2. Chemical, Nutritional, and Technological Interest of DFAs
Besides its traditional involvement in the preparation of homemade products such as pastries, caramelization is nowadays an important industrial process used for the production of food materials and colouring additives, mainly from sucrose but also from
d-glucose, maltose, maltodextrins and starch syrups [
20,
21]. Products having various properties are commercially available, depending on the starting material and the temperature and duration of the heating process. Food grade acids are frequently used as caramelization promoters. In any case, caramelization results in pH lowering, which self-catalyses the caramelization process. Protonic activation of the sugar precursor then occurs, leading to formation of volatiles (e.g. 2-hydromethylfurfural, HMF), coloured polymers (melanoidins) and a major fraction of oligosaccharidic nature whose structure remained unknown until recent times [
18,
19,
20,
21].
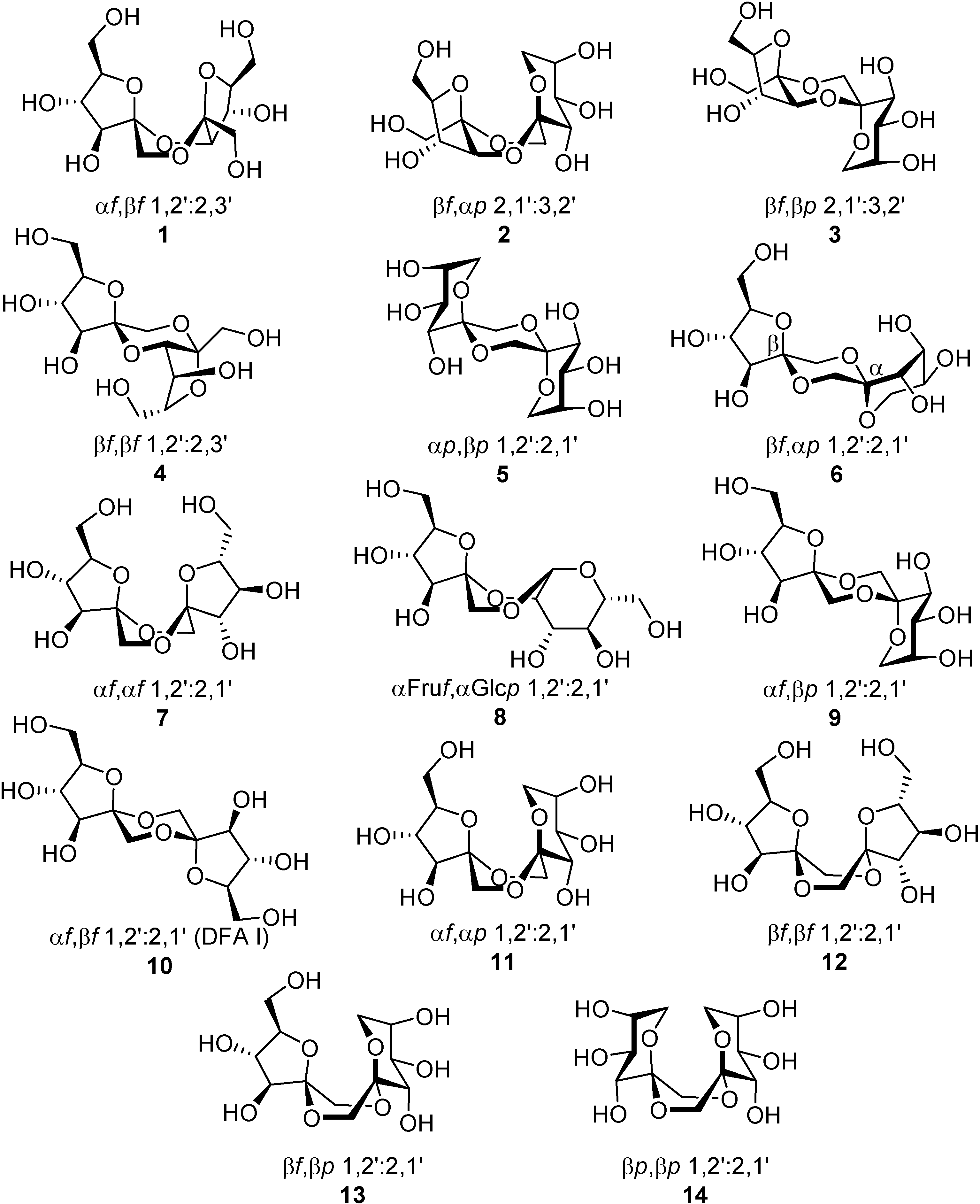
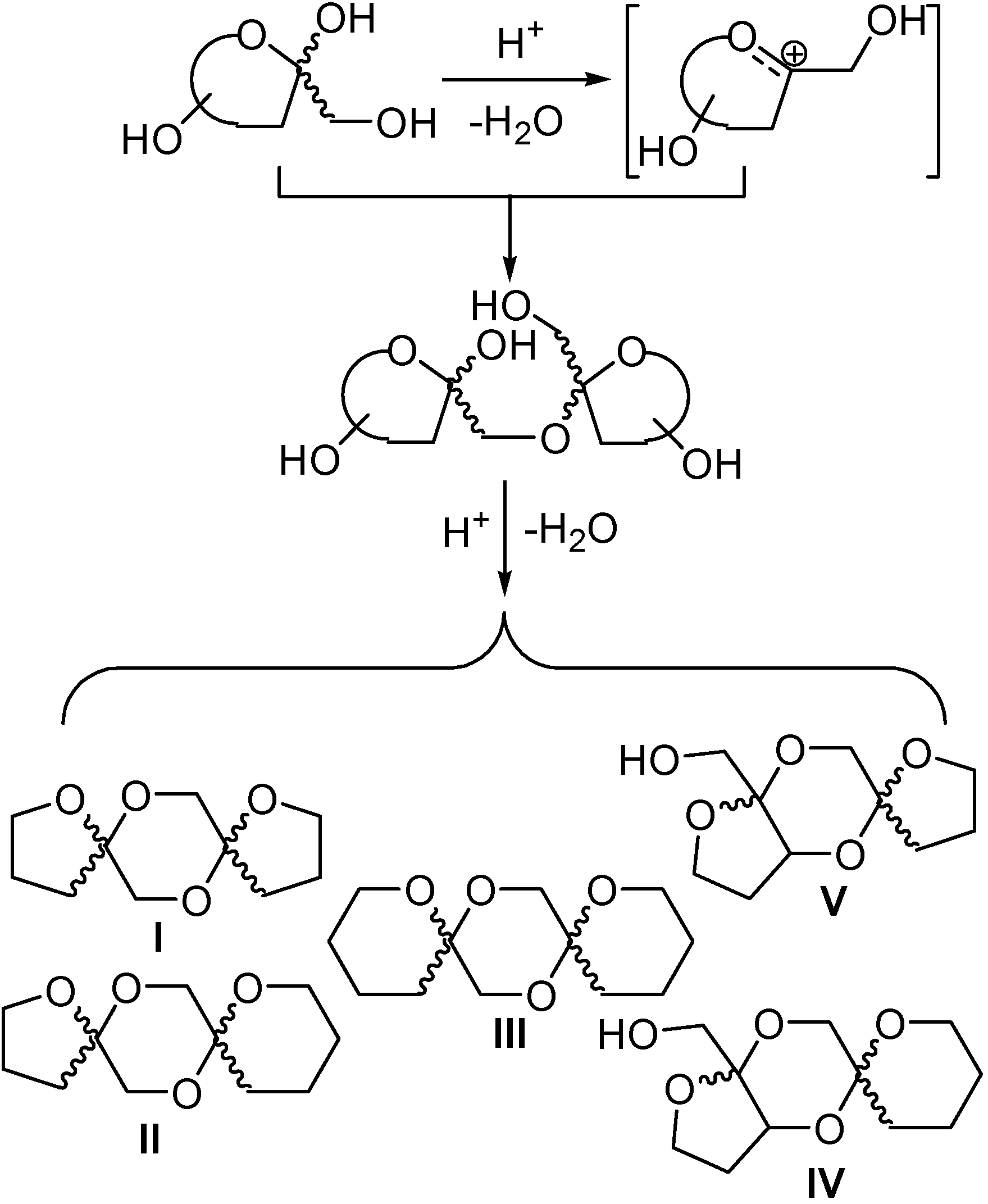
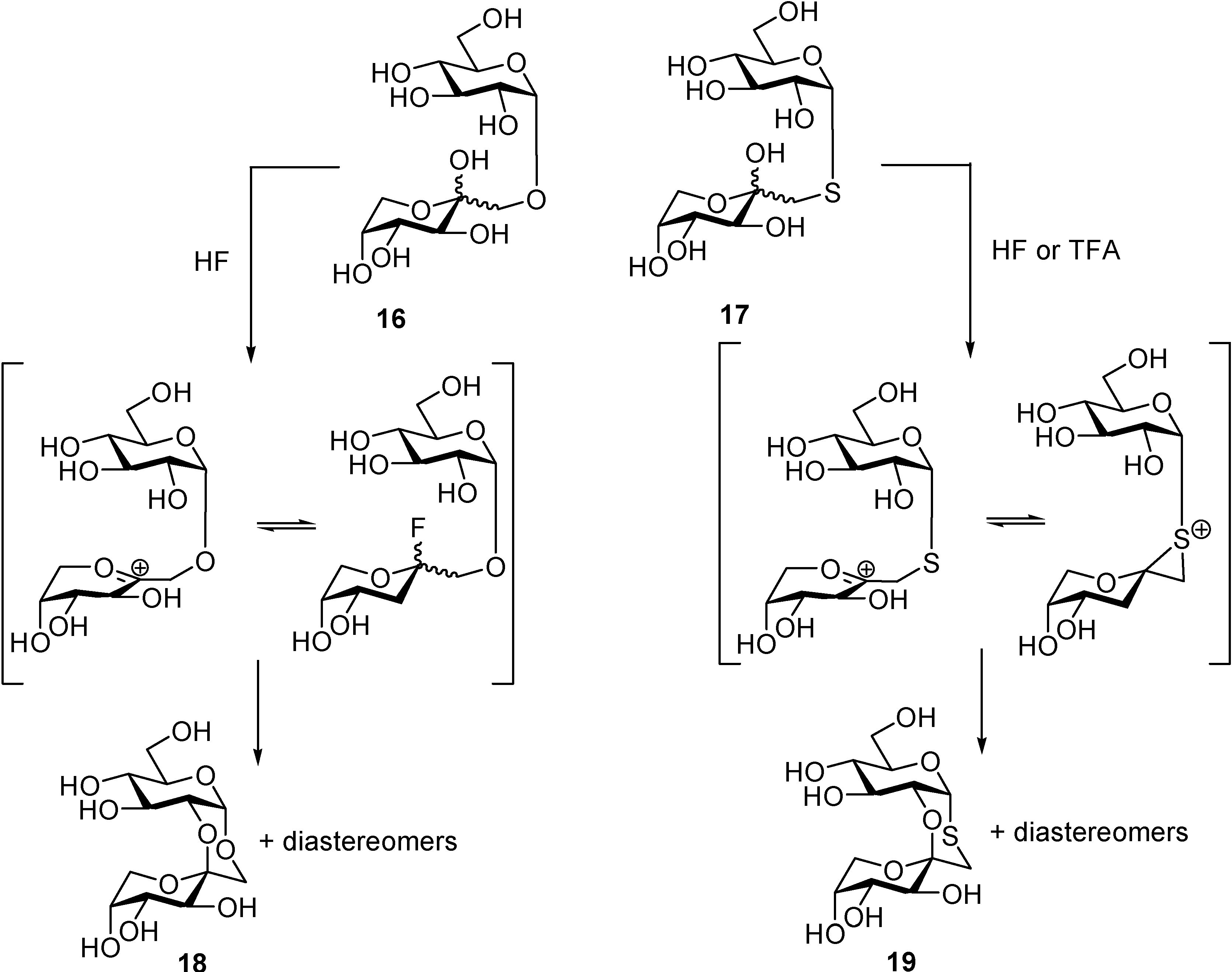
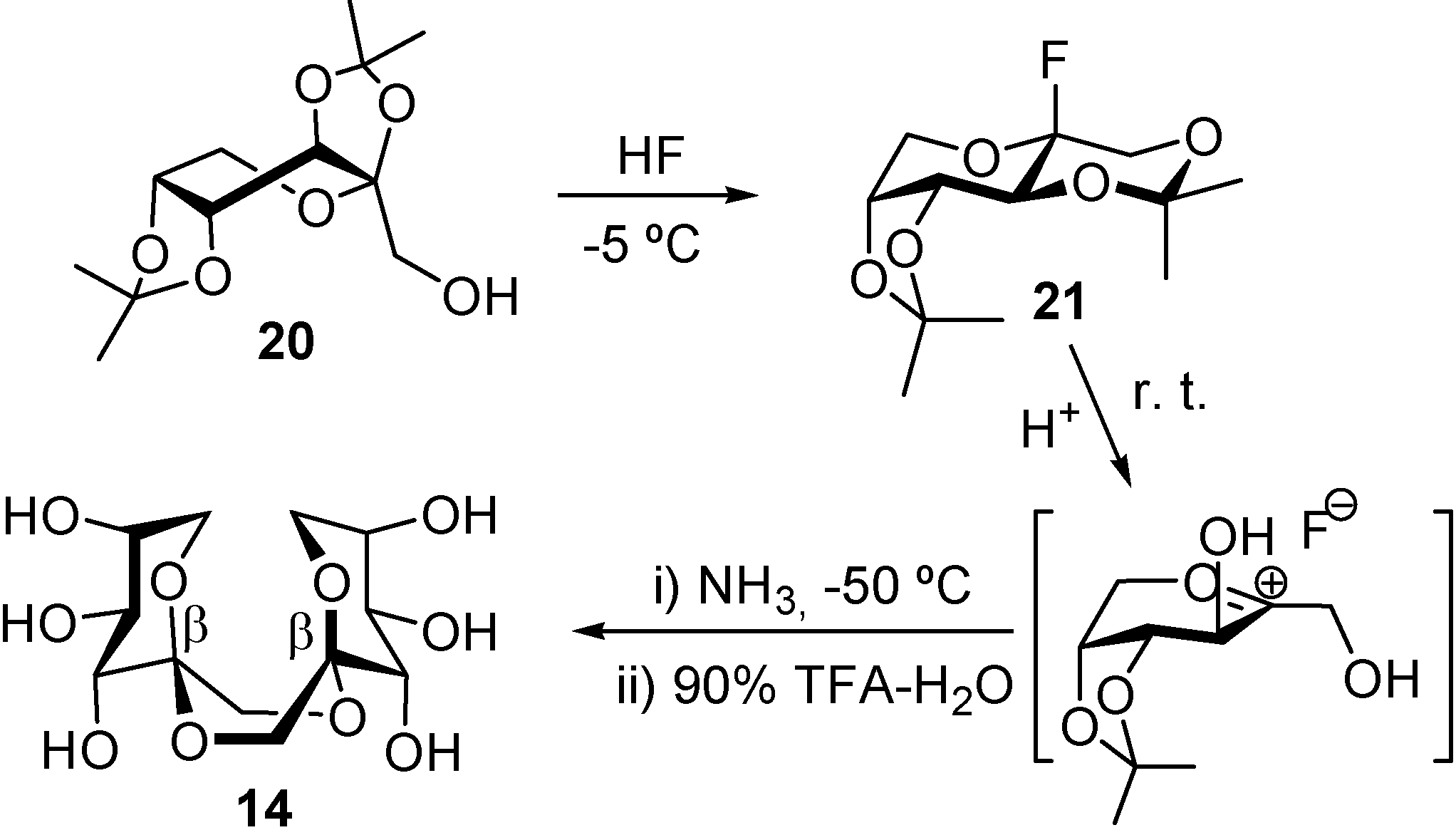
DFAs are formed upon thermal or protonic activation of fructose and fructose-containing disaccharides (sucrose) and oligosaccharides (levan and inulin), which includes caramelization of these raw materials. Under such conditions, a fructosyloxacarbenium cation is generated, which undergoes
in situ glycosylation by reacting with a second fructose unit. A transient ketodisaccharide is then formed, which undergoes intramolecular spiroketalization to furnish the DFA tricyclic core. A complex distribution of isomers that differ in the ring size, linking position, and stereochemistry at the ketal stereocenters is generally obtained. For instance, up to five different tricyclic cores and 13 DFA isomers have been identified in the disaccharidic fraction of fructose caramel (
Figure 1). An additional mixed dianhydride containing a glucose subunit [α-
d-fructofuranose-α-
d-glucopyranose-1,2’:2,1’- dianhydride, (
8)] has been identified in the disaccharidic fraction of sucrose caramel.
Figure 1.
DFA structures identified in the disaccharide fraction of fructose and sucrose caramel.
Figure 1.
DFA structures identified in the disaccharide fraction of fructose and sucrose caramel.
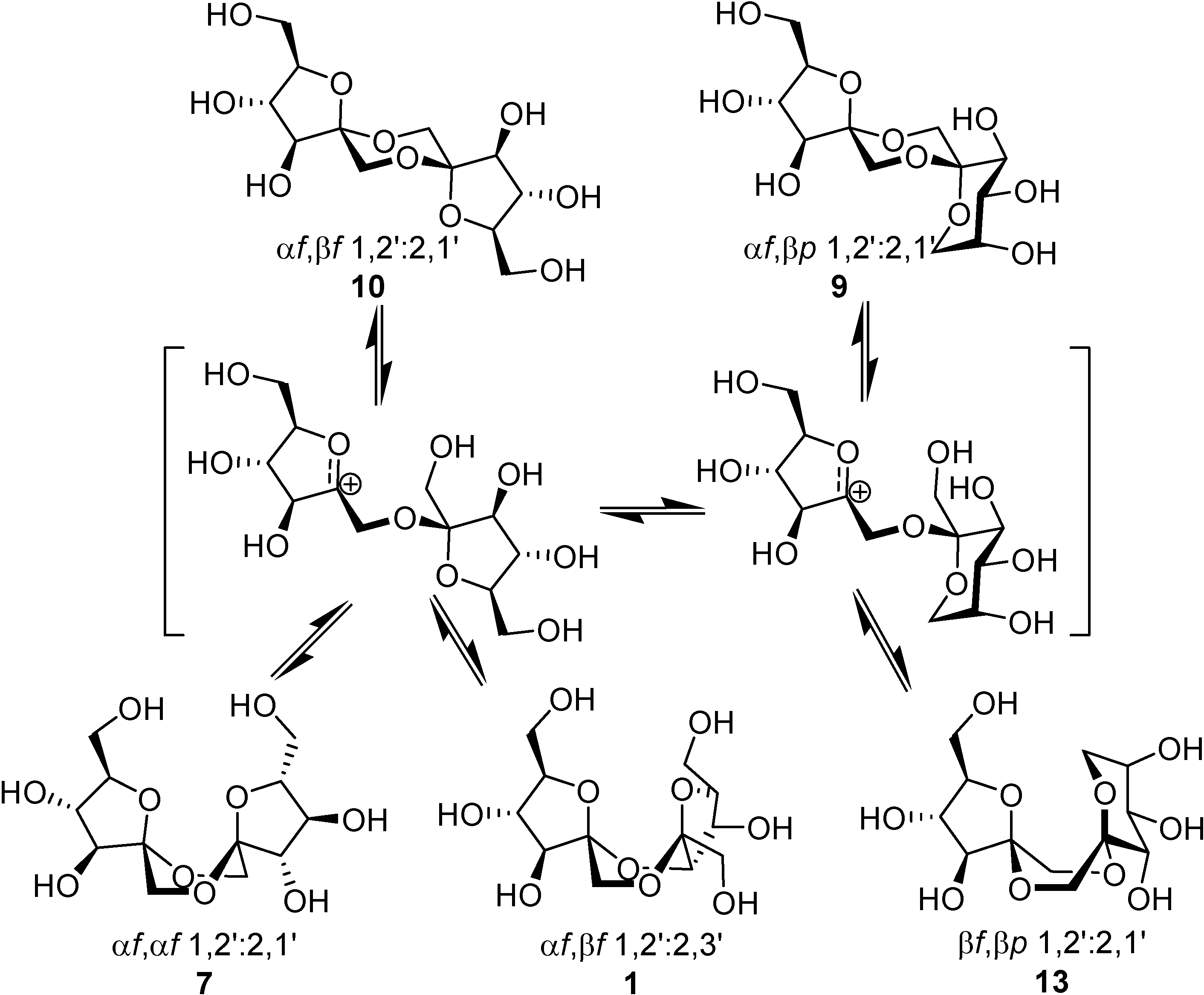
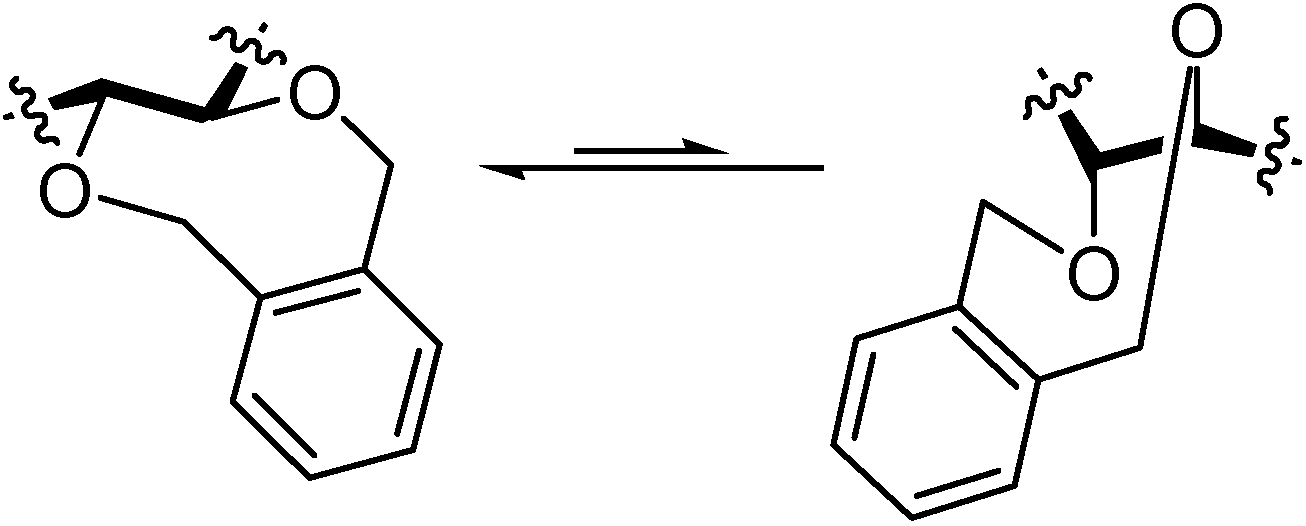
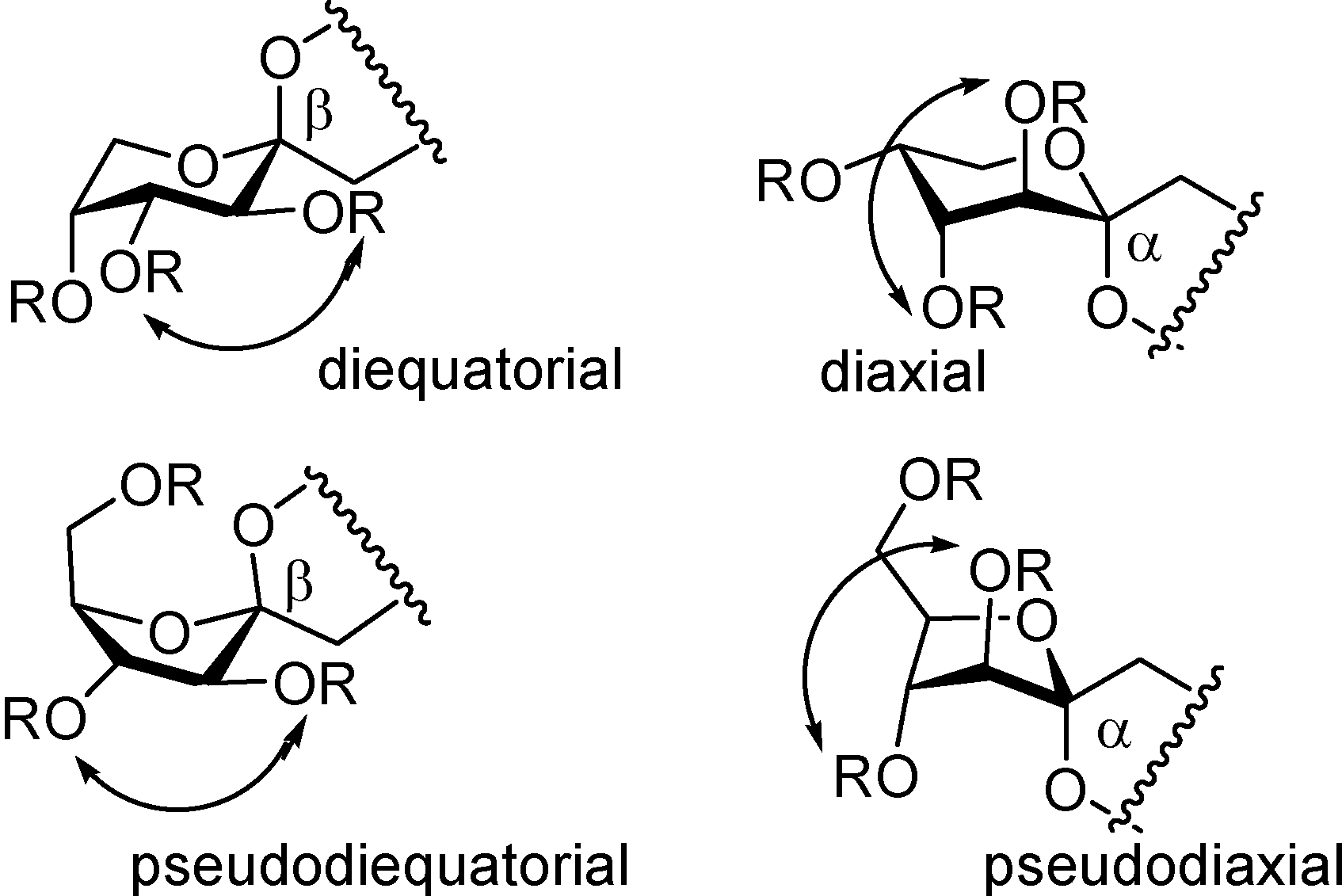
Spiroketalization is a reversible process under normal (thermal or protonic) caramelization conditions. Consequently, the isomeric distribution of DFAs in the mixture can be rationalized in terms of kinetic and thermodynamic control. Therefore, as observed by Manley-Harris and Richards for inulin pyrolysis [
21,
22], the relative abundance of isomeric DFAs is a balance between their rate of formation and disappearance. Dispiro-difuranose isomers [e.g., di-α-
d-fructofuranose-1,2’:2,1’- dianhydride (
7) or α-
d-fructofuranose-β-
d-fructofuranose-1,2’:2,1’-dianhydride (
10)] are more abundant when short reaction times are applied, while longer reaction times favor the thermodynamically more stable pyranose isomers [e.g., α-
d-fructofuranose-β-
d-fructopyranose- 1,2’:2,1’-dianhydride (
9)] and monospiro compounds [e.g., α-
d-fructofuranose-β-
d-fructofuranose- 1,2’:2,3’-dianhydride (
1)] (
Figure 2).
Figure 2.
DFA isomerization reactions.
Figure 2.
DFA isomerization reactions.
Since DFA formation is associated with heating of carbohydrate materials, DFAs are potentially present in any carbohydrate-rich foodstuff that has been subjected to cooking or that contains caramel or another cooked sugar as an additive. Actually, DFAs have been postulated as chemical markers and tracers of some food products. Defaye and co-workers have developed a GC-based analytical method that unequivocally assesses the authenticity and origin of caramels by observing the DFA fraction pattern in the chromatogram [
23]. The same procedure has been applied to detect honey adulteration with commercial syrups [
24] and to differentiate between natural- and sugar-roasted torrefacto coffee [
25].
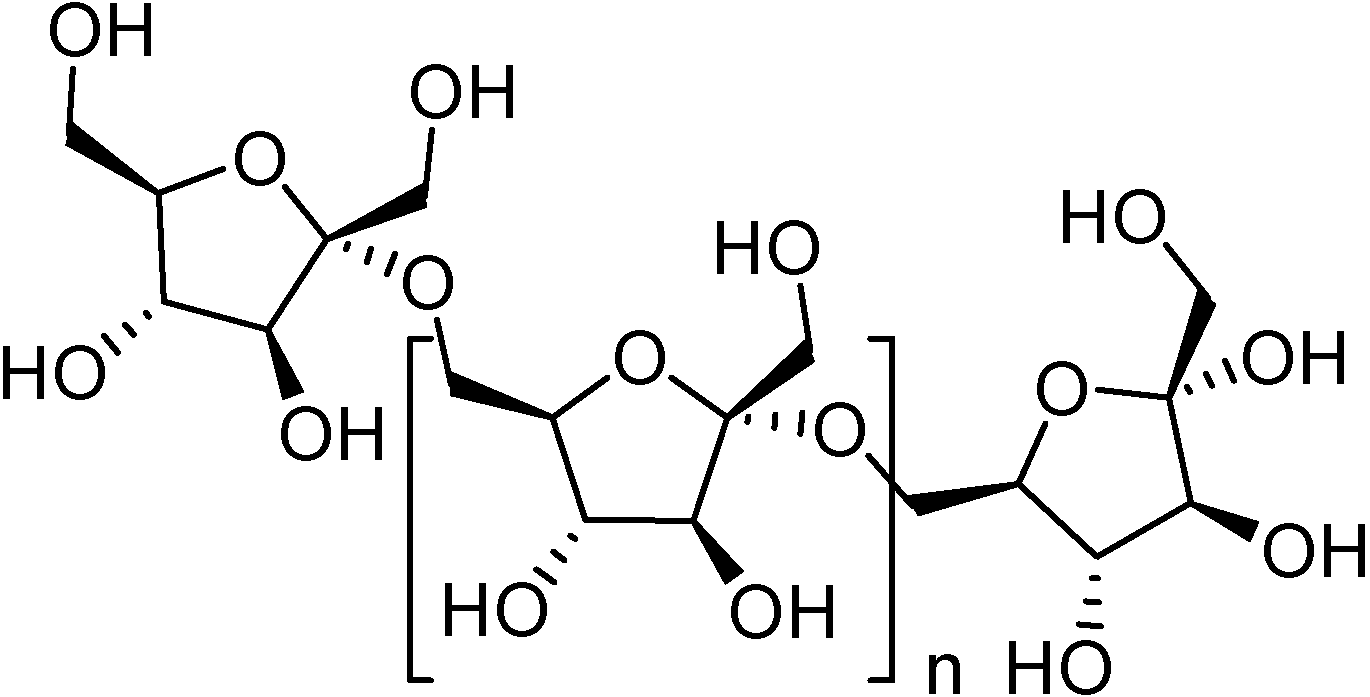
Nutritional studies on DFAs reveal some similarities between this family of cyclic fructodisaccharides and the reducing fructooligosaccharides (FOS) used as prebiotics in the food industry. FOS are mixtures of linear oligosaccharides containing five to ten β-(2→1)-linked fructose units (
Figure 3). They are non-digestible substances and behave as soluble alimentary fibers, facilitating intestinal motility. In mammals, including humans, FOS are known to exert a well studied bifidogenic effect in the colon [
26,
27] and are therefore extensively used as food additives [
28,
29].
Figure 3.
Structure of FOS.
Figure 3.
Structure of FOS.
In most of the patent files dealing with DFAs, they are rated as low-caloric sweeteners. Bifidogenic, anticariogenic and anti-tooth decaying effects have been also claimed. Moreover, DFAs promote
in vitro growth of bifidobacteria [
30]. Some of the benefits associated to a healthy
Bifidobacteria spp. population in the digestive tract of mammals are carcinogenesis inhibition [
31], the decrease of blood pressure and blood cholesterol levels [
32], vitamin B-complex synthesis stimulation [
33] or inhibition of proliferation of undesirable bacteria such as
Clostridium perfringens or
Escherichia coli, among others [
34,
35]. Some studies have revealed that the use of DFA-containing products in animal feeding (i.e., fowls) protect against digestive tract infections [
36]. These prebiotic effects are the basis of several inventions, like that described by Stoppok
et al. consisting of a beverage containing DFAs as dietary fiber [
37].
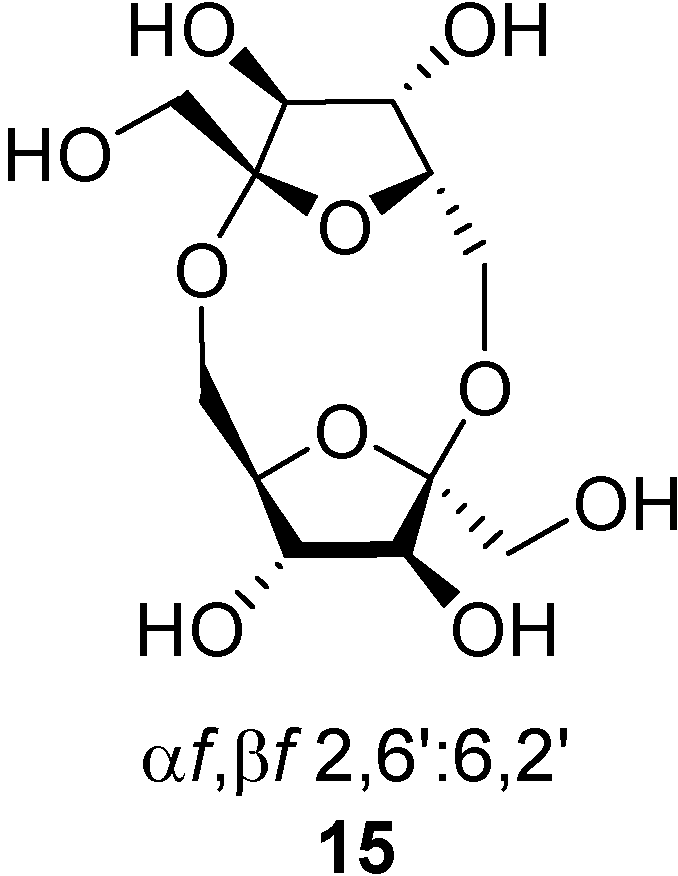
Several reports state that certain DFAs isomers are involved in intestinal metal cation absorption. For instance, Hara and Tomita have demonstrated that ingestion of α-
d-fructofuranose-β-
d-fructofuranose 1,2’:2,3’-dianhydride (
1, also known as DFA III) in daily diet increases the absorption of calcium, magnesium and zinc in rats [
38,
39,
40,
41,
42,
43]. The same authors observed that ingestion of DFA III and DFA IV [di-β-
d-fructofuranose-2,6’:6,2’-dianhydride (
15), a non-spiroacetalic DFA isomer,
Figure 4] [
44] affects the epithelial tissue and activates the passage of tight junctions in vitro, thereby promoting mineral absorption in the small and large intestine of rats [
45,
46]. α-
d-Fructofuranose β-
d-fructofuranose-1,2’:2,3’-dianhydride (
1) also prevents tannic acid-induced suppression of iron absorption and contributes to bone strength, thereby protecting against anaemia [
47] and preventing osteoporosis [
48]. Interestingly, these properties are characteristic of certain DFAs and have not been observed in other fructooligosaccharides.
Figure 4.
Structure of the non-spiroacetalic di-β-d-fructofuranose 2,6’:6,2’ dianhydride (15).
Figure 4.
Structure of the non-spiroacetalic di-β-d-fructofuranose 2,6’:6,2’ dianhydride (15).
Very recently, preliminary results concerning the effects of DFAs in the human diet have been reported. DFA III, one of the very few isomers available at technical scale, increases iron absorption and retention in female college students [
49] and a similar effect has been reported for calcium in healthy men [
50].
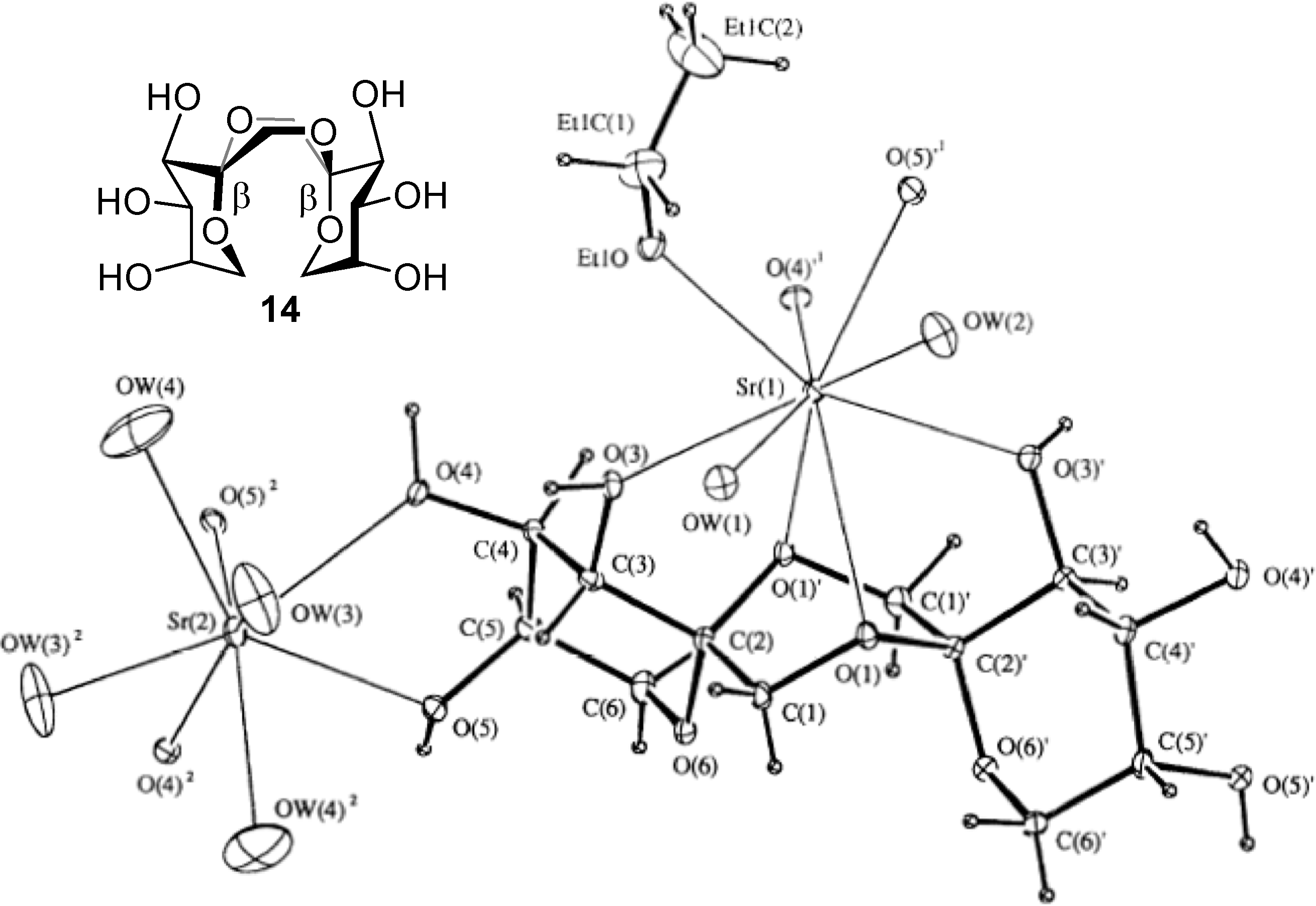
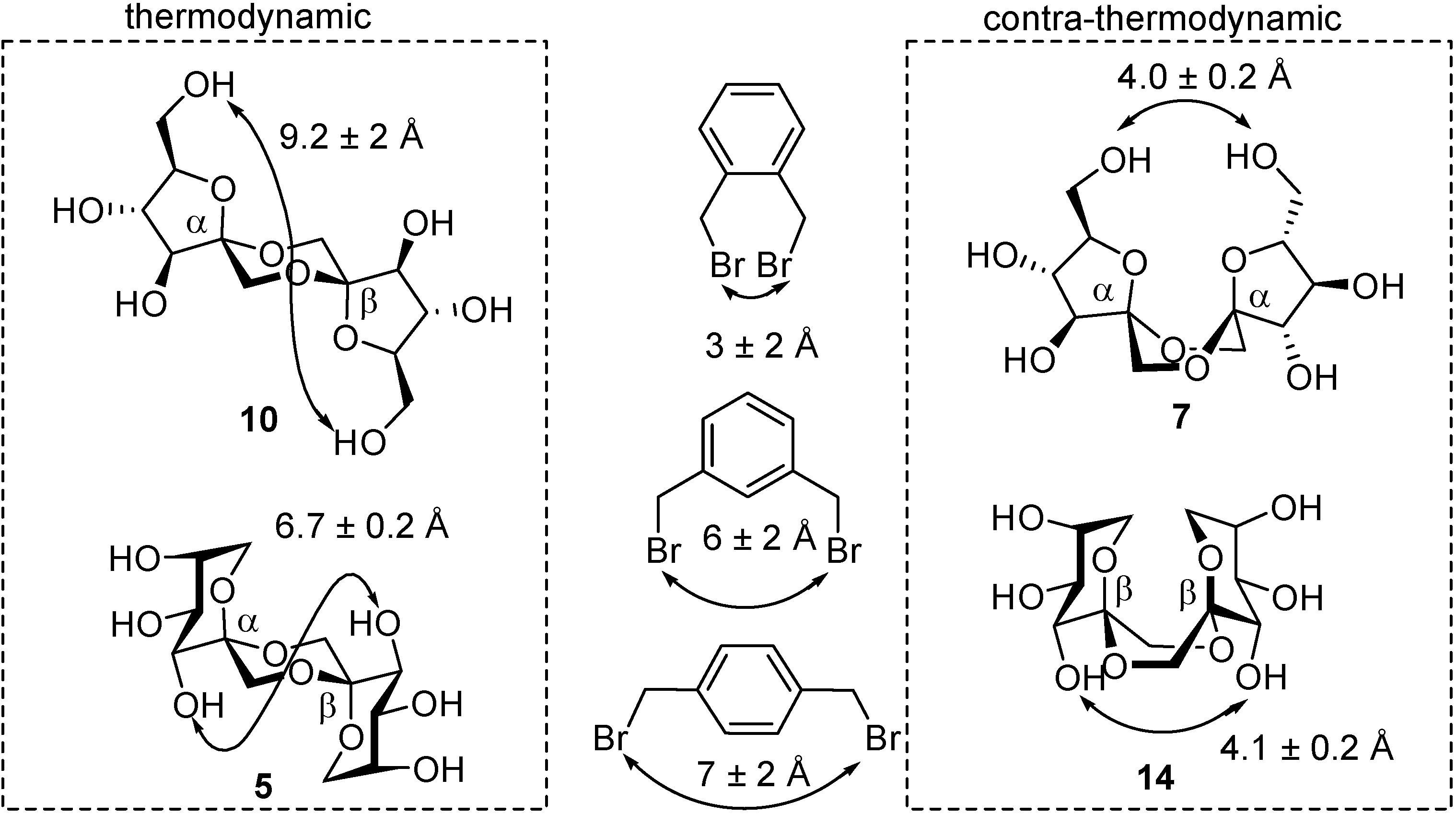
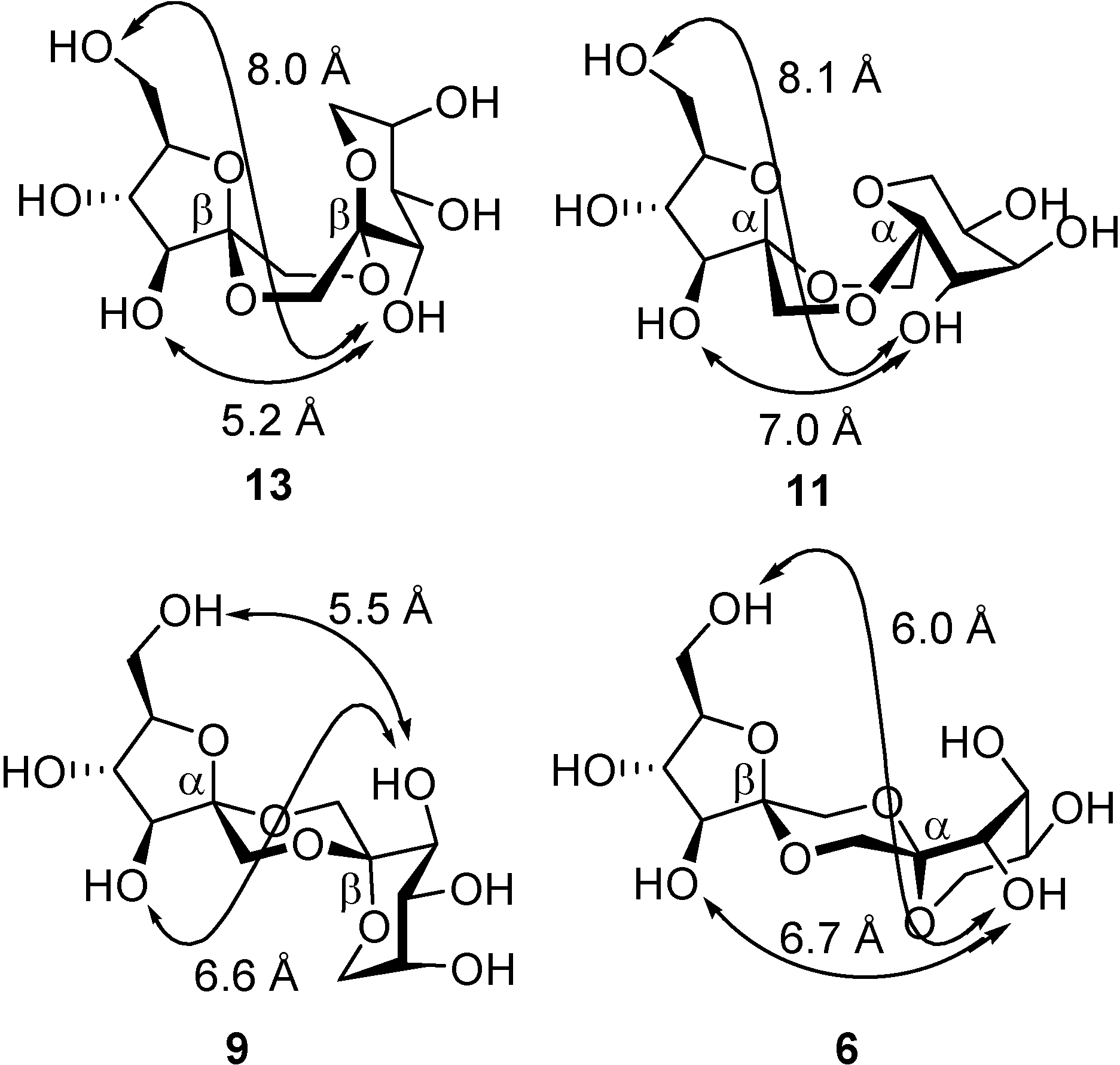
From a technical point of view, other than nutritional, the most interesting features of DFAs are probably the high rigidity of their structure and their hydrophilicity, which is shared with many sucrochemicals [
51,
52]. These characteristics can be exploited, for instance, in the preparation of surfactants, hydrophilic polymers, or complexing agents, to mention just a few. Indeed,
C2-symmetric
d-fructose-1,2’:2,1’-dianhydrides have been reported to form complexes with metal cations such as Ca
2+ and Sr
2+ [
53]. A conformational study carried out by Pedersen and coworkers revealed that a boat conformation in the central 1,4-dioxane ring is predominant in these isomers due to the interplay of the anomeric effect at the spiroketal centers and the exo-anomeric effect related to the fructose moieties [
54]. The boat (or skew-boat) conformer is the only one detected both in the solid state and in solution for di-β-
d-fructopyranose-1,2’:2,1’-dianhydride (
14), for instance [
53]. This arrangement allows both dioxane oxygen atoms to coordinate to a metal cation simultaneously and, at the same time, fixes a favorable orientation of the hydroxyl groups to participate in complex stabilization (
Figure 5). It has been postulated that such cation complexing DFAs might find an application as detergent builders or co-builders because of their ion-chelating and sequestering properties, representing an alternative to the banned tripolyphosphates [
55].
Figure 5.
X-Ray structure of the Sr
2+−di-β-fructopyranose-1,2’:2,1’-dianhydride (
14) complex (reproduced from reference [
53], with permission of NRC Research Press).
Figure 5.
X-Ray structure of the Sr
2+−di-β-fructopyranose-1,2’:2,1’-dianhydride (
14) complex (reproduced from reference [
53], with permission of NRC Research Press).
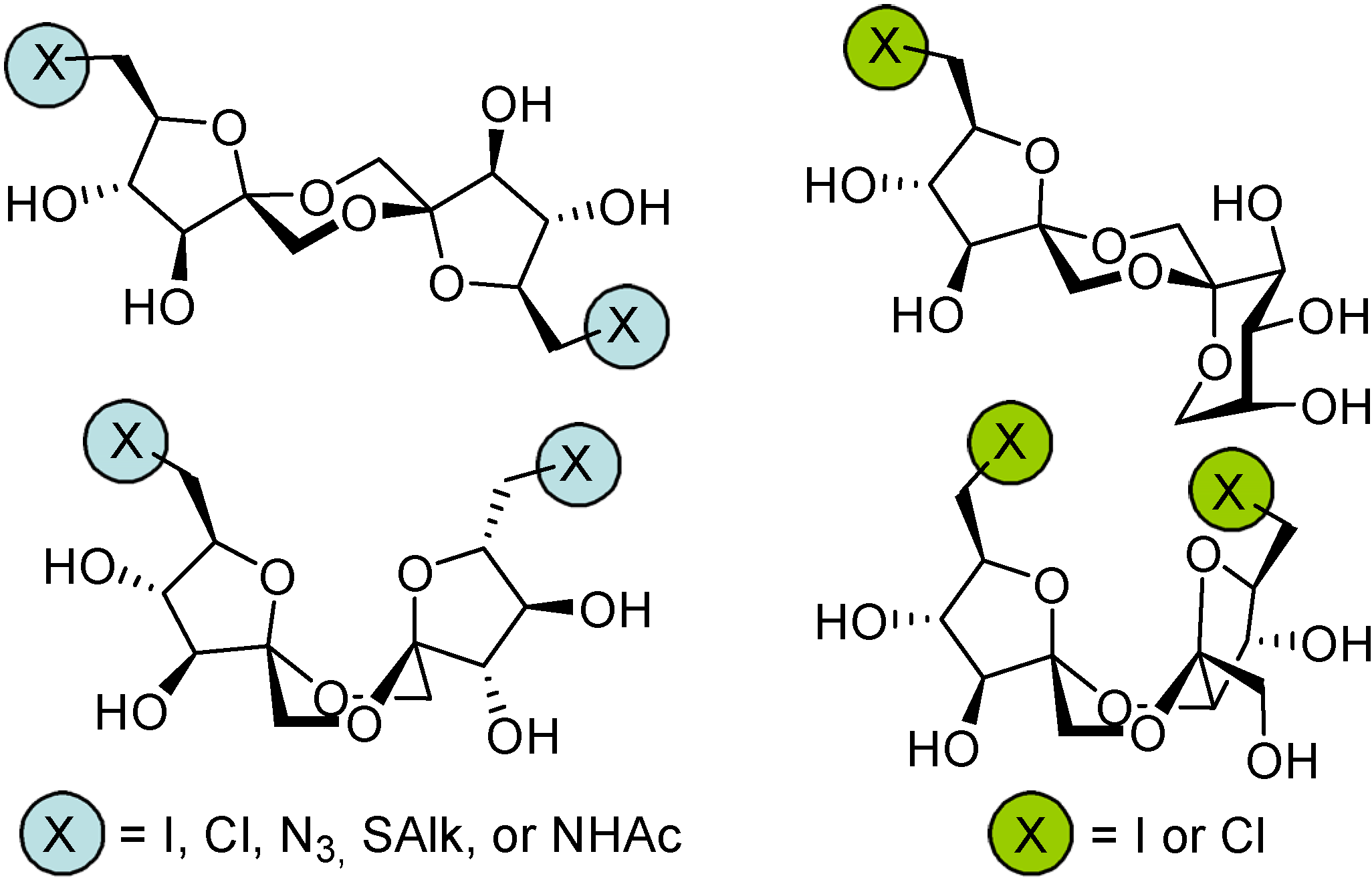
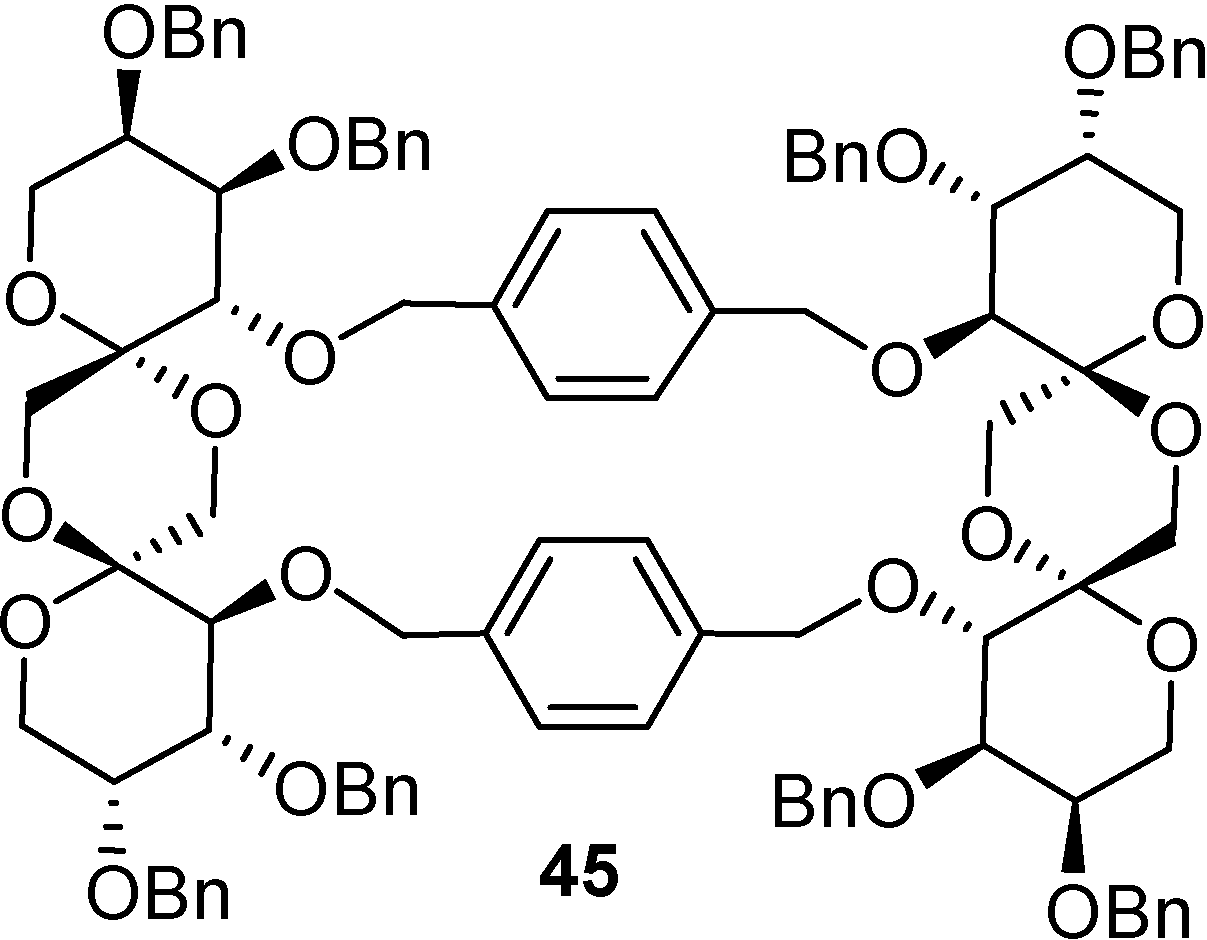
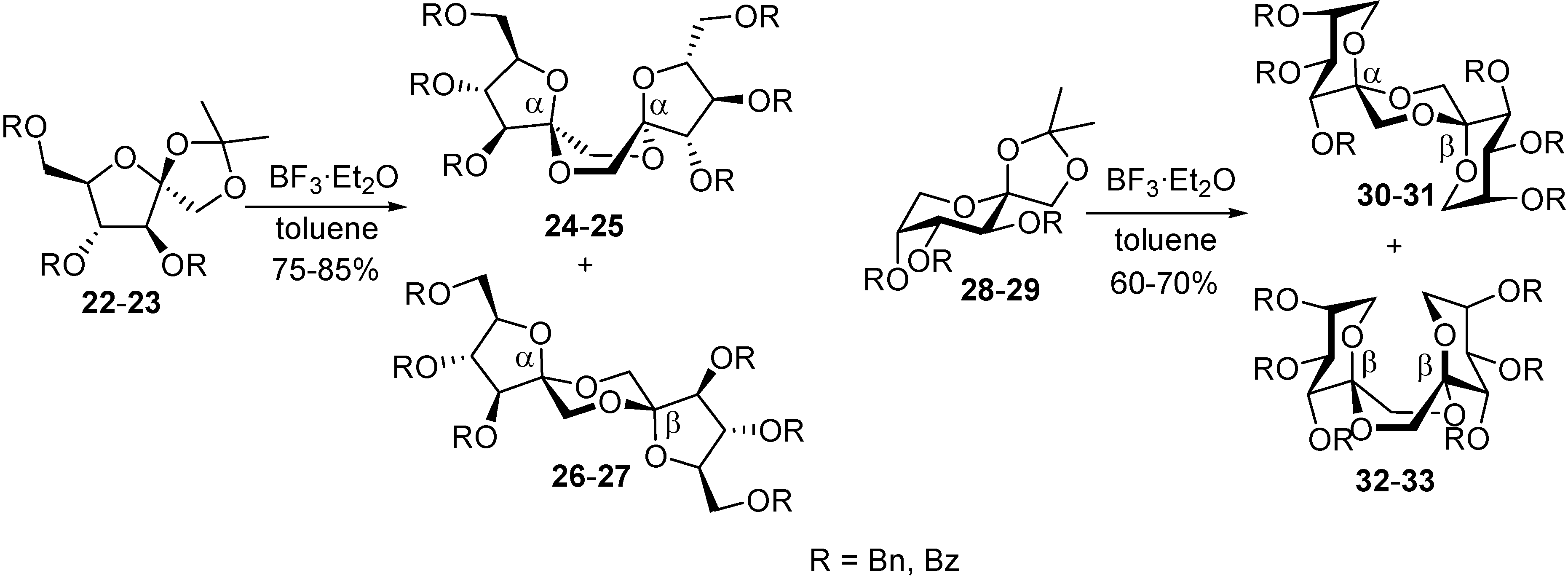
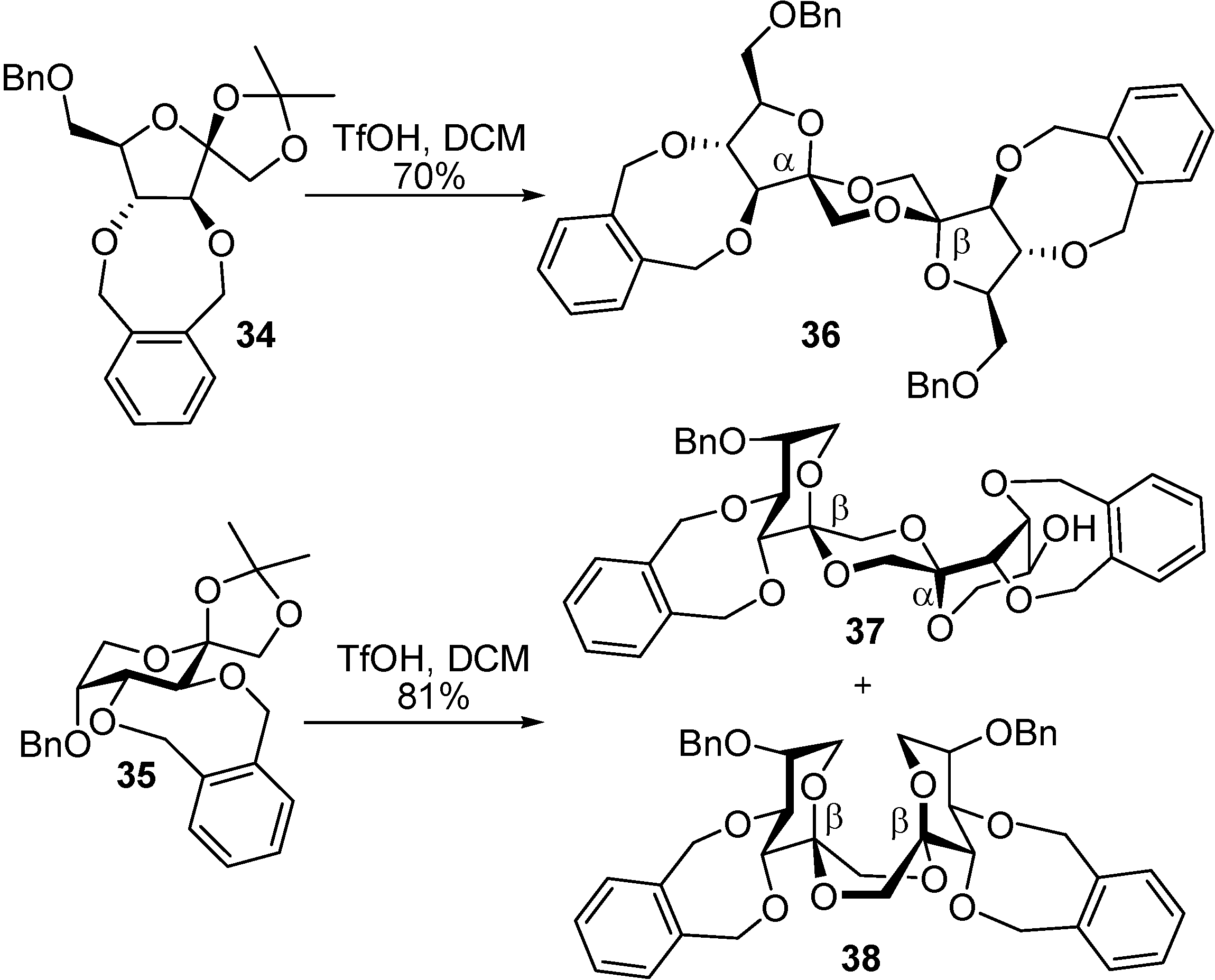
DFAs are also attractive synthetic intermediates. Their stability towards heat and acid hydrolysis makes them compatible with a wide range of reaction conditions. Applications as building blocks or scaffolds for chemical synthesis have been outlined. For instance, taking advantage of reported strategies for the selective functionalization of sucrose and fructooligosaccharides, García Fernández and Defaye have reported the preparation of libraries of DFA derivatives bearing diverse functional groups on their primary positions (i. e. halogens, azide, thioether, amine, or amide) (
Figure 6) [
56]. Some of the members of these libraries have shown remarkable properties as liquid crystals, while others were excellent precursors of hydrophilic polymers or non-cationic surfactants.
A much greater body of knowledge must be gained about the effects of ingesting the different isomeric DFAs. The lack of efficient production and purification methodologies to obtain pure DFA isomers has been a major limitation for such goal. During the last decade, major advances concerning the stereoselective synthesis of DFAs have been achieved that may significantly change the current status of this kind of spirocompounds in carbohydrate chemistry and biology. The increasing interest of the food industry in developing new “functional products” is motivating pure academic research in the field, which certainly will bring about new applications for this family of carbohydrate spiroketals.
Figure 6.
DFA derivative libraries [
56].
Figure 6.
DFA derivative libraries [
56].
Expectation for commercial viability of DFAs is further sustained by the fact that the raw materials from which they can be obtained are readily available from comparatively inexpensive agricultural feedstocks. DFA-producing oligosaccharides such as inulin or levan (consisting of β2→1 and β2→6-linked fructosides, respectively) are abundant starting materials. They are energy storage carbohydrates in higher plants, such as Jerusalem artichoke or chicory for the former and ryegrass or cocksfoot for the latter. Chicory, in particular, might eventually be considered to replace sugar beet in certain geographical areas where climate restrictions hampers the cultivation of the latter. Chicory tolerates cold, diseases and desiccation, while crop yields are much higher than those for corn or sugar beet (up to 45 tons per ha) [
57,
58]. Efficient large scale preparation of these oligosaccharides using sucrose industry technologies has been demonstrated. For instance, inulin has been produced in yields up to 46%, which compares very favorably with the 20-25% yield of sucrose obtained from sugar beet [
59].
4. Enzymatic Strategies towards the Synthesis of DFAs
The first evidence of specific DFA formation in higher plants dates back to 1933, when Schlubach and Knoop [
94] isolated a compound tentatively identified as α-fructofuranose β-fructofuranose 1,2’:2,1’-dianhydride (
10, also known as DFA I) from Jerusalem artichoke. The investigation of DFAs as natural products has been very limited until the mid-70s, when Tanaka and Uchiyama reported the isolation of an extracellular inulinase from
Arthrobacter ureafaciens, a soil bacteria that synthesized α-fructofuranose β-fructofuranose 1,2’:2,3’-dianhydride (DFA III) from inulin [
95,
96].
Table 5.
Nomenclature of DFA-producing enzymes.
Table 5.
Nomenclature of DFA-producing enzymes.
| EC 4.2.2.16 | |
|---|
| common name | levan fructotransferase (DFA IV-forming) |
| product | di-β-D-fructofuranose-2,6':2',6-dianhydride(non spiroketalic DFA) |
| EC 4.2.2.17 (formerly EC 2.4.1.200) | |
| common name | inulin fructotransferase (DFA I-forming) |
| product | α-D-fructofuranose β-D-fructofuranose-1,2':2,1'-dianhydride |
| EC 4.2.2.18 (formerly EC 2.4.1.93) | |
| common name | inulin fructotransferase (DFA III-forming) |
| product | α-D-fructofuranose β-D-fructofuranose-1,2':2,3'-dianhydride |
These seminal results have prompted much interest in the biosynthetic routes to DFAs, which however has been almost exclusively located in Japan. Unfortunately, a quantity of otherwise relevant papers was published in local journals, which limited their scope and accessibility. In a more general revision of the subject, Uchiyama has collected the most significant contributions to microbial biosynthesis and degradation of fructans up to 1993 [
97]. In 2000 Saito and Tomita [
98] reviewed particular issues concerning mass production of DFAs following biosynthetic routes and, very recently, Kawamura and Uchiyama have collated the recent advances in the enzymatic production of DFAs and cyclofructans [
99].
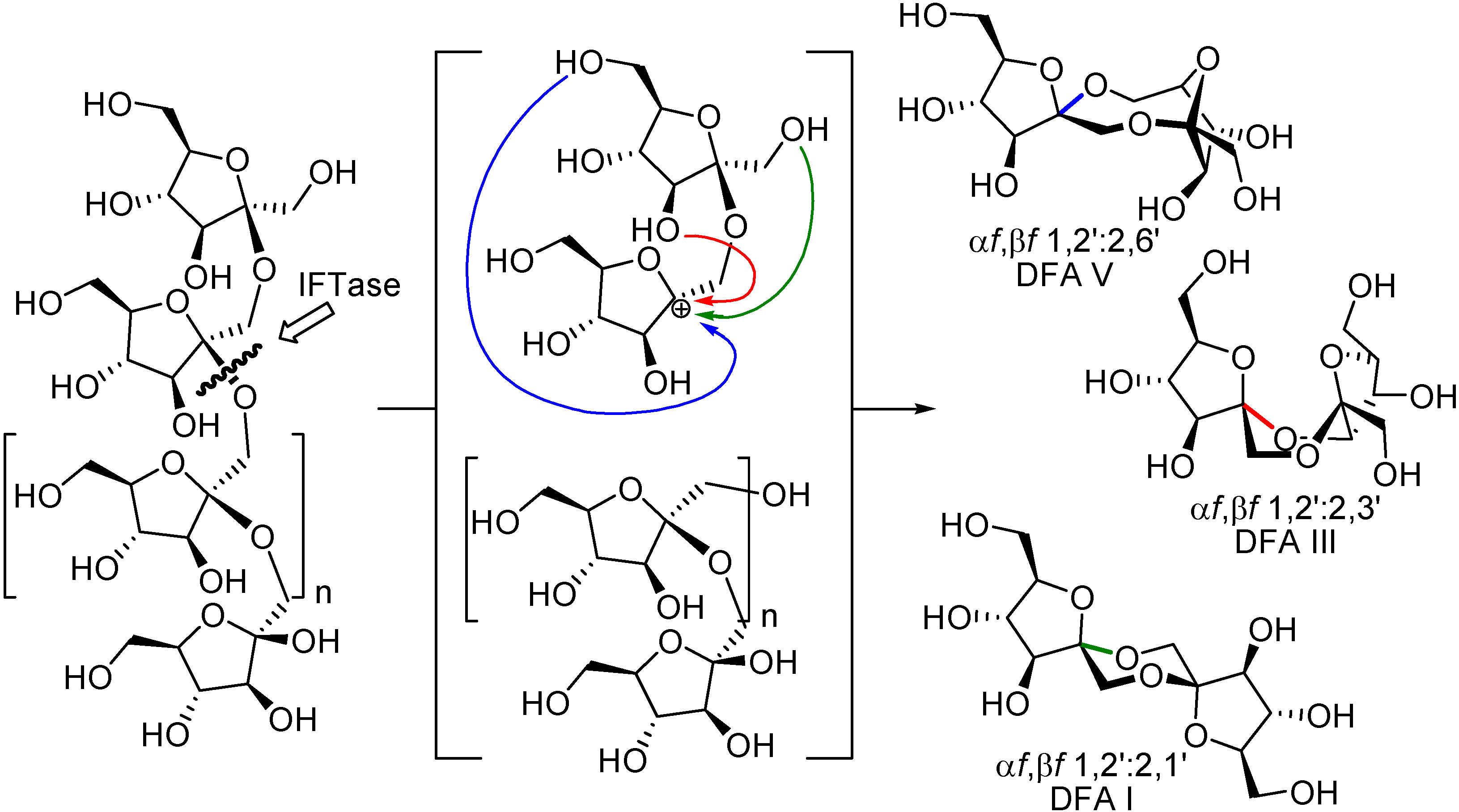
Since Tanaka and Uchiyama first report on the isolation of a DFA III-forming enzyme, several microorganisms, many of which belong to the
Arthrobacter genus, have been shown to produce enzymes that promote the transformation of fructans (inulin or levan) into DFAs (
Table 5). As a unique feature, and in stark contrast with common fructan decomposing enzymes, these enzymes catalyze an intramolecular transglycosylation reaction through which the second glycosidic bond from the non-reducing fructose is transferred to the terminal residue, thus releasing a DFA molecule (
Figure 14). This mechanism operates for both, inulin degrading (IFTase) [
100] and levan degrading (LFTase) enzymes [
101,
102,
103]. Though, IFTase have long been classified as “transferases”, they have been recently re-categorized as “lyases” (EC 4) since glycon transfer occurs intramolecularly through an elimination reaction [
104].
Figure 14.
IFTase-catalyzed synthesis of DFAs.
Figure 14.
IFTase-catalyzed synthesis of DFAs.
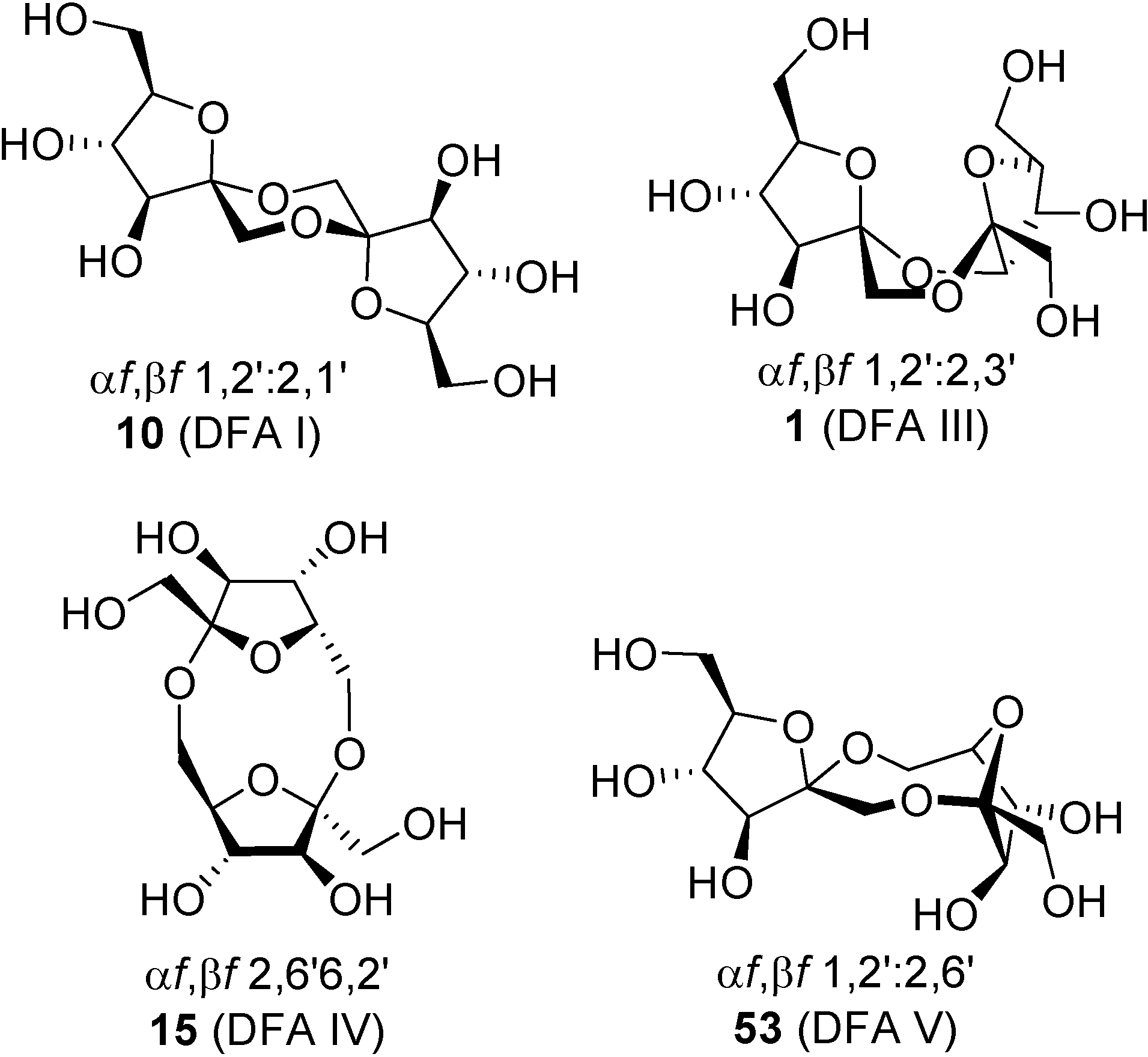
Up to now, four DFA isomers have been obtained by enzymatic degradation of fructans: α-
d-fructofuranose-β-
d-fructofuranose-1,2’:2,3’-dianhydride (
1, DFA III), α-
d-fructofuranose-β-
d-fructofuranose-1,2’:2,6’-dianhydride (
53, DFA V), α-
d-fructofuranose-β-
d-fructofuranose-1,2’:2,1’-dianhydride (
10, DFA I), and the non-spiroketal di-β-fructofuranose-2,6’:6,2’-dianhydride (
15, DFA IV) (
Figure 15). Though enzymatic synthesis is less versatile than chemical, exploitation of biocatalyzed processes for the synthesis of certain isomers may easily overcome synthetic drawbacks, especially concerning scaling up. Most of the reports on enzymatic synthesis of DFAs are related to DFA III and DFA IV. In fact their synthesis has become an issue of industrial interest [
98]. Reports on DFA I are more scarce and, so far, the DFA V-forming enzyme has not been characterized.
Isolation of DFA I-forming enzymes from
Arthrobacter spp. was first reported by Kobayashi and coworkers [
105]. The enzyme optimal operating conditions were pH 6 and 40 ºC and it was strongly inhibited by Hg
2+, Fe
3+, and other metal cations, a common feature for most of these enzymes [
106].
Streptomyces spp. have also been shown to be a source of DFA I-forming enzyme [
107]. More recently, Haraguchi and coworkers have reported new DFA I-forming enzymes from
Arthrobacter spp. with slightly prolonged heat stability [
108,
109], a feature that is mandatory when technical scale synthesis of this isomer is demanded. In the case of DFA V, reports are even more frugal. DFA V has been identified as a transient product during early stages of inulin thermolysis [
22] and its structure confirmed as its per-O-acetate [
110]. DFA V-producing enzyme has not been isolated yet and only evidences of its actions have been described [
111].
Figure 15.
DFA isomers available by biosynthetic routes.
Figure 15.
DFA isomers available by biosynthetic routes.
Enzymatic preparation is far more developed in the case of DFA III and the non-spiroketalic DFA IV [
34,
36]. The abundance of reports on isolation, characterization and even gene sequencing and engineering has provided a broader pool of DFAs III and IV-producing enzymes, which might be useful tools to face the ultimate challenge: industrial enzymatic biosynthesis of DFAs. Generally, enzymes dedicated to industrial production require high productivity and thermal stability. Yokota and coworkers have pioneered this field. These authors reported the isolation of an extracellular DFA III-producing IFTase from
Arthrobacter sp. H65-7 featuring high productivity (up to 90 U mL
-1) and thermal tolerance (retention of 80% activity after treatment at 70 ºC) [
112,
113]. Saito and Tomita have also reported the isolation and characterization of a LFTase from
Arthrobacternicotinovarorans GS-9 well suited for mass production of DFA IV [
114]. As a major advantage, enzyme excretion can be stimulated by their specific substrates and are easily purified. Furthermore, substrate conversion yields in a variety of conditions are excellent (
Table 6). Minor traces of fructose and short oligofructosides (1-kestose, nystose, and fructofuranosyl nystose) were removed by fermenting the reaction solution with baker yeast. As a drawback, the productive enzyme could not be recycled.
Table 6.
Enzymatic production of DFA III and IV.
Table 6.
Enzymatic production of DFA III and IV.
| Substrate | Product | Yield (%) |
|---|
| inulin, 50 g/L | DFA III | 93.0 |
| inulin, 250 g/L | DFA III | 82.5 |
| levan, 20 g/L | DFA IV | 75.5 |
| levan, 40 g/L | DFA IV | 73.3 |
Recently, enzymes featuring improved thermal stability have been reported. In the particular case of DFA III-forming enzymes, although temperature range has not been significantly raised, enzymes remain active after prolonged heating [
115,
116]. In all cases, substrate selectivity and product outcome profile still remain exquisite.
Similarly, improvements on LFTases performance in DFA IV formation have been achieved. However, only in few of these cases are the DFA yield and purity as high as in the case of the inulin converting enzymes [
117]. These enzymes are generally more easily inactivated by the presence of metal cations (Mn
2+, Cu
2+, Fe
3+ or Ag
+) [
118] and are very sensitive to substrate origin (molecular weight and branching) [
119,
120].
The search for efficient DFA-producing enzymes has lead to the development of a DFA IV production system directly from sucrose in a single culture by using a levan producing
B. subtilis as a host strain for the expression of LFTase gene [
121]. Furthermore, intensive efforts have led to the isolation of new enzymes, such as those reverting DFA formation [
122].
As evidence of the increasing interest in the development of DFA producing technologies, in the last decade, a number of genes encoding for DFA-forming enzymes (IFTases and LFTases) have been cloned [
123,
124]. Gene homology evaluation and site-directed manipulation might open new possibilities to engineer enzymes producing additional DFA isomers or featuring improved performance. A genetically engineered
E. coli has been reported to produce a DFA III-forming enzyme with twice as much activity compared to that of the original strain [
125]. A similar modification in LFTase-encoding gene resulted in a 5-fold increase on the DFA IV-forming enzyme activity [
126]. However, although these evident advances increased enzyme production, they did not improve enzyme turnover.
Lee and coworkers have tried to improve enzyme turnover by immobilizing a LFTase on different solid supports [
127]. Immobilized enzyme activity was comparable to that of the enzyme in solution and, in the best conditions, 60% of the activity was preserved after 20 catalytic cycles. Haraguchi and coworkers have also reported a heat-stable immobilized IFTase, which activity is virtually unaffected after repeated use (up to 8 cycles) [
128]. The authors suggest that a reactor using this immobilized enzyme might be envisioned for industrial production of DFA III. More recently, aiming at developing a method for commercial production of DFA III, Vorlop and coworkers have reported a genetically engineered enzyme tolerant to prolonged heating and, thus, well suited to function in continuous processes [
129]. Enzyme expression in
E. coli was extremely efficient (1760 U mL
-1). Moreover, the enzyme could be entrapped in calcium alginate hydrogels to enable the preparation of homogeneous enzyme-active beads (196 U mL
-1).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}













