

Entropy and Fractal Techniques for Monitoring Fish Behaviour and Welfare in Aquacultural Precision Fish Farming—A Review

Abstract
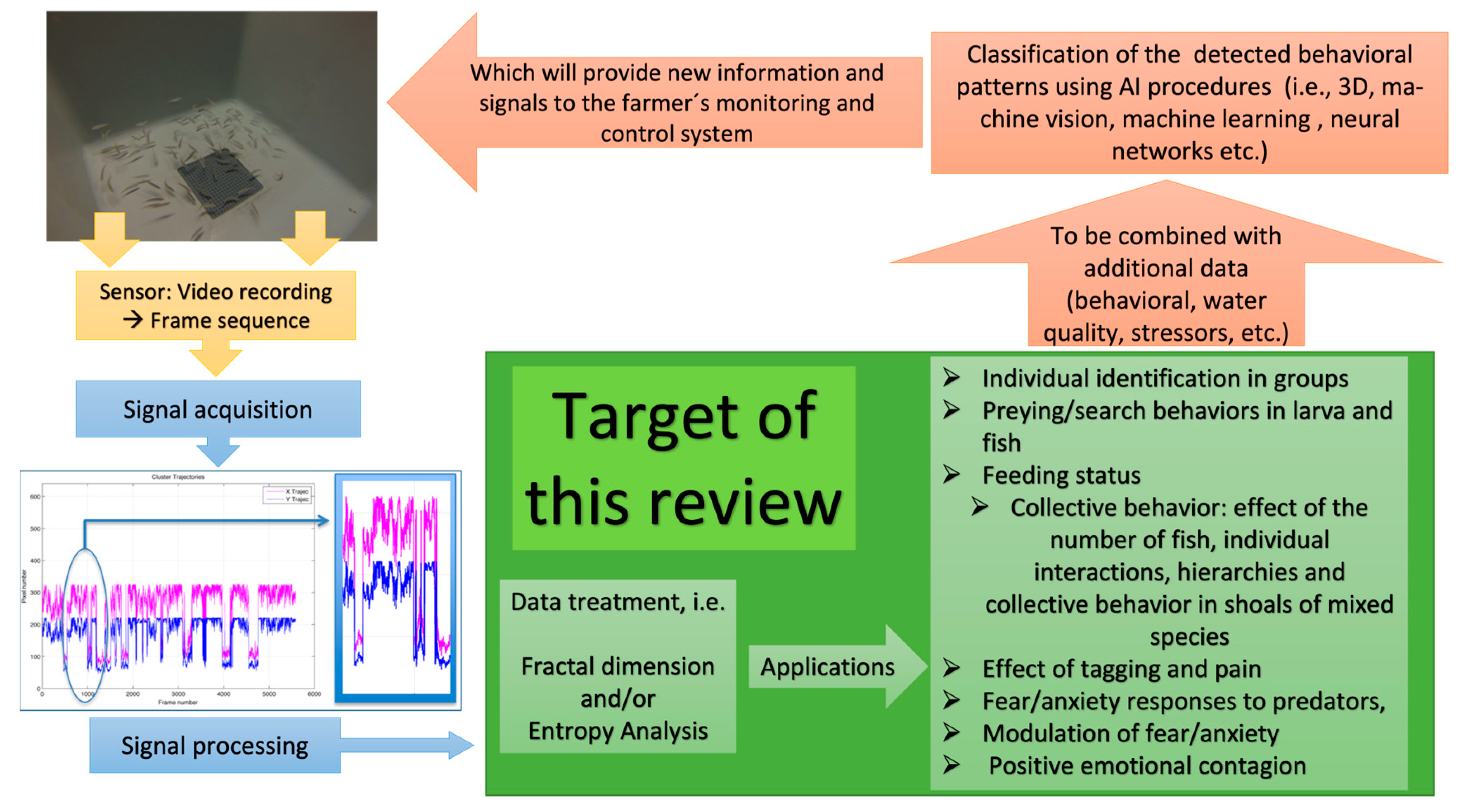
:1. Introduction
1.1. Applications of Entropy and Fractal Analyses to Fish Behaviour Studies
1.2. Aim of the Work
2. Methodology Followed for the Review
3. Targeted Applications
3.1. Individual Identification in Groups
3.2. Preying/Feeding Search Behaviour
3.2.1. Larvae
3.2.2. Fish
3.3. Feeding Status
3.4. Collective Behaviour
3.5. Effect of the Number of Fish
3.6. Collective Behaviour and Individual Interactions
3.7. Collective Behaviour and Hierarchies
3.8. Collective Behaviour and Mixed Shoal Species
3.9. Tagging and Pain
3.10. Fear/Anxiety Responses to Predators
3.11. Modulation of Fear/Anxiety Responses
3.12. Psychoactive Drugs for Anxiety Modulation
3.13. Positive Emotional Contagion
4. Prospects and Research Requirements
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Reference | Species | Methods, FD, and/or Entropy | Individual/Collective Behaviour | Main Findings |
|---|---|---|---|---|
| Individual identification in groups. | ||||
| [46] | Mature goldfish (Carassius auratus). | The mean swimming velocity and five nonlinear parameters: two measures of the FD (characteristic FD and the Richardson dimension) the Lempel–Ziv complexity, the Hurst exponent, and the degree of relative dispersion. | Individual swimming patterns in groups. | Discriminant analysis of the six measures showed that each fish displayed a very different and highly individual swimming profile, which allowed the researchers to distinguish each individual fish within groups. |
| [101] | Zebrafish (Danio rerio). | 2D video recording of the interaction between the fish and a robotic replica of 4 zebrafish by TE as affected by 3 concentrations of caffeine: 0 (control), 5, 25, and 50 mg/L. | Individual response of individually placed fish to a replica of a shoal of conspecifics and its modulation by caffeine. | The TE was always higher from the replica to the fish, but the difference was only significant in fish exposed to at least 25–50 mg caffeine/mL. |
| Preying/feeding search behaviours—Larvae | ||||
| [64] | Pink anemonefish (Amphiprion perideraion) larvae. | Fractal analysis of prey search patterns. | Individual swimming and searching behaviour in groups (up to 10 larva) prior to and after the start of feeding. | FD studies show that the larvae forage using at least one type of search behaviour for ranging and another for local searches depending on the age of the larvae and the prey’s abundance |
| [65] | Malabar grouper (Epinephelus malabaricus) larvae | 3D-recorded swimming behaviour of the larvae in presence of prey, including FD. | Early feeding: individual larval prey-searching behaviour in groups of 4 larvae. | Without prey, the FDs of the horizontal and vertical projections of larvae indicated increased activity on vertical axis. In the presence of prey, the FD showed similarly complex activity in both dimensions, indicating an optimization of the search volume and, therefore, the need to consider 3D search behaviour. |
| Preying/feeding search behaviours—Fish | ||||
| [44,45] | Computer simulation. | Chaos and fractals. | Modelling of fish schools’ chaotic movements in the presence of prey. | The motion of the fish school and its fractal properties depend on the predation rate. |
| [66] | Several—Pressure (depth)-sensitive-data-logging-tagged basking sharks (Cetorhinus maximus), small spotted catshark (Scyliorhinus canicular), bigeye tuna (Thunnus obesus), Atlantic cod (Gadus morhua), leatherback turtles (Dermochelys coriacea), penguins (Spheniscus magellanicus), and southern elephant seals (Mirounga leonine). | Simulation of searches was used to test the hypothesis that foraging success (biomass consumed per distance moved) by optimal Lévy walkers in fractal (natural) prey distributions exceeded prey acquisition rates within random prey fields. | Different marine predators’ searching behaviour in the wild. | Optimal search patterns (Lévy walks) seem to arise as a function of the underlying distribution of the prey field, i.e., the predator’s prey-searching patterns are a result of the prey distribution it encounters. |
| [67] | Zebrafish (Danio rerio). | The number of fish is not indicated. The behaviour of the groups was 2D-video-recorded, and the treated images were processed and used to create a database of different behaviours that was further used to train a VGG-16 network. Two-dimensional image entropy (calculated according to SE) was applied to describe the changes of the status of fish. Two statuses were examined: “Normal” (shoaling) and “Abnormal” (response to feeding). | Collective behaviour in a group. | The procedure can correctly classify fish into their real status. The image entropy (SE) increases when the fish transfer from a shoaling to a schooling-type of behaviour. |
| Feeding status | ||||
| [61]. Abstract in English and original paper in Chinese. | Not given. | Real-life RAS farm settings. Use of a database of recoded videos to classify the fish into 3 categories: non-eating, weakly eating, and strongly eating using a 5-layer classification neural network (CNN) optimized by a cross-entropy loss function. | Classification of fish according to three categories. It was not indicated if individual fish were classified. | There was a lack of relevant information in the English abstract, including the species used, whether the classification system applies to individuals or groups, and whether the model was tested with data different from those used to develop the CNN. |
| Collective behaviour | ||||
| [51] | Species/organisms/social bodies displaying collective behaviour. | Study of collective behaviours by applying the causal entropic (CE) framework of [69]. | To understand the origin of collective behaviour from a purely entropic point of view and make testable predictions about the fundamental form of social interactions | The CE principle gives a purely statistical prediction for many of the emergent properties of collective behaviour even in the absence of a detailed understanding of the mechanisms of interaction between individuals. |
| [70] | Nile tilapia (Oreochromis niloticus). | Video recording of a shoal and calculation of its dispersion (estimated by optical flow, entropy, and statistical parameters), velocity, and turning angle. | Shoal behaviour and welfare: construction of an efficient method to recognize special shoal behaviours and provide reliable theoretical support for online smart supervision in aquaculture, especially in RAS, to ensure fish welfare. | The proposed KEM model to detect a specific behaviour (evacuation of gastro-intestinal contents) showed good performance with respect to detecting emergent gathering and scattering behaviours of the shoal. |
| [72] | Freshwater rummy-nose tetra (Hemigrammus rhodostomus). | Video recording of the fish trajectories and assessment of the collective interactions by TE. | Collective behaviour and interactions: flow in a fish school during collective U-turn swimming changes are exposed by TE. | Two different information flows were identified: an informative flow (positive TE) from fish that have already turned to fish that are turning and a misinformative flow (negative TE) from fish that have not turned yet to fish that are turning. |
| Number of fish | ||||
| [74] | Japanese horse mackerel (Trachurus japonicas) | HD video recording using a camera that is mounted on top of a tank containing 1 to 24 fish and a simulation with robots under the same conditions. | Number of fish (1–24) and FD. Evaluation of a simulation of behaviour using robots and comparison of the fishes’ behaviour under the same conditions. Measures, among other parameters, the FD of the system. | The trajectories of the virtual robot differed from those of real fish: while the value of FD in the real fish decreases with increasing number of fish, the opposite occurred when using robots. |
| [36] | European seabass (Dicentrarchus labrax). | 2D video recording of the behaviour of fish estimated by the SE of their trajectory exposed to two variables: number of fish (n = 1 to 50) and a hit in the tank. | Individual and collective (2–50) behaviour of groups in response to two different stressors: number of fish and a hit in the tank. | The SE of the shoaling behaviour increased concomitantly with the number of fish (power function). In addition, the SE entropy increased after the hit in the tank (schooling response) for all fish groups but particularly so for individual fish and groups with only 2–5 fish. |
| Collective behaviour and individual interactions | ||||
| [75] | Medaka (Oryzias latipes). | Simple genetic algorithm, the nearest neighbour distance, polarization, the expanse, and the FD. | Individual schooling behaviour in groups of 5 fish and its mathematical modelling. | Modelling the behaviour of the medaka school using a simple genetic algorithm with the fitness defined by 4 variables: the nearest neighbour distance, polarization, the expanse, and the FD of the school’s centre of mass. |
| [43] | Computer simulation. | Order and flexibility of the school based on attraction, repulsion, and parallel orientation behaviour of individuals. | Motion of fish schools. | School’s order and flexibility are affected by the number of neighbours interacting and by the randomness of individual motion. Schooling fish have evolved specialized ability to establish both school order and flexibility at low Nb,max (=3). |
| [76] | Chicken grunt (Parapristipoma trilineatum). | Higuchi’s FD analysis of digital video images time series. | Effect of the number of fish and the presence of structures in the tank on the behaviour of the schools. | There was a difference in behaviour of schools with 1–5 fish compared to those of 10–25 fish. |
| [78] | Three-spined sticklebacks (Gasterosteus aculeatus). | Video recording of the shoaling behaviour of fish from 13 different populations and estimation of the SE and log-likelihood analysis of the population distributions. | Shoaling behaviour: Assessment of inter- and intra-populational variation in shoaling behaviour of 13 different populations of wild sticklebacks. | Traditional behavioural measures did not reveal populational differences in shoaling behaviour but SE analyses identified population-specific clustering patterns consistent with individual-specific behavioural patterns. |
| [79] | X-ray tetra (Pristella maxillaris). | Recording and comparing, via linear and non-linear (entropy) measurements, the behaviour of fish groups exposed to an alarm cue (macerated conspecifics) and to food versus the controls | Collective behaviour of groups in responses to an alarm cue and food. | Exposure to the alarm cue induced the strongest responses with wide ranging effects on the behaviour of the fish and changes in the entropy, mutual information, and entropy rate, indicating that their movements became more unpredictable after exposure to the alarm both in terms of changes in displacement and changes in velocity over short time periods. |
| [80] | Juvenile zebrafish (Danio rerio) | Groups of 60, 80, or 100 juvenile zebrafish were 2D-video-recorded and their individual position, velocity, and acceleration values were tracked (Idtracker.ai). The statistics calculated from the videos included the following parameters: distance to centre of arena, local polarization, inter-individual distance, and probability of finding another animal around a focal one. Cross-entropy loss was one of the parameters used to train the deep network when estimating the probability of a fish turning right or left. The purpose of the work was to develop a model (using deep attention networks) to understand the rules of collective behaviour and predict the sides a fish will turn toward. | Modelling individual behaviour of the collective according to interaction with conspecifics. | The network indicated that the number of interacting individuals is between 8–22, with 1–10 more important ones, particularly if some move at higher speed in front or to the sides or if they are very close or on a collision path. |
| [81] | X-ray tetras (Pristella maxillaris). | 2D video recording of the behaviour of groups of fish with different ratios of hungry/satiated fish (8/0, 6/2, 4/4, 2/6, and 0/8). Analysis of individual and group speed, group cohesion, and polarization, as well as pair-wise TE between individuals in each group. | Individual and collective behaviours in groups of 8 fish and their interactions according to the hunger status of their individual members. | The nutritional status of individuals within the groups impacts both individual and group behaviour, and members of heterogeneous groups adapt their behaviour to facilitate collective motion. |
| Collective behaviour and hierarchies | ||||
| [38] | Zebrafish (Danio rerio). | Individual trajectories of each fish in groups of 2–10 zebrafish were established by idTracking [84] and the distribution of leadership was quantified by calculating the entropy associated with the time series of all the leaders. | Collective behaviour: identification of leadership in groups of zebrafish. | Any fish could potentially lead the collective movements in the shoal. The predictor of a fish’s tendency to lead seemed to be mostly its mobility, regardless of its position in the hierarchy, although some individuals led collective movements more often than others, which, over time, may result in the development of specialized roles. |
| [85] | Ayus (Plecoglossus altivelis) and Boids (artificial life simulation) models. | 2D video recording of groups of n = 2–5 fish and analysis of Boids models to analyse the integrity of fish groups via three parameters: Mutual information (MI), TE, and integrated information theory (IIT 3.0). | Use of individual interactions and dynamics of the collective behaviour in the school to assess the integrity of the system. | IIT 3.0 identifies intrinsic differences in the behaviour of schools of 2–5 fish and a discontinuity in the group integration between systems of n = 2–3 fish and n = 4–5. For n < 4 groups, integrity requires an intact visual field. For n > 4 groups, integrity has tolerance for some blind spots. |
| Collective behaviour in mixed-species shoals | ||||
| [86] | Three-spined sticklebacks (Gasterosteus aculeatus), nine-spined sticklebacks (Pungitius pungitius), and roach (Rutilus rutilus) | Video recording of the behaviour of individuals and shoals composed of individuals from the same and different species using linear (mean of the median speeds of each fish, mean distance between all fish, and mean polarization of the group during each trial) and nonlinear (TE calculated on heading updates and differences for each pair of individuals within each group across all relevant samples) methods. | Collective behaviour and interactions in environments with mixed-species shoals. | Single-species groups were more polarized than mixed-species groups, for which there were differences between treatments in mean pairwise TE. Species-specific differences were noted in: (1) the use of information within the mixed-species groups and in (2) differences in the responses to conspecifics and heterospecifics in groups of mixed species. |
| Tagging and pain | ||||
| [23] | European seabass (Dicentrarchus labrax). | The FD (Higuchi, Katz, and Katz–Castiglioni) and Shannon and Permutation entropy of the schooling response to a percussive force applied to the tank) of groups (n = 81) of VIE-tagged and non-tagged fish. | Collective schooling behaviours of tagged/non-tagged fish. | Negligible effect of VIE-tagging on the SE and very small effect on the permutation entropy. On the other hand, the Katz–Castiglioni FD of tagged fish decreased somewhere between 4–15% depending on the window length. |
| [88] | Zebrafish (Danio rerio). | Videorecording of the individual and group behaviours exhibited by groups of 3 individuals: two non-tagged and one (focal subject) that was either non-tagged (control condition) or sham-, purple-, blue-, or yellow-tagged using traditional behavioural parameters of shoaling and schooling activities and information theoreticl measure of social interaction by TE | Effect of tagging on the individual and collective behaviour and interactions in groups of 3 fish. | Tagging did not affect the shoaling and schooling tendencies of the fish, but it significantly increased individual speed of the tagged subjects and of the group. TE. however, showed altered levels of interactions between individuals and that yellow-tagged fish were less likely to influence others. |
| [90] | Female zebrafish (Danio rerio). | Effect of fin clipping (with and without administration of the anaesthetic lidocaine), PIT tagging, and a standard pain test on the complexity of the 3D swimming patterns of individual fish estimated by their FD values. The results were compared to those of control and sham-treated zebrafish. | Effect of the selected pain treatments on the FD of 3D swimming trajectories of individual fish that were individually tested. | The FD in treated groups showed a reduced degree pf complexity in the fishes’ trajectories, while the FD of the control and sham-treated fishes did not change over time. Anesthetizing the fish with lidocaine prior to fin clipping restored the complexity of the fish behaviour to that seen in control fish. FD was useful for estimating lack of welfare/presence of pain. |
| [91] | Zebrafish (Danio rerio). | Effect of fin amputation, lidocaine treatment, and fin regeneration on the complexity of 3D swimming patterns (analysed by idTracker) of individual fish estimated via their FD and through meandering entropy analyses. In addition, principal component and hierarchical clustering analyses of collective behaviour performance were calculated. The results were compared to those of control and sham-treated zebrafish. | Effect of the selected treatments on the individual behaviour of fish individually tested (fin amputation) and collective behaviour in groups (n = 6, for lidocaine treatment and regeneration experiments). | Amputation of caudal fin resulted in more dramatic effects that fish recovered from even before full regeneration of the fin. Lidocaine treatment did not accelerate recuperation and induced minor (sedative) side-effects. |
| Fear/anxiety responses to predators | ||||
| [94] | Zebrafish (Danio rerio) and life-sized robot replica of the zebrafish. | The behaviours of a fish–conspecific and a fish–robot system were recorded by a web camera placed above the surface of the water. The SE was used to calculate the TE between the fish–conspecific and the fish–robot systems. | Testing of TE as a measure of directional information in fish–fish and fish–robot interactions. | Validation of TE as an information-theoretical measure with which to compute directional information flow in social behaviour. |
| [95] | Zebrafish (Danio rerio). | TE was used to study the relationship between fish and robots imitating these fish but differing in size. | Behavioural response of individual fish to replica of differing in size. | TE showed that the fish adjust their behaviour in response to variations in the size of the fish replicas, avoiding larger replicas and being attracted to and influenced by smaller ones. Similar-sized replica did not elicit significant responses. |
| [96] | Zebrafish (Danio rerio) as prey, robotic predators, and red tiger oscar fish (Astronotus ocellatus) as live predators. | Video recording of prey fish and their interactions with a robotic predator and with a real life predatory fish surrounding it. Analysis of causal interactions by TE | Individual behaviour. Predator–prey interactions: Testing of TE as a tool to identify and quantify causal prey–predator behavioural interactions in real fish systems. | When a living predator surrounds the prey (as in open fish farming settings), a two-way interaction is established, as shown by TE: the predator watches and responds to the behaviour of the prey, which displays an avoidance response. |
| [97] | Prey (rosy bitterling, Rhodeus ocellatus) and its predator (northern snakehead, Channa argus). | Videorecording of the behaviour of the prey and the predator as they interact when the prey is placed in a concentric tank surrounded by a larger tank containing the predator. The fish can interact with each other visually and by feeling the ripples in the water produced by the swimming movements of the other. | Use of TE to acquire information on the interactions between the two individual fish. | The prey’s TE was generally significantly greater than the predator’s (i.e., information flows from the predator to the prey) regardless of the set of coarse-grained parameters chosen, indicating that the prey was more vigilant with respect to the predator’s position than vice versa. The prey was also vigilant while the predator moved aimlessly. |
| [98] | Male and female zebrafish (Danio rerio) acting as prey and a 3D-printed robotic replica of its allopatric predator the red tiger oscar fish (Astronotus ocellatus). | Videorecording of the motion of the prey fish in the vertical and horizontal axes in response to the movements of the replica predator controlled under closed- and open-loop conditions. The fear response to the predator is computed by two avoidance-related parameters: (1) the average distance between the replica and the fish and (2) the time spent by the focal fish in the half of the water column opposite to that occupied by the replica. | Individual behaviour of individual fish in response to a replica predator. TE quantifies the response of the prey in terms of “stationary”, “swimming”, and “attacking” motion patterns of the predator. | The zebrafish quickly adjusted their behaviour to avoid the predator’s attacks. TE revealed that the state of the robot affected the vertical position of the fish only in the closed-loop control condition and not its position in the horizontal axis. |
| Modulation of fear/anxiety | ||||
| [99] | Zebrafish (Danio rerio). Data from [100]. | Data from [100]. | Modelling of geotaxis by using spatial entropy (SE) to identify the extent of the volume of water occupied by a fish and in which part of the tank most of the activity takes place. | Mathematical model to quantify geotaxis. |
| [101] | Zebrafish (Danio rerio). | 2D videorecording of the interaction between the fish and a robotic replica of 4 zebrafish according to TE as affected by 3 concentrations of caffeine: 5, 25, and 50 mg/L and compared to an untreated control | Individual response of individually placed fish to a replica of a shoal of conspecifics and its modulation by caffeine. | The TE was always higher from the replica to the fish but only in fish exposed to at least 25–50 mg caffeine/mL the difference was significant. |
| [102] | Zebrafish (Danio rerio) and a fear-eliciting stimulus consisting of three zebrafish replicas with a synchronized and polarized motion moving in 3D trajectories via a robotic platform. | 3D tracking of zebrafish responses in the presence and absence of ethanol (0, control group, and two test groups with 0.25% and 1.00% concentrations of ethanol/water), which is known to interfere with fear conditioning in different species. | Modulation of conditioned fear-responses. Individual responses. The parameters assessed were avoidance index, geotaxis, freezing, spatial entropy (SE), and average speed, acceleration, and angular speed. | Ethanol significantly lowered the intensity of anxiety-related behaviours (i.e., spatial avoidance and geotaxis). The effects on spatial entropy (SE) and average speed and acceleration were marginal, time-dependent, and only present at the highest dose (1.00%). There was no effect on freezing or average angular speed. |
| [103] | Zebrafish (Danio rerio) and a robotic stimulus mimicking a sympatric predator of zebrafish (Indian pond heron, Ardeola grayii) programmed via a microcontroller that regularly struck the water’s surface in a lateral compartment. | 3D tracking of zebrafish behaviour in the presence and absence of two anxiolytic compounds. The experimental groups included: drug-free control groups and those treated with three different concentrations of citalopram (30, 50, and 100 mg/L) and ethanol (0.25%, 0.50%, and 1.00%). | Modulation of fear/anxiety. Individual responses. The parameters assessed were avoidance index, geotaxis, freezing, spatial entropy (SE), and average speed, acceleration, and angular speed. | In the absence of the conditioning stimulus, zebrafish displayed a conditioned geotaxis that was reduced in a linear dose–response-oriented manner by citalopram and in a U-shaped dose–response manner by ethanol. |
| Positive emotional contagion | ||||
| [106] | Zebrafish (Danio rerio). | 2D recording of zebrafish administered a solution of citalopram of 0 (control), 30, and 100 mg/L. The fish were tested individually and in groups of n = 5, where only 1 of them had been citalopram-treated. | Testing of positive emotional contagion by analysing geotaxis, group cohesion, coordination, and causal interactions (by TE) for individuals (individually assessed) and for groups. | Group cohesion and coordination were not affected by the treatment. Changes in geotaxis were consistent with alleviation of anxiety in citalopram-treated individuals and in groups where only one member was treated. TE indicated that emotional contagion was directional: the treated individual influenced untreated fish, but not vice versa |
| Recognition of abnormal behaviour | ||||
| [37] | Species not declared | Video recording under real farming conditions and development of a method to detect abnormal behaviours. | Detection of abnormal behaviour in real-life situation on a farm. | Softmax cross-entropy loss and weight attenuation L2 regularization were used to optimize a convolution 3D model, which, consequently, proved to be able to successfully analyse real-life aquaculture videos and detect abnormal behaviours. |
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- EATiP. A Review of the Strategic Research and Innovation Agenda. In Our Vision for the Future of European Aquaculture; European Aquaculture Technology and Innovation Platform: Liege, Belgium, 2017; Available online: https://eatip.eu/wp-content/uploads/2018/02/EATIP-SRIA-2017.pdf (accessed on 21 March 2023).
- Eguiraun, H.; Izagirre, U.; Martinez, I. A Paradigm Shift in Safe Seafood Production: From Contaminant Detection to Fish Monitoring—Application of Biological Warning Systems to Aquaculture. Trends Food Sci. Technol. 2015, 43, 104–113. [Google Scholar] [CrossRef]
- EC. A Sustainable Food System for the European Union; 2020; ISBN 978-3-9820301-3-5. Available online: https://www.sapea.info/wp-content/uploads/sustainable-food-system-report.pdf (accessed on 21 March 2023).
- EU. Identification of Emerging Technologies and Breakthrough; 2022; ISBN 9789294691224. Available online: https://eic.ec.europa.eu/system/files/2022-02/EIC-Emerging-Tech-and-Breakthrough-Innov-report-2022-1502-final.pdf (accessed on 21 March 2023).
- Føre, M.; Frank, K.; Norton, T.; Svendsen, E.; Alfredsen, J.A.; Dempster, T.; Eguiraun, H.; Watson, W.; Stahl, A.; Sunde, L.M.; et al. Precision Fish Farming: A New Framework to Improve Production in Aquaculture. Biosyst. Eng. 2018, 173, 176–193. [Google Scholar] [CrossRef]
- Li, D.; Wang, G.; Du, L.; Zheng, Y.; Wang, Z. Recent Advances in Intelligent Recognition Methods for Fish Stress Behavior. Aquac. Eng. 2022, 96, 102222. [Google Scholar] [CrossRef]
- Marques, T.A.; Thomas, L.; Martin, S.W.; Mellinger, D.K.; Ward, J.A.; Moretti, D.J.; Harris, D.; Tyack, P.L. Estimating Animal Population Density Using Passive Acoustics. Biol. Rev. 2013, 88, 287–309. [Google Scholar] [CrossRef] [Green Version]
- Føre, M.; Svendsen, E.; Alfredsen, J.A.; Uglem, I.; Bloecher, N.; Sveier, H.; Sunde, L.M.; Frank, K. Using Acoustic Telemetry to Monitor the Effects of Crowding and Delousing Procedures on Farmed Atlantic Salmon (Salmo salar). Aquaculture 2018, 495, 757–765. [Google Scholar] [CrossRef]
- Rose, C.S.; Stoner, A.W.; Matteson, K. Use of High-Frequency Imaging Sonar to Observe Fish Behaviour near Baited Fishing Gears. Fish Res. 2005, 76, 291–304. [Google Scholar] [CrossRef]
- Gesto, M.; Zupa, W.; Alfonso, S.; Spedicato, M.T.; Lembo, G.; Carbonara, P. Using Acoustic Telemetry to Assess Behavioral Responses to Acute Hypoxia and Ammonia Exposure in Farmed Rainbow Trout of Different Competitive Ability. Appl. Anim. Behav. Sci. 2020, 230, 105084. [Google Scholar] [CrossRef]
- Halvorsen, M.B.; Zeddies, D.G.; Ellison, W.T.; Chicoine, D.R.; Popper, A.N. Effects of Mid-Frequency Active Sonar on Hearing in Fish. J. Acoust. Soc. Am. 2012, 131, 599–607. [Google Scholar] [CrossRef]
- Kruusmaa, M.; Gkliva, R.; Tuhtan, J.A.; Tuvikene, A.; Alfredsen, J.A. Salmon Behavioural Response to Robots in an Aquaculture Sea Cage. R Soc. Open Sci. 2020, 7, 191220. [Google Scholar] [CrossRef] [Green Version]
- Kruusmaa, M.; Rieucau, G.; Montoya, J.C.C.; Markna, R.; Handegard, N.O. Collective Responses of a Large Mackerel School Depend on the Size and Speed of a Robotic Fish but Not on Tail Motion. Bioinspir. Biomim. 2016, 11, 56020. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, S.; Liu, J.; Wang, H.; Zhu, J.; Li, D.; Zhao, R. Application of Machine Learning in Intelligent Fish Aquaculture: A Review. Aquaculture 2021, 540, 736724. [Google Scholar] [CrossRef]
- Gladju, J.; Kamalam, B.S.; Kanagaraj, A. Applications of Data Mining and Machine Learning Framework in Aquaculture and Fisheries: A Review. Smart Agric. Technol. 2022, 2, 100061. [Google Scholar] [CrossRef]
- Saberioon, M.; Gholizadeh, A.; Cisar, P.; Pautsina, A.; Urban, J. Application of Machine Vision Systems in Aquaculture with Emphasis on Fish: State-of-the-Art and Key Issues. Rev. Aquac. 2017, 9, 369–387. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Yu, H.; Fang, X.; Song, L.; Li, D.; Chen, Y. Computer Vision Models in Intelligent Aquaculture with Emphasis on Fish Detection and Behavior Analysis: A Review. Arch. Comput. Methods Eng. 2021, 28, 2785–2816. [Google Scholar] [CrossRef]
- Wei, Y.; Wei, Q.; An, D. Intelligent Monitoring and Control Technologies of Open Sea Cage Culture: A Review. Comput. Electron. Agric. 2020, 169, 105119. [Google Scholar] [CrossRef]
- Andrady, A.L. Microplastics in the Marine Environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef]
- Brausch, J.M.; Connors, K.A.; Brooks, B.W.; Rand, G.M. Human Pharmaceuticals in the Aquatic Environment: A Review of Recent Toxicological Studies and Considerations for Toxicity Testing. Rev. Environ. Contam. Toxicol. 2012, 218, 1–99. [Google Scholar] [CrossRef]
- Brodin, T.; Piovano, S.; Fick, J.; Klaminder, J.; Heynen, M.; Jonsson, M. Ecological Effects of Pharmaceuticals in Aquatic Systems—Impacts through Behavioural Alterations. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguiraun, H.; López-de-Ipiña, K.; Martinez, I. Application of Entropy and Fractal Dimension Analyses to the Pattern Recognition of Contaminated Fish Responses in Aquaculture. Entropy 2014, 16, 6133–6151. [Google Scholar] [CrossRef] [Green Version]
- Gavrilescu, M.; Demnerová, K.; Aamand, J.; Agathos, S.; Fava, F. Emerging Pollutants in the Environment: Present and Future Challenges in Biomonitoring, Ecological Risks and Bioremediation. N. Biotechnol. 2015, 32, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Herbert-Read, J.E.; Kremer, L.; Bruintjes, R.; Radford, A.N.; Ioannou, C.C. Anthropogenic Noise Pollution from Pile-Driving Disrupts the Structure and Dynamics of Fish Shoals; 2017; Volume 284, ISBN 0000000154703. Available online: https://research-information.bris.ac.uk/ws/portalfiles/portal/134650896/20171627.full.pdf (accessed on 21 March 2023).
- Jacquin, L.; Petitjean, Q.; Côte, J.; Laffaille, P.; Jean, S. Effects of Pollution on Fish Behavior, Personality, and Cognition: Some Research Perspectives. Front. Ecol. Evol. 2020, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Llorca, M.; Farré, M. Current Insights into Potential Effects of Micro-Nanoplastics on Human Health by in-Vitro Tests. Front. Toxicol. 2021, 3, 752140. [Google Scholar] [CrossRef] [PubMed]
- Mattsson, K.; Johnson, E.V.; Malmendal, A.; Linse, S.; Hansson, L.A.; Cedervall, T. Brain Damage and Behavioural Disorders in Fish Induced by Plastic Nanoparticles Delivered through the Food Chain. Sci. Rep. 2017, 7, 11452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoye, C.O.; Addey, C.I.; Oderinde, O.; Okoro, J.O.; Uwamungu, J.Y.; Ikechukwu, C.K.; Okeke, E.S.; Ejeromedoghene, O.; Odii, E.C. Toxic Chemicals and Persistent Organic Pollutants Associated with Micro-and Nanoplastics Pollution. Chem. Eng. J. Adv. 2022, 11, 100310. [Google Scholar] [CrossRef]
- Suryanto, M.E.; Yang, C.-C.; Audira, G.; Vasquez, R.D.; Roldan, M.J.M.; Ger, T.; Hsiao, C. Evaluation of Locomotion Complexity in Zebrafish after Exposure to Twenty Antibiotics by Fractal Dimension and Entropy Analysis. Antibiotics 2022, 11, 1059. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhu, J.; Wang, A.; Wang, J.; Wu, Y.; Yao, W. Early Detection of Cyanide, Organophosphate and Rodenticide Pollution Based on Locomotor Activity of Zebrafish Larvae. PeerJ 2021, 9, e12703. [Google Scholar] [CrossRef]
- Weilgart, L.S. The Impacts of Anthropogenic Ocean Noise on Cetaceans and Implications for Management. Can. J. Zool. 2007, 85, 1091–1116. [Google Scholar] [CrossRef]
- Bae, M.J.; Park, Y.S. Biological Early Warning System Based on the Responses of Aquatic Organisms to Disturbances: A Review. Sci. Total Environ. 2014, 466–467, 635–649. [Google Scholar] [CrossRef]
- Brodin, T.; Fick, J.; Jonsson, M.; Klaminder, J. Dilute Concentrations of a Psychiatric Drug Alter Behavior of Fish from Natural Populations. Science (1979) 2013, 339, 814–815. [Google Scholar] [CrossRef]
- Eguiraun, H.; Casquero, O.; Sørensen, A.J.; Martinez, I. Reducing the Number of Individuals to Monitor Shoaling Fish Systems—Application of the Shannon Entropy to Construct a Biological Warning System Model. Front. Physiol. 2018, 9, 493. [Google Scholar] [CrossRef] [Green Version]
- Chase, I.D.; Tovey, C.; Spangler-Martin, D.; Manfredonia, M. Individual Differences versus Social Dynamics in the Formation of Animal Dominance Hierarchies. Proc. Natl. Acad. Sci. USA 2002, 99, 5744–5749. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Li, X.H.; Xie, X.Y.; Zhao, Y.C. Abnormal Behavior Recognition of Underwater Fish Body Based on C3D Model. In Proceedings of the 6th International Conference on Machine Learning and Soft Computing, Haikou, China, 15–17 January 2022; pp. 92–97. Available online: https://dl.acm.org/doi/pdf/10.1145/3523150.3523165 (accessed on 21 March 2023). [CrossRef]
- Collignon, B.; Séguret, A.; Chemtob, Y.; Cazenille, L.; Halloy, J. Collective Departures and Leadership in Zebrafish. PLoS ONE 2019, 14, e0216798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komareji, M.; Bouffanais, R. Resilience and Controllability of Dynamic Collective Behaviors. PLoS ONE 2013, 8, e82578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, S.D.; Radford, A.N.; Nedelec, S.L.; Ferrari, M.C.O.; Chivers, D.P.; McCormick, M.I.; Meekan, M.G. Anthropogenic Noise Increases Fish Mortality by Predation. Nat. Commun. 2016, 7, 10544. [Google Scholar] [CrossRef] [Green Version]
- Mwaffo, V.; Butail, S.; Porfiri, M. Analysis of Pairwise Interactions in a Maximum Likelihood Sense to Identify Leaders in a Group. Front. Robot. AI 2017, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, S.; Harcourt, J.L.; Johnstone, R.A.; Manica, A. Initiative, Personality and Leadership in Pairs of Foraging Fish. PLoS ONE 2012, 7, e36606. [Google Scholar] [CrossRef]
- Inada, Y.; Kawachi, K. Order and Flexibility in the Motion of Fish Schools. J. Theor. Biol. 2002, 214, 371–387. [Google Scholar] [CrossRef] [Green Version]
- Tikhonov, D.A.; Enderlein, J.; Malchow, H.; Medvinsky, A.B. Chaos and Fractals in Fish School Motion. Chaos Solitons Fractals 2001, 12, 277–288. [Google Scholar] [CrossRef]
- Tikhonov, D.A.; Malchow, H. Chaos and Fractals in Fish School Motion, II. Chaos Solitons Fractals 2003, 16, 287–289. [Google Scholar] [CrossRef]
- Neumeister, H.; Cellucci, C.J.; Rapp, P.E.; Korn, H.; Faber, D.S. Dynamical Analysis Reveals Individuality of Locomotion in Goldfish. J. Exp. Biol. 2004, 207, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Mandelbrot, B.B. The Fractal Geometry of Nature; W.H. Freeman: San Francisco, CA, USA, 1982; ISBN 978-0716711865. [Google Scholar]
- Zmeskal, O.; Dzik, P.; Vesely, M. Entropy of Fractal Systems. Comput. Math. Appl. 2013, 66, 135–146. [Google Scholar] [CrossRef]
- Schreiber, T. Measuring Information Transfer. Phys. Rev. Lett. 2000, 85, 461–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butail, S.; Mwaffo, V.; Porfiri, M. Model-Free Information-Theoretic Approach to Infer Leadership in Pairs of Zebrafish. Phys. Rev. E 2016, 93, 042411. [Google Scholar] [CrossRef] [Green Version]
- Mann, R.P.; Garnett, R. The Entropic Basis of Collective Behaviour. J. R. Soc. Interface 2015, 12, 20150037. [Google Scholar] [CrossRef]
- Kitano, H. Computational Systems Biology. Nature 2002, 420, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Spasic, S.; Savic, A.; Nikolic, L.; Budimir, S.; Janosevic, D.; Mitrovic, A. Applications of Higuchi’s Fractal Dimension in the Analysis of Biological Signals. In Proceedings of the 2012 20th Telecommunications Forum, TELFOR 2012—Proceedings, Belgrade, Serbia, 20–22 November 2012; pp. 639–641. [Google Scholar] [CrossRef]
- Costa, M.; Goldberger, A.L.; Peng, C.K. Multiscale Entropy Analysis of Biological Signals. Phys. Rev. E Stat. Nonlin. Soft Matter. Phys. 2005, 71, 021906. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Mandelbrot, B. How Long Is the Coast of Britain? Statistical Self-Similarity and Fractional Dimension. Science (1979) 1967, 156, 636–638. [Google Scholar] [CrossRef] [Green Version]
- Kith, K.; Sourina, O.; Kulish, V.; Khoa, N.M. An Algorithm for Fractal Dimension Calculation Based on Renyi Entropy for Short Time Signal Analysis. In Proceedings of the ICICS 2009—Conference Proceedings of the 7th International Conference on Information, Communications and Signal Processing, Macau, China, 8–10 December 2009; pp. 1–5. [Google Scholar] [CrossRef]
- Ezeiza, A.; López de Ipiña, K.; Hernández, C.; Barroso, N. Enhancing the Feature Extraction Process for Automatic Speech Recognition with Fractal Dimensions. Cognit. Comput. 2013, 5, 545–550. [Google Scholar] [CrossRef]
- Sekine, M.; Tamura, T.; Akay, M.; Fujimoto, T.; Togawa, T.; Fukui, Y. Discrimination of Walking Patterns Using Wavelet-Based Fractal Analysis. IEEE Trans. Neural Syst. Rehabil. Eng. 2002, 10, 188–196. [Google Scholar] [CrossRef]
- Alados, C.L.; Escos, J.M.; Emlen, J.M. Fractal Structure of Sequential Behaviour Patterns: An Indicator of Stress. Anim. Behav. 1996, 51, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Lihong, X.; Shijing, L. Fine-Grained Classification Algorithm of Fish Feeding State Based on Optical Flow Method. Nongye Gongcheng Xuebao/Trans. Chin. Soc. Agric. Eng. 2021, 37, 238–244. [Google Scholar] [CrossRef]
- Iqbal, U.; Li, D.; Akhter, M. Intelligent Diagnosis of Fish Behavior Using Deep Learning Method. Fishes 2022, 7, 201. [Google Scholar] [CrossRef]
- Kong, Q.; Du, R.; Duan, Q.; Zhang, Y.; Chen, Y.; Li, D.; Xu, C.; Li, W.; Liu, C. A Recurrent Network Based on Active Learning for the Assessment of Fish Feeding Status. Comput. Electron. Agric. 2022, 198, 106979. [Google Scholar] [CrossRef]
- Coughlin, D.J.; Strickler, J.R.; Sanderson, B. Swimming and Search Behaviour in Clownfish, Amphiprion perideraion, Larvae. Anim. Behav. 1992, 44, 427–440. [Google Scholar] [CrossRef]
- Mahjoub, M.S.; Souissi, S.; Schmitt, F.G.; Nan, F.H.; Hwang, J.S. Anisotropy and Shift of Search Behavior in Malabar Grouper (Epinephelus malabaricus) Larvae in Response to Prey Availability. Hydrobiologia 2011, 666, 215–222. [Google Scholar] [CrossRef]
- Sims, D.W.; Southall, E.J.; Humphries, N.E.; Hays, G.C.; Bradshaw, C.J.A.; Pitchford, J.W.; James, A.; Ahmed, M.Z.; Brierley, A.S.; Hindell, M.A.; et al. Scaling Laws of Marine Predator Search Behaviour. Nature 2008, 451, 1098–1102. [Google Scholar] [CrossRef]
- Zhang, B.; Xie, F.; Han, F. Fish Population Status Detection Based on Deep Learning System. In Proceedings of the 2019 IEEE International Conference on Mechatronics and Automation, ICMA 2019, Tianjin, China, 4–7 August 2019; pp. 81–85. [Google Scholar] [CrossRef]
- Surowiecki, J. The Wisdom Of Crowds: Why the Many Are Smarter than the Few and How Collective Wisdom Shapes Business, Economics, Society and Nations; 1st Abacus Edition; Little Brown: Boston, MA, USA, 2004; ISBN 978-0349116051. [Google Scholar]
- Wissner-Gross, A.D.; Freer, C.E. Causal Entropic Forces. Phys. Rev. Lett. 2013, 110, 168702. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Gu, Z.; Shi, M.; Lu, H.; Li, J.; Shen, M.; Ye, Z.; Zhu, S. Spatial Behavioral Characteristics and Statistics-Based Kinetic Energy Modeling in Special Behaviors Detection of a Shoal of Fish in a Recirculating Aquaculture System. Comput. Electron. Agric. 2016, 127, 271–280. [Google Scholar] [CrossRef]
- Tunstrøm, K.; Katz, Y.; Ioannou, C.C.; Huepe, C.; Lutz, M.J.; Couzin, I.D. Collective States, Multistability and Transitional Behavior in Schooling Fish. PLoS Comput. Biol. 2013, 9, e1002915. [Google Scholar] [CrossRef] [Green Version]
- Crosato, E.; Jiang, L.; Lecheval, V.; Lizier, J.T.; Wang, X.R.; Tichit, P.; Theraulaz, G.; Prokopenko, M. Informative and Misinformative Interactions in a School of Fish. Swarm Intell. 2018, 12, 283–305. [Google Scholar] [CrossRef] [Green Version]
- Calovi, D.S.; Lopez, U.; Schuhmacher, P.; Chaté, H.; Sire, C.; Theraulaz, G. Collective Response to Perturbations in a Data-Driven Fish School Model. J. R. Soc. Interface 2015, 12, 20141362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushida, K.; Fukuda, K.; Sugino, R.; Morizumi, N. Computational Algorithm for Fish Schooling Using Bitmap Image. In Proceedings of the SICE Annual Conference, Nagoya, Japan, 14–17 September 2013; pp. 937–942. [Google Scholar]
- Hiramatsu, K.; Shikasho, S.; Mori, K. Mathematical Modeling of Fish Schooling of Japanese Medaka Using Basic Behavioral Patterns. J. Fac. Agric. Kyushu Univ. 2000, 45, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Takagi, T.; Hiraishi, T. Video Analysis of Fish Schooling Behavior in Finite Space Using a Mathematical Model. Fish Res. 2003, 60, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, T. Approach to an Irregular Time Series on the Basis of the Fractal Theory. Physica D 1988, 31, 277–283. [Google Scholar] [CrossRef]
- Wark, A.R.; Wark, B.J.; Lageson, T.J.; Peichel, C.L. Novel Methods for Discriminating Behavioral Differences between Stickleback Individuals and Populations in a Laboratory Shoaling Assay. Behav. Ecol. Sociobiol. 2011, 65, 1147–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaerf, T.M.; Dillingham, P.W.; Ward, A.J.W. The Effects of External Cues on Individual and Collective Behavior of Shoaling Fish. Sci. Adv. 2017, 7, e1603201. [Google Scholar] [CrossRef] [Green Version]
- Heras, F.J.H.; Romero-Ferrero, F.; Hinz, R.C.; De Polavieja, G.G. Deep Attention Networks Reveal the Rules of Collective Motion in Zebrafish. PLoS Comput. Biol. 2019, 15, e1007354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.D.M.; Burns, A.L.J.; Crosato, E.; Lizier, J.; Prokopenko, M.; Schaerf, T.M.; Ward, A.J.W. Conformity in the Collective: Differences in Hunger Affect Individual and Group Behavior in a Shoaling Fish. Behav. Ecol. 2019, 30, 968–974. [Google Scholar] [CrossRef]
- King, A.J.; Johnson, D.D.P.; Van Vugt, M. The Origins and Evolution of Leadership. Curr. Biol. 2009, 19, R911–R916. [Google Scholar] [CrossRef] [Green Version]
- Petit, O.; Bon, R. Decision-Making Processes: The Case of Collective Movements. Behav. Process. 2010, 84, 635–647. [Google Scholar] [CrossRef]
- Pérez-Escudero, A.; Vicente-Page, J.; Hinz, R.C.; Arganda, S.; De Polavieja, G.G. IdTracker: Tracking Individuals in a Group by Automatic Identification of Unmarked Animals. Nat. Methods 2014, 11, 743–748. [Google Scholar] [CrossRef]
- Niizato, T.; Sakamoto, K.; Mototake, Y.I.; Murakami, H.; Tomaru, T.; Hoshika, T.; Fukushima, T. Finding Continuity and Discontinuity in Fish Schools via Integrated Information Theory. PLoS ONE 2020, 15, e0229573. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.J.W.; Schaerf, T.M.; Burns, A.L.J.; Lizier, J.T.; Crosato, E.; Prokopenko, M.; Webster, M.M. Cohesion, Order and Information Flow in the Collective Motion of Mixed-Species Shoals. R. Soc. Open Sci. 2018, 5, 181132. [Google Scholar] [CrossRef] [Green Version]
- Brennan, N.P.; Leber, K.M.; Blankenship, H.L.; Ransier, J.M.; DeBruler, R. An Evaluation of Coded Wire and Elastomer Tag Performance in Juvenile Common Snook under Field and Laboratory Conditions. N. Am. J. Fish Manag. 2005, 25, 437–445. [Google Scholar] [CrossRef]
- Ruberto, T.; Clément, R.J.G.; Spinello, C.; Neri, D.; MacRì, S.; Porfiri, M. The Tagging Procedure of Visible Implant Elastomers Influences Zebrafish Individual and Social Behavior. Zebrafish 2018, 15, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Frommen, J.G.; Hanak, S.; Schmidl, C.A.; Thünken, T. Visible Implant Elastomer Tagging Influences Social Preferences of Zebrafish (Danio rerio). Behaviour 2015, 152, 1765–1777. [Google Scholar] [CrossRef] [Green Version]
- Deakin, A.G.; Spencer, J.W.; Cossins, A.R.; Young, I.S.; Sneddon, L.U. Welfare Challenges Influence the Complexity of Movement: Fractal Analysis of Behaviour in Zebrafish. Fishes 2019, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Audira, G.; Suryanto, M.E.; Chen, K.H.C.; Vasquez, R.D.; Roldan, M.J.M.; Yang, C.C.; Der Hsiao, C.; Huang, J.C. Acute and Chronic Effects of Fin Amputation on Behavior Performance of Adult Zebrafish in 3D Locomotion Test Assessed with Fractal Dimension and Entropy Analyses and Their Relationship to Fin Regeneration. Biology 2022, 11, 969. [Google Scholar] [CrossRef]
- Zhang, Z.; Fu, Y.; Zhang, Z.; Zhang, X.; Chen, S. A Comparative Study on Two Territorial Fishes: The Influence of Physical Enrichment on Aggressive Behavior. Animals 2021, 11, 1868. [Google Scholar] [CrossRef]
- Näslund, J.; Rosengren, M.; Del Villar, D.; Gansel, L.; Norrgård, J.R.; Persson, L.; Winkowski, J.J.; Kvingedal, E. Hatchery Tank Enrichment Affects Cortisol Levels and Shelter-Seeking in Atlantic Salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2013, 70, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Butail, S.; Ladu, F.; Spinello, D.; Porfiri, M. Information Flow in Animal-Robot Interactions. Entropy 2014, 16, 1315–1330. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, T.; Mwaffo, V.; Showler, A.; Macrì, S.; Butail, S.; Porfiri, M. Zebrafish Response to 3D Printed Shoals of Conspecifics: The Effect of Body Size. Bioinspir. Biomim. 2016, 11, 026003. [Google Scholar] [CrossRef]
- Neri, D.; Ruberto, T.; Cord-Cruz, G.; Porfiri, M. Information Theory and Robotics Meet to Study Predator-Prey Interactions. Chaos Interdiscip. J. Nonlinear Sci. 2017, 27, 073111. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Nie, L.J.; Fu, S.J. Information Dynamics in the Interaction between a Prey and a Predator Fish. Entropy 2015, 17, 7230–7241. [Google Scholar] [CrossRef] [Green Version]
- Spinello, C.; Yang, Y.; Macrì, S.; Porfiri, M. Zebrafish Adjust Their Behavior in Response to an Interactive Robotic Predator. Front. Robot. AI 2019, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Burbano, D.A.L.; Porfiri, M. Modeling Zebrafish Geotaxis as a Feedback Control Process. In Proceedings of the American Control Conference 2021, New Orleans, LA, USA, 25–28 May 2021; pp. 660–665. [Google Scholar] [CrossRef]
- Macrì, S.; Clément, R.J.G.; Spinello, C.; Porfiri, M. Comparison between Two- and Three-Dimensional Scoring of Zebrafish Response to Psychoactive Drugs: Identifying When Three-Dimensional Analysis Is Needed. PeerJ 2019, 7, e7893. [Google Scholar] [CrossRef]
- Ladu, F.; Mwaffo, V.; Li, J.; Macrì, S.; Porfiri, M. Acute Caffeine Administration Affects Zebrafish Response to a Robotic Stimulus. Behav. Brain Res. 2015, 289, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Macrì, S.; Karakaya, M.; Spinello, C.; Porfiri, M. Zebrafish Exhibit Associative Learning for an Aversive Robotic Stimulus. Lab. Anim. 2020, 49, 259–264. [Google Scholar] [CrossRef]
- Clément, R.J.G.; Macrì, S.; Porfiri, M. Design and Development of a Robotic Predator as a Stimulus in Conditioned Place Aversion for the Study of the Effect of Ethanol and Citalopram in Zebrafish. Behav. Brain Res. 2020, 378, 1–10. [Google Scholar] [CrossRef]
- Cianca, V.; Bartolini, T.; Porfiri, M.; Macrì, S. A Robotics-Based Behavioral Paradigm to Measure Anxiety-Related Responses in Zebrafish. PLoS ONE 2013, 8, e69661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahashi, W.; Ohtsuki, H. When Is Emotional Contagion Adaptive? J. Theor. Biol. 2015, 380, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Burbano Lombana, D.A.; Macrì, S.; Porfiri, M. Collective Emotional Contagion in Zebrafish. Front. Behav. Neurosci. 2021, 15, 730372. [Google Scholar] [CrossRef] [PubMed]
| Queries | Documents |
|---|---|
| (TITLE-ABS-KEY (fish AND behav*) AND TITLE-ABS-KEY (fractal* OR entropy)) | 143 |
| (TITLE-ABS-KEY (aquacult*) AND TITLE-ABS-KEY (fractal* OR entropy)) | 87 |
| (TITLE-ABS-KEY ("Fish behavio*") AND TITLE-ABS-KEY (entropy)) | 11 |
| (TITLE-ABS-KEY ("Fish behavio*") AND TITLE-ABS-KEY (fractal)) | 9 |
| (TITLE-ABS-KEY ("collective behaviour" OR "collective behavior") AND TITLE-ABS-KEY (fish) AND TITLE-ABS-KEY (welfare OR stress* OR health OR disease)) | 23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eguiraun, H.; Martinez, I. Entropy and Fractal Techniques for Monitoring Fish Behaviour and Welfare in Aquacultural Precision Fish Farming—A Review. Entropy 2023, 25, 559. https://doi.org/10.3390/e25040559
Eguiraun H, Martinez I. Entropy and Fractal Techniques for Monitoring Fish Behaviour and Welfare in Aquacultural Precision Fish Farming—A Review. Entropy. 2023; 25(4):559. https://doi.org/10.3390/e25040559
Chicago/Turabian StyleEguiraun, Harkaitz, and Iciar Martinez. 2023. "Entropy and Fractal Techniques for Monitoring Fish Behaviour and Welfare in Aquacultural Precision Fish Farming—A Review" Entropy 25, no. 4: 559. https://doi.org/10.3390/e25040559