Potent Effects of Flavonoid-Rich Extract from Rosa laevigata Michx Fruit against Hydrogen Peroxide-Induced Damage in PC12 Cells via Attenuation of Oxidative Stress, Inflammation and Apoptosis


Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of the TFs on H2O2-Induced Injury in PC12 Cells
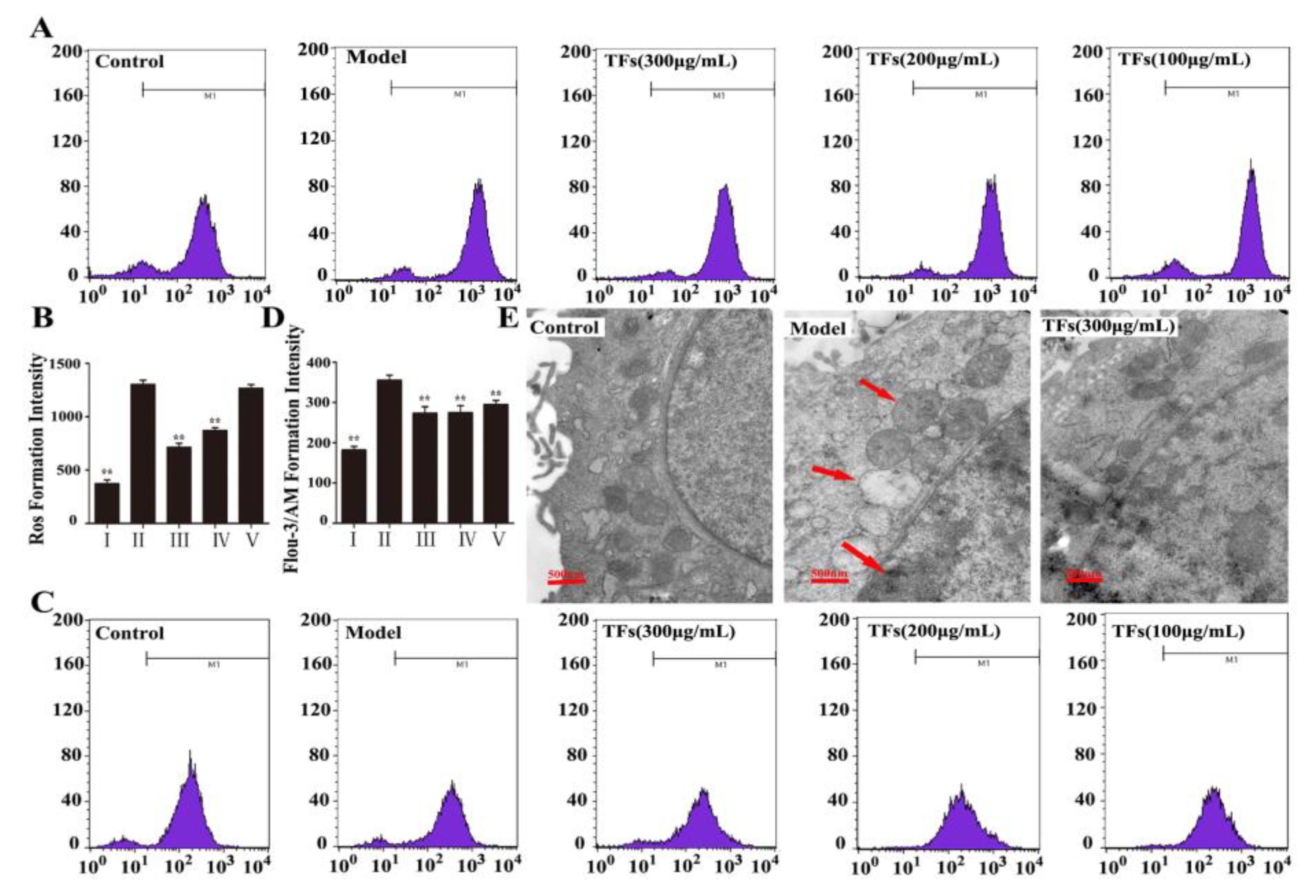
2.2. Effects of the TFs on the Levels of H2O2-Induced ROS and Ca2+ Release
2.3. Effects of the TFs on Ultrastructure Changes Caused by H2O2

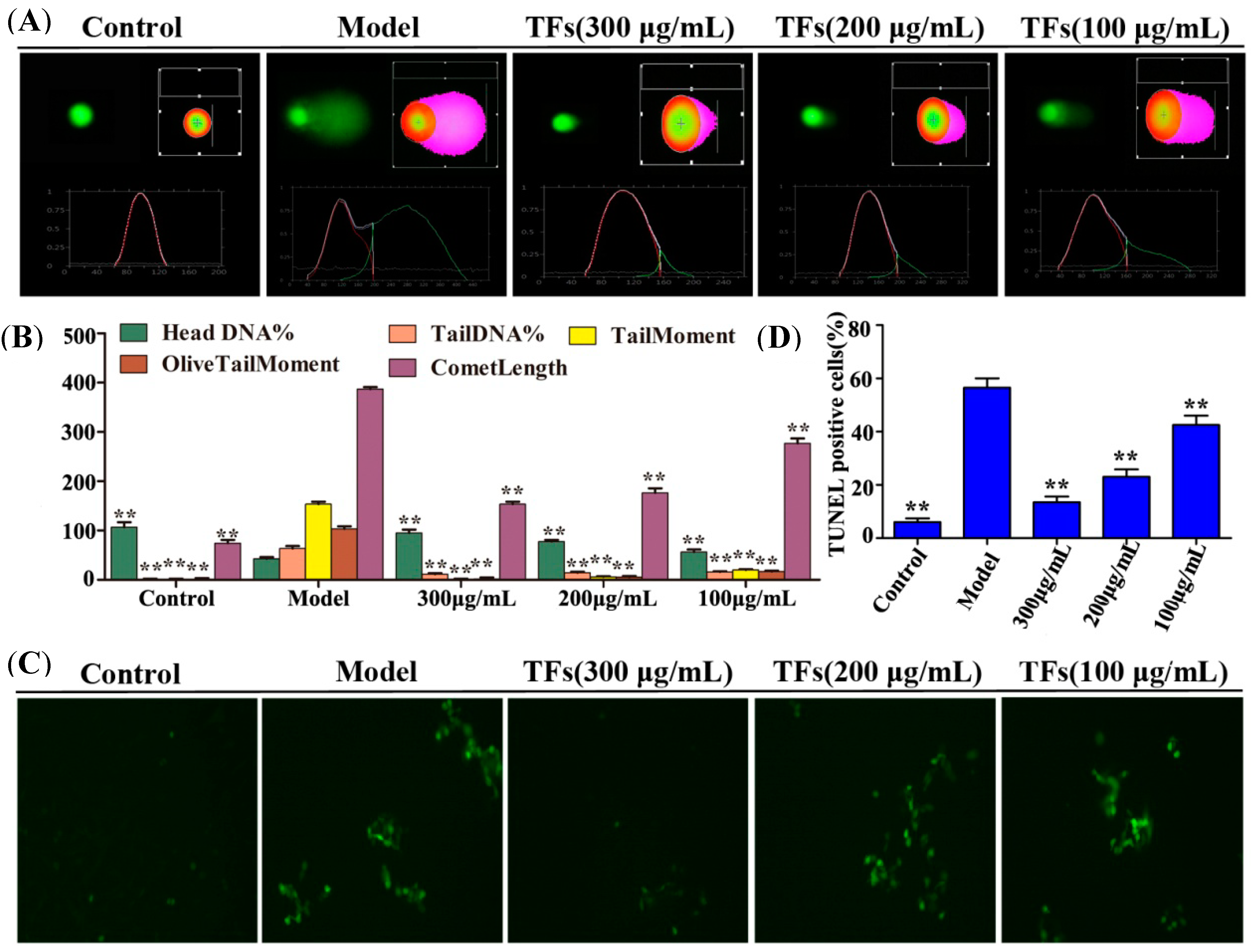
2.4. TFs Alleviated H2O2-Induced DNA Damage
2.5. TFs Reduced H2O2-Induced Apoptosis of PC12 Cells


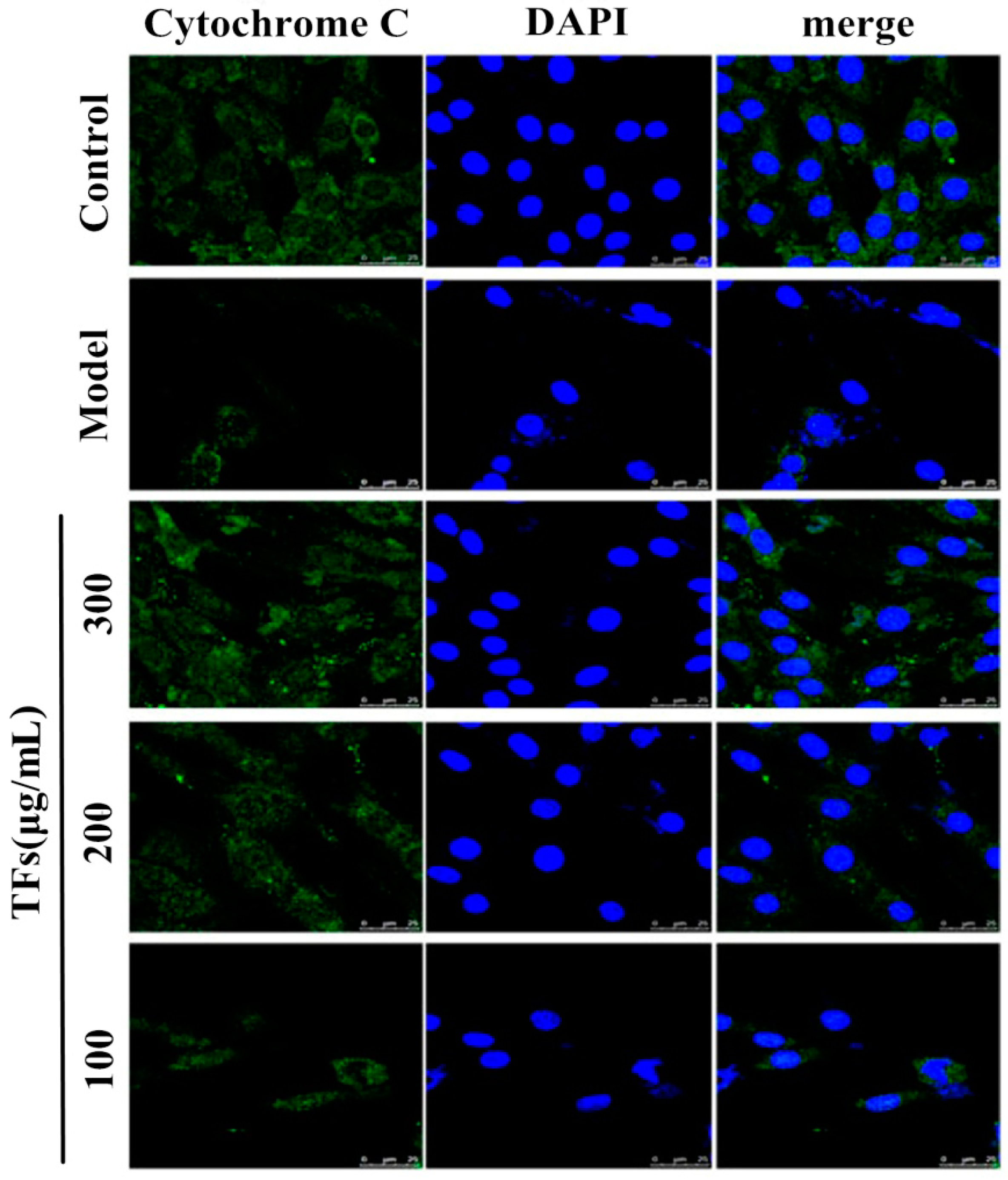
2.6. TFs Inhibited Cytochrome C Release
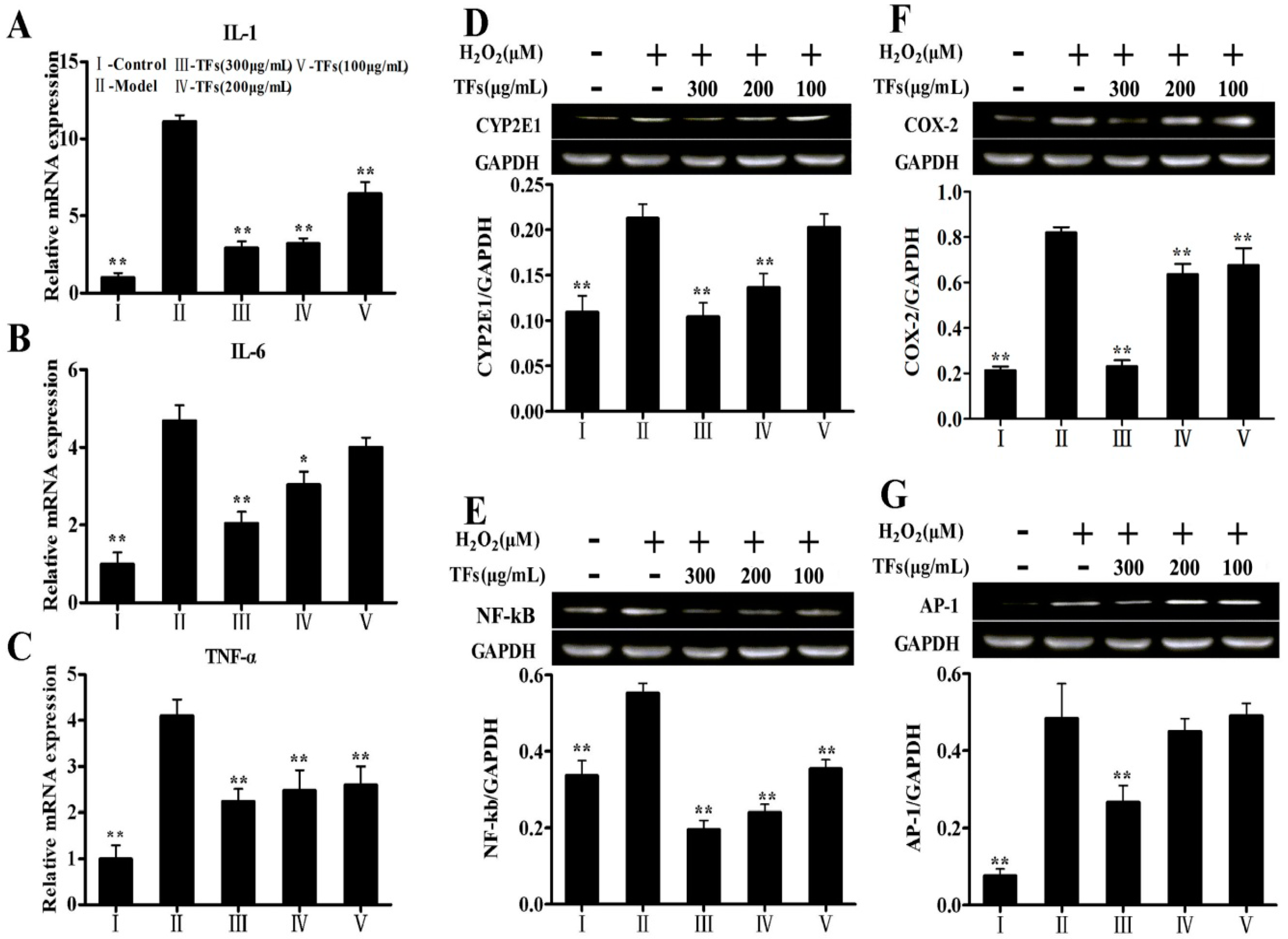
2.7. Effects of the TFs on the Expressions of H2O2-Induced Inflammatory Mediators

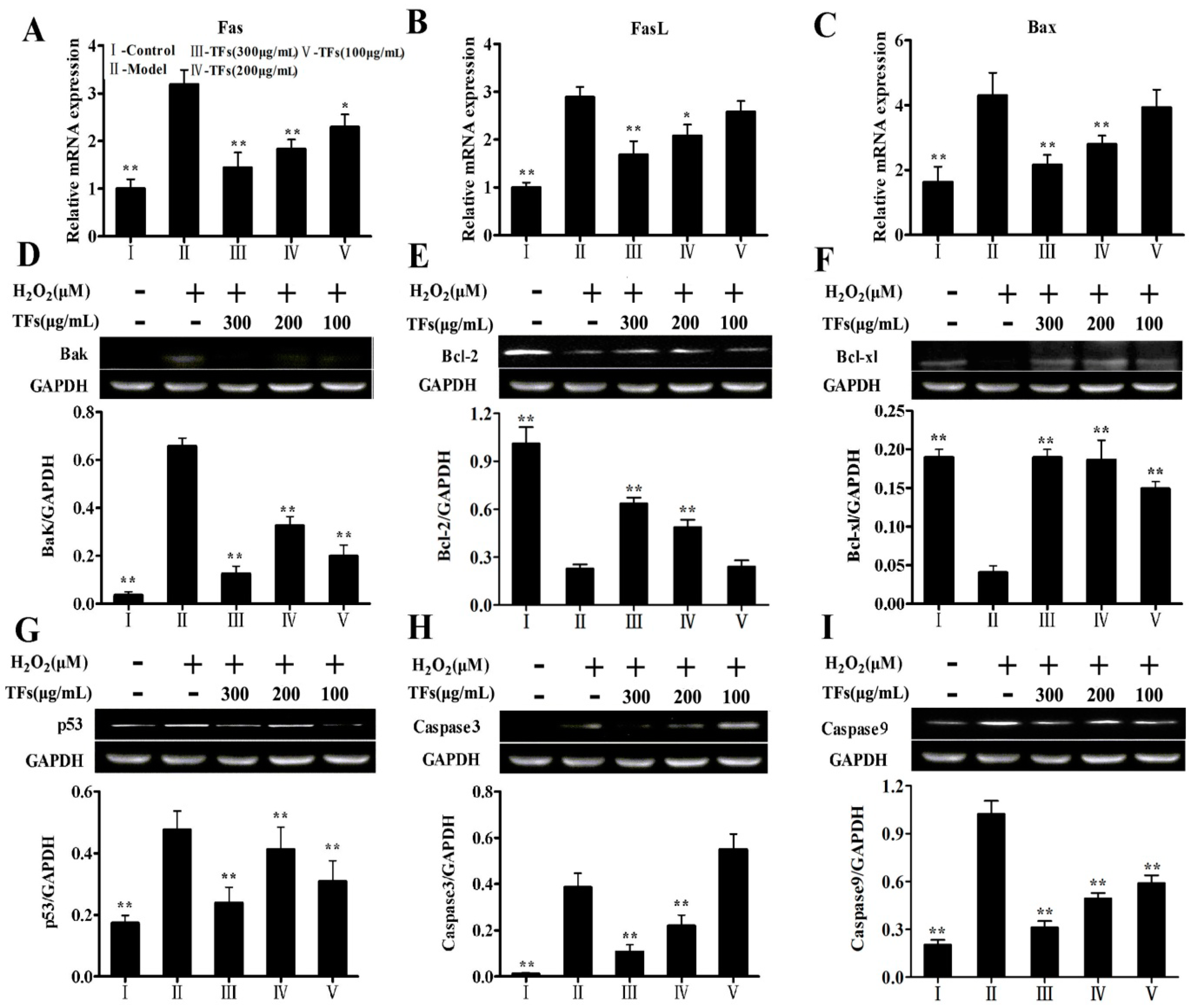
2.8. Effects of the TFs on the Expressions of H2O2-Induced Apoptotic Molecules
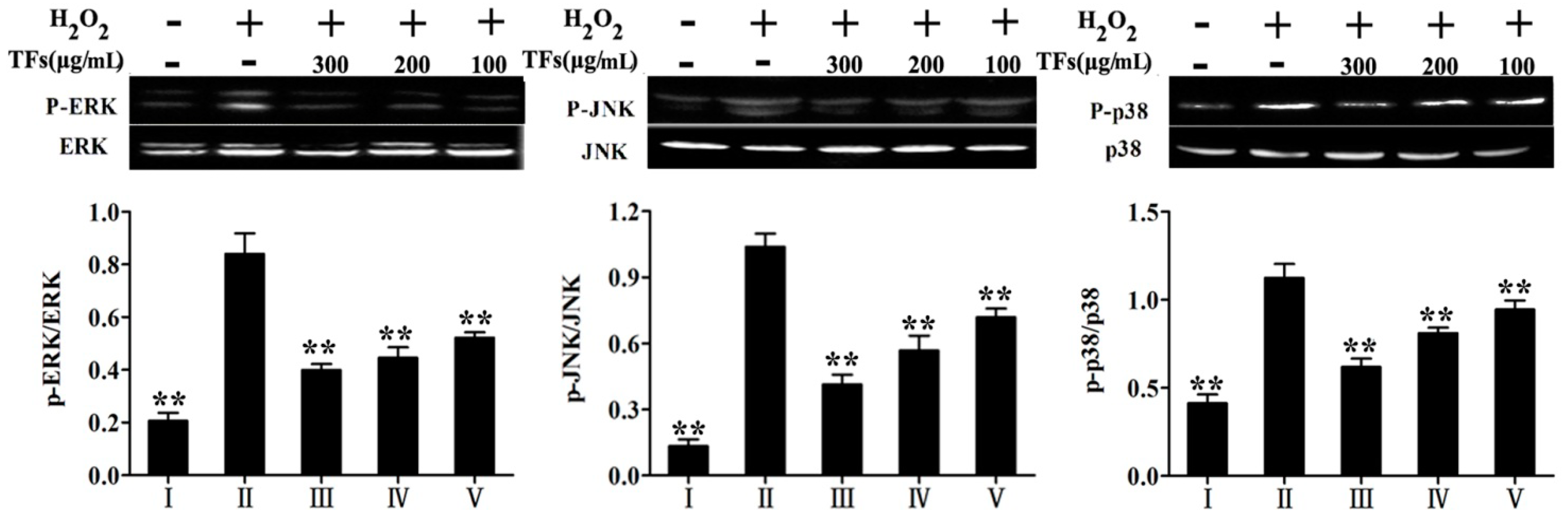
2.9. TFs Attenuated H2O2-Induced the Levels of MAPKs Phosphorylation
2.10. Discussion



3. Experimental
3.1. Chemicals and Reagents
3.2. Herbal Material and Preparation of the Total Flavonols
3.3. Cell Culture
3.4. Assay of Cell Viability
3.5. AO/EB and DAPI Staining
3.6. Single Cell Gel Electrophoresis Assay
3.7. TUNEL Assay
3.8. Detection of Cytochrome C Release
3.9. Detection of Intracellular ROS Accumulation and Ca2+ Release
3.10. Transmission Electron Microscope (TEM) Assay
3.11. Quantitative Real-time PCR Analyses
3.12. Western Blotting Assay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Dilutions | Company |
|---|---|---|---|
| GAPDH | rat | 1:5,000 | Proteintech Group, Chicago, IL, USA |
| CYP2E1 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| COX-2 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| HMGB-1 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| NF-κB | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| AP-1 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL,.USA |
| Bak | rabbit | 1:1,000 | Proteintech Group, Chicago, IL,.USA |
| Bcl-2 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| Bcl-x | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| p53 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| Caspase-3 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| Caspase-9 | rabbit | 1:1,000 | Proteintech Group, Chicago, IL, USA |
| p-p38 | rabbit | 1:1,000 | Bioworld Technology, St. Louis Park, MN, USA |
| p-38 | rabbit | 1:1,000 | Bioworld Technology, St. Louis Park, MN, USA |
| p-ERK | rabbit | 1:1,000 | Bioworld Technology, St. Louis Park, MN, USA |
| ERK | rabbit | 1:1,000 | Bioworld Technology, St. Louis Park, MN, USA |
| p-JNK | rabbit | 1:1,000 | Bioworld Technology, St. Louis Park, MN, USA |
| JNK | rabbit | 1:1,000 | Bioworld Technology, St. Louis Park, MN, USA |
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Andersen, J.K. Oxidative stress in neurodegeneration: Cause or consequence. Nat. Med. 2004, 10, S18–S25. [Google Scholar] [CrossRef]
- Heistad, D.D. Oxidative stress and vascular disease: 2005 Duff lecture. Arterioscl. Throm. Vas. 2006, 26, 689–695. [Google Scholar] [CrossRef]
- Devasagayam, T.P.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Physician. Indian 2004, 52, 794–804. [Google Scholar]
- Hool, L.C.; Corry, B. Redox control of calcium channels: From mechanisms to therapeutic opportunities. Antioxid. Redox Sign. 2007, 9, 409–435. [Google Scholar] [CrossRef]
- Haorah, J.; Ramirez, S.H.; Schall, K.; Smith, D.; Pandya, R.; Persidsky, Y. Oxidative stress activates protein tyrosine kinase and matrix metalloproteinases leading to blood-brain barrier dysfunction. J. Neurochem. 2007, 101, 566–576. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef]
- Yu, Y.; Du, J.R.; Wang, C.Y.; Qian, Z.M. Protection against hydrogen peroxide-induced injury by Z-ligustilide in PC12 cells. Exp. Brain Res. 2008, 184, 307–312. [Google Scholar] [CrossRef]
- Liu, S.L.; Lin, X.; Shi, D.Y.; Cheng, J.; Wu, C.Q.; Zhang, Y.D. Reactive oxygen species stimulated human hepatoma cell proliferation via cross-talk between PI3-K/PKB and JNK signaling pathways. Arch. Biochem. Biophys. 2002, 406, 173–182. [Google Scholar] [CrossRef]
- Sen, C.K.; Packer, L. Antioxidant and redox regulation of gene transcription. FASEB J. 1996, 10, 709–720. [Google Scholar]
- Jonas, W.B. Alternative medicine-learning from the past, examining the present, advancing to the future. J. Am. Med. Assoc. 1998, 280, 1616–1618. [Google Scholar] [CrossRef]
- Xu, J.D.; Yang, Y. Traditional Chinese medicine in the Chinese health care system. Health Policy 2009, 90, 133–139. [Google Scholar]
- Wang, S.P.; Wu, X.; Tan, M.; Gong, J.; Tan, W.; Bian, B.L.; Chen, M.; Wang, Y. Fighting fire with fire: Poisonous Chinese herbal medicine for cancer therapy. J. Ethnopharmacol. 2012, 140, 33–45. [Google Scholar] [CrossRef]
- Lim, G.P.; Chu, T.; Yang, F.; Beech, W.; Frautschy, S.A.; Cole, G.M. The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J. Neurosci. 2001, 21, 8370–8377. [Google Scholar]
- Gelinas, S.; Martinoli, M.G. Neuroprotective effect of estradiol and phytoestro-gens on MPP+-induced cytotoxicity in neuronal PC12 cells. J. Neurosci. Res. 2002, 70, 90–96. [Google Scholar]
- Park, S.Y.; Kim, H.S.; Cho, E.K.; Kwon, B.Y.; Phark, S.; Hwang, K.W.; Sul, D. Curcumin protected PC12 cells against beta-amyloid-induced toxicity through the inhibition of oxidative damage and tau hyperphosphorylation. Food Chem. Toxicol. 2008, 46, 2881–2887. [Google Scholar]
- Scapagnini, G.; Foresti, R.; Calabrese, V.; Giuffrida Stella, A.M.; Green, C.J.; Motterlini, R. Caffeic acid phenethyl ester and curcumin: A novel class of heme oxygenase-1 inducers. Mol. Pharmacol. 2002, 61, 554–561. [Google Scholar]
- Chen, C.Y.; Jang, J.H.; Li, M.H.; Surh, Y.J. Resveratrol upregulates heme oxygenase-1 expression via activation of NF-E2-related factor 2 in PC12 cells. Biochem. Biophys. Res. Commun. 2005, 331, 993–1000. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, B.; Han, X.; Xu, L.; Qi, Y.; Yin, L.; Xu, Y.; Zhao, Y.; Liu, K.; Peng, J. Protection of the flavonoid fraction from Rosa laevigata Michx fruit against carbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. 2013, 55, 60–69. [Google Scholar]
- Zhang, S.; Zheng, L.; Dong, D.; Xu, L.; Yin, L.; Qi, Y.; Han, X.; Lin, Y.; Liu, K.; Peng, J. Effects of flavonoids from Rosa laevigata Michx fruit against high-fat diet-induced non-alcoholic fatty liver disease in rats. Food Chem. 2013, 141, 2108–2116. [Google Scholar] [CrossRef]
- Jia, Y.; Ji, L.; Zhang, S.; Xu, L.; Yin, L.; Li, L.; Zhao, Y.; Peng, J. Total flavonoids from Rosa laevigata Michx fruit attenuates hydrogen peroxide induced injury in human umbilical vein endothelial cells. Food Chem. Toxicol. 2012, 50, 3133–3141. [Google Scholar] [CrossRef]
- Liu, S.; Han, Y.; Zhang, T.; Yang, Z. Protective effect of trifluoperazine on hydrogen peroxide-induced apoptosis in PC12 cells. Brain Res. Bull. 2011, 84, 183–188. [Google Scholar] [CrossRef]
- Zhang, S.; Qi, Y.; Xu, Y.; Han, X.; Peng, J.; Liu, K.; Sun, C. Protective effect of flavonoid-rich extract from Rosa laevigata Michx on cerebral ischemia-reperfusion injury through suppression of apoptosis and inflammation. Neurochem. Int. 2013, 63, 522–532. [Google Scholar] [CrossRef]
- Hinshaw, D.B.; Miller, M.T.; Omann, G.M.; Beals, T.F.; Hyslop, P.A. A cellular model of oxidant-mediated neuronal injury. Brain Res. 1993, 615, 13–26. [Google Scholar]
- Lu, Q.P.; Tian, L. Fas mRNA expression and calcium influx change in H2O2- induced apoptotic hepatocytes in vitro. World J. Gastroenterol. 2005, 11, 534–537. [Google Scholar]
- Sato, H.; Takeo, T.; Liu, Q.; Nakano, K.; Osanai, T.; Suga, S.; Wakui, M.; Wu, J. Hydrogen peroxide mobilizes Ca2+ through two distinct mechanisms in rat hepatocytes. Acta Pharmacol. Sin. 2009, 30, 78–89. [Google Scholar] [CrossRef]
- Gao, Z.; Huang, K.; Xu, H. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in HSSY5Y cells. Pharmacol. Res. 2001, 43, 173–178. [Google Scholar] [CrossRef]
- Ren, G.; Qiao, H.X.; Yang, J.; Zhou, C.X. Protective effects of steroids from Allium chinense against H2O2-induced oxidative stress in rat cardiac H9C2 cells. Phytother. Res. 2010, 24, 404–409. [Google Scholar] [CrossRef]
- Cao, Q.; Mak, K.; Lieber, C. Cytochrome P4502E1 primes macrophages to increase TNF-alpha production in response to liposaccharide. Am. J. Physiol. 2005, 289, G95–G107. [Google Scholar]
- Mukhopadhyay, P.; Rajesh, M.; Horváth, B.; Bátkai, S.; Park, O.; Tanchian, G.; Gao, R.Y.; Patel, V.; Wink, D.A.; Liaudet, L.; et al. Cannabidiol protects against hepatic ischemia/reperfusion injury by attenuating inflammatory signaling and response, oxidative/nitrative stress, and cell death. Free Radic. Biol. Med. 2011, 50, 1368–1381. [Google Scholar] [CrossRef]
- Liu, S.F.; Newton, R.; Evans, T.W.; Barnes, P.J. Differential regulation of cyclooxygenase-1 and cyclo-oxygenase-2 gene expression by lipopolysaccharide treatment in vivo in the rat. Clin. Sci. 1996, 90, 301–306. [Google Scholar]
- Silva, A.; Gomes, O. Oxidative DNA damage protection and repair by polyphen -olic compounds in PC12 cells. Eur. J. Pharmacol. 2008, 601, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef]
- Hollstein, M.; Sidransky, D.; Vogelstein, B.; Harris, C. P53 mutations in human cancers. Science 1991, 253, 49–53. [Google Scholar]
- Wang, I.; Lin-Shiau, S.; Lin, J. Induction of apoptosis by apigenin and related flavonoids through cytochrome c release and activation of caspase-9 and caspase-3 in leukaemia HL-60 cells. Eur. J. Cancer 1999, 35, 1517–1525. [Google Scholar] [CrossRef]
- Kong, M.; Ba, M.; Liang, H.; Ma, L.; Yu, Q.; Yu, T.; Wang, Y. 5'-Aza-dC sensitizes paraquat toxic effects on PC12 cell. Neurosci. Lett. 2012, 524, 35–39. [Google Scholar] [CrossRef]
- Yao, Z.; Wood, N.W. Cell death pathways in Parkinson’s disease: Role of mitochondria. Antioxid. Redox Sign. 2009, 11, 2135–2149. [Google Scholar]
- Ghibelli, L.; Diederich, M. Multistep and multitask bax activation. Mitochondrion 2010, 10, 604–613. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.J. Specificity of receptor tyrosine kinase signaling: Transient versus sustained extracellular signal-regulated kinase activation. Cell 1995, 80, 179–185. [Google Scholar]
- Davies, S.P.; Reddy, H.; Caivano, M.; Cohen, P. Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem. J. 2000, 351, 95–105. [Google Scholar]
- Zhang, S.; Zheng, L.; Xu, L.; Sun, H.; Li, H.; Yao, J.; Li, K.; Peng, J. Subchronic toxicity study of the total flavonoids from Rosa laevigata Michx fruit in rats. Regul. Toxicol. Pharmacol. 2012, 62, 221–230. [Google Scholar] [CrossRef]
- Beloosh, T.A.; John, A.; Tariq, S.; Raza, H. Increased mitochondrial stress and modulation of mitochondrial respiratory enzyme activities in acetaminophen-induced toxicity in mouse macrophage cells. Food Chem. Toxicol. 2010, 48, 2624–2632. [Google Scholar]
- Sample Availability: Samples of compounds total flavonoids (TFs) from Rosa laevigata Michx fruit are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, M.; Xu, Y.; Han, X.; Liang, C.; Yin, L.; Xu, L.; Qi, Y.; Zhao, Y.; Peng, J.; Sun, C. Potent Effects of Flavonoid-Rich Extract from Rosa laevigata Michx Fruit against Hydrogen Peroxide-Induced Damage in PC12 Cells via Attenuation of Oxidative Stress, Inflammation and Apoptosis. Molecules 2014, 19, 11816-11832. https://doi.org/10.3390/molecules190811816
Liu M, Xu Y, Han X, Liang C, Yin L, Xu L, Qi Y, Zhao Y, Peng J, Sun C. Potent Effects of Flavonoid-Rich Extract from Rosa laevigata Michx Fruit against Hydrogen Peroxide-Induced Damage in PC12 Cells via Attenuation of Oxidative Stress, Inflammation and Apoptosis. Molecules. 2014; 19(8):11816-11832. https://doi.org/10.3390/molecules190811816
Chicago/Turabian StyleLiu, Min, Youwei Xu, Xu Han, Chen Liang, Lianhong Yin, Lina Xu, Yan Qi, Yanyan Zhao, Jinyong Peng, and Changkai Sun. 2014. "Potent Effects of Flavonoid-Rich Extract from Rosa laevigata Michx Fruit against Hydrogen Peroxide-Induced Damage in PC12 Cells via Attenuation of Oxidative Stress, Inflammation and Apoptosis" Molecules 19, no. 8: 11816-11832. https://doi.org/10.3390/molecules190811816
APA StyleLiu, M., Xu, Y., Han, X., Liang, C., Yin, L., Xu, L., Qi, Y., Zhao, Y., Peng, J., & Sun, C. (2014). Potent Effects of Flavonoid-Rich Extract from Rosa laevigata Michx Fruit against Hydrogen Peroxide-Induced Damage in PC12 Cells via Attenuation of Oxidative Stress, Inflammation and Apoptosis. Molecules, 19(8), 11816-11832. https://doi.org/10.3390/molecules190811816