
Detection of Anticipatory Dynamics between a Pair of Zebrafish

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiments of Fish Pairs
2.2. Time Lag Mutual Information (TLMI)
2.3. Detection of Direction of Information Flow (DIF)
2.3.1. Granger Causality (GC)
2.3.2. Liang’s T Method
2.3.3. Transfer Entropy (TE)
2.4. Simulation of Anticipatory Data
3. Results
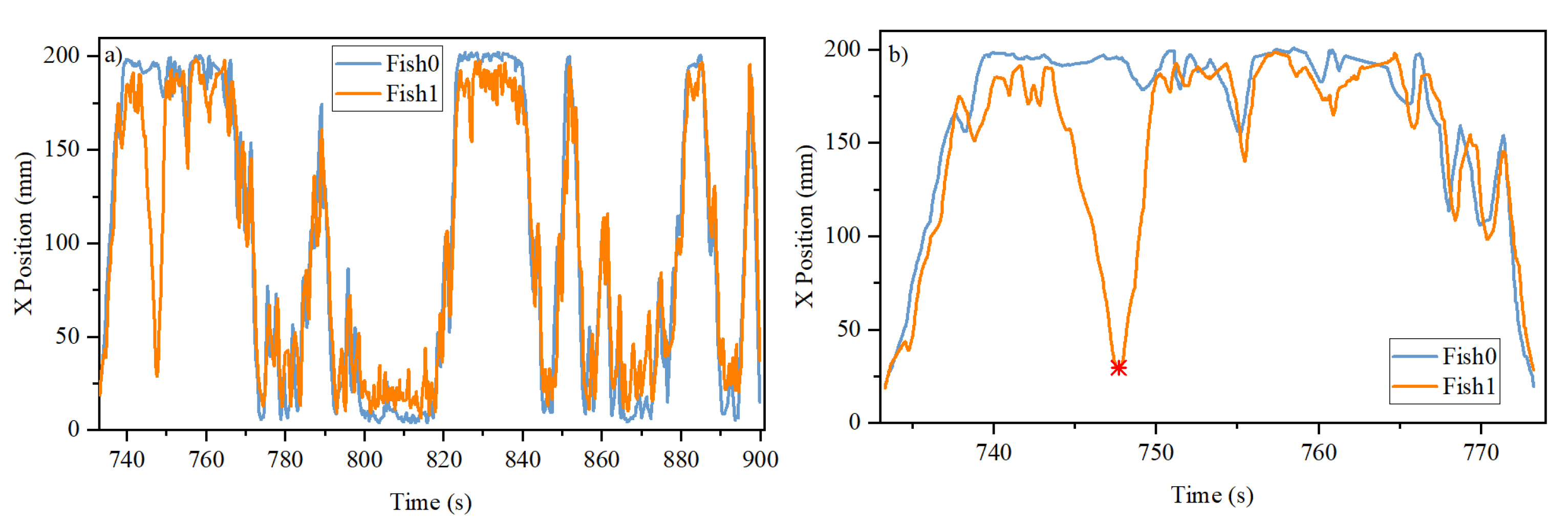
3.1. Trajectories in One- and Three-Door Tanks
3.2. Time Lag Mutual Information from the X Component (xTLMI)
3.3. yTLMI and DIF Detection
3.4. Summary of Experimental Results
3.5. DIF Detection by Statistical Methods
3.6. Testing of DIF Detection with Simulated Anticipatory Data
4. Limitations
4.1. Predictive and True Causality
4.2. Detection of Anticipatory Dynamics
4.3. Issues of Entropy Based Causality
4.4. Correlation and Causation
4.5. Problem of Bi-Directional Coupling
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations and Symbols
| AD | Anticipatory dynamics |
| DIF | Direction of information flow |
| Expt | Experiment |
| GC | Granger causality |
| NGD | Negative group delay |
| OU | Ornstein–Uhlenbeck |
| TE | Transfer Entropy |
| xTLMI | Time lag mutual information from the x component |
| Time series from the trajectories | |
| Source and target time series in the NGD model | |
| W | Ornstein–Uhlenbeck noise |
| P | Probability distribution |
| Error (residuals) | |
| F-test | |
| p | p-value |
| T | Value of Liang’s T mnethod |
| C | Covariance |
| Value of transfer entropy | |
| H | Entropy |
References
- Fodor, É.; Marchetti, M.C. The statistical physics of active matter: From self-catalytic colloids to living cells. Phys. A Stat. Mech. Its Appl. 2018, 504, 106–120. [Google Scholar] [CrossRef]
- Voss, H.U. Signal prediction by anticipatory relaxation dynamics. Phys. Rev. E 2016, 93, 030201. [Google Scholar] [CrossRef] [PubMed]
- Voss, H.U. Anticipating chaotic synchronization. Phys. Rev. E 2000, 61, 5115. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.J., II; Brivanlou, I.H.; Jordan, T.A.; Meister, M. Anticipation of moving stimuli by the retina. Nature 1999, 398, 334. [Google Scholar] [CrossRef] [PubMed]
- Chou, P.Y.; Chien, J.F.; Chen, K.S.; Huang, Y.T.; Chen, C.C.; Chan, C. Anticipation and negative group delay in a retina. Phys. Rev. E 2021, 103, L020401. [Google Scholar] [CrossRef] [PubMed]
- Vicsek, T.; Czirók, A.; Ben-Jacob, E.; Cohen, I.; Shochet, O. Novel type of phase transition in a system of self-driven particles. Phys. Rev. Lett. 1995, 75, 1226. [Google Scholar] [CrossRef] [PubMed]
- Katz, Y.; Tunstrøm, K.; Ioannou, C.C.; Huepe, C.; Couzin, I.D. Inferring the structure and dynamics of interactions in schooling fish. Proc. Natl. Acad. Sci. USA 2011, 108, 18720–18725. [Google Scholar] [CrossRef]
- Hlaváčková-Schindler, K.; Paluš, M.; Vejmelka, M.; Bhattacharya, J. Causality detection based on information—Theoretic approaches in time series analysis. Phys. Rep. 2007, 441, 1–46. [Google Scholar] [CrossRef]
- Vastano, J.A.; Swinney, H.L. Information transport in spatiotemporal systems. Phys. Rev. Lett. 1988, 60, 1773. [Google Scholar] [CrossRef]
- Van der Walt, S.; Schönberger, J.L.; Nunez-Iglesias, J.; Boulogne, F.; Warner, J.D.; Yager, N.; Gouillart, E.; Yu, T. scikit-image: Image processing in Python. PeerJ 2014, 2, e453. [Google Scholar] [CrossRef]
- Wiener, N. Cybernetics or Control and Communication in the Animal and the Machine; MIT Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Barnett, L.; Seth, A.K. The MVGC multivariate Granger causality toolbox: A new approach to Granger-causal inference. J. Neurosci. Methods 2014, 223, 50–68. [Google Scholar] [CrossRef] [PubMed]
- San Liang, X. Unraveling the cause-effect relation between time series. Phys. Rev. E 2014, 90, 052150. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, T. Measuring information transfer. Phys. Rev. Lett. 2000, 85, 461. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.G.; Valentini, G.; Walker, S.I.; Levin, M. Inform: Efficient information-theoretic analysis of collective behaviors. Front. Robot. AI 2018, 5, 60. [Google Scholar] [CrossRef] [PubMed]
- Wibral, M.; Vicente, R.; Lindner, M. Transfer entropy in neuroscience. In Directed Information Measures in Neuroscience; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3–36. [Google Scholar]
- Li, J.; Convertino, M. Inferring ecosystem networks as information flows. Sci. Rep. 2021, 11, 7094. [Google Scholar] [CrossRef] [PubMed]
- Pearl, J. Models, Reasoning and Inference; Cambridge University Press: Cambridge, UK, 2000; Volume 19, p. 3. [Google Scholar]
- Hyttinen, A.; Eberhardt, F.; Hoyer, P.O. Experiment selection for causal discovery. J. Mach. Learn. Res. 2013, 14, 3041–3071. [Google Scholar]
- Zenil, H.; Kiani, N.A.; Tegnér, J. Low-algorithmic-complexity entropy-deceiving graphs. Phys. Rev. E 2017, 96, 012308. [Google Scholar] [CrossRef]
- Hahs, D.W.; Pethel, S.D. Distinguishing anticipation from causality: Anticipatory bias in the estimation of information flow. Phys. Rev. Lett. 2011, 107, 128701. [Google Scholar] [CrossRef]
- Porta, A.; Castiglioni, P.; Di Rienzo, M.; Bassani, T.; Bari, V.; Faes, L.; Nollo, G.; Cividjan, A.; Quintin, L. Cardiovascular control and time domain Granger causality: Insights from selective autonomic blockade. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2013, 371, 20120161. [Google Scholar] [CrossRef]
- Gelpi, F.; Bari, V.; Cairo, B.; De Maria, B.; Tonon, D.; Rossato, G.; Faes, L.; Porta, A. Dynamic cerebrovascular autoregulation in patients prone to postural syncope: Comparison of techniques assessing the autoregulation index from spontaneous variability series. Auton. Neurosci. 2022, 237, 102920. [Google Scholar] [CrossRef]
- Porta, A.; Catai, A.M.; Takahashi, A.C.; Magagnin, V.; Bassani, T.; Tobaldini, E.; Van De Borne, P.; Montano, N. Causal relationships between heart period and systolic arterial pressure during graded head-up tilt. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 300, R378–R386. [Google Scholar] [CrossRef] [PubMed]
- Nagy, M.; Akos, Z.; Biro, D.; Vicsek, T. Hierarchical group dynamics in pigeon flocks. Nature 2010, 464, 890. [Google Scholar] [CrossRef] [PubMed]
- Bashirzade, A.A.; Zabegalov, K.N.; Volgin, A.D.; Belova, A.S.; Demin, K.A.; de Abreu, M.S.; Babchenko, V.Y.; Bashirzade, K.A.; Yenkoyan, K.B.; Tikhonova, M.A.; et al. Modeling neurodegenerative disorders in zebrafish. Neurosci. Biobehav. Rev. 2022, 138, 104679. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment Number | Number of Passages | ||
|---|---|---|---|
| Left | Middle | Right | |
| 1 | 28 | 19 | 33 |
| 2 | 28 | 20 | 20 |
| 3 | 51 | 16 | 31 |
| 4 | 20 | 6 | 18 |
| Probability | 43.8% | 21.0% | 35.2% |
| Properties of Cross xTLMI | Properties of Cross yTLMI | ||||||
|---|---|---|---|---|---|---|---|
| Expt Number | Fish Pair | Peak Position(s) | Peak Height | Decay Time(s) | Peak Position(s) | Peak Height | Decay Time(s) |
| 1 | A | −0.83 | 0.22 | 0.57 | −1.10 | 0.41 | 6.13 |
| 2 | B | 0.63 | 0.09 | 0.53 | −3.87 | 0.35 | 7.90 |
| 3 | B | −0.23 | 0.31 | 0.53 | −0.53 | 0.48 | 6.03 |
| 4 | C | −0.60 | 0.12 | 0.87 | −0.73 | 0.19 | 3.97 |
| 5 | D | 0.50 | 0.09 | 0.47 | −3.30 | 0.28 | 9.30 |
| 6 | D | −0.63 | 0.12 | 0.60 | −11.17 | 0.20 | 19.97 |
| 7 | E | −0.77 | 0.20 | 0.40 | −1.43 | 0.36 | 19.97 |
| 8 | F | 0.47 | 0.13 | 0.50 | −0.53 | 0.26 | 2.53 |
| 9 | F | −0.27 | 0.16 | 0.53 | −1.13 | 0.19 | 2.47 |
| 10 | G | −0.63 | 0.22 | 0.63 | −1.73 | 0.39 | 13.47 |
| 11 | H | −0.60 | 0.26 | 0.63 | −1.37 | 0.26 | 4.67 |
| 12 | I | −0.30 | 0.17 | 1.23 | −0.67 | 0.22 | 7.40 |
| 13 | J | −0.43 | 0.25 | 0.47 | −0.53 | 0.34 | 4.93 |
| 14 | L | 0.57 | 0.23 | 0.83 | −0.33 | 0.30 | 17.53 |
| 15 | L | −0.67 | 0.27 | 1.03 | −1.33 | 0.31 | 9.47 |
| 16 | M | −0.40 | 0.21 | 0.87 | −1.43 | 0.23 | 3.73 |
| Expt Number | Fish | Order | F (MVGC) | p-Value (MVGC) | Liang’s T | |||
|---|---|---|---|---|---|---|---|---|
| Pair | (BIC) | |||||||
| 1 | A | 20 | ||||||
| 2 | B | 20 | ||||||
| 3 | B | 20 | ||||||
| 4 | C | 15 | ||||||
| 5 | D | 11 | ||||||
| 6 | D | 18 | ||||||
| 7 | E | 20 | ||||||
| 8 | F | 14 | ||||||
| 9 | F | 20 | ||||||
| 10 | G | 20 | ||||||
| 11 | H | 20 | ||||||
| 12 | I | 10 | ||||||
| 13 | J | 20 | ||||||
| 14 | L | 10 | ||||||
| 15 | L | 10 | ||||||
| 16 | M | 20 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.-J.; Ko, I.-S.; Lin, C.-A.; Chen, C.-J.; Wu, J.-S.; Chan, C.K. Detection of Anticipatory Dynamics between a Pair of Zebrafish. Entropy 2024, 26, 13. https://doi.org/10.3390/e26010013
Chen W-J, Ko I-S, Lin C-A, Chen C-J, Wu J-S, Chan CK. Detection of Anticipatory Dynamics between a Pair of Zebrafish. Entropy. 2024; 26(1):13. https://doi.org/10.3390/e26010013
Chicago/Turabian StyleChen, Wei-Jie, I-Shih Ko, Chi-An Lin, Chun-Jen Chen, Jiun-Shian Wu, and C. K. Chan. 2024. "Detection of Anticipatory Dynamics between a Pair of Zebrafish" Entropy 26, no. 1: 13. https://doi.org/10.3390/e26010013