
Sleep Spindle-Related EEG Connectivity in Children with Attention-Deficit/Hyperactivity Disorder: An Exploratory Study

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. EEG Recordings and Pre-Processing
2.3. Generalized Partial Direct Coherence (gPDC)
2.4. Statistics
3. Results
3.1. Number of Connections
3.2. Nodes Degrees and Principal Component Analysis
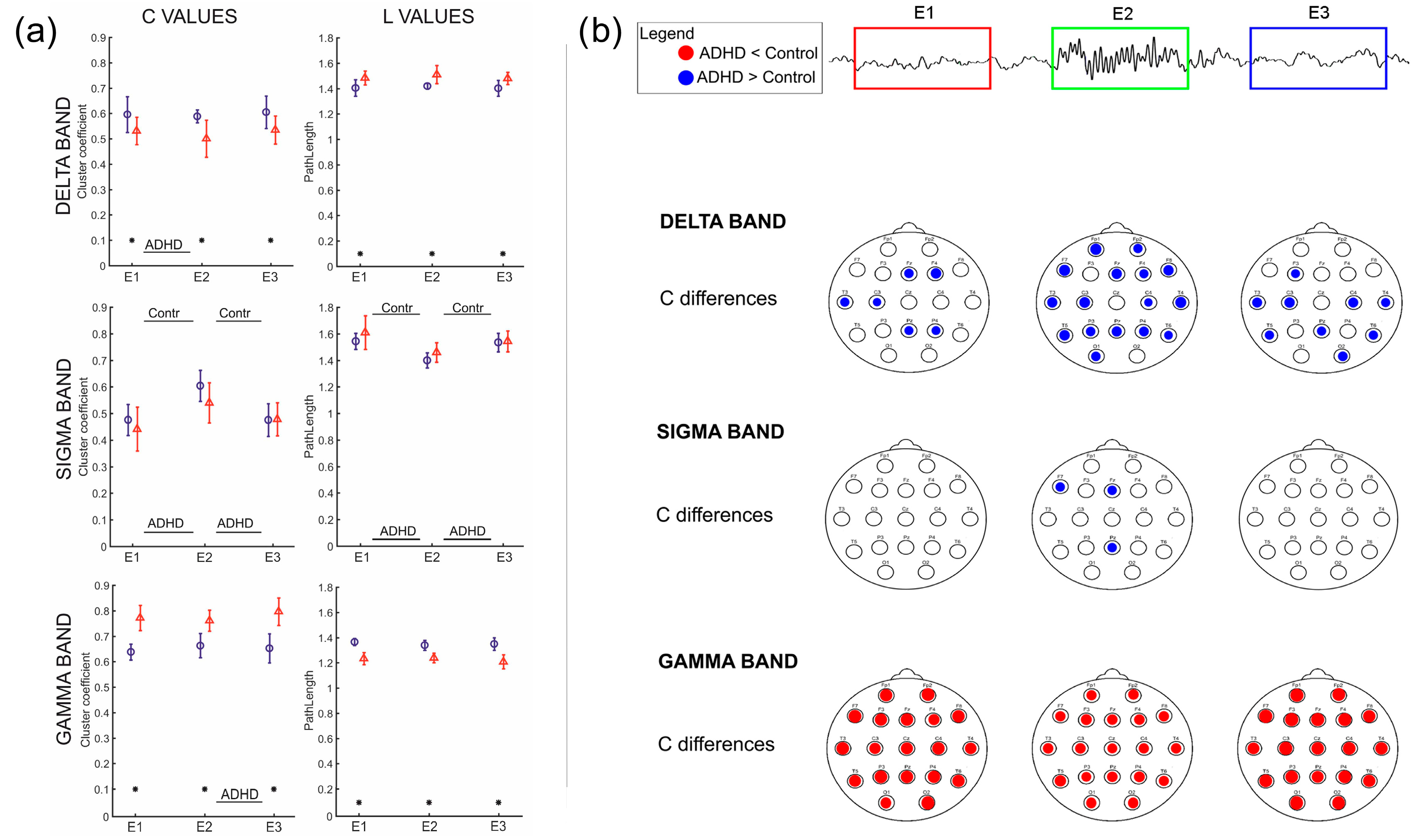
3.3. Clustering Coefficient and Path Length
4. Discussion
4.1. Neurophysiological Characteristics and Functional Connectivity—Sleep EEG in ADHD
4.2. Dynamic Modifications of the Neural Network in ADHD and Controls across Frequency Bands: Different Balances between Different Functions?
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 3rd ed.; DSM-III-R; American Psychiatric Association Publishing: Washington, DC, USA, 1987. [Google Scholar]
- Thome, J.; Ehlis, A.C.; Fallgatter, A.J.; Krauel, K.; Lange, K.W.; Riederer, P.; Romanos, M.; Taurines, R.; Tucha, O.; Uzbekov, M.; et al. Biomarkers for Attention-Deficit/Hyperactivity Disorder (ADHD). A Consensus Report of the WFSBP Task Force on Biological Markers and the World Federation of ADHD. World J. Biol. Psychiatry 2012, 13, 379–400. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.M.; Rugino, T.A.; Hornig, M.; Stein, M.A. Integration of an EEG Biomarker with a Clinician’s ADHD Evaluation. Brain Behav. 2015, 5, e00330. [Google Scholar] [CrossRef]
- Chang, M.-Y.; Ouyang, C.-S.; Chiang, C.-T.; Yang, R.-C.; Wu, R.-C.; Wu, H.-C.; Lin, L.-C. A New Method of Diagnosing Attention-Deficit Hyperactivity Disorder in Male Patients by Quantitative EEG Analysis. Clin. EEG Neurosci. 2019, 50, 339–347. [Google Scholar] [CrossRef]
- Chiarenza, G.A. Quantitative EEG in Childhood Attention Deficit Hyperactivity Disorder and Learning Disabilities. Clin. EEG Neurosci. 2021, 52, 144–155. [Google Scholar] [CrossRef]
- McVoy, M.; Lytle, S.; Fulchiero, E.; Aebi, M.E.; Adeleye, O.; Sajatovic, M. A Systematic Review of Quantitative EEG as a Possible Biomarker in Child Psychiatric Disorders. Psychiatry Res. 2019, 279, 331–344. [Google Scholar] [CrossRef]
- Du, Y.; Fu, Z.; Calhoun, V.D. Classification and Prediction of Brain Disorders Using Functional Connectivity: Promising but Challenging. Front. Neurosci. 2018, 12, 525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cheng, W.; Liu, Z.; Zhang, K.; Lei, X.; Yao, Y.; Becker, B.; Liu, Y.; Kendrick, K.M.; Lu, G.; et al. Neural, Electrophysiological and Anatomical Basis of Brain-Network Variability and Its Characteristic Changes in Mental Disorders. Brain 2016, 139, 2307–2321. [Google Scholar] [CrossRef]
- Lenartowicz, A.; Loo, S.K. Use of EEG to Diagnose ADHD. Curr. Psychiatry Rep. 2014, 16, 498. [Google Scholar] [CrossRef]
- Loo, S.K.; Makeig, S. Clinical Utility of EEG in Attention-Deficit/Hyperactivity Disorder: A Research Update. Neurotherapeutics 2012, 9, 569–587. [Google Scholar] [CrossRef] [PubMed]
- Alba, G.; Pereda, E.; Mañas, S.; Méndez, L.D.; González, A.; González, J.J. Electroencephalography Signatures of Attention-Deficit/Hyperactivity Disorder: Clinical Utility. Neuropsychiatr. Dis. Treat. 2015, 11, 2755–2769. [Google Scholar] [CrossRef]
- Kirkland, A.E.; Holton, K.F. Measuring Treatment Response in Pharmacological and Lifestyle Interventions Using Electroencephalography in ADHD: A Review. Clin. EEG Neurosci. 2019, 50, 256–266. [Google Scholar] [CrossRef]
- Michelini, G.; Jurgiel, J.; Bakolis, I.; Cheung, C.H.M.; Asherson, P.; Loo, S.K.; Kuntsi, J.; Mohammad-Rezazadeh, I. Atypical Functional Connectivity in Adolescents and Adults with Persistent and Remitted ADHD during a Cognitive Control Task. Transl. Psychiatry 2019, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Ghaderi, A.H.; Nazari, M.A.; Shahrokhi, H.; Darooneh, A.H. Functional Brain Connectivity Differences between Different ADHD Presentations: Impaired Functional Segregation in ADHD-Combined Presentation but Not in ADHD-Inattentive Presentation. Basic Clin. Neurosci. 2017, 8, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, P.; Covino, A.; Cristaldi, L.; Frosolone, M.; Gargiulo, L.; Mancino, F.; Mantile, F.; Moccaldi, N. A Systematic Review on Feature Extraction in Electroencephalography-Based Diagnostics and Therapy in Attention Deficit Hyperactivity Disorder. Sensors 2022, 22, 4934. [Google Scholar] [CrossRef]
- Ahmadi, M.; Kazemi, K.; Kuc, K.; Cybulska-Klosowicz, A.; Helfroush, M.S.; Aarabi, A. Disrupted Functional Rich-Club Organization of the Brain Networks in Children with Attention-Deficit/Hyperactivity Disorder, a Resting-State EEG Study. Brain Sci. 2021, 11, 938. [Google Scholar] [CrossRef]
- Alim, A.; Imtiaz, M.H. Automatic Identification of Children with ADHD from EEG Brain Waves. Signals 2023, 4, 193–205. [Google Scholar] [CrossRef]
- Shephard, E.; Tye, C.; Ashwood, K.L.; Azadi, B.; Johnson, M.H.; Charman, T.; Asherson, P.; McLoughlin, G.; Bolton, P.F. Oscillatory Neural Networks Underlying Resting-State, Attentional Control and Social Cognition Task Conditions in Children with ASD, ADHD and ASD+ADHD. Cortex 2019, 117, 96–110. [Google Scholar] [CrossRef]
- Konofal, E.; Lecendreux, M.; Cortese, S. Sleep and ADHD. Sleep Med. 2010, 11, 652–658. [Google Scholar] [CrossRef]
- Berry, R.B.; Brooks, R.; Gamaldo, C.E.; Harding, S.M.; Marcus, C.L.; Vaughn, B.V. The AASM Manual for the Scoring of Sleep and Associated Events. Am. Acad. Sleep Med. 2013, 53, 1689–1699. [Google Scholar]
- Mikoteit, T.; Brand, S.; Perren, S.; von Wyl, A.; von Klitzing, K.; Kurath, J.; Holsboer-Trachsler, E.; Hatzinger, M. Visually Detected Non−rapid Eye Movement Stage 2 Sleep Spindle Density at Age Five Years Predicted Prosocial Behavior Positively and Hyperactivity Scores Negatively at Age Nine Years. Sleep Med. 2018, 48, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Gruber, R.; Cassoff, J. The Interplay Between Sleep and Emotion Regulation: Conceptual Framework Empirical Evidence and Future Directions. Curr. Psychiatry Rep. 2014, 16, 500. [Google Scholar] [CrossRef] [PubMed]
- Koo, P.C.; Marshall, L. Neuroscience: A Sleep Rhythm with Multiple Facets. Curr. Biol. 2016, 26, R813–R815. [Google Scholar] [CrossRef] [PubMed]
- Astori, S.; Wimmer, R.D.; Lüthi, A. Manipulating Sleep Spindles—Expanding Views on Sleep, Memory, and Disease. Trends Neurosci. 2013, 36, 738–748. [Google Scholar] [CrossRef]
- Timofeev, I. Neuronal Plasticity and Thalamocortical Sleep and Waking Oscillations. Prog. Brain Res. 2011, 193, 121–144. [Google Scholar] [CrossRef]
- Fogel, S.M.; Smith, C.T. The Function of the Sleep Spindle: A Physiological Index of Intelligence and a Mechanism for Sleep-Dependent Memory Consolidation. Neurosci. Biobehav. Rev. 2011, 35, 1154–1165. [Google Scholar] [CrossRef]
- Fogerson, P.M.; Huguenard, J.R. Tapping the Brakes: Cellular and Synaptic Mechanisms That Regulate Thalamic Oscillations. Neuron 2016, 92, 687–704. [Google Scholar] [CrossRef]
- Krishnan, G.P.; Rosen, B.Q.; Chen, J.Y.; Muller, L.; Sejnowski, T.J.; Cash, S.S.; Halgren, E.; Bazhenov, M. Thalamocortical and Intracortical Laminar Connectivity Determines Sleep Spindle Properties. PLoS Comput. Biol. 2018, 14, 1006171. [Google Scholar] [CrossRef] [PubMed]
- Hobson, J.A.; Pace-Schott, E.F. The Cognitive Neuroscience of Sleep: Neuronal Systems, Consciousness and Learning. Nat. Rev. Neurosci. 2002, 3, 679–693. [Google Scholar] [CrossRef]
- Cirelli, C.; Tononi, G. Cortical Development, Electroencephalogram Rhythms, and the Sleep/Wake Cycle. Biol. Psychiatry 2015, 77, 1071–1078. [Google Scholar] [CrossRef]
- Astolfi, L.; Cincotti, F.; Mattia, D.; Marciani, M.G.; Baccalà, L.A.; De Vico Fallani, F.; Salinari, S.; Ursino, M.; Zavaglia, M.; Babiloni, F. Assessing Cortical Functional Connectivity by Partial Directed Coherence: Simulations and Application to Real Data. IEEE Trans. Biomed. Eng. 2006, 53, 1802–1812. [Google Scholar] [CrossRef]
- Astolfi, L.; Cincotti, F.; Mattia, D.; Marciani, M.G.; Baccala, L.A.; Fallani, F.D.V.; Salinari, S.; Ursino, M.; Zavaglia, M.; Ding, L.; et al. Comparison of Different Cortical Connectivity Estimators for High-Resolution EEG Recordings. Hum. Brain Mapp. 2007, 28, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Faes, L.; Erla, S.; Nollo, G. Measuring Connectivity in Linear Multivariate Processes: Definitions, Interpretation, and Practical Analysis. Comput. Math. Methods Med. 2012, 2012, 140513. [Google Scholar] [CrossRef]
- Von Bünau, P.; Meinecke, F.C.; Scholler, S.; Müller, K.R. Finding Stationary Brain Sources in EEG Data. In Proceedings of the 2010 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, EMBC’10, Buenos Aires, Argentina, 31 August–4 September 2010; pp. 2810–2813. [Google Scholar]
- Miladinović, A.; Ajčević, M.; Jarmolowska, J.; Marusic, U.; Colussi, M.; Silveri, G.; Battaglini, P.P.; Accardo, A. Effect of Power Feature Covariance Shift on BCI Spatial-Filtering Techniques: A Comparative Study. Comput. Methods Programs Biomed. 2021, 198, 105808. [Google Scholar] [CrossRef]
- Delorme, A.; Mullen, T.; Kothe, C.; Akalin Acar, Z.; Bigdely-Shamlo, N.; Vankov, A.; Makeig, S. EEGLAB, SIFT, NFT, BCILAB, and ERICA: New Tools for Advanced EEG Processing. Comput. Intell. Neurosci. 2011, 2011, 130714. [Google Scholar] [CrossRef]
- Mullen, T.R. The Dynamic Brain: Modeling Neural Dynamics and Interactions from Human Electrophysiological Recordings; University of California: San Diego, CA, USA, 2014; ISBN 9781321234619. [Google Scholar]
- Baccalá, L.A.; Sameshima, K. Partial Directed Coherence: A New Concept in Neural Structure Determination. Biol. Cybern. 2001, 84, 463–474. [Google Scholar] [CrossRef]
- Silfverhuth, M.J.; Hintsala, H.; Kortelainen, J.; Seppänen, T. Experimental Comparison of Connectivity Measures with Simulated EEG Signals. Med. Biol. Eng. Comput. 2012, 50, 683–688. [Google Scholar] [CrossRef]
- Baccala, L.A.; Sameshima, K.; Takahashi, D.Y.Y. Generalized Partial Directed Coherence. In Proceedings of the 2007 15th International Conference on Digital Signal Processing, Cardiff, UK, 1–4 July 2007; pp. 163–166. [Google Scholar]
- Rubinov, M.; Sporns, O. Complex Network Measures of Brain Connectivity: Uses and Interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chen, Y.; Lin, P.; Wang, J. Small-World Brain Functional Networks in Children with Attention-Deficit/Hyperactivity Disorder Revealed by EEG Synchrony. Clin. EEG Neurosci. 2015, 46, 183–191. [Google Scholar] [CrossRef]
- Ahmadlou, M.; Adeli, H.; Adeli, A. Graph Theoretical Analysis of Organization of Functional Brain Networks in ADHD. Clin. EEG Neurosci. 2012, 43, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Furlong, S.; Cohen, J.R.; Hopfinger, J.; Snyder, J.; Robertson, M.M.; Sheridan, M.A. Resting-State EEG Connectivity in Young Children with ADHD. J. Clin. Child Adolesc. Psychol. 2021, 50, 746–762. [Google Scholar] [CrossRef]
- Liu, X.; Sun, L.; Zhang, D.; Wang, S.; Hu, S.; Fang, B.; Yan, G.; Sui, G.; Huang, Q.; Wang, S. Phase-Amplitude Coupling Brain Networks in Children with Attention-Deficit/Hyperactivity Disorder. Clin. EEG Neurosci. 2022, 53, 399–405. [Google Scholar] [CrossRef]
- Buzsáki, G.; Wang, X.-J. Mechanisms of Gamma Oscillations. Annu. Rev. Neurosci. 2012, 35, 203–225. [Google Scholar] [CrossRef]
- Hashemi, N.S.; Dehnavi, F.; Moghimi, S.; Ghorbani, M. Slow Spindles Are Associated with Cortical High Frequency Activity. Neuroimage 2019, 189, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Ueda, R.; Takeichi, H.; Kaga, Y.; Oguri, M.; Saito, Y.; Nakagawa, E.; Maegaki, Y.; Inagaki, M. Atypical Gamma Functional Connectivity Pattern during Light Sleep in Children with Attention Deficit Hyperactivity Disorder. Brain Dev. 2020, 42, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Stam, C.J. Characterization of Anatomical and Functional Connectivity in the Brain: A Complex Networks Perspective. Int. J. Psychophysiol. 2010, 77, 186–194. [Google Scholar] [CrossRef]
- Liao, X.; Vasilakos, A.V.; He, Y. Small-World Human Brain Networks: Perspectives and Challenges. Neurosci. Biobehav. Rev. 2017, 77, 286–300. [Google Scholar] [CrossRef]
- Vecchio, F.; Miraglia, F.; Gorgoni, M.; Ferrara, M.; Iberite, F.; Bramanti, P.; De Gennaro, L.; Rossini, P.M. Cortical Connectivity Modulation during Sleep Onset: A Study via Graph Theory on EEG Data. Hum. Brain Mapp. 2017, 38, 5456–5464. [Google Scholar] [CrossRef]
- Berkman, J.M. Sleep in Children with Developmental Disabilities. Med. Health. R. I. 2006, 89, 94–96. [Google Scholar] [PubMed]
- Doran, S.M.; Harvey, M.T.; Horner, R.H. Sleep and Developmental Disabilities: Assessment, Treatment, and Outcome Measures. Ment. Retard. 2006, 44, 13–27. [Google Scholar] [CrossRef]
- Darchia, N.; Campbell, I.G.; Basishvili, T.; Eliozishvili, M.; Tchintcharauli, T.; Oniani, N.; Sakhelashvili, I.; Shetekauri, T.; Oniani, T.; Feinberg, I. Longitudinal Assessment of NREM Sleep EEG in Typically Developing and Medication-Free ADHD Adolescents: First Year Results. Sleep Med. 2021, 80, 171–175. [Google Scholar] [CrossRef]
- Biancardi, C.; Sesso, G.; Masi, G.; Faraguna, U.; Sicca, F. Sleep EEG Microstructure in Children and Adolescents with Attention Deficit Hyperactivity Disorder: A Systematic Review and Meta-Analysis. Sleep 2021, 44, zsab006. [Google Scholar] [CrossRef]
- Castelnovo, A.; Lividini, A.; Bernardi, G.; Pezzoli, V.; Foderaro, G.; Ramelli, G.P.; Manconi, M.; Miano, S. Sleep Power Topography in Children with Attention Deficit Hyperactivity Disorder (ADHD). Children 2022, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Cortese, S.; Faraone, S.V.; Konofal, E.; Lecendreux, M. Sleep in Children With Attention-Deficit/Hyperactivity Disorder: Meta-Analysis of Subjective and Objective Studies. J. Am. Acad. Child Adolesc. Psychiatry 2009, 48, 894–908. [Google Scholar] [CrossRef]
- Miano, S.; Parisi, P.; Villa, M.P. The Sleep Phenotypes of Attention Deficit Hyperactivity Disorder: The Role of Arousal during Sleep and Implications for Treatment. Med. Hypotheses 2012, 79, 147–153. [Google Scholar] [CrossRef]
- Achard, S.; Salvador, R.; Whitcher, B.; Suckling, J.; Bullmore, E. A Resilient, Low-Frequency, Small-World Human Brain Functional Network with Highly Connected Association Cortical Hubs. J. Neurosci. 2006, 26, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Supekar, K.; Musen, M.; Menon, V. Development of Large-Scale Functional Brain Networks in Children. PLoS Biol. 2009, 7, e1000157. [Google Scholar] [CrossRef]
- Salvador, R.; Suckling, J.; Schwarzbauer, C.; Bullmore, E. Undirected Graphs of Frequency-Dependent Functional Connectivity in Whole Brain Networks. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Titone, S.; Samogin, J.; Peigneux, P.; Swinnen, S.P.; Mantini, D.; Albouy, G. Frequency-dependent Connectivity in Large-scale Resting-state Brain Networks during Sleep. Eur. J. Neurosci. 2023. [Google Scholar] [CrossRef]
- Steriade, M.; Contreras, D.; Amzica, F. Synchronized Sleep Oscillations and Their Paroxysmal Developments. Trends Neurosci. 1994, 17, 201–207. [Google Scholar] [CrossRef]
- Valderrama, M.; Crépon, B.; Botella-Soler, V.; Martinerie, J.; Hasboun, D.; Alvarado-Rojas, C.; Baulac, M.; Adam, C.; Navarro, V.; Le Van Quyen, M. Human Gamma Oscillations during Slow Wave Sleep. PLoS ONE 2012, 7, 33477. [Google Scholar] [CrossRef]
- Cox, R.; Van Driel, J.; De Boer, M.; Talamini, L.M. Slow Oscillations during Sleep Coordinate Interregional Communication in Cortical Networks. J. Neurosci. 2014, 34, 16890–16901. [Google Scholar] [CrossRef] [PubMed]
- Piantoni, G.; Halgren, E.; Cash, S.S. Spatiotemporal Characteristics of Sleep Spindles Depend on Cortical Location. Neuroimage 2017, 146, 236–245. [Google Scholar] [CrossRef]
- Mills, K.L.; Bathula, D.; Dias, T.G.C.; Iyer, S.P.; Fenesy, M.C.; Musser, E.D.; Stevens, C.A.; Thurlow, B.L.; Carpenter, S.D.; Nagel, B.J.; et al. Altered Cortico-Striatal-Thalamic Connectivity in Relation to Spatial Working Memory Capacity in Children with ADHD. Front. Psychiatry 2012, 3, 2. [Google Scholar] [CrossRef]
- Welton, T.; Kent, D.A.; Auer, D.P.; Dineen, R.A. Reproducibility of Graph-Theoretic Brain Network Metrics: A Systematic Review. Brain Connect. 2015, 5, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Telesford, Q.K.; Simpson, S.L.; Burdette, J.H.; Hayasaka, S.; Laurienti, P.J. The Brain as a Complex System: Using Network Science as a Tool for Understanding the Brain. Brain Connect. 2011, 1, 295–308. [Google Scholar] [CrossRef]
- Liu, S.; Guo, J.; Meng, J.; Wang, Z.; Yao, Y.; Yang, J.; Qi, H.; Ming, D. Abnormal EEG Complexity and Functional Connectivity of Brain in Patients with Acute Thalamic Ischemic Stroke. Comput. Math. Methods Med. 2016, 2016, 2582478. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; Kazemi, K.; Kuc, K.; Cybulska-Klosowicz, A.; Zakrzewska, M.; Racicka-Pawlukiewicz, E.; Helfroush, M.S.; Aarabi, A. Cortical Source Analysis of Resting State EEG Data in Children with Attention Deficit Hyperactivity Disorder. Clin. Neurophysiol. 2020, 131, 2115–2130. [Google Scholar] [CrossRef]
- Florian, G.; Pfurtscheller, G. Dynamic spectral analysis of event-related EEG data. Electroencephalogr. Clin. Neurophysiol. 1995, 95, 393–396. [Google Scholar] [CrossRef]
- Jansen, B.H.; Bourne, J.R.; Ward, J.W. Autoregressive estimation of short segment spectra for computerized EEG analysis. IEEE Trans. Biomed. Eng. 1981, 28, 630–638. [Google Scholar] [CrossRef]
- De Brito, C.S.; Baccalá, L.A.; Takahashi, D.Y.; Sameshima, K. Asymptotic behavior of generalized partial directed coherence. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2010, 2010, 1718–1721. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanus, C.; Miladinović, A.; De Dea, F.; Skabar, A.; Stecca, M.; Ajčević, M.; Accardo, A.; Carrozzi, M. Sleep Spindle-Related EEG Connectivity in Children with Attention-Deficit/Hyperactivity Disorder: An Exploratory Study. Entropy 2023, 25, 1244. https://doi.org/10.3390/e25091244
Zanus C, Miladinović A, De Dea F, Skabar A, Stecca M, Ajčević M, Accardo A, Carrozzi M. Sleep Spindle-Related EEG Connectivity in Children with Attention-Deficit/Hyperactivity Disorder: An Exploratory Study. Entropy. 2023; 25(9):1244. https://doi.org/10.3390/e25091244
Chicago/Turabian StyleZanus, Caterina, Aleksandar Miladinović, Federica De Dea, Aldo Skabar, Matteo Stecca, Miloš Ajčević, Agostino Accardo, and Marco Carrozzi. 2023. "Sleep Spindle-Related EEG Connectivity in Children with Attention-Deficit/Hyperactivity Disorder: An Exploratory Study" Entropy 25, no. 9: 1244. https://doi.org/10.3390/e25091244
APA StyleZanus, C., Miladinović, A., De Dea, F., Skabar, A., Stecca, M., Ajčević, M., Accardo, A., & Carrozzi, M. (2023). Sleep Spindle-Related EEG Connectivity in Children with Attention-Deficit/Hyperactivity Disorder: An Exploratory Study. Entropy, 25(9), 1244. https://doi.org/10.3390/e25091244