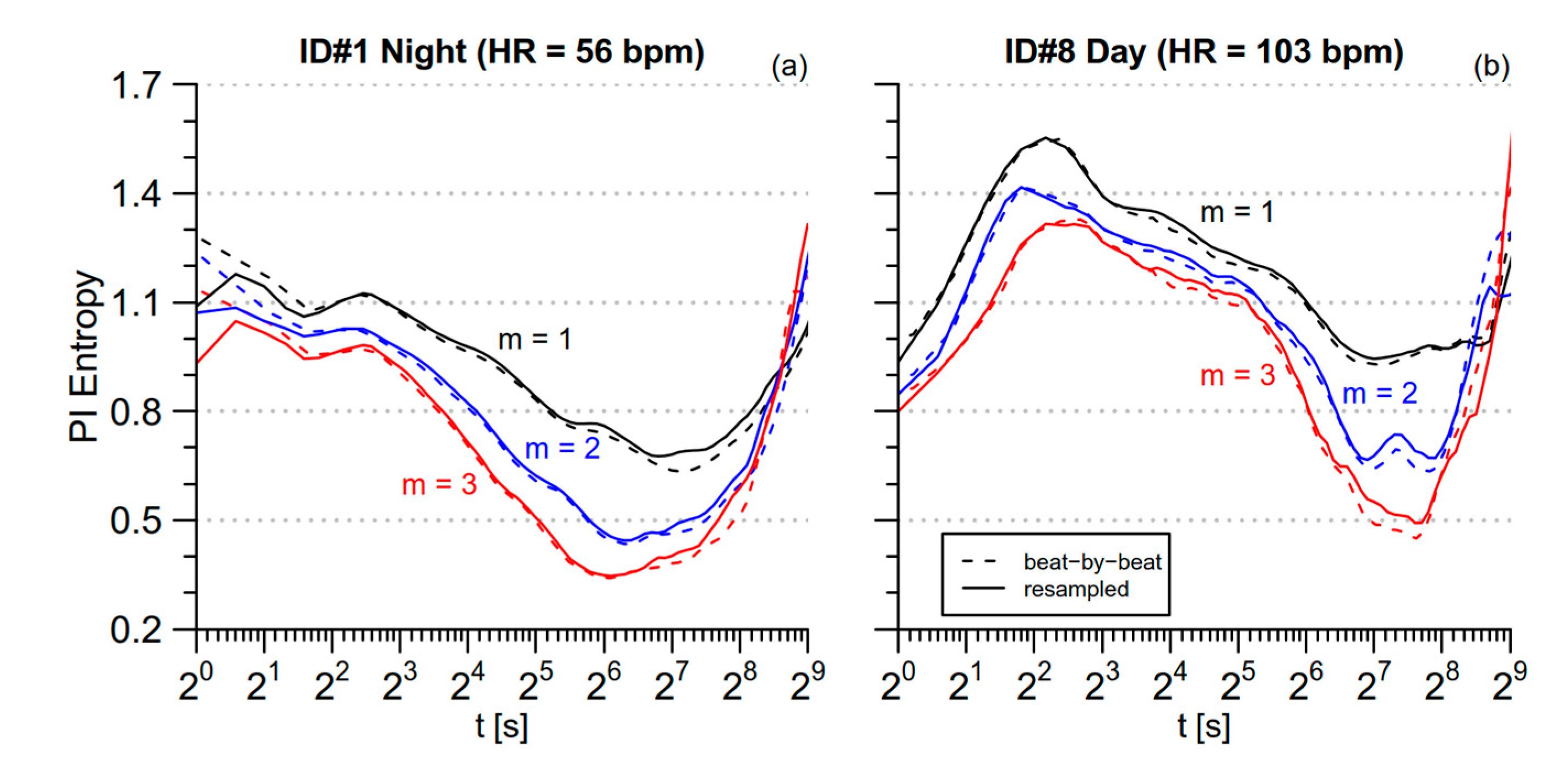
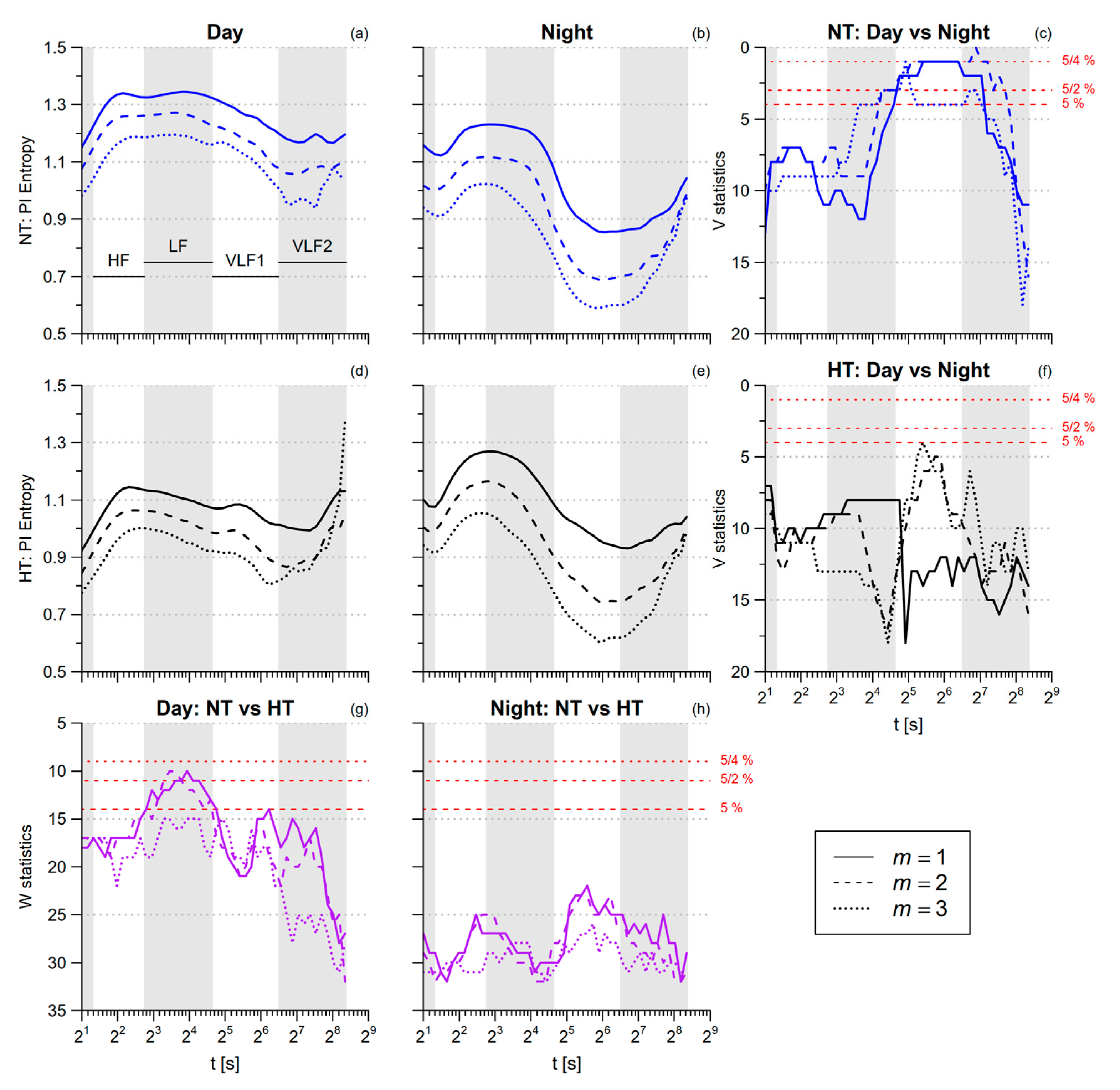
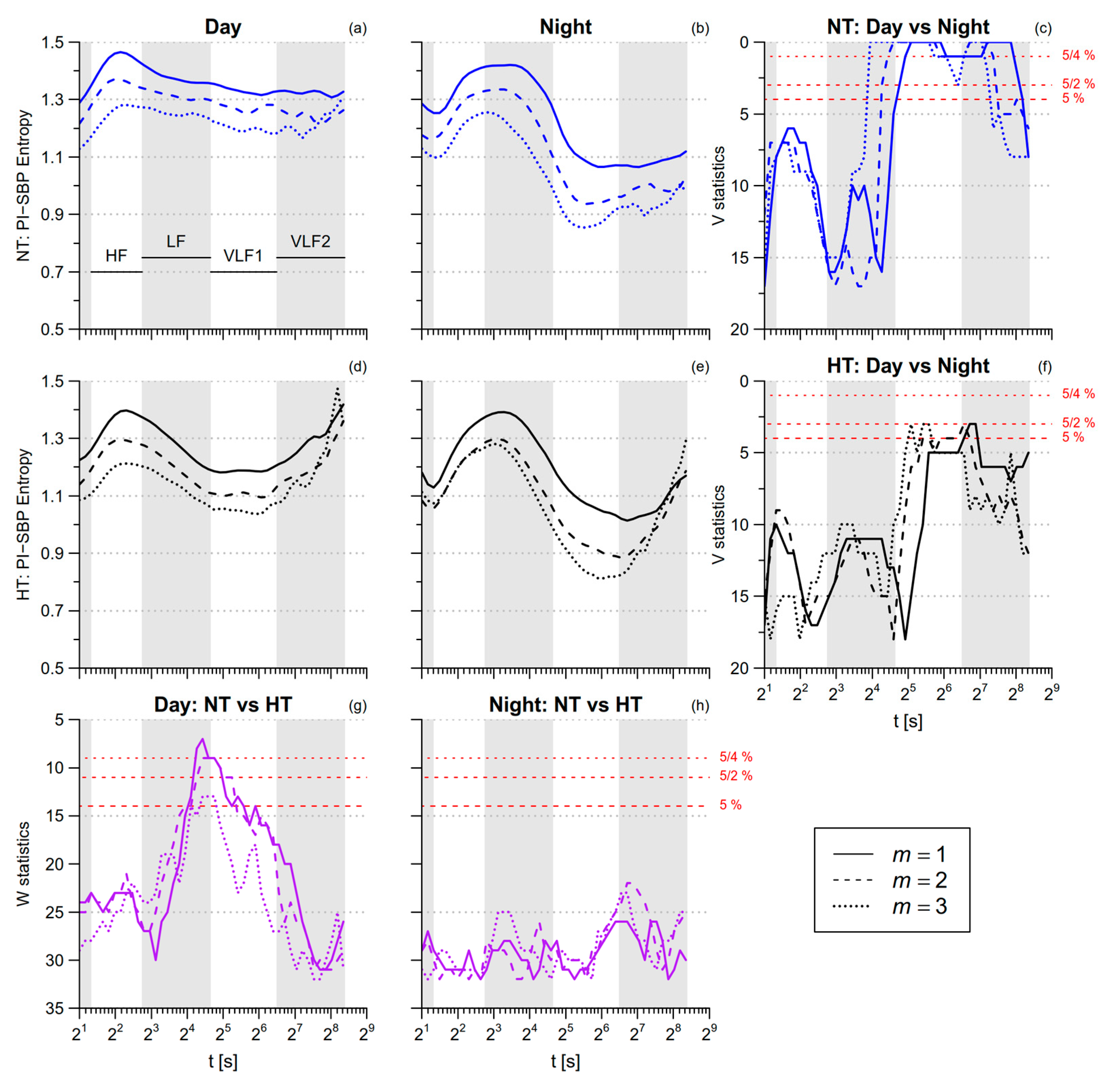
3.1. PI Entropy
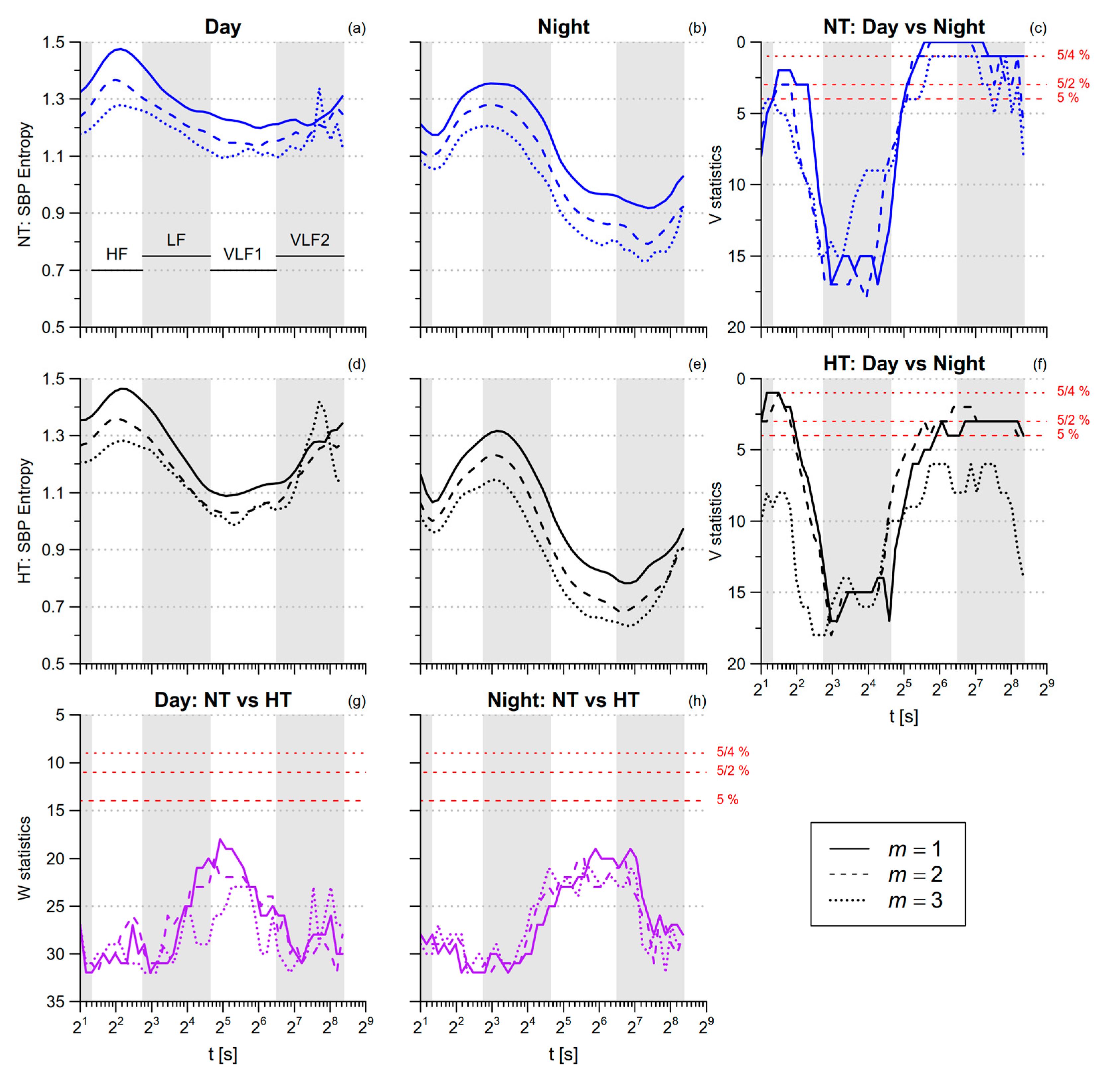
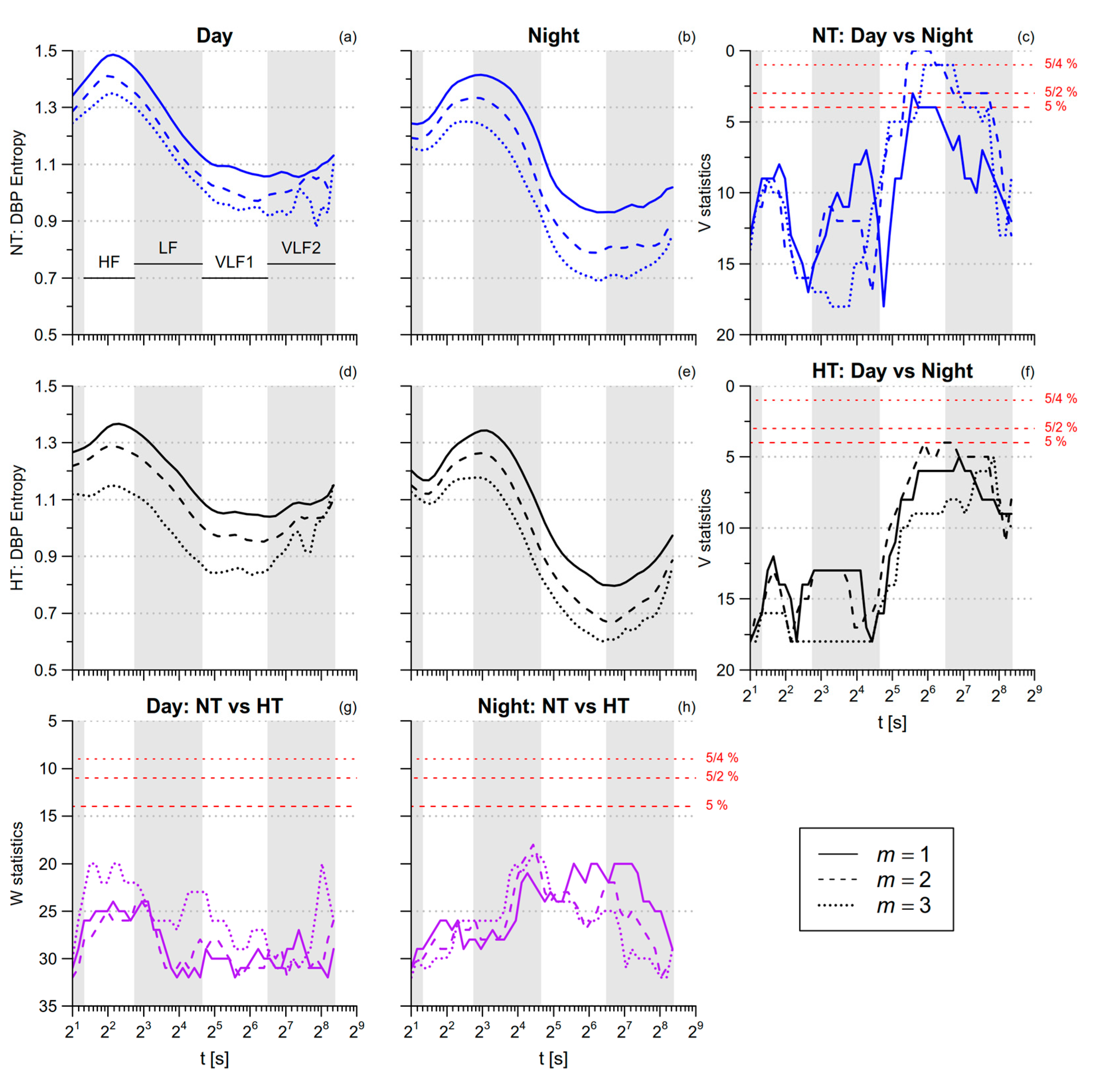
3.2. Blood Pressure Entropy
3.3. SBP-PI Cross-Entropy
4. Discussion
4.1. Day-Night Modulations in Normotensive Subjects
4.2. Hypertension and Entropy
4.3. Limitations and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pincus, S.M.; Gladstone, I.M.; Ehrenkranz, R.A. A regularity statistic for medical data analysis. J. Clin. Monit. 1991, 7, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Pincus, S.M. Approximate entropy as a measure of system complexity. Proc. Natl. Acad. Sci. USA 1991, 88, 2297–2301. [Google Scholar] [CrossRef]
- Richman, J.S.; Moorman, J.R. Physiological time-series analysis using approximate entropy and sample entropy. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H2039–H2049. [Google Scholar] [CrossRef]
- Pincus, S.M.; Mulligan, T.; Iranmanesh, A.; Gheorghiu, S.; Godschalk, M.; Veldhuis, J.D. Older males secrete luteinizing hormone and testosterone more irregularly, and jointly more asynchronously, than younger males. Proc. Natl. Acad. Sci. USA 1996, 93, 14100–14105. [Google Scholar] [CrossRef]
- Costa, M.; Goldberger, A.L.; Peng, C.-K. Multiscale Entropy Analysis of Complex Physiologic Time Series. Phys. Rev. Lett. 2002, 89, 068102. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Goldberger, A.L.; Peng, C.-K. Multiscale entropy analysis of biological signals. Phys. Rev. E 2005, 71, 021906. [Google Scholar] [CrossRef]
- Valencia, J.F.; Porta, A.; Vallverdu, M.; Claria, F.; Baranowski, R.; Orlowska-Baranowska, E.; Caminal, P. Refined Multiscale Entropy: Application to 24-h Holter Recordings of Heart Period Variability in Healthy and Aortic Stenosis Subjects. IEEE Trans. Biomed. Eng. 2009, 56, 2202–2213. [Google Scholar] [CrossRef]
- Wu, S.-D.; Wu, C.-W.; Lee, K.-Y.; Lin, S.-G. Modified multiscale entropy for short-term time series analysis. Physica A 2013, 392, 5865–5873. [Google Scholar] [CrossRef]
- Trunkvalterova, Z.; Javorka, M.; Tonhajzerova, I.; Javorkova, J.; Lazarova, Z.; Javorka, K.; Baumert, M. Reduced short-term complexity of heart rate and blood pressure dynamics in patients with diabetes mellitus type 1: Multiscale entropy analysis. Physiol. Meas. 2008, 29, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, E.; Kiyono, K.; Hayano, J.; Yamamoto, Y.; Inamasu, J.; Yamamoto, M.; Ichikawa, T.; Sobue, Y.; Harada, M.; Ozaki, Y. Multiscale Entropy of the Heart Rate Variability for the Prediction of an Ischemic Stroke in Patients with Permanent Atrial Fibrillation. PLoS ONE 2015, 10, e0137144. [Google Scholar] [CrossRef]
- Ho, Y.-L.; Lin, C.; Lin, Y.-H.; Lo, M.-T. The Prognostic Value of Non-Linear Analysis of Heart Rate Variability in Patients with Congestive Heart Failure—A Pilot Study of Multiscale Entropy. PLoS ONE 2011, 6, e18699. [Google Scholar] [CrossRef] [PubMed]
- Bari, V.; Valencia, J.F.; Vallverdú, M.; Girardengo, G.; Marchi, A.; Bassani, T.; Caminal, P.; Cerutti, S.; George, A.L.; Brink, P.A.; et al. Multiscale Complexity Analysis of the Cardiac Control Identifies Asymptomatic and Symptomatic Patients in Long QT Syndrome Type 1. PLoS ONE 2014, 9, e93808. [Google Scholar] [CrossRef]
- Pan, W.-Y.; Su, M.-C.; Wu, H.-T.; Lin, M.-C.; Tsai, I.-T.; Sun, C.-K. Multiscale Entropy Analysis of Heart Rate Variability for Assessing the Severity of Sleep Disordered Breathing. Entropy 2015, 17, 231–243. [Google Scholar] [CrossRef]
- Chen, C.-H.; Huang, P.-W.; Tang, S.-C.; Shieh, J.-S.; Lai, D.-M.; Wu, A.-Y.; Jeng, J.-S. Complexity of Heart Rate Variability Can Predict Stroke-In-Evolution in Acute Ischemic Stroke Patients. Sci. Rep. 2015, 5, 17552. [Google Scholar] [CrossRef]
- Chen, C.-H.; Tang, S.-C.; Lee, D.-Y.; Shieh, J.-S.; Lai, D.-M.; Wu, A.-Y.; Jeng, J.-S. Impact of Supratentorial Cerebral Hemorrhage on the Complexity of Heart Rate Variability in Acute Stroke. Sci. Rep. 2018, 8, 11473. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.-C.; Jen, H.-I.; Lin, Y.-H.; Hung, C.-S.; Jou, W.-J.; Huang, P.-W.; Shieh, J.-S.; Ho, Y.-L.; Lai, D.-M.; Wu, A.-Y.; et al. Complexity of heart rate variability predicts outcome in intensive care unit admitted patients with acute stroke. J. Neurol. Neurosurg. Psychiatry 2015, 86, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Norris, P.R.; Anderson, S.M.; Jenkins, J.M.; Williams, A.E.; Morris, J.A.J. Heart Rate Multiscale Entropy at Three Hours Predicts Hospital Mortality in 3,154 Trauma Patients. Shock 2008, 30, 17. [Google Scholar] [CrossRef] [PubMed]
- Valenza, G.; Nardelli, M.; Bertschy, G.; Lanata, A.; Scilingo, E.P. Mood states modulate complexity in heartbeat dynamics: A multiscale entropy analysis. EPL 2014, 107, 18003. [Google Scholar] [CrossRef]
- Riganello, F.; Larroque, S.K.; Bahri, M.A.; Heine, L.; Martial, C.; Carrière, M.; Charland-Verville, V.; Aubinet, C.; Vanhaudenhuyse, A.; Chatelle, C.; et al. A Heartbeat Away From Consciousness: Heart Rate Variability Entropy Can Discriminate Disorders of Consciousness and Is Correlated With Resting-State fMRI Brain Connectivity of the Central Autonomic Network. Front. Neurol. 2018, 9, 1–18. [Google Scholar] [CrossRef]
- Silva, L.E.V.; Lataro, R.M.; Castania, J.A.; da Silva, C.A.A.; Valencia, J.F.; Murta, L.O.; Salgado, H.C.; Fazan, R.; Porta, A. Multiscale entropy analysis of heart rate variability in heart failure, hypertensive, and sinoaortic-denervated rats: classical and refined approaches. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R150–R156. [Google Scholar] [CrossRef]
- Vandendriessche, B.; Peperstraete, H.; Rogge, E.; Cauwels, P.; Hoste, E.; Stiedl, O.; Brouckaert, P.; Cauwels, A. A Multiscale Entropy-Based Tool for Scoring Severity of Systemic Inflammation. Crit. Care Med. 2014, 42, e560–e569. [Google Scholar] [CrossRef]
- Angelini, L.; Maestri, R.; Marinazzo, D.; Nitti, L.; Pellicoro, M.; Pinna, G.D.; Stramaglia, S.; Tupputi, S.A. Multiscale analysis of short term heart beat interval, arterial blood pressure, and instantaneous lung volume time series. Artif. Intell. Med. 2007, 41, 237–250. [Google Scholar] [CrossRef]
- Castiglioni, P.; Coruzzi, P.; Bini, M.; Parati, G.; Faini, A. Multiscale Sample Entropy of Cardiovascular Signals: Does the Choice between Fixed- or Varying-Tolerance among Scales Influence Its Evaluation and Interpretation? Entropy 2017, 19, 590. [Google Scholar] [CrossRef]
- Turianikova, Z.; Javorka, K.; Baumert, M.; Calkovska, A.; Javorka, M. The effect of orthostatic stress on multiscale entropy of heart rate and blood pressure. Physiol. Meas. 2011, 32, 1425–1437. [Google Scholar] [CrossRef]
- Heart rate variability: Standards of measurement, physiological interpretation and clinical use. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Circulation 1996, 93, 1043–1065. [CrossRef]
- Radaelli, A.; Mancia, G.; De Carlini, C.; Soriano, F.; Castiglioni, P. Patterns of cardiovascular variability after long-term sino-aortic denervation in unanesthetized adult rats. Sci. Rep. 2019, 9, 1232. [Google Scholar] [CrossRef] [PubMed]
- Parati, G.; Castiglioni, P.; Faini, A.; Di Rienzo, M.; Mancia, G.; Barbieri, R.; Saul, J.P. Closed-Loop Cardiovascular Interactions and the Baroreflex Cardiac Arm: Modulations over the 24 Hours and the Effect of Hypertension. Front. Physiol. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Govindan, R.B.; Wilson, J.D.; Eswaran, H.; Lowery, C.L.; Preißl, H. Revisiting sample entropy analysis. Physica A 2007, 376, 158–164. [Google Scholar] [CrossRef]
- Eke, A.; Herman, P.; Kocsis, L.; Kozak, L.R. Fractal characterization of complexity in temporal physiological signals. Physiol. Meas. 2002, 23, R1-38. [Google Scholar] [CrossRef]
- Wilcoxon, F. Individual Comparisons by Ranking Methods. Biometr. Bull. 1945, 1, 80–83. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Porta, A.; Bari, V.; Marchi, A.; De Maria, B.; Castiglioni, P.; di Rienzo, M.; Guzzetti, S.; Cividjian, A.; Quintin, L. Limits of permutation-based entropies in assessing complexity of short heart period variability. Physiol. Meas. 2015, 36, 755–765. [Google Scholar] [CrossRef]
- Porta, A.; Castiglioni, P.; Bari, V.; Bassani, T.; Marchi, A.; Cividjian, A.; Quintin, L.; Di Rienzo, M. K-nearest-neighbor conditional entropy approach for the assessment of the short-term complexity of cardiovascular control. Physiol. Meas. 2013, 34, 17–33. [Google Scholar] [CrossRef]
- de Boer, R.W.; Karemaker, J.M.; Strackee, J. Relationships between short-term blood-pressure fluctuations and heart-rate variability in resting subjects. I: A spectral analysis approach. Med. Biol. Eng. Comput. 1985, 23, 352–358. [Google Scholar] [CrossRef]
- Radaelli, A.; Bernardi, L.; Valle, F.; Leuzzi, S.; Salvucci, F.; Pedrotti, L.; Marchesi, E.; Finardi, G.; Sleight, P. Cardiovascular autonomic modulation in essential hypertension. Effect of tilting. Hypertension 1994, 24, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Castiglioni, P.; Parati, G.; Omboni, S.; Mancia, G.; Imholz, B.P.; Wesseling, K.H.; Di Rienzo, M. Broad-band spectral analysis of 24 h continuous finger blood pressure: comparison with intra-arterial recordings. Clin. Sci. 1999, 97, 129–139. [Google Scholar] [PubMed]
- Constant, I.; Laude, D.; Murat, I.; Elghozi, J.L. Pulse rate variability is not a surrogate for heart rate variability. Clin. Sci. 1999, 97, 391–397. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p Value | ||||||
|---|---|---|---|---|---|---|
| Day | Night | Group | Time | Time *Group | ||
| PI SampEn | ||||||
| m = 1 | NT | 1.02 (0.21) * | 1.31 (0.31) | 0.25 | <0.001 | 0.10 |
| HT | 0.84 (0.14) ** | 1.32 (0.27) | ||||
| m = 2 | NT | 0.94 (0.23) * | 1.22 (0.29) | 0.31 | <0.001 | 0.07 |
| HT | 0.75 (0.15) ** | 1.27 (0.27) | ||||
| m = 3 | NT | 0.88 (0.23) * | 1.06 (0.21) | 0.50 | <0.001 | 0.06 |
| HT | 0.69 (0.17) ** | 1.14 (0.30) | ||||
| SBP SampEn | ||||||
| m = 1 | NT | 1.29 (0.19) | 1.41 (0.30) | 0.45 | <0.05 | >0.99 |
| HT | 1.37 (0.21) | 1.45 (0.29) | ||||
| m = 2 | NT | 1.25 (0.19) | 1.37 (0.29) | 0.49 | <0.05 | 0.92 |
| HT | 1.30 (0.18) | 1.42 (0.29) | ||||
| m = 3 | NT | 1.18 (0.18) | 1.24 (0.25) | 0.34 | 0.19 | 0.83 |
| HT | 1.25 (0.18) | 1.27 (0.28) | ||||
| DBP SampEn | ||||||
| m = 1 | NT | 1.25 (0.24) | 1.35 (0.32) | 0.83 | 0.18 | 0.47 |
| HT | 1.26 (0.26) | 1.31 (0.29) | ||||
| m = 2 | NT | 1.20 (0.25) | 1.30 (0.33) | 0.83 | 0.14 | 0.68 |
| HT | 1.19 (0.27) | 1.26 (0.30) | ||||
| m = 3 | NT | 1.17 (0.25) | 1.25 (0.32) | 0.90 | 0.16 | 0.68 |
| HT | 1.16 (0.27) | 1.23 (0.31) | ||||
| SBP-PI cross-SampEn | ||||||
| m = 1 | NT | 1.22 (0.13) * | 1.47 (0.25) | 0.78 | <0.01 | 0.86 |
| HT | 1.20 (0.15) * | 1.43 (0.28) | ||||
| m = 2 | NT | 1.19 (0.15) * | 1.46 (0.28) | 0.70 | <0.01 | 0.67 |
| HT | 1.15 (0.14) ** | 1.42 (0.28) | ||||
| m = 3 | NT | 1.13 (0.15) * | 1.33 (0.21) | 0.83 | <0.01 | 0.63 |
| HT | 1.11 (0.15) ** | 1.30 (0.27) | ||||
| HF | LF | VLF1 | VLF2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day | Night | Day | Night | Day | Night | Day | Night | ||
| PI mMSE | |||||||||
| m = 1 | NT | 1.31 (0.25) | 1.17 (0.25) | 1.34 (0.19) | 1.19 (0.29) | 1.27 (0.24) | 0.93 (0.25) | 1.18 (0.22) | 0.90 (0.24) |
| HT | 1.05 (0.29) | 1.19 (0.33) | 1.12 (0.25) | 1.21 (0.35) | 1.08 (0.23) | 1.01 (0.26) | 1.03 (0.20) | 0.96 (0.21) | |
| m = 2 | NT | 1.23 (0.24) | 1.06 (0.21) | 1.26 (0.20) | 1.06 (0.25) | 1.17 (0.29) | 0.75 (0.25) | 1.08 (0.31) | 0.77 (0.22) |
| HT | 0.97 (0.27) | 1.10 (0.35) | 1.04 (0.25) | 1.06 (0.35) | 0.99 (0.25) | 0.81 (0.25) | 0.92 (0.23) | 0.81 (0.24) | |
| m = 3 | NT | 1.14 (0.24) | 0.96 (0.16) | 1.18 (0.22) | 0.92 (0.19) | 1.11 (0.34) | 0.64 (0.25) | 1.00 (0.42) | 0.70 (0.21) |
| HT | 0.89 (0.25) | 1.02 (0.34) | 0.97 (0.25) | 0.94 (0.34) | 0.92 (0.25) | 0.67 (0.23) | 0.88 (0.29) | 0.70 (0.25) | |
| SBP mMSE | |||||||||
| m = 1 | NT | 1.44 (0.19) | 1.26 (0.28) | 1.31 (0.18) | 1.30 (0.26) | 1.22 (0.18) | 1.03 (0.24) | 1.24 (0.24) | 0.95 (0.26) |
| HT | 1.41 (0.15) | 1.18 (0.36) | 1.30 (0.15) | 1.23 (0.37) | 1.11 (0.18) | 0.90 (0.29) | 1.19 (0.22) | 0.84 (0.25) | |
| m = 2 | NT | 1.34 (0.21) | 1.20 (0.28) | 1.23 (0.20) | 1.21 (0.25) | 1.15 (0.19) | 0.92 (0.24) | 1.20 (0.33) | 0.84 (0.25) |
| HT | 1.33 (0.17) | 1.11 (0.36) | 1.21 (0.17) | 1.12 (0.36) | 1.05 (0.19) | 0.79 (0.27) | 1.14 (0.26) | 0.74 (0.26) | |
| m = 3 | NT | 1.25 (0.19) | 1.13 (0.27) | 1.19 (0.22) | 1.11 (0.23) | 1.11 (0.23) | 0.85 (0.24) | 1.16 (0.45) | 0.78 (0.26) |
| HT | 1.21 (0.15) | 1.05 (0.35) | 1.14 (0.19) | 1.03 (0.35) | 1.00 (0.20) | 0.73 (0.27) | 1.16 (0.31) | 0.69 (0.27) | |
| DBP mMSE | |||||||||
| m = 1 | NT | 1.45 (0.25) | 1.34 (0.29) | 1.28 (0.18) | 1.33 (0.30) | 1.08 (0.22) | 1.01 (0.23) | 1.08 (0.25) | 0.95 (0.27) |
| HT | 1.32 (0.23) | 1.25 (0.30) | 1.26 (0.24) | 1.24 (0.30) | 1.08 (0.27) | 0.92 (0.21) | 1.06 (0.31) | 0.84 (0.19) | |
| m = 2 | NT | 1.38 (0.27) | 1.28 (0.31) | 1.20 (0.22) | 1.22 (0.32) | 1.00 (0.26) | 0.86 (0.27) | 1.02 (0.29) | 0.82 (0.28) |
| HT | 1.27 (0.25) | 1.20 (0.31) | 1.18 (0.27) | 1.14 (0.30) | 0.99 (0.28) | 0.79 (0.20) | 0.99 (0.31) | 0.73 (0.20) | |
| m = 3 | NT | 1.32 (0.29) | 1.21 (0.31) | 1.16 (0.26) | 1.11 (0.32) | 0.96 (0.30) | 0.77 (0.30) | 0.96 (0.33) | 0.74 (0.29) |
| HT | 1.19 (0.24) | 1.14 (0.31) | 1.10 (0.27) | 1.05 (0.28) | 0.93 (0.28) | 0.71 (0.19) | 0.97 (0.29) | 0.66 (0.18) | |
| SBP-PI mXMSE | |||||||||
| m = 1 | NT | 1.41 (0.20) | 1.33 (0.21) | 1.39 (0.12) | 1.38 (0.18) | 1.34 (0.14) | 1.13 (0.16) | 1.33 (0.14) | 1.08 (0.19) |
| HT | 1.32 (0.16) | 1.26 (0.29) | 1.31 (0.10) | 1.33 (0.30) | 1.20 (0.11) | 1.11 (0.24) | 1.25 (0.16) | 1.06 (0.26) | |
| m = 2 | NT | 1.33 (0.20) | 1.25 (0.21) | 1.32 (0.13) | 1.26 (0.16) | 1.28 (0.15) | 0.98 (0.18) | 1.25 (0.20) | 0.99 (0.21) |
| HT | 1.24 (0.18) | 1.18 (0.29) | 1.22 (0.11) | 1.21 (0.28) | 1.12 (0.14) | 0.96 (0.23) | 1.16 (0.24) | 0.97 (0.31) | |
| m = 3 | NT | 1.23 (0.18) | 1.17 (0.18) | 1.26 (0.14) | 1.14 (0.14) | 1.21 (0.15) | 0.89 (0.19) | 1.20 (0.28) | 0.94 (0.22) |
| HT | 1.14 (0.16) | 1.10 (0.29) | 1.14 (0.12) | 1.10 (0.26) | 1.06 (0.16) | 0.84 (0.20) | 1.11 (0.30) | 0.95 (0.34) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castiglioni, P.; Parati, G.; Faini, A. Information-Domain Analysis of Cardiovascular Complexity: Night and Day Modulations of Entropy and the Effects of Hypertension. Entropy 2019, 21, 550. https://doi.org/10.3390/e21060550
Castiglioni P, Parati G, Faini A. Information-Domain Analysis of Cardiovascular Complexity: Night and Day Modulations of Entropy and the Effects of Hypertension. Entropy. 2019; 21(6):550. https://doi.org/10.3390/e21060550
Chicago/Turabian StyleCastiglioni, Paolo, Gianfranco Parati, and Andrea Faini. 2019. "Information-Domain Analysis of Cardiovascular Complexity: Night and Day Modulations of Entropy and the Effects of Hypertension" Entropy 21, no. 6: 550. https://doi.org/10.3390/e21060550
APA StyleCastiglioni, P., Parati, G., & Faini, A. (2019). Information-Domain Analysis of Cardiovascular Complexity: Night and Day Modulations of Entropy and the Effects of Hypertension. Entropy, 21(6), 550. https://doi.org/10.3390/e21060550