
Novel Facet of an Old Dietary Molecule? Direct Influence of Caffeine on Glucose and Biogenic Amine Handling by Human Adipocytes

 , , ,
, , ,  , , and
, , and 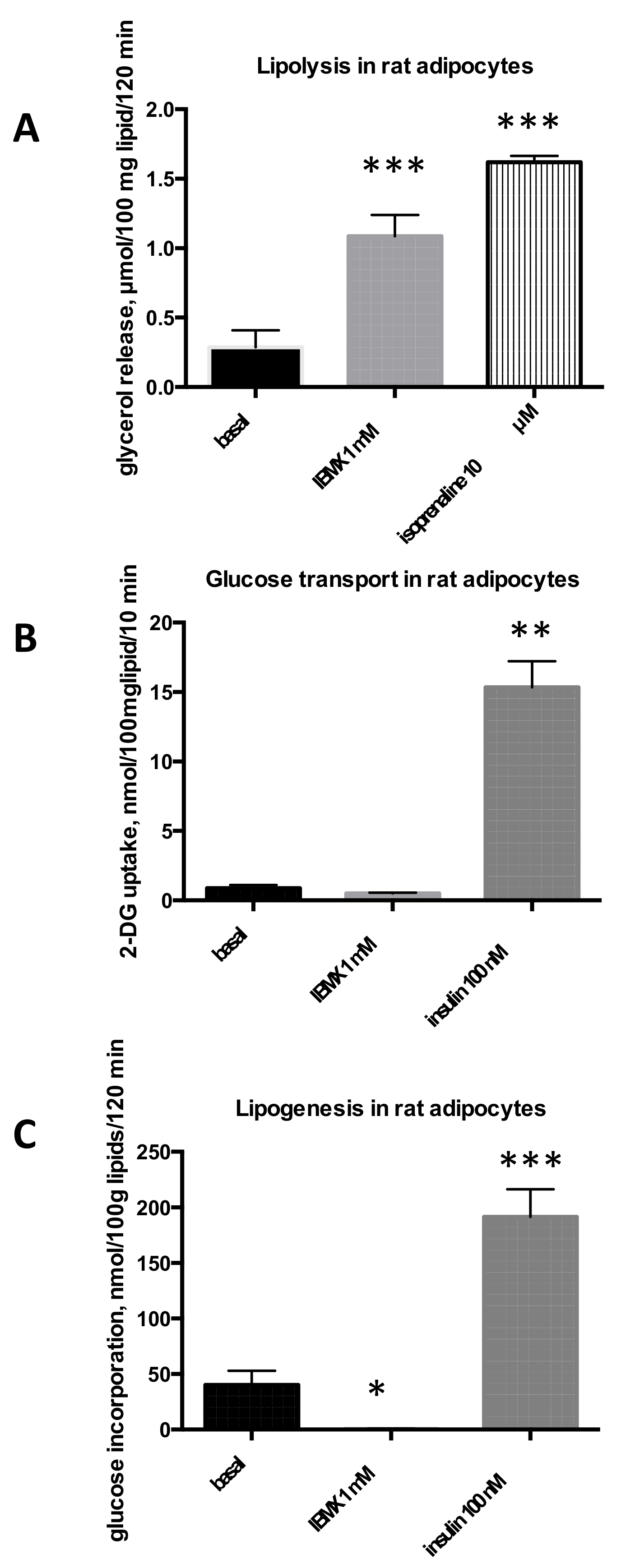{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
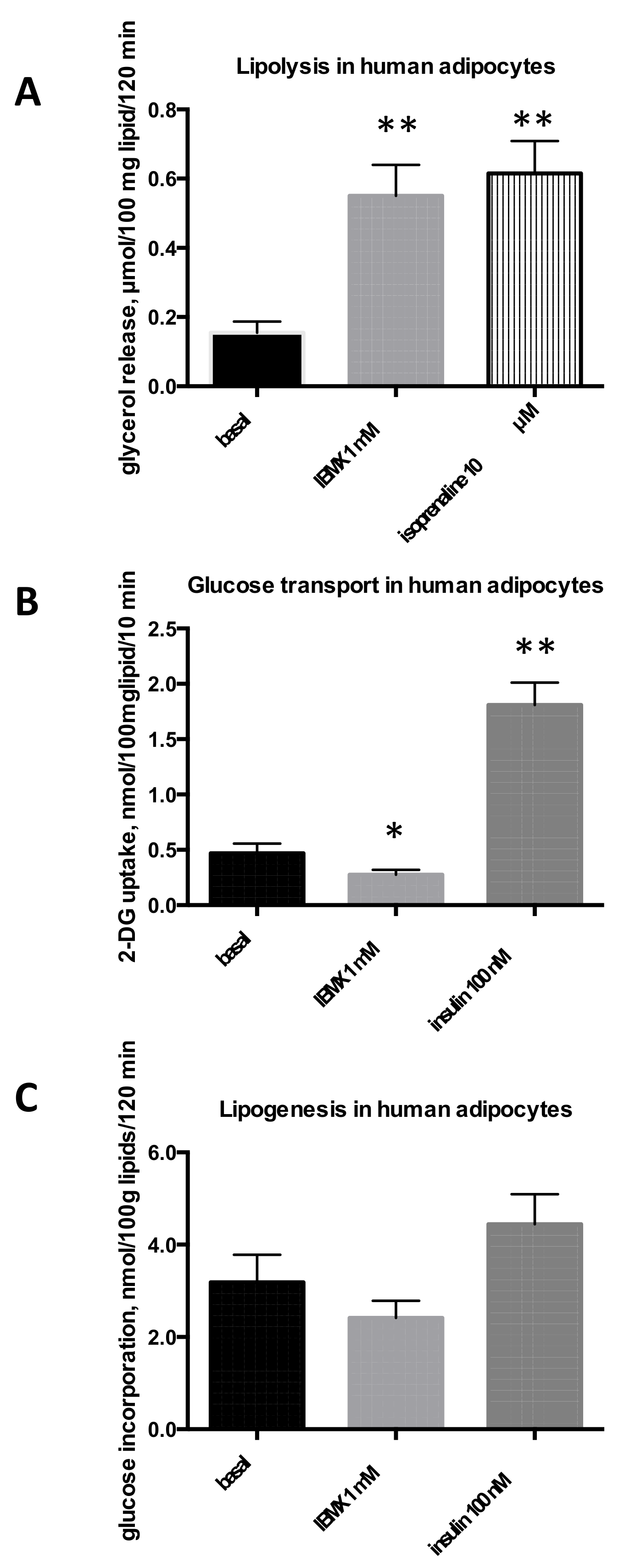
2.1. Influence of IBMX on Lipolysis, Glucose Uptake and Lipogenesis in Rat, Mouse, and Human Adipose Cells
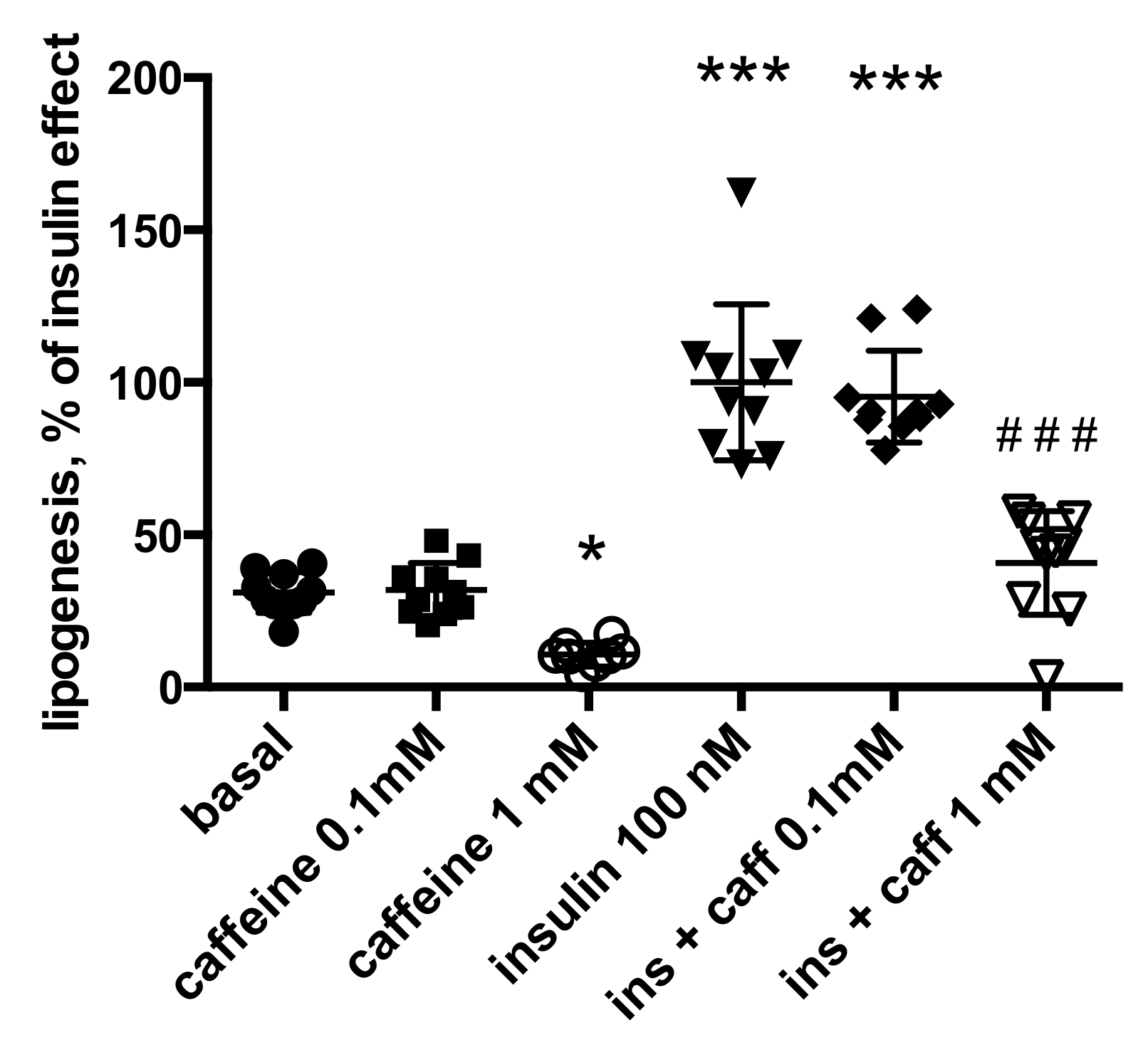
2.2. Effects of Caffeine on Glucose Uptake and Lipogenesis in Murine Adipose Cells
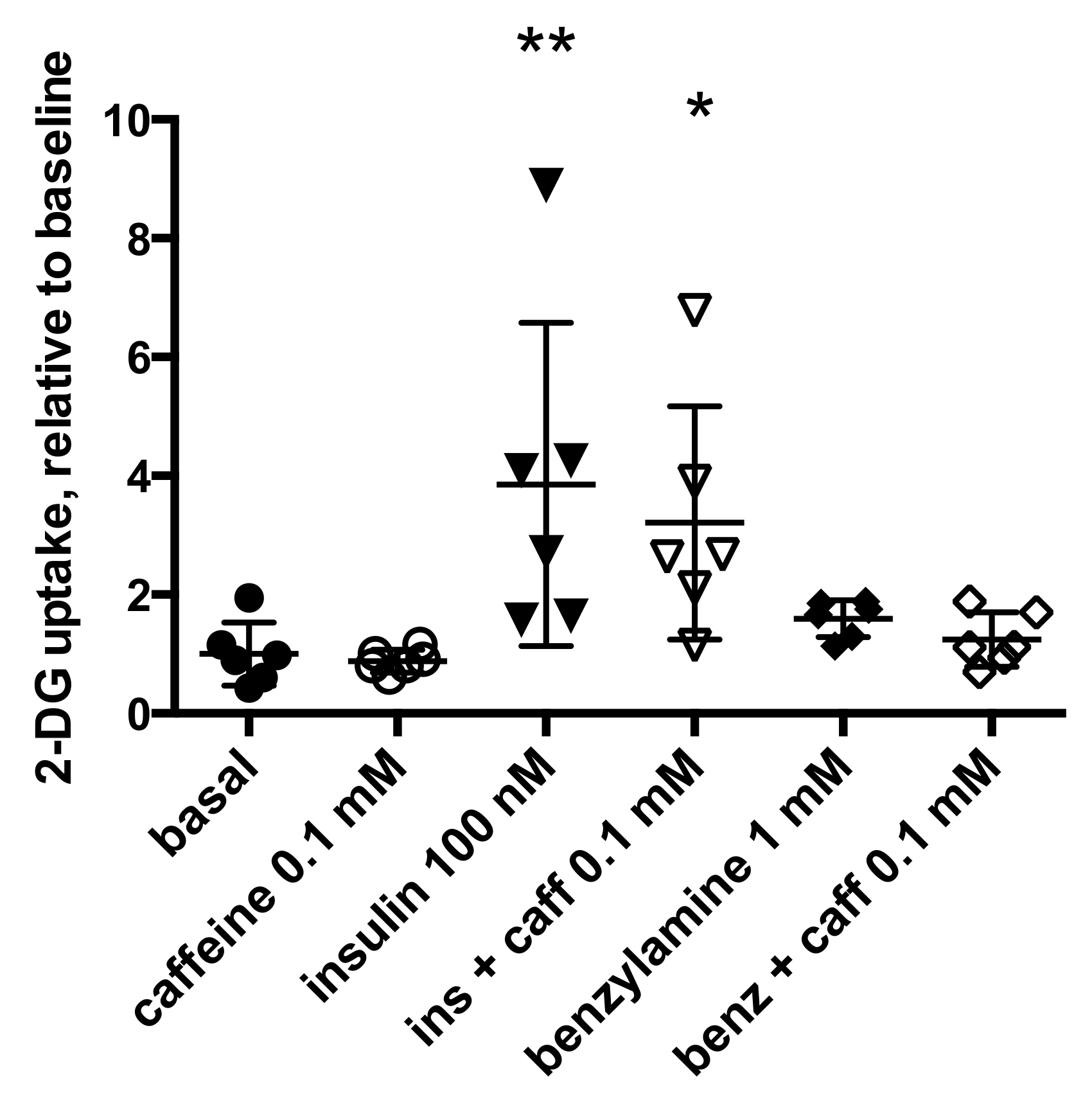
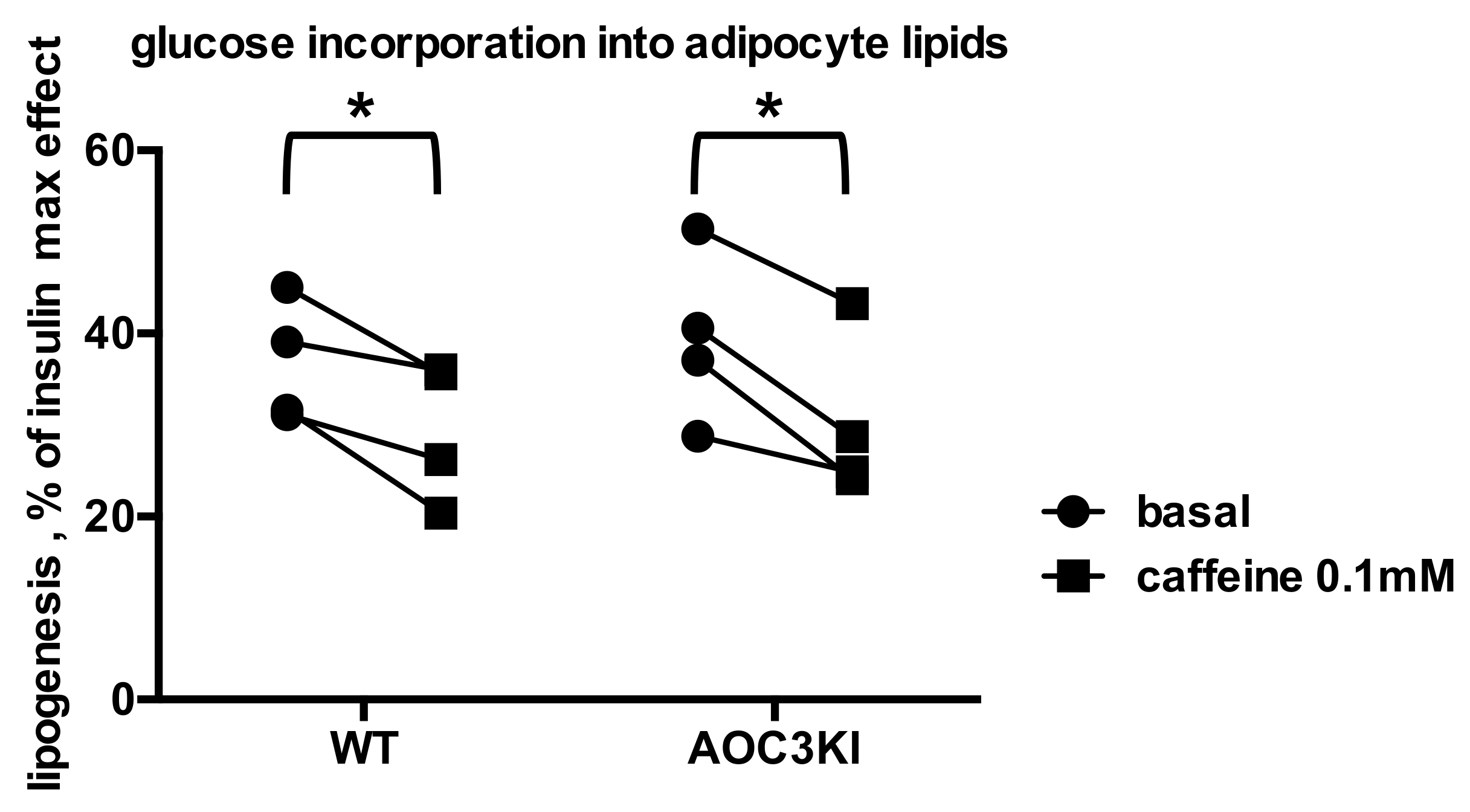
2.3. Is Caffeine Inhibiting Insulin Stimulation of Adipocyte Anabolism in Humans?
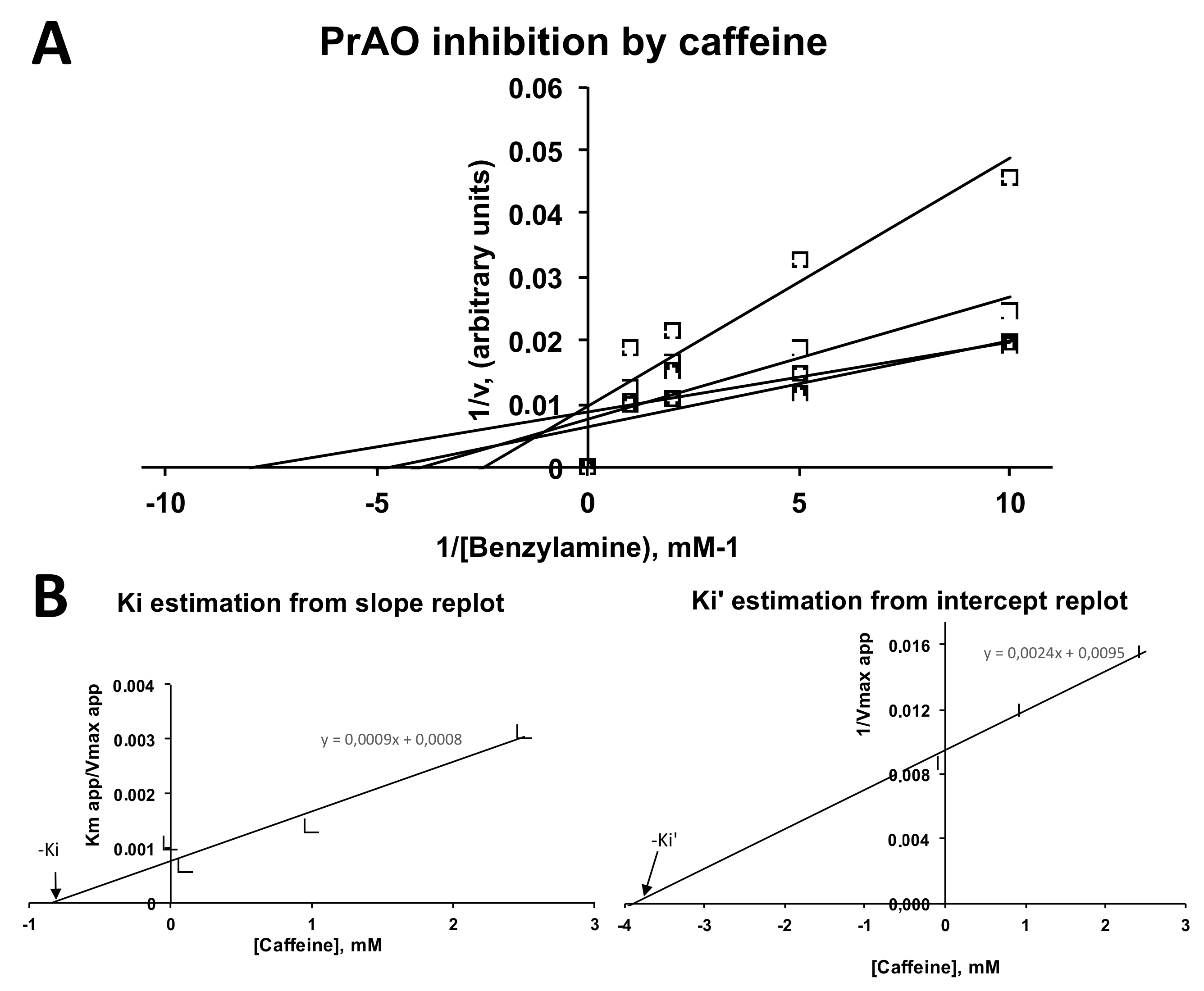
2.4. Inhibition by Caffeine of PrAO and MAO Activities in Human Adipose Tissue
2.5. Exploring the Mechanisms Involved in Caffeine Inhibition of Lipogenesis
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Animal Models
4.3. Patients and Adipose Tissue Biopsies
4.4. Preparations of Functional Adipocytes from Rodent and Human Adipose Depots
4.5. Lipolysis and Lipogenesis Assessment in Rodent and Human Adipose Cells
4.6. Glucose Transport Assay in Rodent and Human Adipose Cells
4.7. Amine Oxidase Activities in Human Adipose Tissue
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Barone, J.J.; Roberts, H.R. Caffeine consumption. Food Chem. Toxicol. 1996, 34, 119–129. [Google Scholar] [CrossRef]
- Mitchell, D.C.; Knight, C.A.; Hockenberry, J.; Teplansky, R.; Hartman, T.J. Beverage caffeine intakes in the U.S. Food Chem. Toxicol. 2014, 63, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Biesterbos, J.W.H.; Sijm, D.; van Dam, R.; Mol, H.G.J. A health risk for consumers: The presence of adulterated food supplements in the Netherlands. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2019, 36, 1273–1288. [Google Scholar] [CrossRef] [PubMed]
- Beyer, L.A.; Hixon, M.L. Review of animal studies on the cardiovascular effects of caffeine. Food Chem. Toxicol. 2018, 118, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Che, B.; Wang, L.; Zhang, Z.; Zhang, Y.; Deng, Y. Distribution and accumulation of caffeine in rat tissues and its inhibition on semicarbazide-sensitive amine oxidase. Neurotoxicology 2012, 33, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Dulloo, A.G.; Seydoux, J.; Girardier, L. Potentiation of the thermogenic antiobesity effects of ephedrine by dietary methylxanthines: Adenosine antagonism or phosphodiesterase inhibition? Metabolism 1992, 41, 1233–1241. [Google Scholar] [CrossRef]
- Shechter, Y. Differential effects of two phosphodiesterase inhibitors on fat cell metabolism. Endocrinology 1984, 115, 1787–1791. [Google Scholar] [CrossRef]
- Monteiro, J.P.; Alves, M.G.; Oliveira, P.F.; Silva, B.M. Structure-Bioactivity Relationships of Methylxanthines: Trying to Make Sense of All the Promises and the Drawbacks. Molecules 2016, 21, 974. [Google Scholar] [CrossRef] [Green Version]
- Velickovic, K.; Wayne, D.; Leija, H.A.L.; Bloor, I.; Morris, D.E.; Law, J.; Budge, H.; Sacks, H.; Symonds, M.E.; Sottile, V. Caffeine exposure induces browning features in adipose tissue in vitro and in vivo. Sci. Rep. 2019, 9, 9104. [Google Scholar] [CrossRef] [Green Version]
- Panchal, S.K.; Wong, W.Y.; Kauter, K.; Ward, L.C.; Brown, L. Caffeine attenuates metabolic syndrome in diet-induced obese rats. Nutrition 2012, 28, 1055–1062. [Google Scholar] [CrossRef]
- Steinfelder, H.J.; Petho-Schramm, S. Methylxanthines inhibit glucose transport in rat adipocytes by two independent mechanisms. Biochem. Pharmacol. 1990, 40, 1154–1157. [Google Scholar] [CrossRef]
- Akiba, T.; Yaguchi, K.; Tsutsumi, K.; Nishioka, T.; Koyama, I.; Nomura, M.; Yokogawa, K.; Moritani, S.; Miyamoto, K. Inhibitory mechanism of caffeine on insulin-stimulated glucose uptake in adipose cells. Biochem. Pharmacol. 2004, 68, 1929–1937. [Google Scholar] [CrossRef]
- Nakabayashi, H.; Hashimoto, T.; Ashida, H.; Nishiumi, S.; Kanazawa, K. Inhibitory effects of caffeine and its metabolites on intracellular lipid accumulation in murine 3T3-L1 adipocytes. BioFactors 2008, 34, 293–302. [Google Scholar] [CrossRef]
- Chernogubova, E.; Cannon, B.; Bengtsson, T. Norepinephrine increases glucose transport in brown adipocytes via beta3-adrenoceptors through a cAMP, PKA, and PI3-kinase-dependent pathway stimulating conventional and novel PKCs. Endocrinology 2004, 145, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Dallner, O.S.; Chernogubova, E.; Brolinson, K.A.; Bengtsson, T. Beta3-adrenergic receptors stimulate glucose uptake in brown adipocytes by two mechanisms independently of glucose transporter 4 translocation. Endocrinology 2006, 147, 5730–5739. [Google Scholar] [CrossRef] [Green Version]
- Mukaida, S.; Evans, B.A.; Bengtsson, T.; Hutchinson, D.S.; Sato, M. Adrenoceptors promote glucose uptake into adipocytes and muscle by an insulin-independent signaling pathway involving mechanistic target of rapamycin complex 2. Pharmacol. Res. 2017, 116, 87–92. [Google Scholar] [CrossRef]
- Bour, S.; Daviaud, D.; Grès, S.; Lefort, C.; Prévot, D.; Zorzano, A.; Wabitsch, M.; Saulnier-Blache, J.S.; Valet, P.; Carpéné, C. Adipogenesis-related increase of semicarbazide-sensitive amine oxidase and monoamine oxidase in human adipocytes. Biochimie 2007, 89, 916–925. [Google Scholar] [CrossRef]
- Olivieri, A.; Rico, D.; Khiari, Z.; Henehan, G.; O’Sullivan, J.; Tipton, K. From caffeine to fish waste: Amine compounds present in food and drugs and their interactions with primary amine oxidase. J. Neural Transm. 2011, 118, 1079–1089. [Google Scholar] [CrossRef]
- Salmi, M.; Jalkanen, S. Vascular Adhesion Protein-1: A Cell Surface Amine Oxidase in Translation. Antioxid. Redox Signal. 2019, 30, 314–332. [Google Scholar] [CrossRef]
- Vakal, S.; Jalkanen, S.; Dahlström, K.M.; Salminen, T.A. Human Copper-Containing Amine Oxidases in Drug Design and Development. Molecules 2020, 25, 1293. [Google Scholar] [CrossRef] [Green Version]
- Carpéné, C.; Grès, S.; Rascalou, S. The amine oxidase inhibitor phenelzine limits lipogenesis in adipocytes without inhibiting insulin action on glucose uptake. J. Neural Transm. 2013, 120, 997–1003. [Google Scholar] [CrossRef]
- Carpéné, C.; Mercader, J.; Le Gonidec, S.; Schaak, S.; Mialet-Perez, J.; Zakaroff-Girard, A.; Galitzky, J. Body fat reduction without cardiovascular changes in mice after oral treatment by the MAO inhibitor phenelzine. Br. J. Pharmacol. 2018, 175, 2428–2440. [Google Scholar] [CrossRef] [Green Version]
- Carpéné, C.; Boulet, N.; Chaplin, A.; Mercader, J. Past, Present and Future Anti-Obesity Effects of Flavin-Containing and/or Copper-Containing Amine Oxidase Inhibitors. Medicines 2019, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Petzer, A.; Pienaar, A.; Petzer, J.P. The interactions of caffeine with monoamine oxidase. Life Sci. 2013, 93, 283–287. [Google Scholar] [CrossRef]
- Olivieri, A.; Tipton, K. Inhibition of bovine plasma semicarbazide-sensitive amine oxidase by caffeine. J. Biochem. Mol. Toxicol. 2011, 25, 26–27. [Google Scholar] [CrossRef]
- Shanahan, P.; O’Sullivan, J.; Tipton, K.F.; Kinsella, G.K.; Ryan, B.J.; Henehan, G.T.M. Theobromine and related methylxanthines as inhibitors of Primary Amine Oxidase. J. Food Biochem. 2018, 43, e12697. [Google Scholar] [CrossRef] [Green Version]
- Haj Ahmed, W.; Peiro, C.; Fontaine, J.; Ryan, B.J.; Kinsella, G.K.; O’Sullivan, J.; Grolleau, J.L.; Henehan, G.T.M.; Carpéné, C. Methylxanthines Inhibit Primary Amine Oxidase and Monoamine Oxidase Activities of Human Adipose Tissue. Medicines 2020, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Papukashvili, D.; Rcheulishvili, N.; Deng, Y. Attenuation of Weight Gain and Prevention of Associated Pathologies by Inhibiting SSAO. Nutrients 2020, 12, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papukashvili, D.; Rcheulishvili, N.; Deng, Y. Beneficial impact of Semicarbazide-Sensitive Amine Oxidase inhibition on the potential cytotoxicity of creatine supplementation in type 2 diabetes mellitus. Molecules 2020, 25, 2029. [Google Scholar] [CrossRef] [PubMed]
- Weston, C.J.; Shepherd, E.L.; Claridge, L.C.; Rantakari, P.; Curbishley, S.M.; Tomlinson, J.W.; Hubscher, S.G.; Reynolds, G.M.; Aalto, K.; Anstee, Q.M.; et al. Vascular adhesion protein-1 promotes liver inflammation and drives hepatic fibrosis. J. Clin. Investig. 2015, 125, 501–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonan, T.; Lukas, S.; Peet, G.W.; Pelletier, J.; Panzenbeck, M.; Hanidu, A.; Mazurek, S.; Wasti, R.; Rybina, I.; Roma, T.; et al. The oxidase activity of vascular adhesion protein-1 (VAP-1) is essential for function. Am. J. Clin. Exp. Immunol. 2013, 2, 172–185. [Google Scholar]
- Jargaud, V.; Bour, S.; Tercé, F.; Collet, X.; Valet, P.; Bouloumié, A.; Guillemot, J.-C.; Mauriège, P.; Jalkanen, S.; Stolen, C.; et al. Obesity of mice lacking VAP-1/SSAO by Aoc3 gene deletion is reproduced in mice expressing a mutated vascular adhesion protein-1 (VAP-1) devoid of amine oxidase activity. J. Physiol. Biochem. 2021, 77, 141–154. [Google Scholar] [CrossRef]
- Rosak, C.; Hittelman, K.J. Characterization of lipolytic responses of isolated white adipocytes from hamsters. Biochim. Biophys. Acta 1977, 496, 458–474. [Google Scholar] [CrossRef]
- Scotini, E.; Carpenedo, F.; Fassina, G. New derivatives of methyl-xanthines: Effect of thiocaffeine thiotheophylline and 8-phenyltheophylline on lipolysis and on phosphodiesterase activities. Pharmacol. Res. Commun. 1983, 15, 131–143. [Google Scholar] [CrossRef]
- Colitti, M.; Stefanon, B. Different anti-adipogenic effects of bio-compounds on primary visceral pre-adipocytes and adipocytes. EXCLI J. 2016, 15, 362–377. [Google Scholar] [CrossRef]
- Gomez-Zorita, S.; Belles, C.; Briot, A.; Fernandez-Quintela, A.; Portillo, M.P.; Carpéné, C. Pterostilbene Inhibits Lipogenic Activity similar to Resveratrol or Caffeine but Differently Modulates Lipolysis in Adipocytes. Phytother. Res. 2017, 31, 1273–1282. [Google Scholar] [CrossRef]
- Morin, N.; Lizcano, J.M.; Fontana, E.; Marti, L.; Smih, F.; Rouet, P.; Prévot, D.; Zorzano, A.; Unzeta, M.; Carpéné, C. Semicarbazide-sensitive amine oxidase substrates stimulate glucose transport and inhibit lipolysis in human adipocytes. J. Pharmacol. Exp. Ther. 2001, 297, 563–572. [Google Scholar]
- Fredholm, B.B.; Lindgren, E. The effect of alkylxanthines and other phosphodiesterase inhibitors on adenosine-receptor mediated decrease in lipolysis and cyclic AMP accumulation in rat fat cells. Acta Pharmacol. Toxicol. 1984, 54, 64–71. [Google Scholar] [CrossRef]
- Carpéné, C.; Mauriège, P.; Boulet, N.; Biron, S.; Grolleau, J.L.; Garcia-Barrado, M.J.; Iglesias-Osma, M.C. Methylamine Activates Glucose Uptake in Human Adipocytes Without Overpassing Action of Insulin or Stimulating its Secretion in Pancreatic Islets. Medicines 2019, 6, 89. [Google Scholar] [CrossRef] [Green Version]
- Carpéné, C.; Les, F.; Casedas, G.; Peiro, C.; Fontaine, J.; Chaplin, A.; Mercader, J.; Lopez, V. Resveratrol Anti-Obesity Effects: Rapid Inhibition of Adipocyte Glucose Utilization. Antioxidants 2019, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Willson, C. The clinical toxicology of caffeine: A review and case study. Toxicol. Rep. 2018, 5, 1140–1152. [Google Scholar] [CrossRef]
- Horrigan, L.A.; Kelly, J.P.; Connor, T.J. Immunomodulatory effects of caffeine: Friend or foe? Pharmacol. Ther. 2006, 111, 877–892. [Google Scholar] [CrossRef]
- Dray, C.; Daviaud, D.; Guigne, C.; Valet, P.; Castan-Laurell, I. Caffeine reduces TNFalpha up-regulation in human adipose tissue primary culture. J. Physiol. Biochem. 2007, 63, 329–336. [Google Scholar] [CrossRef]
- Wikoff, D.; Welsh, B.T.; Henderson, R.; Brorby, G.P.; Britt, J.; Myers, E.; Goldberger, J.; Lieberman, H.R.; O’Brien, C.; Peck, J.; et al. Systematic review of the potential adverse effects of caffeine consumption in healthy adults, pregnant women, adolescents, and children. Food Chem. Toxicol. 2017, 109, 585–648. [Google Scholar] [CrossRef]
- Beach, C.A.; Bianchine, J.R.; Gerber, N. The excretion of caffeine in the semen of men: Pharmacokinetics and comparison of the concentrations in blood and semen. J. Clin. Pharmacol. 1984, 24, 120–126. [Google Scholar] [CrossRef]
- Mandel, H.G. Update on caffeine consumption, disposition and action. Food Chem. Toxicol. 2002, 40, 1231–1234. [Google Scholar] [CrossRef]
- Martínez-López, S.; Sarriá, B.; Baeza, G.; Mateos, R.; Bravo-Clemente, L. Pharmacokinetics of caffeine and its metabolites in plasma and urine after consuming a soluble green/roasted coffee blend by healthy subjects. Food Res. Int. 2014, 64, 125–133. [Google Scholar] [CrossRef]
- Gans, J.H. Comparative toxicities of dietary caffeine and theobromine in the rat. Food Chem. Toxicol. 1984, 22, 365–369. [Google Scholar] [CrossRef]
- Sugiura, C.; Zheng, G.; Liu, L.; Sayama, K. Catechins and Caffeine Promote Lipid Metabolism and Heat Production through the Transformation of Differentiated 3T3-L1 Adipocytes from White to Beige Adipocytes. J. Food Sci. 2020, 85, 192–200. [Google Scholar] [CrossRef]
- Kim, A.R.; Yoon, B.K.; Park, H.; Seok, J.W.; Choi, H.; Yu, J.H.; Choi, Y.; Song, S.J.; Kim, A.; Kim, J.W. Caffeine inhibits adipogenesis through modulation of mitotic clonal expansion and the AKT/GSK3 pathway in 3T3-L1 adipocytes. BMB Rep. 2016, 49, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Lee, M.Y.; Park, H.M.; Park, Y.K.; Shon, J.C.; Liu, K.H.; Lee, C.H. Urine and serum metabolite profiling of rats fed a high-fat diet and the anti-obesity effects of caffeine consumption. Molecules 2015, 20, 3107–3128. [Google Scholar] [CrossRef] [Green Version]
- Emami, M.R.; Khorshidi, M.; Zarezadeh, M.; Safabakhsh, M.; Rezagholizadeh, F.; Alizadeh, S. Acute effects of caffeine ingestion on glycemic indices: A systematic review and meta-analysis of clinical trials. Complement. Ther. Med. 2019, 44, 282–290. [Google Scholar] [CrossRef]
- Battram, D.S.; Bugaresti, J.; Gusba, J.; Graham, T.E. Acute caffeine ingestion does not impair glucose tolerance in persons with tetraplegia. J. Appl. Physiol. (1985) 2007, 102, 374–381. [Google Scholar] [CrossRef] [Green Version]
- Elovaara, H.; Kidron, H.; Parkash, V.; Nymalm, Y.; Bligt, E.; Ollikka, P.; Smith, D.J.; Pihlavisto, M.; Salmi, M.; Jalkanen, S.; et al. Identification of two imidazole binding sites and key residues for substrate specificity in human primary amine oxidase AOC3. Biochemistry 2011, 50, 5507–5520. [Google Scholar] [CrossRef]
- Les, F.; Carpéné, C.; Arbones-Mainar, J.M.; Decaunes, P.; Valero, M.S.; Lopez, V. Pomegranate juice and its main polyphenols exhibit direct effects on amine oxidases from human adipose tissue and inhibit lipid metabolism in adipocytes. J. Funct. Foods 2017, 33, 323–331. [Google Scholar] [CrossRef]
- Carpéné, C.; Les, F.; Esteve, D.; Galitzky, J. Short-term effects of obestatin on hexose uptake and triacylglycerol breakdown in human subcutaneous adipocytes. World J. Diabetes 2018, 9, 25–32. [Google Scholar] [CrossRef]
- Carpéné, C.; Galitzky, J.; Belles, C.; Zakaroff-Girard, A. Mechanisms of the antilipolytic response of human adipocytes to tyramine, a trace amine present in food. J. Physiol. Biochem. 2018, 74, 623–633. [Google Scholar] [CrossRef]
- Leroux, M.; Lemery, T.; Boulet, N.; Briot, A.; Zakaroff, A.; Bouloumié, A.; Andrade, F.; Pérez-Matute, P.; Arbones-Mainar, J.M.; Carpéné, C. Effects of the amino acid derivatives, β-hydroxy-β-methylbutyrate, taurine, and N-methyltyramine, on triacylglycerol breakdown in fat cells. J. Physiol. Biochem. 2019, 75, 263–273. [Google Scholar] [CrossRef]
- Moody, A.J.; Stan, M.A.; Stan, M.; Gliemann, J. A simple free fat cell bioassay for insulin. Horm. Metab. Res. 1974, 6, 12–16. [Google Scholar] [CrossRef]
- Harant-Farrugia, I.; Garcia, J.; Iglesias-Osma, M.C.; Garcia-Barrado, M.J.; Carpéné, C. Is there an optimal dose for dietary linoleic acid? Lessons from essential fatty acid deficiency supplementation and adipocyte functions in rats. J. Physiol. Biochem. 2014, 70, 615–627. [Google Scholar] [CrossRef]
- Carpéné, C.; Gomez-Zorita, S.; Gupta, R.; Gres, S.; Rancoule, C.; Cadoudal, T.; Mercader, J.; Gomez, A.; Bertrand, C.; Iffiu-Soltesz, Z. Combination of low dose of the anti-adipogenic agents resveratrol and phenelzine in drinking water is not sufficient to prevent obesity in very-high-fat diet-fed mice. Eur. J. Nutr. 2014, 53, 1625–1635. [Google Scholar] [CrossRef] [PubMed]
- Carpéné, C.; Galitzky, J.; Saulnier-Blache, J.S. Short-term and rapid effects of lysophosphatidic acid on human adipose cell lipolytic and glucose uptake activities. AIMS Mol. Sci. 2016, 3, 222–237. [Google Scholar] [CrossRef]
- Marti, L.; Morin, N.; Enrique-Tarancon, G.; Prevot, D.; Lafontan, M.; Testar, X.; Zorzano, A.; Carpéné, C. Tyramine and vanadate synergistically stimulate glucose transport in rat adipocytes by amine oxidase-dependent generation of hydrogen peroxide. J. Pharmacol. Exp. Ther. 1998, 285, 342–349. [Google Scholar] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, W.H.; Boulet, N.; Briot, A.; Ryan, B.J.; Kinsella, G.K.; O’Sullivan, J.; Les, F.; Mercader-Barceló, J.; Henehan, G.T.M.; Carpéné, C. Novel Facet of an Old Dietary Molecule? Direct Influence of Caffeine on Glucose and Biogenic Amine Handling by Human Adipocytes. Molecules 2021, 26, 3831. https://doi.org/10.3390/molecules26133831
Ahmed WH, Boulet N, Briot A, Ryan BJ, Kinsella GK, O’Sullivan J, Les F, Mercader-Barceló J, Henehan GTM, Carpéné C. Novel Facet of an Old Dietary Molecule? Direct Influence of Caffeine on Glucose and Biogenic Amine Handling by Human Adipocytes. Molecules. 2021; 26(13):3831. https://doi.org/10.3390/molecules26133831
Chicago/Turabian StyleAhmed, Wiem Haj, Nathalie Boulet, Anaïs Briot, Barry J. Ryan, Gemma K. Kinsella, Jeffrey O’Sullivan, Francisco Les, Josep Mercader-Barceló, Gary T. M. Henehan, and Christian Carpéné. 2021. "Novel Facet of an Old Dietary Molecule? Direct Influence of Caffeine on Glucose and Biogenic Amine Handling by Human Adipocytes" Molecules 26, no. 13: 3831. https://doi.org/10.3390/molecules26133831
APA StyleAhmed, W. H., Boulet, N., Briot, A., Ryan, B. J., Kinsella, G. K., O’Sullivan, J., Les, F., Mercader-Barceló, J., Henehan, G. T. M., & Carpéné, C. (2021). Novel Facet of an Old Dietary Molecule? Direct Influence of Caffeine on Glucose and Biogenic Amine Handling by Human Adipocytes. Molecules, 26(13), 3831. https://doi.org/10.3390/molecules26133831