3.1. Consumption of PGR Induced by Peroxyl Radicals in Presence of Triton X-100 Micelles
Figure 1.
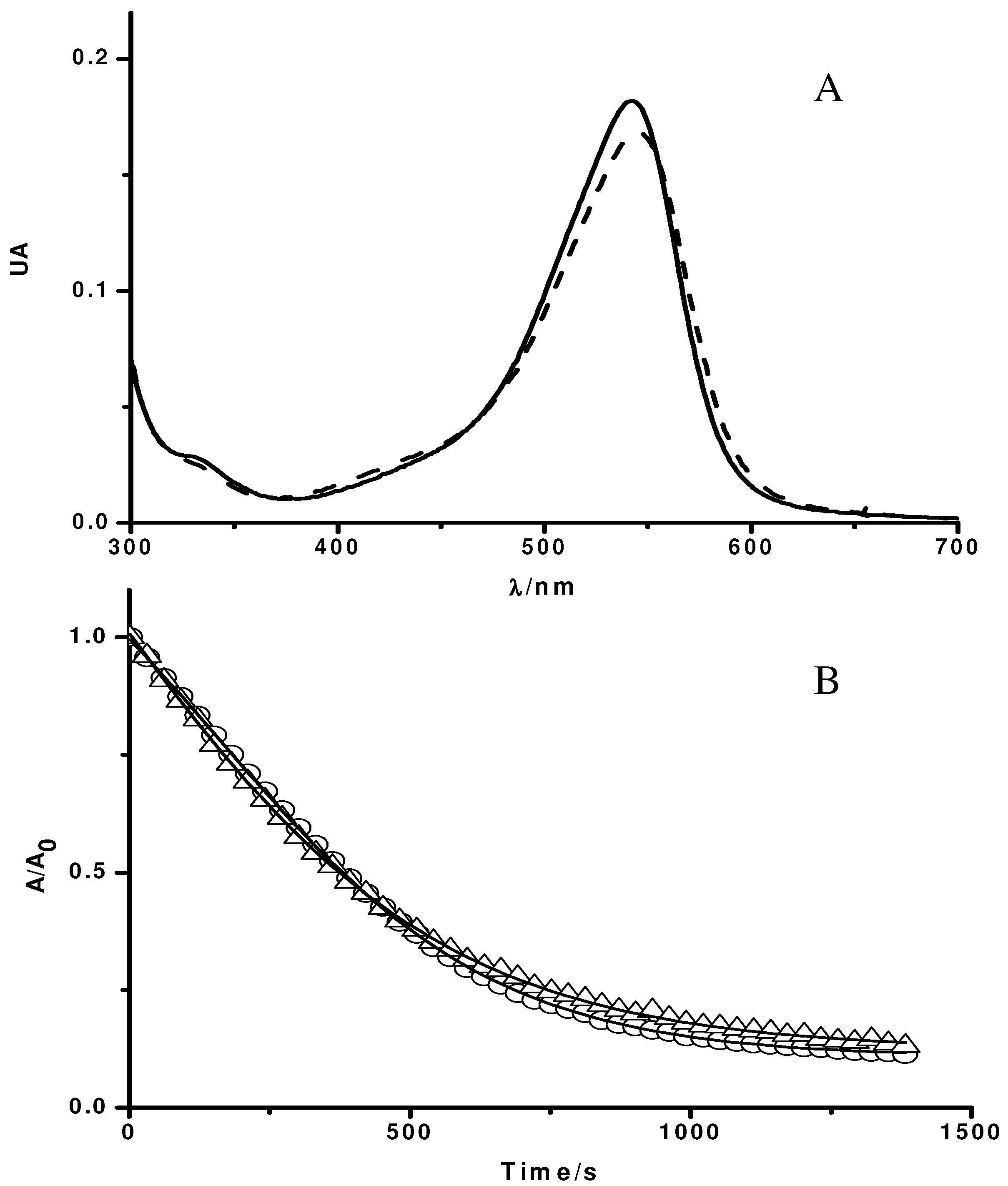
Influence of Triton X-100 micelles on the UV-visible spectrum of PGR (A) and its consumption mediated by peroxyl radicals (B). A: UV-visible spectrum of PGR (5 µM) in phosphate buffer (75 mM, pH 7.4, solid line), and in the presence of Triton X-100 (50 mM) (dashed line); B: Time-course of the consumption of PGR induced by AAPH derived peroxyl radicals in the absence and presence of Triton X-100 micelles. PGR (5 µM) was incubated in phosphate buffer (75 mM, pH 7.4) at 37 °C with AAPH (10 mM) in the absence (○), and presence of Triton X-100 (50 mM, △).
Figure 1.
Influence of Triton X-100 micelles on the UV-visible spectrum of PGR (A) and its consumption mediated by peroxyl radicals (B). A: UV-visible spectrum of PGR (5 µM) in phosphate buffer (75 mM, pH 7.4, solid line), and in the presence of Triton X-100 (50 mM) (dashed line); B: Time-course of the consumption of PGR induced by AAPH derived peroxyl radicals in the absence and presence of Triton X-100 micelles. PGR (5 µM) was incubated in phosphate buffer (75 mM, pH 7.4) at 37 °C with AAPH (10 mM) in the absence (○), and presence of Triton X-100 (50 mM, △).
Figure 1A shows UV-visible spectra of PGR in the absence and presence of Triton X-100 micelles (50 mM). As can be seen, in the absence of Triton X-100 micelles, PGR presented the typical UV-visible spectrum (solid line in
Figure 1), characterized by a band in the visible region with a maximum at 540 nm (ε = 36,000 M
-1cm
-1). In the presence of Triton X-100 micelles, the spectrum of PGR was slightly altered (dashed line in
Figure 1), displacing the maximum of the visible band to 544 nm (ε = 33600 M
-1cm
-1). This behavior would imply that PGR was, at least partially, associated to the micellar pseudophase. In fact, by an ultrafiltration separation process it was observed that, in our experimental conditions (Triton X-100 50 mM, and PGR 5 µM), near 72% of PGR would be associated to Triton X-100 micelles. Nevertheless, the association of PGR to Triton X-100 micelles did not modify the kinetic profile of PGR consumption induced by peroxyl radicals. As is shown in
Figure 1B, the consumption of PGR mediated by AAPH derived peroxyl radicals (evaluated by its absorbance decay at 540 nm) was not affected by the presence of Triton X-100 micelles. This is an unexpected result if it is considered that AAPH and its derived free radicals are mostly in the aqueous phase, while a significant fraction of PGR is associated to the micelles. However, the PGR/peroxyl radical process (Reaction 2,
Scheme 1) is in the zero order kinetic limit [
10,
11] and hence its consumption rate is independent of the PGR concentration present in the aqueous phase. In fact, similar initial rate values (≈ 0.4 µM/min) of PGR decay were estimated in the absence and presence of Triton X-100 micelles. Considering that peroxyl radicals are generated (APPH = 10 mM, at 37 °C) at a rate of 0.8 µM/min [
6], and that each PGR molecule reacts with two peroxyl radicals [
11], the estimated rates of PGR consumption in the absence and presence of Triton X-100 micelles supports the proposal that PGR is in the zero order kinetic limit. Furthermore, PGR adsorbed at the micelles interface could retain its reactivity toward the peroxyl radicals.
3.2. Effect of Single Antioxidants on the PGR Consumption in Presence of Triton X-100 Micelles
To evaluate the effect of Triton X-100 micelles on the protection of PGR afforded by pure compounds, different antioxidants with different hydro/lipophilicity were employed.
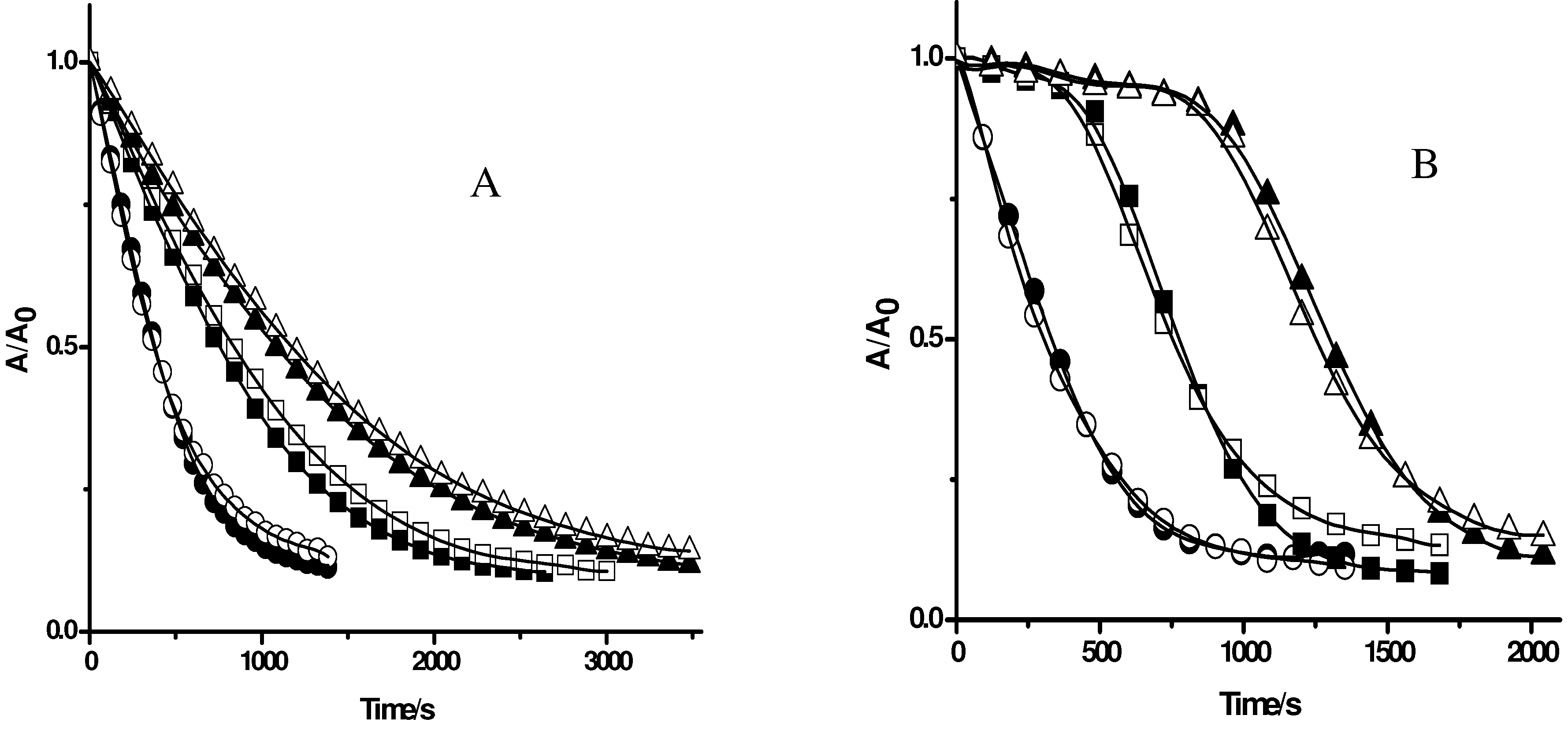
Figure 2 shows the consumption of PGR (5 µM) induced by AAPH derived peroxyl radicals in the presence of Trolox, and ascorbic acid with or without Triton X-100 micelles. As is shown in
Figure 2A, Trolox (50 and 100 µM) inhibited the consumption of PGR in a concentration-dependent way, showing a similar behaviour to that observed in previous studies in aqueous media [
10,
11]. Interestingly, the addition of Triton X-100 micelles did not modify the protection of PGR afforded by Trolox, a result that would be related to the well known hydrophilic property of this antioxidant. In the same way, the addition of Triton X-100 micelles did not alter the protective effect afforded by ascorbic acid, characterized by a neat induction time (
Figure 2B). These similarities can be attributed to the fact that, even in presence of Triton X-100 micelles, all relevant processes take place almost exclusively in the aqueous phase. In contrast with this, when a lipophilic antioxidant as β-carotene was added to a solution containing Triton X-100 micelles, PGR and AAPH, no PGR protection by the antioxidant took place (
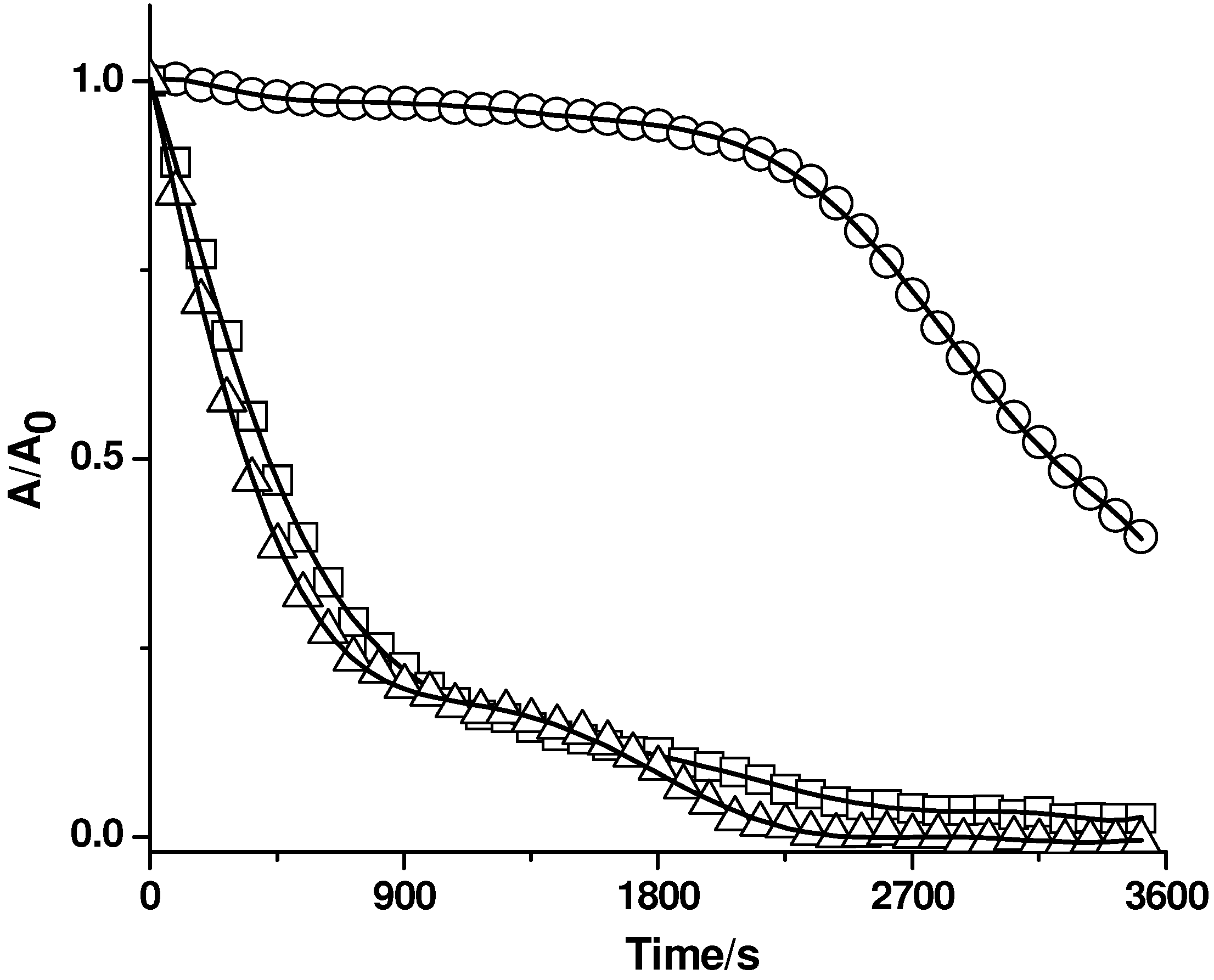
Figure 3). In fact, as it is shown in
Figure 3, the absorption band corresponding to β-carotene (at 460 nm) was not affected by peroxyl radicals during the consumption of PGR. Nonetheless, when PGR was almost totally consumed (c.a. 37 minutes), the visible band (at 460 nm) associated to β-carotene, decayed significantly. These results imply that β-carotene (present in the micelles) was initially protected by PGR (present in the aqueous phase and/or in the micelle-aqueous interface), and only after the total consumption of PGR, β-carotene was consumed by peroxyl radicals (probably in the interface micelle-phosphate buffer). Similarly, β-carotene was efficiently protected by Trolox and ascorbic acid (data not shown), implying that hydrophilic antioxidants react with peroxyl radicals in the aqueous phase, minimizing its interaction with micelle associated targets (as β-carotene). Control experiments with β-carotene micellar solutions revealed no changes in the original absorbance intensity at 460 nm (data not shown) in the time period considered in the presence work.
Figure 2.
Effect of antioxidants on the consumption of PGR induced by AAPH derived peroxyl radicals in the absence (closed symbols) and presence of Triton X-100 micelles (open symbols). PGR (5 µM) was incubated with AAPH (10 mM) in the absence (circles) and presence of. A: Trolox: 50 (squares), and 100 µM (triangles); B: Ascorbic acid: 5 (squares), and 10 µM (triangles).
Figure 2.
Effect of antioxidants on the consumption of PGR induced by AAPH derived peroxyl radicals in the absence (closed symbols) and presence of Triton X-100 micelles (open symbols). PGR (5 µM) was incubated with AAPH (10 mM) in the absence (circles) and presence of. A: Trolox: 50 (squares), and 100 µM (triangles); B: Ascorbic acid: 5 (squares), and 10 µM (triangles).
Figure 3.
Effect of β-carotene on the consumption of PGR induced by AAPH derived peroxyl radicals. PGR (5 µM) was incubated with AAPH (10 mM) and Triton X-100 (50 mM) in the absence and presence of β-carotene (5 µM). The reaction was followed by the consumption of PGR (□, 550 nm) and β-carotene (○, 460 nm). Control experiment without β-carotene (△).
Figure 3.
Effect of β-carotene on the consumption of PGR induced by AAPH derived peroxyl radicals. PGR (5 µM) was incubated with AAPH (10 mM) and Triton X-100 (50 mM) in the absence and presence of β-carotene (5 µM). The reaction was followed by the consumption of PGR (□, 550 nm) and β-carotene (○, 460 nm). Control experiment without β-carotene (△).
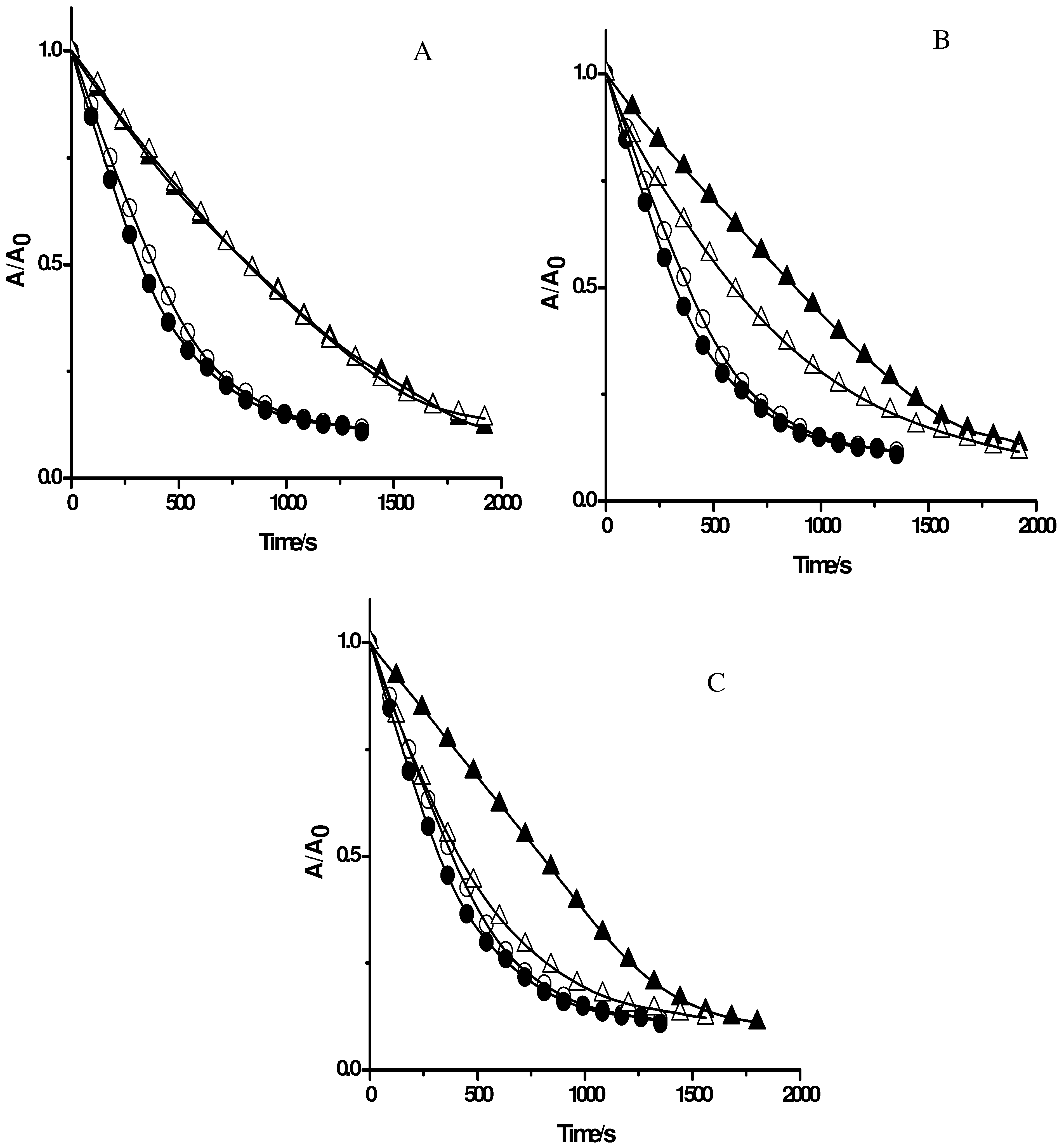
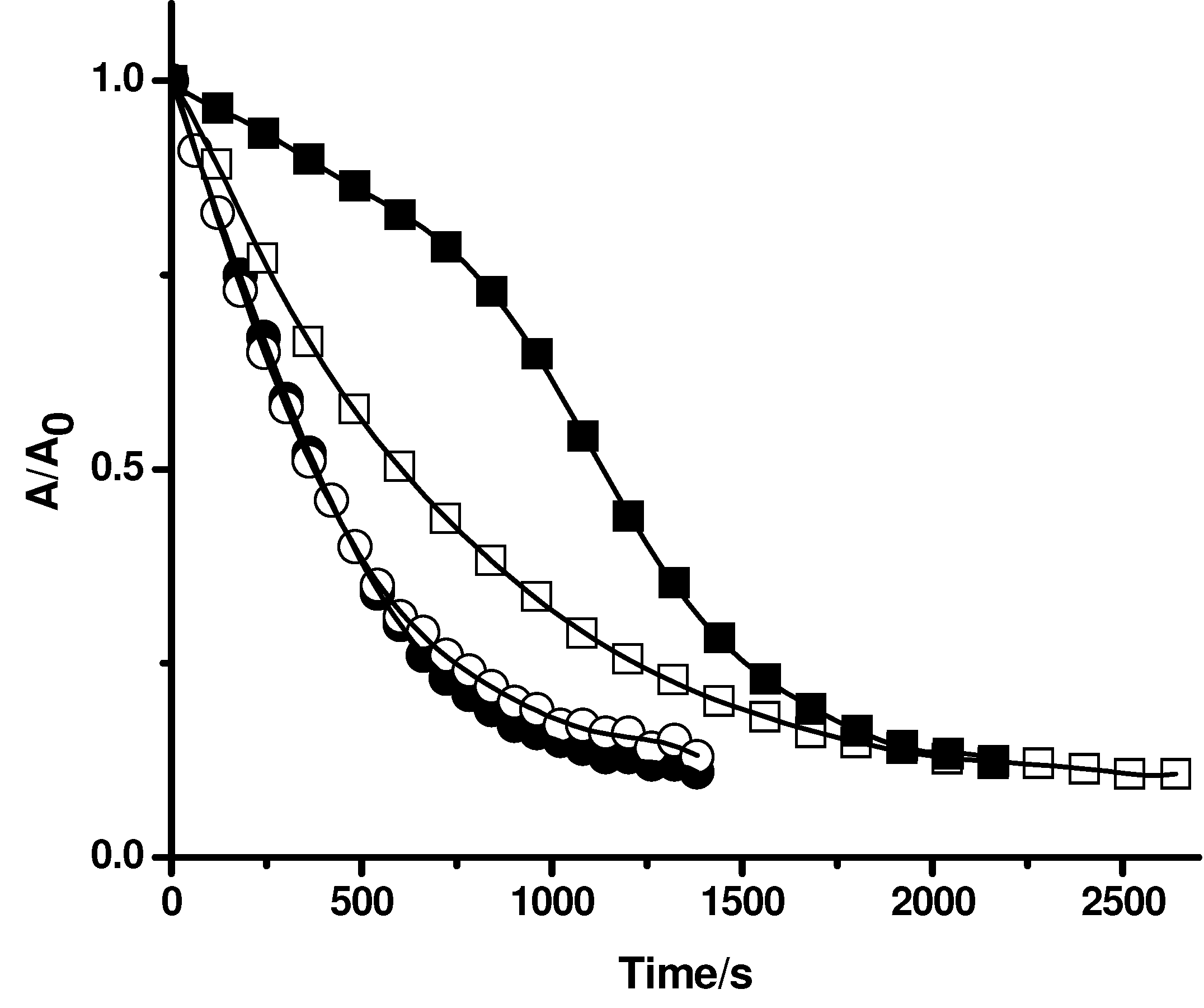
Figure 4.
Effect of gallate derivatives (A gallic acid; B propyl gallate; C octyl gallate) on the consumption of PGR induced by AAPH derived peroxyl radicals in the absence and presence of Triton X-100 micelles. PGR (5 µM) was incubated with AAPH (10 mM) and the gallate derivatives (at 5 µM) in micellar (△) and phosphate solutions (▲). Control experiments without gallate derivatives were carried out in the presence (○) and absence (●) of Triton X-100 micelles.
Figure 4.
Effect of gallate derivatives (A gallic acid; B propyl gallate; C octyl gallate) on the consumption of PGR induced by AAPH derived peroxyl radicals in the absence and presence of Triton X-100 micelles. PGR (5 µM) was incubated with AAPH (10 mM) and the gallate derivatives (at 5 µM) in micellar (△) and phosphate solutions (▲). Control experiments without gallate derivatives were carried out in the presence (○) and absence (●) of Triton X-100 micelles.
To evaluate the influence of Triton X-100 micelles on the protection of PGR afforded by others antioxidants, compounds with a similar reactive centre, but with different degree of lipophilicity, were used. For this purpose, gallic acid and gallate derivatives with different alkyl chain lengths, such as methyl gallate, propyl gallate, and octyl gallate, were employed as model antioxidants. As can be seen in
Figure 4A, the consumption of PGR in the presence of gallic acid at 5 µM concentration was very similar in the absence and presence of Triton X-100 micelles. This result is compatible with those obtained employing Trolox and ascorbic acid (
Figure 2). However, in the case of propyl, and octyl gallate (
Figure 4B, and 4C, respectively) a clear effect of Triton X-100 micelles was observed. In the presence of Triton X-100 micelles the protection afforded by propyl gallate was lower than that the observed in buffer. This would imply that the incorporation of propyl gallate into the micellar pseudophase leads to a lower availability of propyl gallate to react with peroxyl radicals. An extreme case was observed for octyl gallate. The presence of Triton X-100 micelles significantly decreased the observed protection of PGR afforded by this antioxidant in homogeneous medium, indicating that octyl gallate was mostly incorporated in Triton X-100 micelles (
Figure 4C). This corroborates the results obtained with β-carotene (
Figure 3) indicating that the free radical scavengers incorporated into the micelles do not protect PGR from its consumption mediated by peroxyl radicals. Likewise, we assessed the effect of caffeic acid and ethyl caffeate on the consumption of PGR induced by AAPH derived peroxyl radicals. As shown in
Figure 5A, the protective effect of caffeic acid on the consumption of PGR was independent of the presence of Triton X-100 micelles. However, the protection of PGR afforded by ethyl caffeate decreased significantly in the micellar solution (
Figure 5B). A similar behaviour was observed for quercetin (
Figure 6). Quercetin, in phosphate buffer protected efficiently PGR, while in presence of Triton X-100 micelles, its protective effect decreased considerably.
Figure 5.
Effect of caffeic acid (A) and ethyl caffeate (B) on the consumption of PGR induced by AAPH derived peroxyl radicals in the absence and presence of Triton X-100 micelles. PGR (5 µM) was incubated with AAPH (10 mM) and the caffeic acid derivatives (at 250 µM) in micellar (△) and phosphate solutions (▲). Control experiments without caffeic acid derivatives were carried out in the presence (○) and absence (●) of Triton X-100 micelles.
Figure 5.
Effect of caffeic acid (A) and ethyl caffeate (B) on the consumption of PGR induced by AAPH derived peroxyl radicals in the absence and presence of Triton X-100 micelles. PGR (5 µM) was incubated with AAPH (10 mM) and the caffeic acid derivatives (at 250 µM) in micellar (△) and phosphate solutions (▲). Control experiments without caffeic acid derivatives were carried out in the presence (○) and absence (●) of Triton X-100 micelles.
Figure 6.
Effect of quercetin (7 µM) on the consumption of PGR (5 µM) induced by AAPH derived peroxyl radicals in the absence (■) and presence of Triton X-100 micelles (□). Control experiments without quercetin were carried out in the presence (○) and absence (●) of Triton X-100 micelles.
Figure 6.
Effect of quercetin (7 µM) on the consumption of PGR (5 µM) induced by AAPH derived peroxyl radicals in the absence (■) and presence of Triton X-100 micelles (□). Control experiments without quercetin were carried out in the presence (○) and absence (●) of Triton X-100 micelles.
The above results imply that the protection of PGR is strongly influenced by the distribution of the antioxidants in Triton X-100 micelles. In particular, protection by water soluble substrates is similar to that measured in the absence of Triton X-100 micelles. On the other hand, highly hydrophobic substrates do not protect PGR in the presence of Triton X-100 micelles. Antioxidants that partition between both pseudophases (such as quercetin) present an intermediate behaviour. Thus, in the presence of complex mixtures containing antioxidants, the observed effect of Triton X-100 micelles on the kinetic profiles of PGR consumption could be directly related to the percentage of lipophilic antioxidants included in the sample. To study the latter, we estimated the inhibition of PGR consumption induced by AAPH derived peroxyl radicals by complex mixtures such as wines, juices, berry extracts, and tea infusions in the absence and presence of Triton X-100 micelles.
Figure 7 shows the protection of PGR afforded by orange juice (
Figure 7A) and a seed extract of
Eugenia jambolana (
Figure 7B). As can be seen in this figure, orange juice protected PGR throughout a clear induction time. This induction time, originated by the presence of ascorbic acid, was not altered by the presence of Triton X-100 micelles. This result is in accord to those obtained regarding the antioxidant capacity of pure ascorbic acid and implies that the protection of orange juice is almost exclusively related to this antioxidant. Seed extracts of
Eugenia jambolana inhibited efficiently the consumption of PGR induced by peroxyl radicals (
Figure 7B). However, in contrast to orange juice, the effect of the seed extract was dependent of the presence of Triton X-100 micelles. Thus, in the presence of micelles, a lower effect was registered, implying that lipophilic antioxidants contribute to the antioxidant capacity of this extract.
Figure 7.
Effect of complex mixtures on the consumption of PGR (5 µM) induced by AAPH (10 mM) derived peroxyl radicals. A: orange juice (6.7 µL/mL) in the absence (▲) and presence of Triton X-100 micelles (△). B: seed extract of Eugenia jambolana (10 µL/mL) in the absence (▲) and presence of Triton X-100 micelles (△). Control experiments (without additives) were measured in the absence (●) and presence (○) of Triton X-100 micelles.
Figure 7.
Effect of complex mixtures on the consumption of PGR (5 µM) induced by AAPH (10 mM) derived peroxyl radicals. A: orange juice (6.7 µL/mL) in the absence (▲) and presence of Triton X-100 micelles (△). B: seed extract of Eugenia jambolana (10 µL/mL) in the absence (▲) and presence of Triton X-100 micelles (△). Control experiments (without additives) were measured in the absence (●) and presence (○) of Triton X-100 micelles.
3.3. ORAC Values of Single Antioxidants and Complex Mixtures in the Absence and Presence of Triton X-100 Micelles
In order to quantify the effect of Triton X-100 micelles, ORAC-PGR values of pure antioxidants and complex mixtures were estimated in the absence (ORAC-PGR) and presence of Triton X-100 micelles (ORAC-PGR
MIC). Gallate derivatives showed similar ORAC-PGR values, being 11.7, and 9.8 the ORAC-PGR value of gallic acid, and octyl gallate, respectively (
Table 1). The similarity of ORAC-PGR values of gallic acid and gallate derivatives of different hydrophobicity can be related to their common reactive centre. ORAC-PGR
MIC values of the gallate derivatives are also given in
Table 1. It is observed an inverse dependence with the chain length. Thus, ORAC-PGR
MIC index of the gallate derivatives follows the order: Gallic acid > methyl gallate > propyl gallate > octyl gallate.
Table 1.
ORAC-PGR and ORAC-PGRMIC values of pure antioxidants.
Table 1.
ORAC-PGR and ORAC-PGRMIC values of pure antioxidants.
| Compound | ORAC-PGR | ORAC-PGRMIC | ORAC-PGR/ORAC-PGRMIC |
|---|
| Gallic acid | 11.7± 0.5 | 10.9 ± 0.4 | 1.1 ± 0.1 |
| Methyl gallate | 12.0 ± 0.4 | 7.0 ± 0.6 | 1.7 ± 0.2 |
| Propyl gallate | 12.3 ± 0.2 | 6.3 ± 0.5 | 1.9 ± 0.2 |
| Octyl gallate | 9.8 ± 0.3 | 1.6 ± 0.2 | 6.1 ± 0.9 |
| Caffeic acid | 0.4 ± 0.05 | 0.4 ± 0.07 | 1.0 ± 0.3 |
| Ethyl caffeate | 0.4 ± 0.06 | 0.2 ± 0.01 | 2.0 ± 0.4 |
| Quercetin | 11.9 ± 0.4 | 4.1 ± 0.2 | 2.9 ± 0.2 |
| Ascorbic acid | 10.3 ± 0.3 | 10.7 ± 0.5 | 1.0 ± 0.08 |
| β-carotene | ---- | ≈ 0.1 | --- |
| Trolox | 1 | 1 | 1 |
This result can be related to the lipophilicity of gallate derivatives and hence is related to their distribution upon Triton X-100 micelles. Thus, ORAC-PGR
MIC values are inversely related to the distribution of the antioxidants in Triton X-100 micelles. In fact, the reported values for the partition constant of gallate derivatives between Triton X-100 micelles and aqueous solutions range from 97 to 440 M
-1 for methyl and octyl gallate, respectively [
20]. A similar effect of Triton X-100 micelles was observed for the couple caffeic acid / ethyl caffeate (
Table 1). While the ORAC-PGR value of caffeic acid was not altered by Triton X-100 micelles (0.4 in both media), ORAC-PGR
MIC of ethyl caffeate was only one half of its ORAC-PGR value (ORAC-PGR
MIC = 0.2). In addition, ORAC-PGR and ORAC-PGR
MIC values of quercetin and ascorbic acid showed that while the antioxidant capacity of quercetin decreased significantly in presence of Triton X-100 micelles, no effect of micelles was observed for the ascorbic acid antioxidant ability. These results imply that the comparison of ORAC-PGR and ORAC-PGR
MIC values reflects the distribution of the antioxidant in Triton X-100 micelles. Thus, the ratio ORAC-PGR/ORAC-PGR
MIC can be considered as an index related to the lipophilicity of the tested antioxidant(s). As shown in
Table 1, ORAC-PGR/ORAC-PGR
MIC values of gallate derivatives between 1.0 and 6.1 for gallic acid and octyl gallate, respectively, were obtained. In addition, ORAC-PGR/ORAC-PGR
MIC values higher than one were obtained for antioxidants such as ethyl caffeate, and quercetin, compounds that can be considered to partition between the micelles and the dispersium solvent.
The obtained results (collected in
Table 1) employing single antioxidants suggest that ORAC-PGR and ORAC-PGR
MIC assays could be used to estimate the antioxidant capacity and the distribution of antioxidants in Triton X-100 micelles of complex mixtures such as wines, tea infusion, fruit extracts, and juices. As can be seen in
Table 2, ORAC-PGR values of these mixtures ranged from 2 to 36 mM Trolox equivalents, values that are related to the antioxidant activity of the samples. These ORAC-PGR values were influenced in different grades by the presence of Triton X-100 micelles. In fact, ORAC-PGR
MIC values of wines, tea infusions, and seed fruit extracts, were lower than ORAC-PGR values, implying that a fraction of the antioxidants contained in the samples were associated to Triton X-100 micelles. A different result was obtained for orange juice and fruit pulp extracts, where no effect of micelles was observed, indicating minimal incorporation of their antioxidants into the micellar pseudophase. This result is compatible with a predominance of hydrophilic antioxidants such as ascorbic acid in orange juice and fruit extracts.
Table 2.
ORAC-PGR and PRAC-PGRMIC values of complex mixtures a.
Table 2.
ORAC-PGR and PRAC-PGRMIC values of complex mixtures a.
| Complex mixture | ORAC-PGR | ORAC-PGRMIC | ORAC-PGR / ORAC-PGRMIC |
|---|
| Red wine | 36 ± 2 | 24 ± 3 | 1.5 ± 0.3 |
| White wine | 4 ± 1 | 3 ± 0.5 | 1.3 ± 0.5 |
| Tea extract | 16 ± 2 | 11 ± 1 | 1.5 ± 0.3 |
| Seed extract of Eugenia jambolana | 10 ± 1 | 8 ± 0.6 | 1.3 ± 0.2 |
| Pulp extract of Eugenia jambolana | 2 ± 0.3 | 2 ± 0.4 | 1.0 ± 0.3 |
| Seed extract of Myrciaria cauliflora | 15 ± 1 | 12 ± 2 | 1.3 ± 0.3 |
| Pulp extract of Myrciaria cauliflora | 5 ± 1 | 6 ± 1 | 0.8 ± 0.3 |
| Orange juice | 19 ± 3 | 21 ± 2 | 0.9 ± 0.2 |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
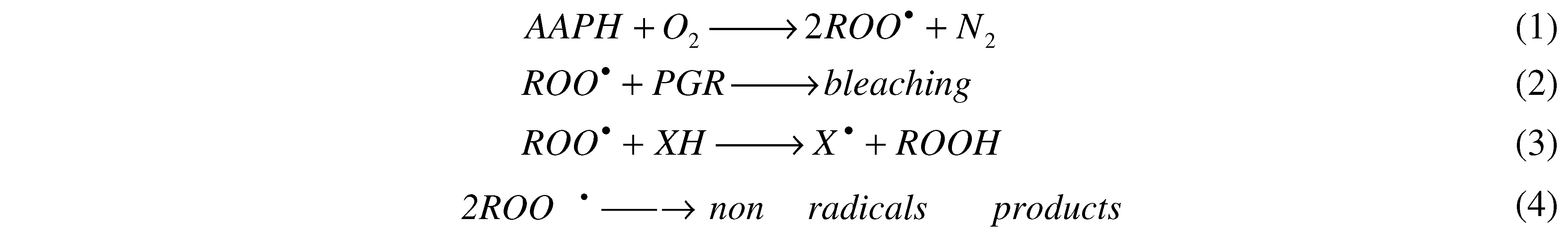{kind=link}
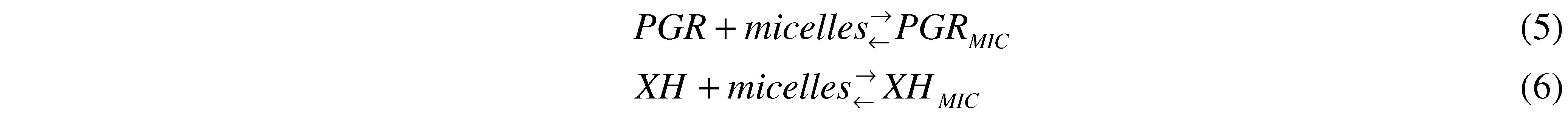{kind=link}

