
Small Endogenous Ligands Modulation of Nerve Growth Factor Bioactivity: A Structural Biology Overview



Abstract
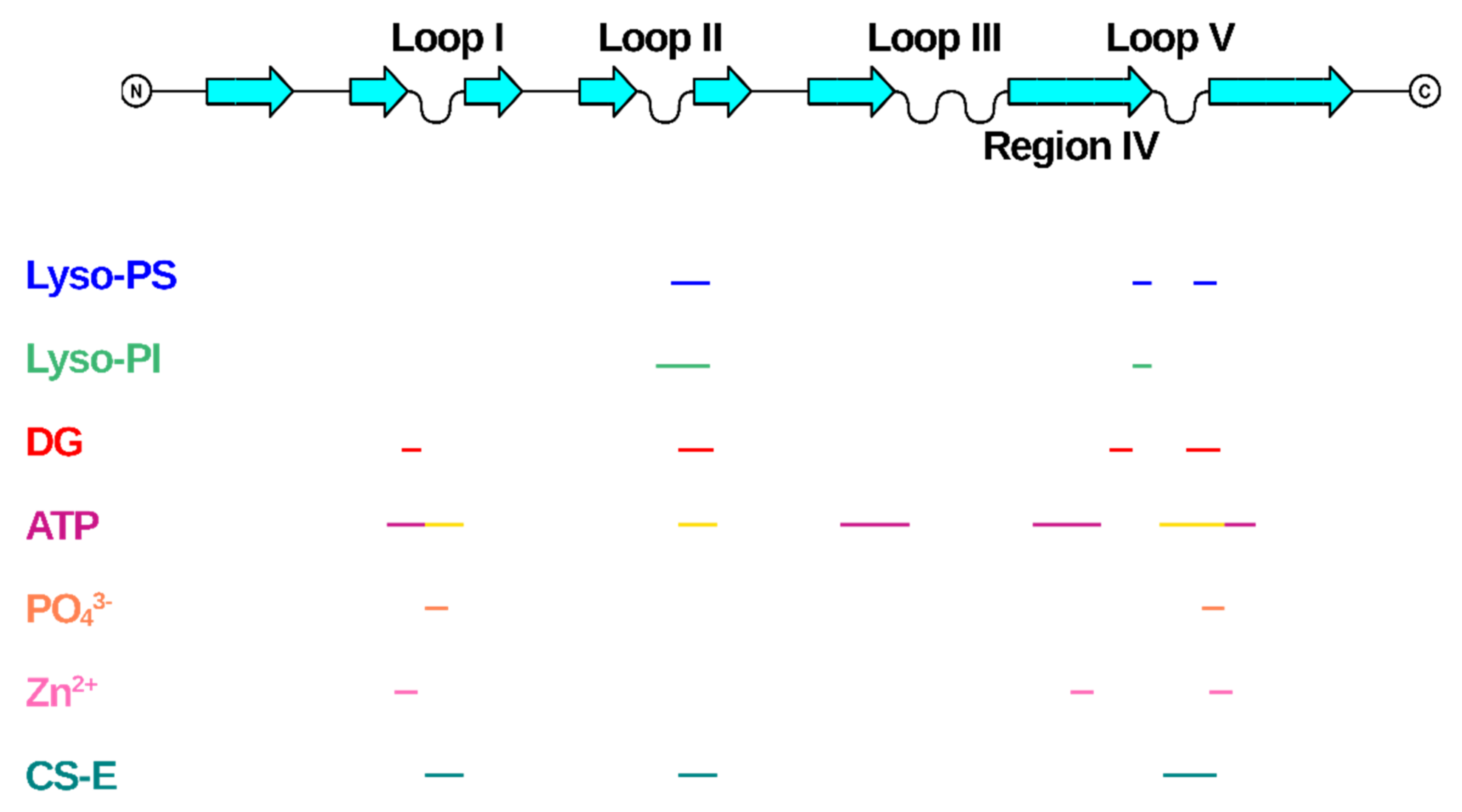
:1. Introduction
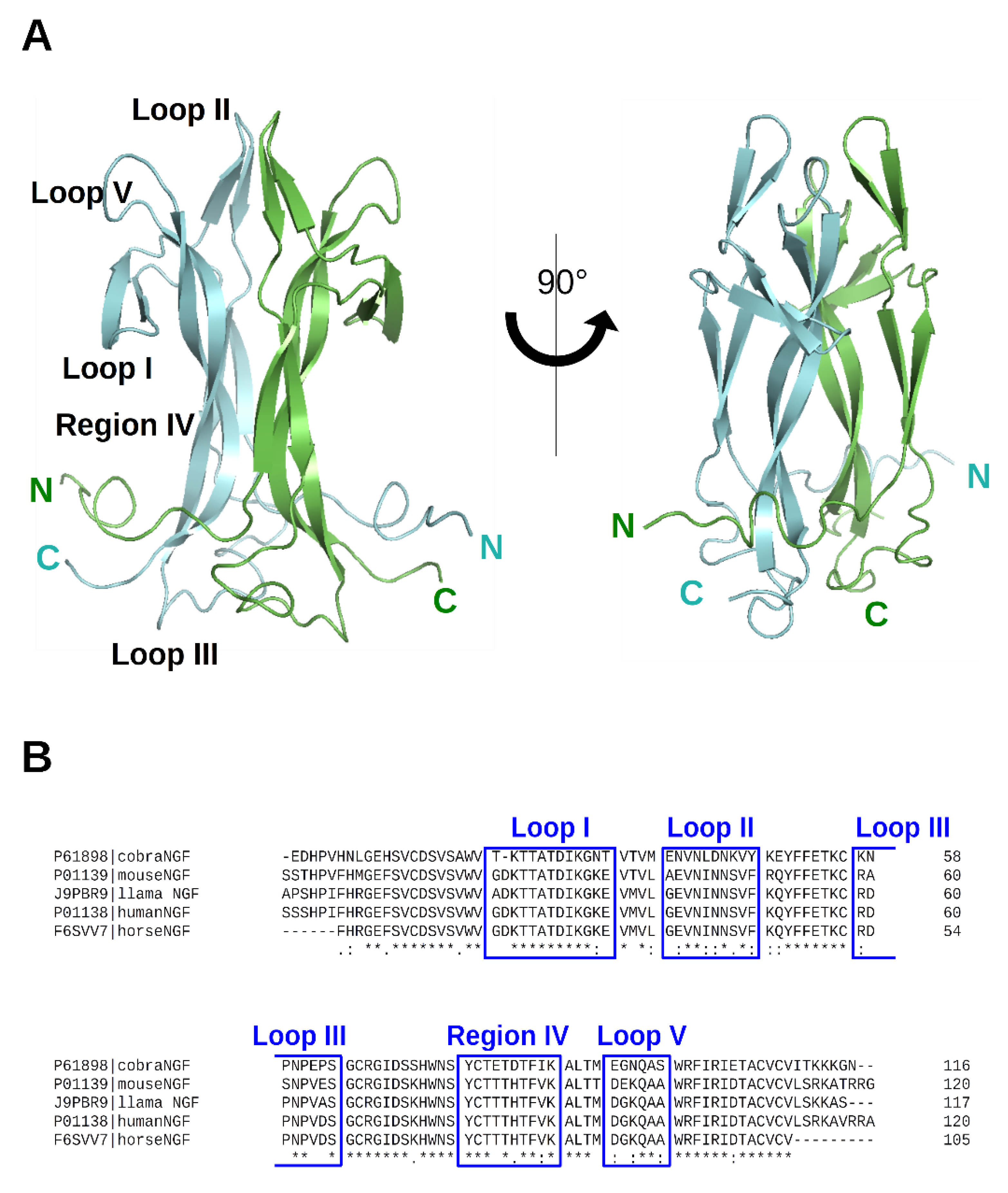
2. NGF and Lipid Molecules
2.1. Biological Context
2.2. Structural Data
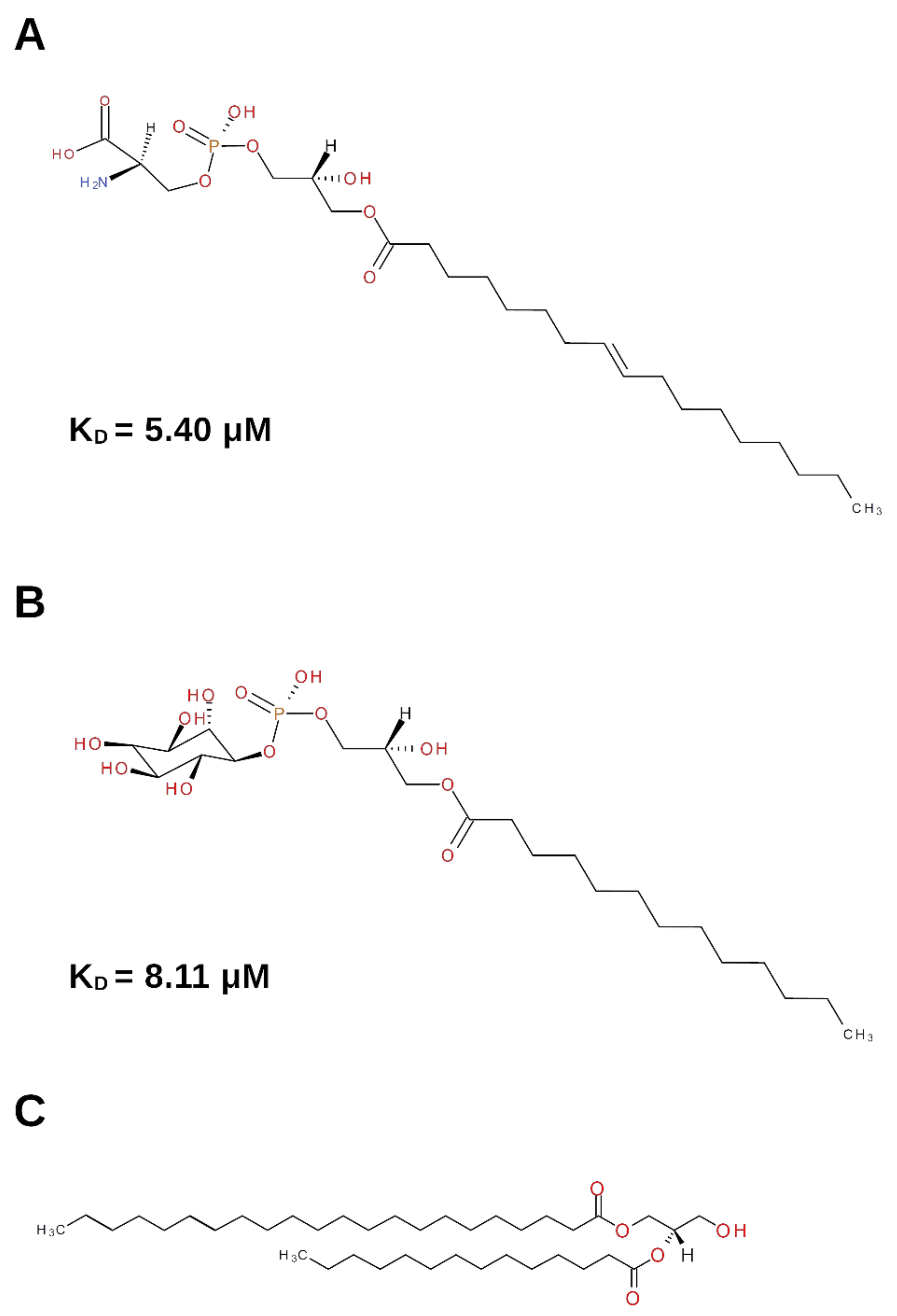
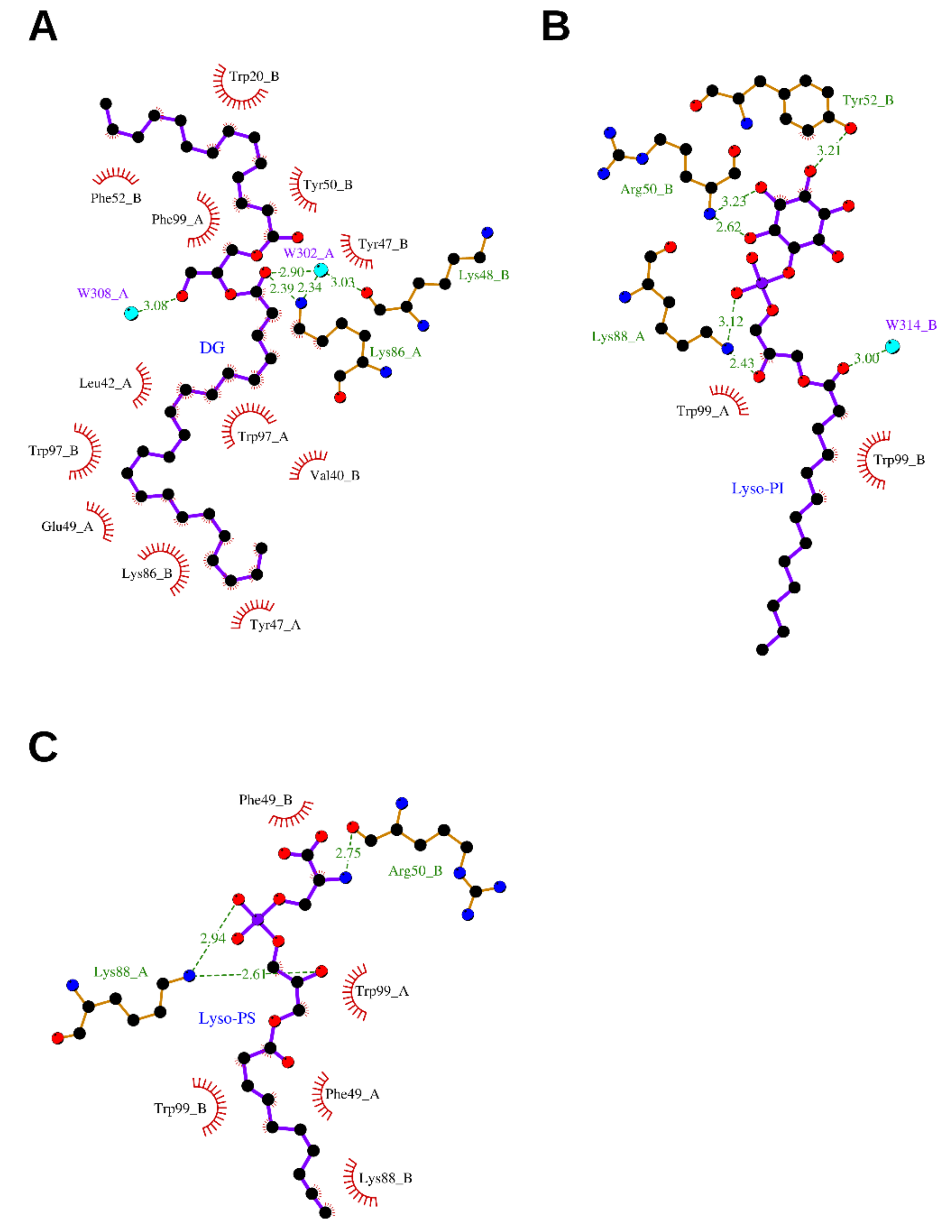
2.2.1. (2.S)-1-Hydroxy-3-(tetradecanoyloxy)propan-2-yl docosanoate—Diacylglycerol (DG)
2.2.2. (2.S)-2-Amino-3-[hydroxy-[(2R)-2-hydroxy-3-octadecanoyloxypropoxy]phosphoryl] oxypropanoic acid—Lysophosphatidylserine (Lyso-PS)
2.2.3. 1-Tridecanoyl-2-hydroxy-sn-glycero-3-phospho-(1′-myo-inositol)—Lysophosphatidylinositol (Lyso-PI)
3. NGF and Molecules Carrying Phosphate Groups
3.1. Biological Context
3.2. Structural Data
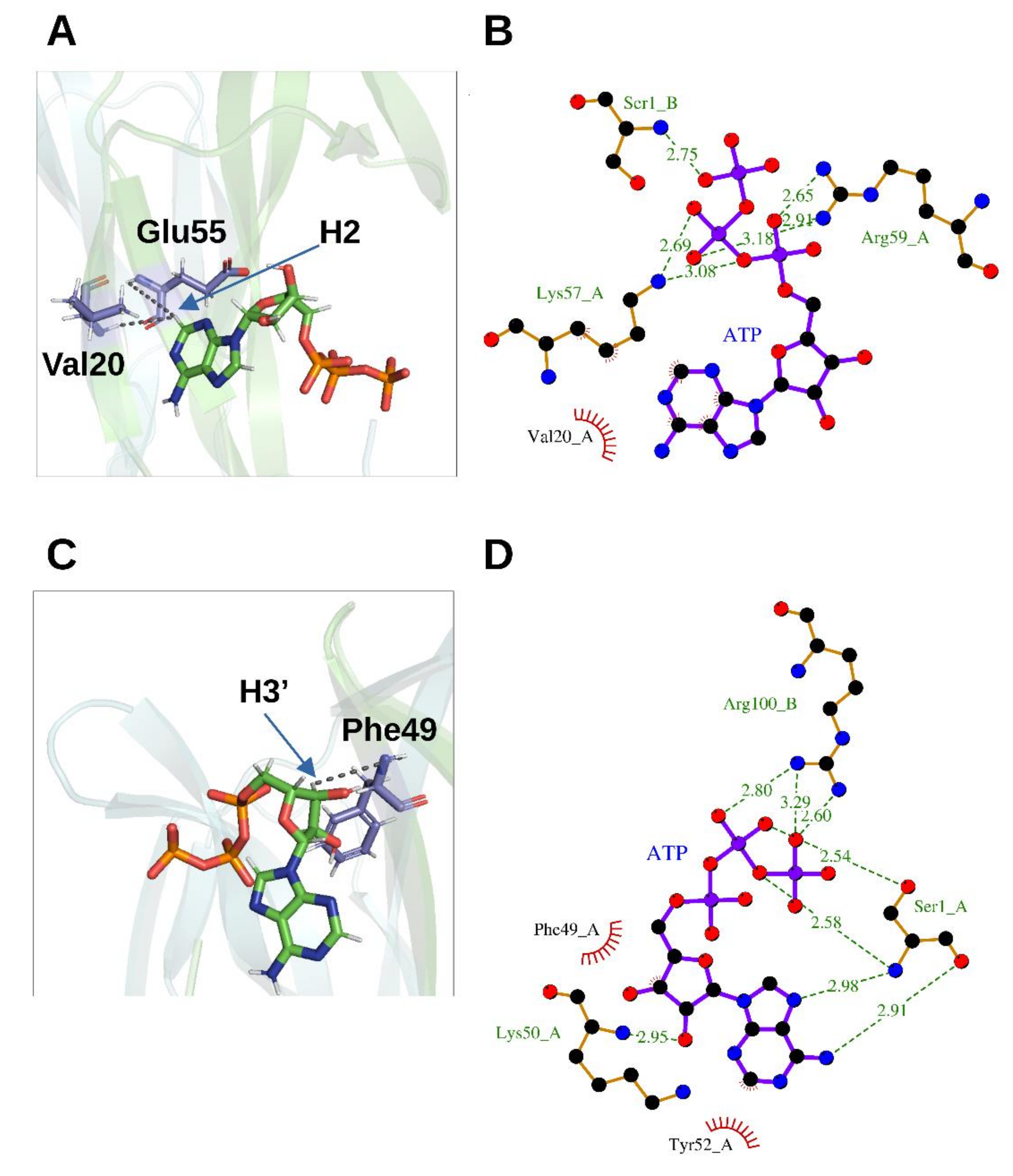
3.2.1. Adenosine-5′-Triphosphate (ATP)
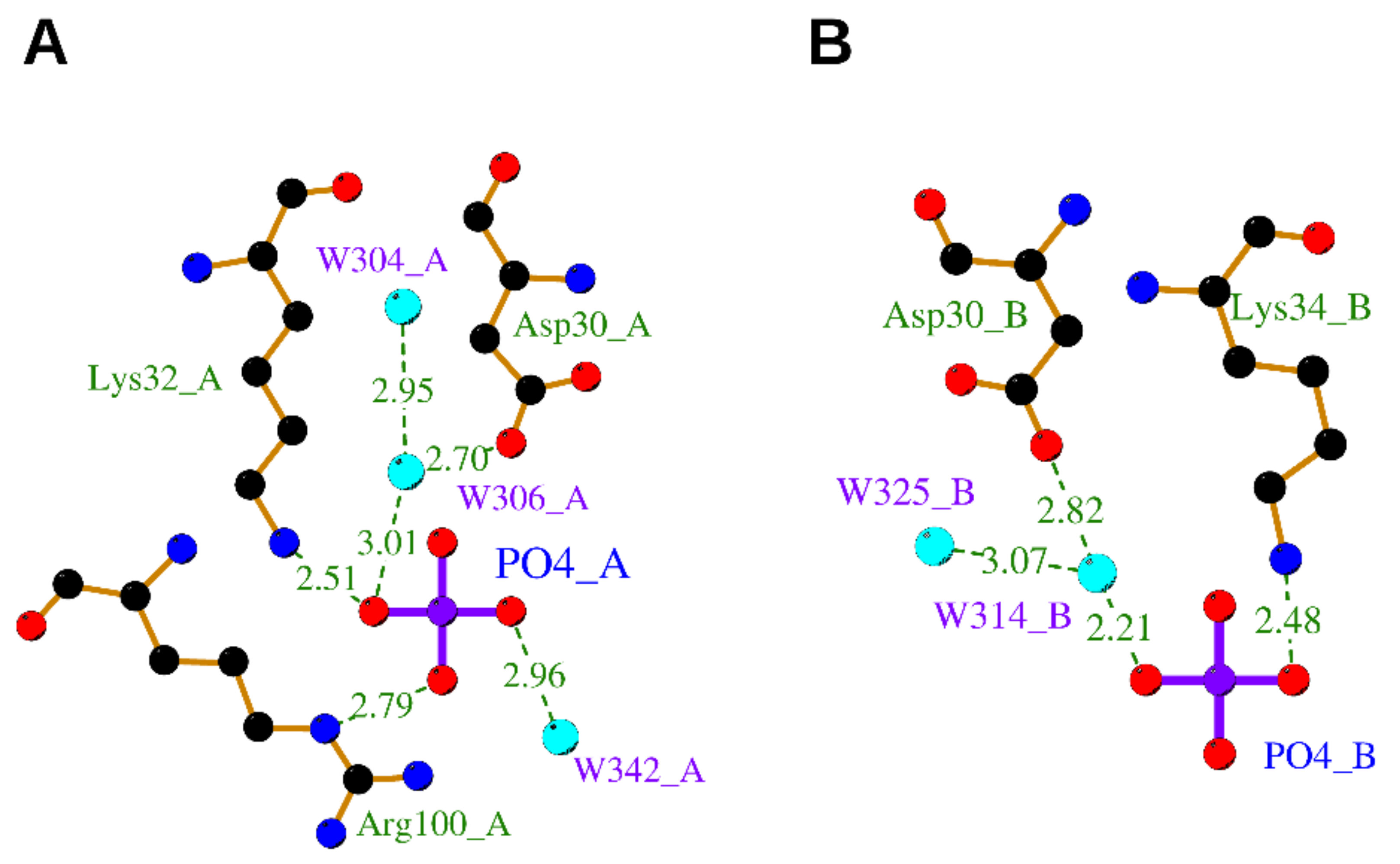
3.2.2. Phosphate Ion
4. NGF and Zinc
4.1. Biological Context
4.2. Structural Data
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levi-Montalcini, R. The nerve growth factor 35 years later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Capsoni, S.; Cattaneo, A. On the molecular basis linking nerve growth factor (NGF) to Alzheimer’s disease. Cell. Mol. Neurobiol. 2006, 26, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, J.H.; Tewari, D.; McMahon, S.B. Neurotrophic factors and their inhibitors in chronic pain treatment. Neurobiol. Dis. 2017, 97, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, F.; Merzel, F.; Cassetta, A.; Ogris, I.; Covaceuszach, S.; Grdadolnik, J.; Lamba, D.; Golič Grdadolnik, S. Endogenous modulators of neurotrophin signaling: Landscape of the transient ATP-NGF interactions. Comput. Struct. Biotechnol. J. 2021, 19, 2938–2949. [Google Scholar] [CrossRef]
- Tong, Q.; Wang, F.; Zhou, H.-Z.; Sun, H.-L.; Song, H.; Shu, Y.-Y.; Gong, Y.; Zhang, W.; Cai, T.; Yang, F.; et al. Structural and functional insights into lipid-bound nerve growth factors. FASEB J. 2012, 26, 3811–3821. [Google Scholar] [CrossRef]
- Sun, H.L.; Jiang, T. The structure of nerve growth factor in complex with lysophosphatidylinositol. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 906–912. [Google Scholar] [CrossRef] [Green Version]
- Ratto, M.H.; Leduc, Y.A.; Valderrama, X.P.; van Straaten, K.E.; Delbaere, L.T.J.; Pierson, R.A.; Adams, G.P. The nerve of ovulation-inducing factor in semen. Proc. Natl. Acad. Sci. USA 2012, 109, 15042–15047. [Google Scholar] [CrossRef] [Green Version]
- Maitra, R.; Shamovsky, I.L.; Wang, W.; Solc, M.; Lawrance, G.; Dostaler, S.M.; Ross, G.M.; Riopelle, R.J. Differential effects of transition metal cations on the conformation and biological activities of nerve growth factor. Neurotox. Res. 2000, 2, 321–341. [Google Scholar] [CrossRef]
- Holland, D.R.; Cousens, L.S.; Meng, W.; Matthews, B.W. Nerve Growth factor in different crystal forms displays structural flexibility and reveals zinc binding sites. J. Mol. Biol. 1994, 239, 385–400. [Google Scholar] [CrossRef]
- Kalnins, G.; Tars, K. Mature horse nerve growth factor at 2.8 angstrom resolution. RCSB PDB Protein Data Bank. To be published. Available online: https://www.rcsb.org/structure/6XUO (accessed on 8 December 2021).
- Piñeiro, R.; Falasca, M. Lysophosphatidylinositol signalling: New wine from an old bottle. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2012, 1821, 694–705. [Google Scholar] [CrossRef] [Green Version]
- Makide, K.; Uwamizu, A.; Shinjo, Y.; Ishiguro, J.; Okutani, M.; Inoue, A.; Aoki, J. Novel lysophosphoplipid receptors: Their structure and function. J. Lipid Res. 2014, 55, 1986–1995. [Google Scholar] [CrossRef] [Green Version]
- Shanbhag, K.; Mhetre, A.; Khandelwal, N.; Kamat, S.S. The lysophosphatidylserines—An emerging class of signalling lysophospholipids. J. Membr. Biol. 2020, 253, 381–397. [Google Scholar] [CrossRef]
- Omi, J.; Kano, K.; Aoki, J. Current knowledge on the biology of lysophosphatidylserine as an emerging bioactive lipid. Cell Biochem. Biophys. 2021, 79, 497–508. [Google Scholar] [CrossRef]
- Bruni, A.; Bigon, E.; Boarato, E.; Mietto, L.; Leon, A.; Toffano, G. Interaction between nerve growth factor and lysophosphatidylserine on rat peritoneal mast cells. FEBS Lett. 1982, 138, 190–192. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, N.; Weskamp, G.; Erne, P.; Otten, U. Nerve growth factor induces mast cell degranulation without changing intracellular calcium levels. FEBS Lett. 1986, 198, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Pearce, F.L.; Thompson, H.L. Some characteristics of histamine secretion from rat peritoneal mast cells stimulated with nerve growth factor. J. Physiol. 1986, 372, 379–393. [Google Scholar] [CrossRef] [Green Version]
- Seebeck, J.; Westenberger, K.; Elgeti, T.; Ziegler, A.; Schütze, S. The exocytotic signaling pathway induced by nerve growth factor in the presence of lyso-phosphatidylserine in rat peritoneal mast cells involves a type D phospholipase. Regul. Pept. 2001, 102, 93–99. [Google Scholar] [CrossRef]
- Horigome, K.; Pryor, J.C.; Bullock, E.D.; Johnson, E.M. Mediator release from mast cells by nerve growth factor. Neurotrophin specificity and receptor mediation. J. Biol. Chem. 1993, 268, 14881–14887. [Google Scholar] [CrossRef]
- Kawamoto, K.; Aoki, J.; Tanaka, A.; Itakura, A.; Hosono, H.; Arai, H.; Kiso, Y.; Matsuda, H. Nerve growth factor activates mast cells through the collaborative interaction with lysophosphatidylserine expressed on the membrane surface of activated platelets. J. Immunol. 2002, 168, 6412–6419. [Google Scholar] [CrossRef] [Green Version]
- Lourenssen, S.; Blennerhassett, M.G. Lysophosphatidylserine potentiates nerve growth factor-induced differentiation of PC12 cells. Neurosci. Lett. 1998, 248, 77–80. [Google Scholar] [CrossRef]
- Alhouayek, M.; Masquelier, J.; Muccioli, G.G. Lysophosphatidylinositols, from cell membrane constituents to GPR55 ligands. Trends Pharmacol. Sci. 2018, 39, 586–604. [Google Scholar] [CrossRef]
- Obara, Y.; Ueno, S.; Yanagihata, Y.; Nakahata, N. Lysophosphatidylinositol causes neurite retraction via GPR55, G13 and RhoA in PC12 cells. PLoS ONE 2011, 6, e24284. [Google Scholar] [CrossRef] [Green Version]
- Wuhanqimuge; Itakura, A.; Matsuki, Y.; Tanaka, M.; Arioka, M. Lysophosphatidylcholine enhances NGF-induced MAPK and Akt signals through the extracellular domain of TrkA in PC12 cells. FEBS Open Bio 2013, 3, 243–251. [Google Scholar] [CrossRef] [Green Version]
- McDonald, N.Q.; Lapatto, R.; Murray-Rust, J.; Gunning, J.; Wlodawer, A.; Blundell, T.L. New protein fold revealed by a 2.3-Å resolution crystal structure of nerve growth factor. Nature 1991, 354, 411–414. [Google Scholar] [CrossRef]
- Ibáñez, C.F. Neurotrophic factors: From structure-function studies to designing effective therapeutics. Trends Biotechnol. 1995, 13, 217–227. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System; Version 2.0; Schrödinger, LLC.: New York, NY, USA.
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Klumpp, S.; Kriha, D.; Bechmann, G.; Maassen, A.; Maier, S.; Pallast, S.; Hoell, P.; Krieglstein, J. Phosphorylation of the growth factors bFGF, NGF and BDNF: A prerequisite for their biological activity. Neurochem. Int. 2006, 48, 131–137. [Google Scholar] [CrossRef]
- König, S.; Hasche, A.; Pallast, S.; Krieglstein, J.; Klumpp, S. Detection of ATP-binding to growth factors. J. Am. Soc. Mass Spectrom. 2008, 19, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Hasche, A.; Ferenz, K.B.; Rose, K.; König, S.; Humpf, H.-U.; Klumpp, S.; Krieglstein, J. Binding of ATP to nerve growth factor: Characterization and relevance for bioactivity. Neurochem. Int. 2010, 56, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Ferenz, K.B.; Rose, K.; König, S.; Krieglstein, J. ATP–NGF-complex, but not NGF, is the neuroprotective ligand. Neurochem. Int. 2011, 59, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Ferenz, K.B.; Gast, R.E.; Rose, K.; Finger, I.E.; Hasche, A.; Krieglstein, J. Nerve growth factor and brain-derived neurotrophic factor but not granulocyte colony-stimulating factor, nimodipine and dizocilpine, require ATP for neuroprotective activity after oxygen–glucose deprivation of primary neurons. Brain Res. 2012, 1448, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rose, K. Interaction of ATP with fibroblast growth factor 2: Biochemical characterization and consequence for growth factor stability. BMC Biochem. 2011, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Gast, R.E.; König, S.; Rose, K.; Ferenz, K.B.; Krieglstein, J. Binding of ATP to vascular endothelial growth factor isoform VEGF-A165 is essential for inducing proliferation of human umbilical vein endothelial cells. BMC Biochem. 2011, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. An introduction to the roles of purinergic signalling in neurodegeneration, neuroprotection and neuroregeneration. Neuropharmacology 2016, 104, 4–17. [Google Scholar] [CrossRef]
- Volonte, C.; Amadio, S.; Cavaliere, F.; D’Ambrosi, N.; Vacca, F.; Bernardi, G. Extracellular ATP and neurodegeneration. Curr. Drug Target. CNS Neurol. Disord. 2003, 2, 403–412. [Google Scholar] [CrossRef]
- Cavaliere, F.; Sancesario, G.; Bernardi, G.; Volonté, C. Extracellular ATP and nerve growth factor intensify hypoglycemia-induced cell death in primary neurons: Role of P2 and NGFRp75 receptors. J. Neurochem. 2002, 83, 1129–1138. [Google Scholar] [CrossRef] [Green Version]
- D’Ambrosi, N.; Murra, B.; Cavaliere, F.; Amadio, S.; Bernardi, G.; Burnstock, G.; Volonté, C. Interaction between ATP and nerve growth factor signalling in the survival and neuritic outgrowth from PC12 cells. Neuroscience 2001, 108, 527–534. [Google Scholar] [CrossRef]
- McConkey, E.H. Molecular evolution, intracellular organization, and the quinary structure of proteins. Proc. Natl. Acad. Sci. USA 1982, 79, 3236–3240. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.; Ebbinghaus, S.; Marcos, J.C. Protein folding and quinary interactions: Creating cellular organisation through functional disorder. FEBS Lett. 2018, 592, 3040–3053. [Google Scholar] [CrossRef] [Green Version]
- Cohen, R.D.; Pielak, G.J. A cell is more than the sum of its (dilute) parts: A brief history of quinary structure. Protein Sci. 2017, 26, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Heo, C.E.; Han, J.Y.; Lim, S.; Lee, J.; Im, D.; Lee, M.J.; Kim, Y.K.; Kim, H.I. ATP kinetically modulates pathogenic tau fibrillations. ACS Chem. Neurosci. 2020, 11, 3144–3152. [Google Scholar] [CrossRef]
- Kang, J.; Lim, L.; Song, J. ATP binds and inhibits the neurodegeneration-associated fibrillization of the FUS RRM domain. Commun. Biol. 2019, 2, 223. [Google Scholar] [CrossRef] [Green Version]
- Dang, M.; Li, Y.; Song, J. ATP biphasically modulates LLPS of SARS-CoV-2 nucleocapsid protein and specifically binds its RNA-binding domain. Biochem. Biophys. Res. Commun. 2021, 541, 50–55. [Google Scholar] [CrossRef]
- Counts, S.E.; Mufson, E.J. The role of nerve growth factor receptors in cholinergic basal forebrain degeneration in prodromal Alzheimer disease. J. Neuropathol. Exp. Neurol. 2005, 64, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Woods, L.T.; Ajit, D.; Camden, J.M.; Erb, L.; Weisman, G.A. Purinergic receptors as potential therapeutic targets in Alzheimer’s disease. Neuropharmacology 2016, 104, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Martín, F.J.; Valera, E.; Casimiro, I.; Merino, J.M. Nerve growth factor increases the sensitivity to zinc toxicity and induces cell cycle arrest in PC12 cells. Brain Res. Bull. 2010, 81, 458–466. [Google Scholar] [CrossRef]
- Kawahara, M.; Mizuno, D.; Koyama, H.; Konoha, K.; Ohkawara, S.; Sadakane, Y. Disruption of zinc homeostasis and the pathogenesis of senile dementia. Metallomics 2014, 6, 209–219. [Google Scholar] [CrossRef]
- Shuttleworth, C.W.; Weiss, J.H. Zinc: New clues to diverse roles in brain ischemia. Trends Pharmacol. Sci. 2011, 32, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Travaglia, A.; Pietropaolo, A.; La Mendola, D.; Nicoletti, V.G.; Rizzarelli, E. The inorganic perspectives of neurotrophins and Alzheimer’s disease. J. Inorg. Biochem. 2012, 111, 130–137. [Google Scholar] [CrossRef]
- Ross, G.M.; Shamovsky, I.L.; Lawrance, G.; Solc, M.; Dostaler, S.M.; Jimmo, S.L.; Weaver, D.; Riopelle, R.J. Zinc alters conformation and inhibits biological activities of nerve growth factor and related neurotrophins. Nat. Med. 1997, 3, 872–878. [Google Scholar] [CrossRef]
- Yang, T.H.; Cleland, J.L.; Lam, X.; Meyer, J.D.; Jones, L.S.; Randolph, T.W.; Manning, M.C.; Carpenter, J.F. Effect of zinc binding and precipitation on structures of recombinant human growth hormone and nerve growth factor. J. Pharm. Sci. 2000, 89, 1480–1485. [Google Scholar] [CrossRef]
- Zhao, G.-H.; Yu, P.; Hu, X.-S.; Zhao, L. Effect of Zn(II) on the structure and biological activity of natural β-NGF. Acta Biochim. Biophys. Sin. 2004, 36, 99–104. [Google Scholar] [CrossRef]
- Pattison, S.E.; Dunn, M.F. Relation of zinc ion to the structure and function of the 7S nerve growth factor protein. Biochemistry 1975, 14, 2733–2739. [Google Scholar] [CrossRef]
- Young, M.; Blanchard, M.H.; Sessions, F.; Boyle, M.D. Subunit structure of high molecular weight mouse nerve growth factor. Biochemistry 1988, 27, 6675–6681. [Google Scholar] [CrossRef]
- Young, M.; Koroly, M.J. Nerve growth factor zymogen. Stoichiometry of the active-site serine and role of zinc(II) in controlling autocatalytic self-activation. Biochemistry 1980, 19, 5316–5321. [Google Scholar] [CrossRef]
- Rao, A.G.; Neet, K.E. Subunit interactions of 7 S nerve growth factor. Gamma-esterase activity, rates, and conformational changes during reassociation. J. Biol. Chem. 1984, 259, 73–79. [Google Scholar] [CrossRef]
- Bax, B.; Blundell, T.L.; Murray-Rust, J.; McDonald, N.Q. Structure of mouse 7S NGF: A complex of nerve growth factor with four binding proteins. Structure 1997, 5, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Dunn, M.F.; Pattison, S.E.; Storm, M.C.; Quiel, E. Comparison of the zinc binding domains in the 7S nerve growth factor and the zinc-insulin hexamer. Biochemistry 1980, 19, 718–725. [Google Scholar] [CrossRef]
- Wang, W.; Post, J.I.; Dow, K.E.; Shin, S.H.; Riopelle, R.J.; Ross, G.M. Zinc and copper inhibit nerve growth factor-mediated protection from oxidative stress-induced apoptosis. Neurosci. Lett. 1999, 259, 115–118. [Google Scholar] [CrossRef]
- Allington, C.; Shamovsky, I.L.; Ross, G.M.; Riopelle, R.J. Zinc inhibits p75NTR-mediated apoptosis in chick neural retina. Cell Death Differ. 2001, 8, 451–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibáñez, C.F.; Ilag, L.L.; Murray-Rust, J.; Persson, H. An extended surface of binding to Trk tyrosine kinase receptors in NGF and BDNF allows the engineering of a multifunctional pan-neurotrophin. EMBO J. 1993, 12, 2281–2293. [Google Scholar] [CrossRef] [PubMed]
- Bond, C.S. TopDraw: A sketchpad for protein structure topology cartoons. Bioinformatics 2003, 19, 311–312. [Google Scholar] [CrossRef] [Green Version]
- Rogers, C.J.; Clark, P.M.; Tully, S.E.; Abrol, R.; Garcia, K.C.; Goddard, W.A.; Hsieh-Wilson, L.C. Elucidating glycosaminoglycan-protein-protein interactions using carbohydrate microarray and computational approaches. Proc. Natl. Acad. Sci. USA 2011, 108, 9747–9752. [Google Scholar] [CrossRef] [Green Version]
- Pulsipher, A.; Griffin, M.E.; Stone, S.E.; Brown, J.M.; Hsieh-Wilson, L.C. Directing neuronal signaling through cell-surface glycan engineering. J. Am. Chem. Soc. 2014, 136, 6794–6797. [Google Scholar] [CrossRef]
- Miller, G.M.; Hsieh-Wilson, L.C. Sugar-dependent modulation of neuronal development, regeneration, and plasticity by chondroitin sulfate proteoglycans. Exp. Neurol. 2015, 274, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, F.; Covaceuszach, S.; Konarev, P.; Gonfloni, S.; Malerba, F.; Schwarz, E.; Svergun, D.; Cattaneo, A.; Lamba, D. Intrinsic structural disorder of mouse proNGF. Proteins: Struct. Funct. Bioinform. 2009, 75, 990–1009. [Google Scholar] [CrossRef]
- Paoletti, F.; Malerba, F.; Kelly, G.; Noinville, S.; Lamba, R.; Cattaneo, A.; Pastore, A. Conformational plasticity of proNGF. PLoS ONE 2011, 6, e22615. [Google Scholar] [CrossRef]
- Yan, R.; Yalinca, H.; Paoletti, F.; Gobbo, F.; Marchetti, L.; Kuzmanic, A.; Lamba, D.; Gervasio, F.L.; Konarev, P.V.; Cattaneo, A.; et al. The structure of the pro-domain of mouse proNGF in contact with the NGF domain. Structure 2019, 27, 78–89.e3. [Google Scholar] [CrossRef] [Green Version]
- Trabjerg, E.; Kartberg, F.; Christensen, S.; Rand, K.D. Conformational characterization of nerve growth factor-β reveals that its regulatory pro-part domain stabilizes three loop regions in its mature part. J. Biol. Chem. 2017, 292, 16665–16676. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Ligand | NGF Species | PDB Entry | Figure | Reference for the Structure | Biological Effect of Ligand/NGF Interaction |
|---|---|---|---|---|---|
| Lyso-PS (lysophosphatidyl-serine) | Mouse | 4EAX |  | [6] | Activation of histamine secretion. Through platelet activation, role in chronic inflammation and wound healing. Enhanced NGF-induced neurite outgrowth in PC12 cells. |
| Lyso-PI (lysophosphatidyl-inositol) | Mouse | 4XPJ |  | [7] | Lyso-PI induces neurite retraction through GPR55 receptor in NGF-differentiated PC12 cells. |
| (2S)-1-hydroxy-3-(tetradecanoyloxy) propan-2-yl docosanoate | Cobra | 4EC7 |  | [6] | Role in histamine release. |
| (PO4)3− | Llama | 4EFV |  | [8] | ATP binding proven to be protective towards neuronal cell death. |
| Zn2+ | Mouse | 1BTG |  | [10] | Induction of secondary structure changes on NGF. Inhibition of TrkA-and p75NTR-mediated effects. |
| Zn2+ | Horse | 6XUO |  | [11] | To be published. |
| Crystal Form | Ion | Protein Ligand | Distance (Å) |
|---|---|---|---|
| P212121 | Zn2+ (1) | His84 (Protomer 1) | 2.4 |
| Asp105 (Protomer 1) | 2.1 | ||
| Zn2+ (2) | His84 (Protomer 2) | 2.3 | |
| Asp105 (Protomer 2) | 2.0 | ||
| C2 | Zn2+ (1) | His 84 (Protomer 3) | 2.0 |
| Asp 105 (Protomer 3) | 2.3 | ||
| Zn2+ (2) | Glu 94 (Protomer 1) | 1.9 | |
| Asp 105 (Protomer 3) | 1.9 | ||
| Zn2+ (3) | His 84 (Protomer 2) | 2.0 | |
| Asp 105 (Protomer 2) | 2.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paoletti, F.; Lamba, D. Small Endogenous Ligands Modulation of Nerve Growth Factor Bioactivity: A Structural Biology Overview. Cells 2021, 10, 3462. https://doi.org/10.3390/cells10123462
Paoletti F, Lamba D. Small Endogenous Ligands Modulation of Nerve Growth Factor Bioactivity: A Structural Biology Overview. Cells. 2021; 10(12):3462. https://doi.org/10.3390/cells10123462
Chicago/Turabian StylePaoletti, Francesca, and Doriano Lamba. 2021. "Small Endogenous Ligands Modulation of Nerve Growth Factor Bioactivity: A Structural Biology Overview" Cells 10, no. 12: 3462. https://doi.org/10.3390/cells10123462
APA StylePaoletti, F., & Lamba, D. (2021). Small Endogenous Ligands Modulation of Nerve Growth Factor Bioactivity: A Structural Biology Overview. Cells, 10(12), 3462. https://doi.org/10.3390/cells10123462