Effects of Climatic Conditions and Soil Properties on Cabernet Sauvignon Berry Growth and Anthocyanin Profiles

Abstract
:1. Introduction
2. Results and Discussion
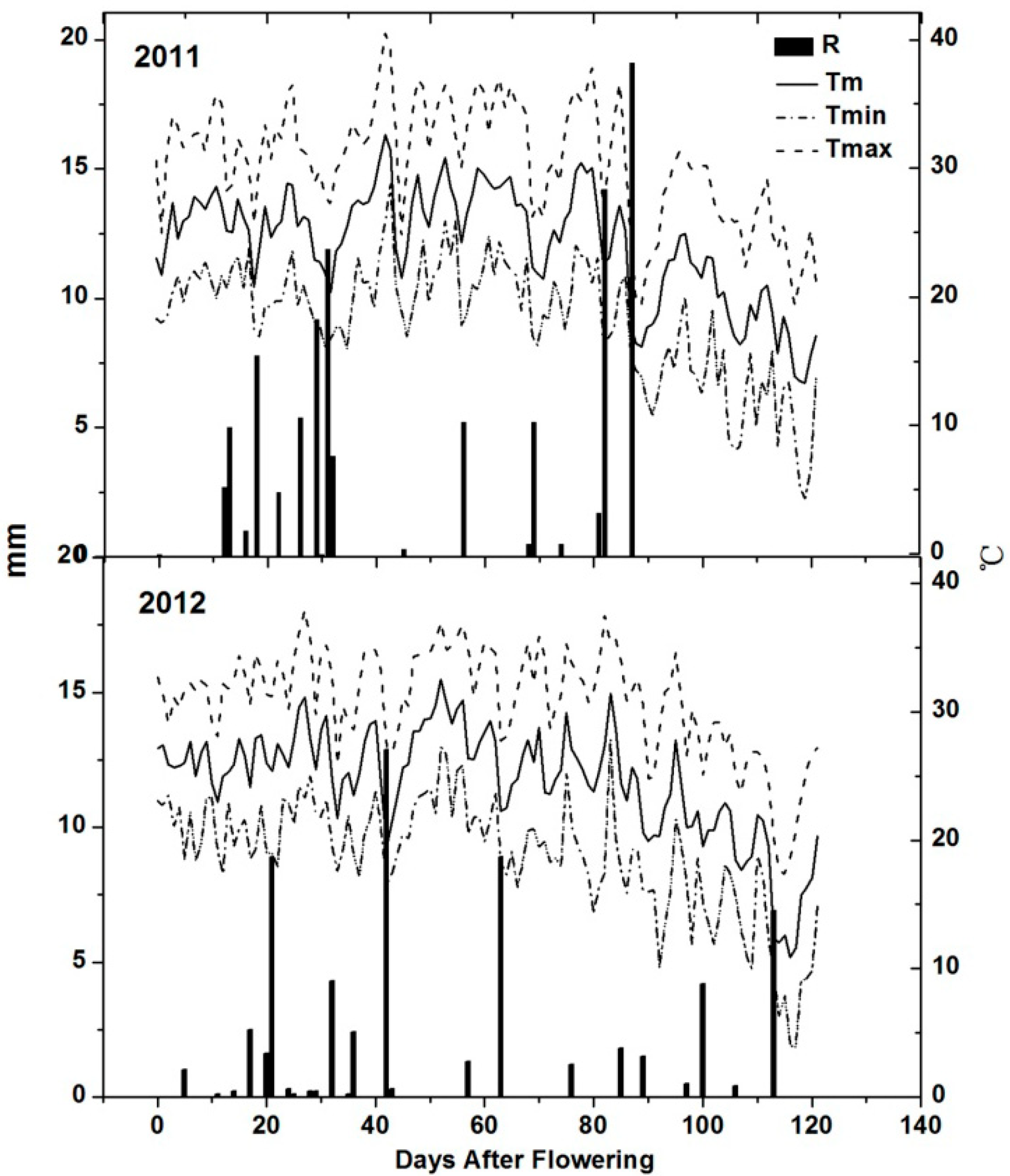
2.1. Climatic Conditions
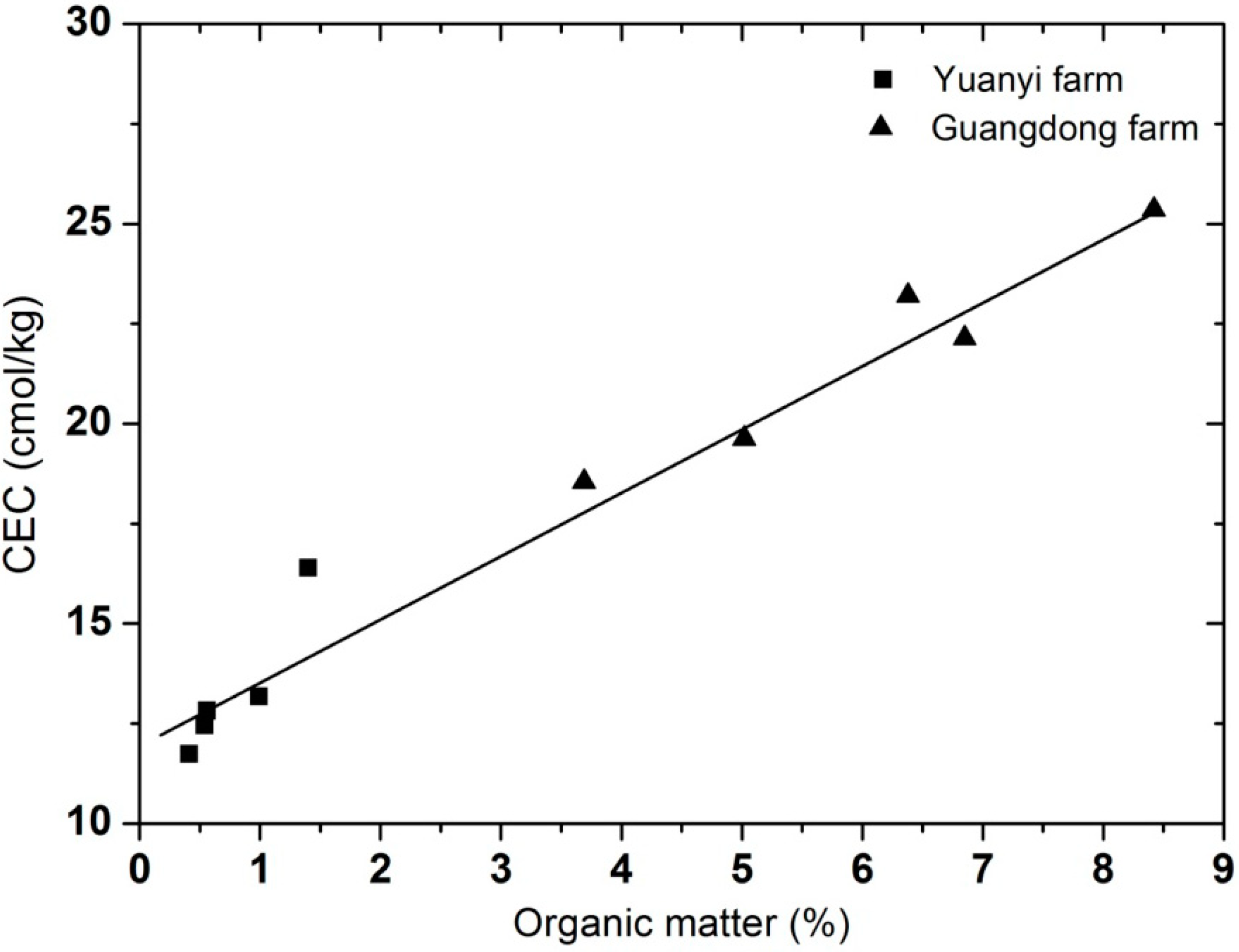
2.2. Soil Chemical and Physical Properties Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Mean Temperature ( °C) | Average Maximum Temperature ( °C) | Average Minimum Temperature ( °C) | Growing Degree Days ( °C) | Sunlight Duration (h) | Rain (mm) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aug-Sep | Apr-Oct | Year | Aug-Sep | Apr-Oct | Year | Aug-Sep | Apr-Oct | Year | Aug-Sep | Apr-Oct | Year | Aug-Sep | Apr-Oct | Year | Aug-Sep | Apr-Oct | Year | |
| 2011 | 22.1 | 20.3 | 8.2 | 28.7 | 26.8 | 13.6 | 16.1 | 14.6 | 3.4 | 740.9 | 2290.6 | 2290.6 | 627.4 | 2173.7 | 2899.0 | 41.2 | 180.1 | 217.8 |
| 2012 | 22.3 | 20.8 | 8.8 | 29.2 | 27.4 | 13.9 | 16.1 | 14.7 | 3.1 | 752.9 | 2356.2 | 2367.2 | 647.3 | 2235.8 | 3102.0 | 29.0 | 103.6 | 179.8 |

| Vineyards | Depth (cm) | Clay (%) | Slit (%) | Sand (%) | Textural Class | pH | EC (ms/cm) | CEC (cmol/kg) | Organic Matter (%) | Bulk Density (kg/m3) | Water Content (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Yuanyi farm | 0–20 | 8.59b | 49.40b | 41.99a | Silt loam | 7.9a | 0.18b | 16.40b | 1.40b | 1.49a | 18.82b |
| 20–40 | 9.82b | 61.66b | 28.52a | Silt loam | 7.8a | 0.20b | 13.18b | 0.99b | 1.65a | 26.49b | |
| 40–60 | 11.37a | 54.13a | 34.50a | Silt loam | 8.1a | 0.19b | 12.82b | 0.56b | 1.41a | 26.83b | |
| 60–80 | 14.63a | 49.87b | 35.49a | Silt loam | 8.0a | 0.20b | 12.46b | 0.54b | 1.50a | 25.61b | |
| 80–100 | 9.02a | 55.72b | 35.26a | Silt loam | 7.9a | 0.18b | 11.74b | 0.41b | 1.55a | 27.97b | |
| Guangdong farm | 0–20 | 15.99a | 63.97a | 20.03b | Silt loam | 7.5b | 0.39a | 23.21a | 6.38a | 0.98b | 29.55a |
| 20–40 | 11.45a | 63.51a | 25.04b | Silt loam | 7.3b | 0.54a | 25.36a | 8.42a | 1.21b | 44.04a | |
| 40–60 | 8.30b | 55.69a | 36.01a | Silt loam | 7.4b | 0.56a | 22.14a | 6.85a | 1.29b | 38.89a | |
| 60–80 | 9.17b | 54.35a | 36.48a | Silt loam | 7.2b | 0.98a | 19.63a | 5.02a | 1.47b | 36.13a | |
| 80–100 | 9.68a | 58.12a | 32.20b | Silt loam | 7.2b | 0.66a | 18.55a | 3.69a | 1.51b | 33.74a |

| Vineyards | Depth (cm) | Total Elements (%) | Effective Elements (mg/kg) | Available Elements (mg/kg) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Na | Ca | Mg | N | P | K | Cu | Fe | Mn | Zn | B | ||
| Yuanyi farm | 0–20 | 0.04b | 0.08a | 1.83a | 21.07b | 3494.46b | 140.62b | 50.86b | 10.10b | 133.29b | 0.75b | 1.61b | 1.01a | 1.40a | 0.47b |
| 20–40 | 0.07b | 0.08a | 1.85a | 33.40b | 3689.73b | 219.12b | 76.93a | 3.09b | 122.70a | 0.85a | 1.09b | 0.59a | 1.29a | 0.11b | |
| 40–60 | 0.04b | 0.08a | 1.80a | 31.33b | 3811.77a | 233.70b | 59.05b | 1.45b | 123.05a | 0.75a | 0.89b | 0.65a | 0.66a | Nd | |
| 60–80 | 0.05b | 0.07a | 1.81a | 39.48b | 3760.86b | 266.62a | 53.24b | 4.33b | 152.14a | 0.72b | 0.83b | 0.65a | 0.38a | Nd | |
| 80–100 | 0.07b | 0.07a | 1.85a | 39.77b | 3800.32a | 295.58a | 55.61b | 4.36b | 134.72a | 0.76b | 1.10b | 0.91a | 0.41a | Nd | |
| Guangdong farm | 0–20 | 0.30a | 0.11a | 1.73b | 51.83a | 3789.53a | 525.98a | 61.74a | 75.42a | 175.10a | 0.85a | 19.68a | 0.58b | 1.35b | 2.51a |
| 20–40 | 0.40a | 0.10a | 1.34b | 57.28a | 3775.11a | 490.15a | 69.34b | 18.25a | 129.57a | 0.58b | 22.65a | 0.45b | 0.82b | 1.03a | |
| 40–60 | 0.30a | 0.06a | 1.40b | 66.10a | 3776.49b | 334.47a | 91.94a | 5.68a | 100.67b | 0.61b | 29.79a | 0.41b | 0.59b | 0.21 | |
| 60–80 | 0.24a | 0.06a | 1.57b | 63.41a | 3824.79a | 265.22a | 86.13a | 8.59a | 78.92b | 0.84a | 33.92a | 0.40b | 0.35b | 0.25 | |
| 80–100 | 0.19a | 0.08a | 1.66b | 52.72a | 3769.12b | 219.11b | 90.66a | 8.93a | 40.57b | 0.85a | 33.26a | 0.39b | 0.26b | 0.75 | |

2.3. Vine Water and Nitrogen Status
| Year | Vineyards | δ13C (‰) | N (%) | Leaf Chlorophyll |
|---|---|---|---|---|
| 2011 | Yuanyi farm | −25.34a | 0.47b | 45.01b |
| Guangdong farm | −26.76b | 0.54a | 46.95a | |
| 2012 | Yuanyi farm | −25.06a | 0.46b | 45.75b |
| Guangdong farm | −26.51b | 0.49a | 46.54a |
2.4. Some Parameters of Vine, Shoot, Cluster and Berries from Two Vineyards in Two Vintages
| Years | Vineyards | Survey of Shoots | Survey of Clusters | Yield (ton/hectare) | Yield/Pruning Weigh (kg/kg) | Leaf Area/Yield (m2/kg) | |||
|---|---|---|---|---|---|---|---|---|---|
| Average Shoot Length (cm) | Shoot Number/m | Cluster Number/Shoot | Cluster Weight (g) | Cluster Compactness (OIV rating) | |||||
| 2011 | Yuanyi farm | 122b a | 13.85a | 1.80a | 107.10b | 4.00b | 9.14a | 5.49a | 1.98a |
| Guangdong farm | 127a | 14.29a | 1.64b | 111.22a | 5.67a | 8.45a | 5.32a | 2.08a | |
| 2012 | Yuanyi farm | 132b | 13.55a | 1.79a | 107.97b | 3.67b | 10.48a | 5.66a | 2.03a |
| Guangdong farm | 136a | 15.38a | 1.58b | 119.20a | 5.33a | 11.59a | 5.78a | 2.12a | |

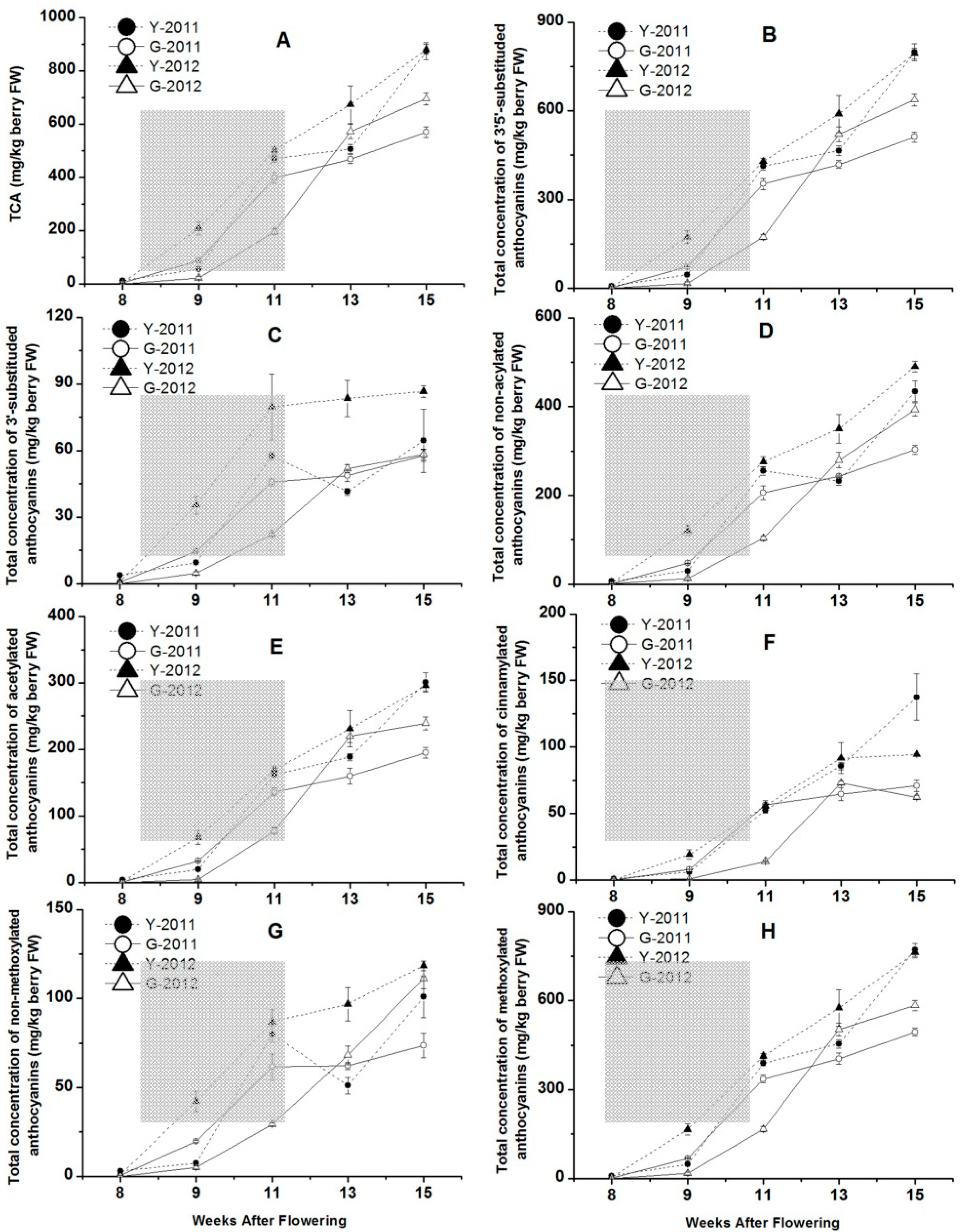
2.5. The Concentrations of Anthocyanins in Grape Skins from Two Vineyards in 2011 and 2012

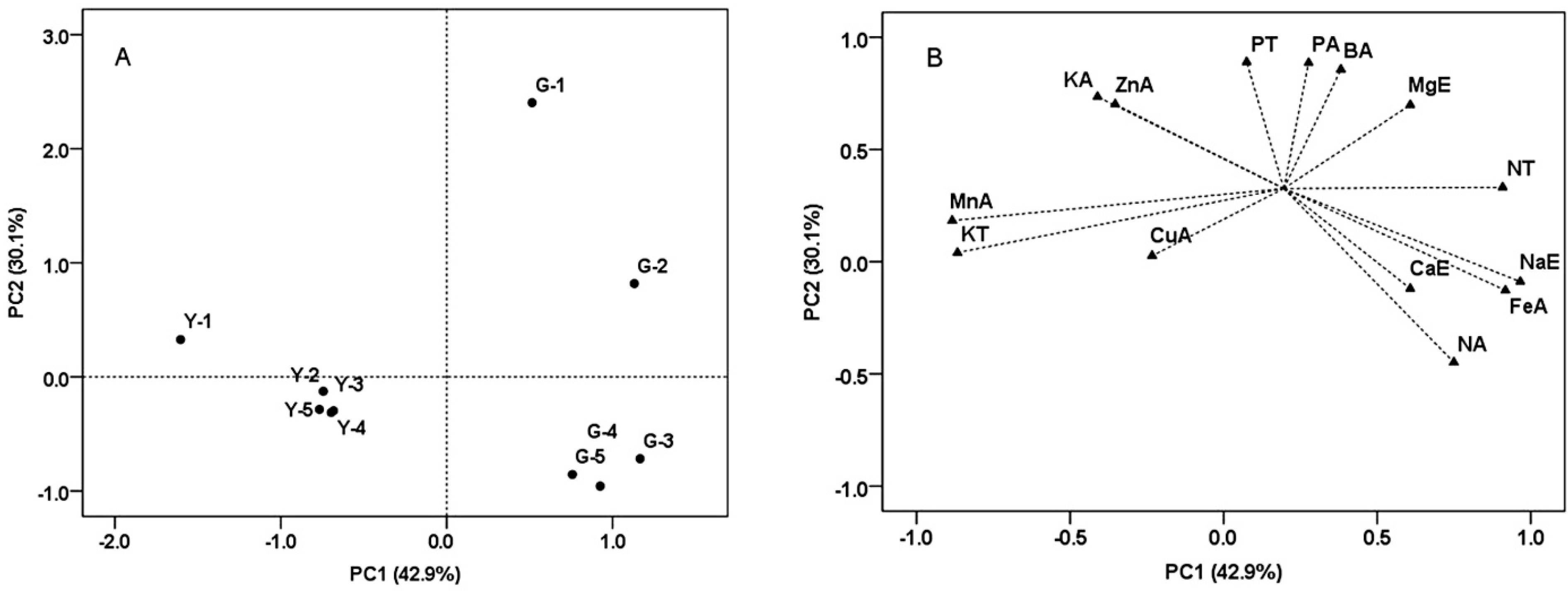
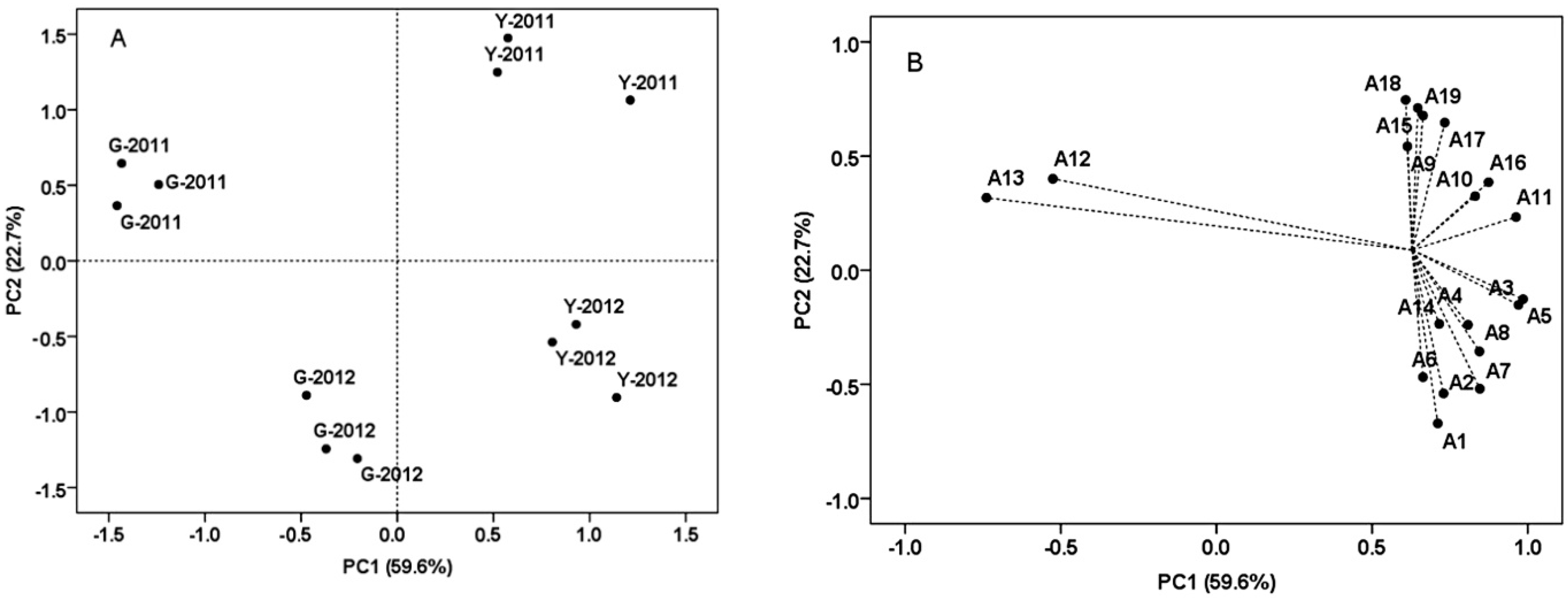
2.6. Principal Component Analysis (PCA) of the Grape Samples from Two Vineyards at Harvest

| Anthocyanins | 2011 | 2012 | ||
|---|---|---|---|---|
| Yuanyi farm (mg/kg berry FW) | Guangdong farm (mg/kg berry FW) | Yuanyi farm (mg/kg berry FW) | Guangdong farm (mg/kg berry FW) | |
| dephinidin-3-O-glucoside (A1) | 52.31 ± 3.7 | 37.65 ± 1.28 | 66.83 ± 0.86 | 63.20 ± 2.48 |
| cyanidin-3-O-glucoside (A2) | 6.97 ± 0.52 | 6.24 ± 0.27 | 8.49 ± 0.75 | 7.31 ± 0.21 |
| petunidin-3-O-glucoside (A3) | 42.36 ± 3.09 | 31.94 ± 1.61 | 43.40 ± 1.35 | 38.92 ± 1.29 |
| peonidin-3-O-glucoside (A4) | 28.71 ± 2.22 | 23.47 ± 1.39 | 39.28 ± 2.22 | 25.05 ± 1.44 |
| malvidin-3-O-glucoside (A5) | 302.77 ± 14.46 | 201.11 ± 6.17 | 332.49 ± 10.28 | 259.38 ± 10.72 |
| dephinidin-3-O-(6-O-acetyl)-glucoside (A6) | 24.94 ± 5.45 | 17.73 ± 5.19 | 28.22 ± 4.10 | 27.84 ± 3.43 |
| cyanidin-3-O-(6-O-acetyl)-glucoside (A7) | 5.45 ± 0.95 | 2.45 ± 0.18 | 7.91 ± 0.51 | 6.13 ± 0.65 |
| petunidin-3-O-(6-O-acetyl)-glucoside (A8) | 17.74 ± 1.76 | 12.51 ± 1.01 | 18.24 ± 0.49 | 17.95 ± 0.92 |
| dephinidin-3-O-(6-O-coumaryl)-glucoside (A9) | 7.67 ± 1.42 | 4.18 ± 0.40 | 4.98 ± 0.67 | 4.88 ± 0.47 |
| peonidin-3-O-(6-O-acetyl)-glucoside (A10) | 18.20 ± 0.86 | 13.99 ± 0.51 | 19.32 ± 0.34 | 12.58 ± 0.58 |
| malvidin-3-O-(6-O-acetyl)-glucoside (A11) | 234.36 ± 8.87 | 148.40 ± 2.63 | 221.82 ± 3.50 | 174.58 ± 5.01 |
| petunidin-3-O-(6-O-caffeoyl)-glucoside (A12) | 0.49 ± 0.15 | 1.07 ± 0.28 | 0.55 ± 0.00 | 0.31 ± 0.01 |
| cyanidin-3-O-(6-O-coumaryl)-glucoside (A13) | 2.05 ± 0.21 | 4.39 ± 0.54 | 2.00 ± 0.74 | 2.00 ± 0.76 |
| malvidin-3-O-(6-O-caffeoyl)-glucoside (A14) | 0.38 ± 0.10 | Nd | 1.21 ± 0.05 | Nd |
| petunidin-3-O-(6-O-coumaryl)-glucoside (A15) | 6.28 ± 0.43 | 3.75 ± 0.23 | 4.36 ± 0.07 | 3.60 ± 0.17 |
| peonidin-3-O-(cis-6-O-coumaryl)-glucoside (A16) | 0.65 ± 0.05 | 0.41 ± 0.01 | 0.64 ± 0.03 | 0.38 ± 0.01 |
| peonidin-3-O-(trans-6-O-coumaryl)-glucoside (A17) | 12.88 ± 1.99 | 6.85 ± 0.39 | 8.95 ± 0.32 | 4.85 ± 0.19 |
| malvidin-3-O-(cis-6-O-coumaryl)-glucoside (A18) | 4.88 ± 0.41 | 2.62 ± 0.16 | 3.65 ± 0.22 | 2.31 ± 0.09 |
| malvidin-3-O-(trans-6-O-coumaryl)-glucoside (A19) | 102.01 ± 16.81 | 47.64 ± 2.77 | 68.04 ± 0.60 | 43.95 ± 1.22 |
3. Experimental Section
3.1. The Experimental Site and Plant Material
3.2. Meteorological Survey
3.3. Soil Sampling and Analysis
3.4. Vine Water and Nutrient Status
3.5. Yield and Grape Composition
3.6. Extraction of Anthocyanins in Grape Skins
3.7. Chemicals and Standards
3.8. HPLC-MS Analyses of Anthocyanins
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgements
Author Contributions
Conflicts of Interest
References
- Downey, M.O.; Dokoozlian, N.K.; Krstic, M.P. Cultural practice and environmental impacts on the flavonoid composition of grapes and wine: A review of recent research. Am. J. Enol. Vitic. 2006, 57, 257–268. [Google Scholar]
- Kennedy, J.A. Managing Wine Quality: Viticulture and Wine Quality; Reynolds, A.G., Ed.; Woodhead Publishing: Cambridge, UK, 2010; pp. 73–104. [Google Scholar]
- Soubeyrand, E.; Basteau, C.; Hilbert, G.; van Leeuwen, C.; Delrot, S.; Gomès, E. Nitrogen supply affects anthocyanin biosynthetic and regulatory genes in grapevine cv. Cabernet-Sauvignon berries. Phytochemistry 2014, 103, 38–49. [Google Scholar] [CrossRef]
- Baldi, A.; Romani, A.; Mulinacci, N.; Vincieri, F.; Caseta, B. HPLC/MS Application to Anthocyanins of Vitis vinifera L. J. Agric. Food. Chem. 1995, 43, 2104–2109. [Google Scholar] [CrossRef]
- Monages, M.; Nunez, V.; Bartolome, B.; Gomez-Cordoves, C. Anthocyanin-derived pigments in Graciano, Tempranillo, and Cabernet Sauvignon wines produced in Spain. Am. J. Enol. Vitic. 2003, 54, 163–169. [Google Scholar]
- Deloire, A.; Vaudour, E.; Carey, V.; Bonnardot, V.; van Leeuwen, C. Grapevine responses to terroir, a global approach. J. Int. Vigne. Vin. 2005, 39, 149–162. [Google Scholar]
- Van Leeuwen, C.; Friant, C.; Friant, F.; Chone, X.; Tregoat, O.; Koundouras, S.; Dubourdieu, D. Influence of climate, soil and cultivar on terroir. Am. J. Enol. Vitic. 2004, 55, 207–217. [Google Scholar]
- Mazza, G. Anthocyanins in grape products. Crit. Rev. Food Sci. Nutr. 1995, 35, 341–371. [Google Scholar] [CrossRef]
- Pereira, G.E.; Gaudillere, J.P.; van Leeuwen, C.; Hilbert, G.; Maucourt, M.; Deborde, C. H1-NMR metabolite fingerprints of grape berry: Comparison of vintage and soil effects in Bordeaux grapevine growing areas. Anal. Chim. Acta 2006, 563, 346–352. [Google Scholar]
- Ferrandino, A.; Lovisolo, C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Cohen, S.D.; Tarara, J.M.; Kennedy, J.A. Diurnal Temperature Range Compression Hastens Berry Development and Modifies Flavonoid Partitioning in Grapes. Am. J. Enol. Vitic. 2012, 63, 112–120. [Google Scholar]
- Bucchetti, B.; Matthews, M.A.; Falginella, L.; Peterlungera, E.; Castellarin, S.D. Effect of water deficit on Merlot grape tannins and anthocyanins across four seasons. Sci. Hortic. 2011, 128, 297–305. [Google Scholar]
- Li, J.H.; Guan, L.; Fan, P.G.; Li, S.H.; Wu, B.H. Effect of Sunlight Exclusion at Different Phenological Stages on Anthocyanin Accumulation in Red Grape Clusters. Am. J. Enol. Vitic. 2013, 64, 349–356. [Google Scholar]
- Castellarin, S.D.; Matthews, M.A.; di Gaspero, G.; Gambetta, G.A. Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 2007, 227, 101–112. [Google Scholar]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell. Environ. 2007, 30, 1381–1399. [Google Scholar]
- Dokoozlian, N.K.; Kliewer, W.M. Influence of light on grape berry growth and composition varies during fruit development. J. Am. Soc. Hortic. Sci. 1996, 121, 869–874. [Google Scholar]
- Hunter, J.J.; Ruffner, H.P.; Volschenk, C.G.; le Roux, D.J. Partial defoliation of Vitis vinifera L. cv. Cabernet Sauvignon/99 Richter: Effect on root growth, canopy efficiency, grape composition, and wine quality. Am. J. Enol. Vitic. 1995, 46, 306–314. [Google Scholar]
- Bergqvist, J.; Dokoozlian, N.; Ebisuda, N. Sunlight exposure and temperature effects on berry growth and composition of Cabernet Sauvignon and Grenache in the central San Joaquin Valley of California. Am. J. Enol. Vitic. 2001, 52, 1–7. [Google Scholar]
- Conradie, W.J. Liming and choice of rootstocks as cultural techniques for vines in acid soils. S. Afr. J. Enol. Vitic. 1983, 4, 39–44. [Google Scholar]
- Josep, M.U.; Xavier, S.; Alicia, Z.; Rosa, M.P. Effects of soil and climatic conditions on grape ripening and wine quality of Cabernet Sauvignon. J. Wine Res. 2010, 21, 1–17. [Google Scholar]
- De Andrés-de Prado, R.; Yuste-Rojas, M.; Sort, X.; Andrés-Lacueva, C.; Torres, M.; Lamuela-Raventós, R.M. Effect of soil type on wines produced from Vitis vinifera L. cv. Grenache in commercial vineyards. J. Agric. Food. Chem. 2007, 55, 779–786. [Google Scholar] [CrossRef]
- Farquhar, G.; Ehleringer, J.; Hubick, K. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Gaudillère, J.P.; van Leeuwen, C.; Ollat, N. Carbon isotope composition of sugars in grapevine, an integrated indicator of vineyard water status. J. Exp. Bot. 2002, 53, 757–763. [Google Scholar]
- Van Leeuwen, C.; Trégoat, O.; Choné, X.; Bois, B.; Pernet, D.; Gaudillère, J.P. Vine water status is a key factor in grape ripening and vintage quality for red Bordeaux wine. How can it be assessed for vineyard management purposes? J. Int. Sci. Vigne. Vin. 2009, 43, 121–134. [Google Scholar]
- King, P.D.; Smart, R.E.; Mcclellan, D.J. Within-vineyard variability in vine vegetative growth, yield, and fruit and wine composition of Cabernet Sauvignon in Hawke’s Bay, New Zealand. Aust. J. Grape Wine Res. 2014, 20, 234–246. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Senchuk, I.V.; van der Reest, C.; de Savigny, C. Use of GPS and GIS for elucidation of the basis for terroir, spatial variation in an Ontario Riesling vineyard. Am. J. Enol. Viti. 2007, 8, 145–162. [Google Scholar]
- Reynolds, A.G.; Edwards, C.G.; Wardle, D.A.; Webster, D.R.; Dever, M. Shoot density affects “Riesling” grapevines I. Vine performance. J. Am. Soc. Hortic. Sci. 1994, 119, 874–880. [Google Scholar]
- Peyrot des Gachons, C.P.; van Leeuwen, C.; Tominaga, T.; Soyer, J.P.; Gaudillère, J.P.; Dubourdieu, D. Influence of water and nitrogen deficit on fruit ripening and aroma potential of Vitis vinifera L. cv Sauvignon blanc in field conditions. J. Sci. Food. Agric. 2005, 85, 73–85. [Google Scholar]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. The effect of maturity and vine water status on grape skin flavonoids. Am. J. Enol. Vitic. 2002, 3, 268–274. [Google Scholar]
- Jackson, D.I.; Lombard, P.B. Environmental and management-practices affecting grape composition and wine quality-a review. Am. J. Enol. Vitic. 1993, 44, 409–430. [Google Scholar]
- Mazza, G.; Fukumoto, L.; Delaquis, P.; Girard, B.; Ewert, B. Anthocyanins, Phenolics, and Color of Cabernet Franc, Merlot, and Pinot Noir Wines from British Columbia. J. Agric. Food Chem. 1999, 47, 4009–4017. [Google Scholar] [CrossRef]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of sunlight and temperature effects on the composition of Vitis Vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Barbagallo, M.G.; Guidoni, S.; Hunter, J.J. Berry size and qualitative characteristics of vitis vinifera. cv. Syrah. S. Afr. J. Enol. Vitic. 2011, 32, 129–136. [Google Scholar]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A; Deloire, A. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Vitic. 2002, 53, 261–267. [Google Scholar]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in winegrape composition: Anthocyanins and tannins. Aust. J. Grape. Wine Res. 2004, 10, 100–107. [Google Scholar]
- Chone, X.; van Leeuwen, C.; Chery, P.; Ribereau-Gayon, P. Terroir influence on water status and nitrogen status of non-irrigated Cabernet Sauvignon (Vitis vinifera). Vegetative development, must and wine composition (example of a Medoc top estate vineyard, Saint Julien area, Bordeaux, 1997). S. Afr. J. Enol. Vitic. 2001, 21, 8–15. [Google Scholar]
- Matsuyama, S.; Tanzawa, F.; Kobayashi, H.; Suzuki, S.; Takata, R.; Saito, H. Leaf Removal Accelerated Accumulation of Delphinidin-based Anthocyanins in “Muscat Bailey A” [Vitis × labruscana (Bailey) and Vitis vinifera (Muscat Hamburg)] Grape Skin. J. Jpn. Soc. Hortic. Sci. 2014, 83, 17–22. [Google Scholar] [CrossRef]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with healthpromoting properties. Annu. Rev. Food. Sci. Technol. 2010, 1, 163–187. [Google Scholar]
- Roggero, J.P.; Coen, S.; Ragonnet, R. High performance liquid chromatography survey on changes in pigment content in ripening grapes of Syrah. An approach to anthocyanin metabolism. Am. J. Enol. Vitic. 1986, 37, 77–83. [Google Scholar]
- Ryan, J.M.; Revilla, E. Anthocyanin composition of Cabernet Sauvignon and Tempranillo grapes at different stages of ripening. J. Agric. Food. Chem. 2003, 51, 3372–3378. [Google Scholar]
- Tarara, J.M.; Lee, J.; Spayd, S.E.; Scagel, C.F. Berry Temperature and Solar Radiation Alter Acylation, Proportion, and Concentration of Anthocyanin in Merlot Grapes. Am. J. Enol. Vitic. 2008, 59, 235–247. [Google Scholar]
- Soil Survey Staff. Soil Survey Manual; Soil Conservation Service: Washington, DC, USA, 1993.
- International Organisation of Vine and Wine. OIV Descriptor List for Grape Varieties and Vitis Species, 2nd ed.; OIV: Paris, France, 2007. [Google Scholar]
- He, J.J.; Liu, Y.X.; Pan, Q.H.; Cui, X.Y.; Duan, C.Q. Different anthocyanin profiles of the skin and the pulp of Yan73 (Muscat Hamburg × Alicante Bouschet) grape berries. Molecules 2010, 15, 1141–1153. [Google Scholar]
- Sample Availability: Samples of grapes are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cheng, G.; He, Y.-N.; Yue, T.-X.; Wang, J.; Zhang, Z.-W. Effects of Climatic Conditions and Soil Properties on Cabernet Sauvignon Berry Growth and Anthocyanin Profiles. Molecules 2014, 19, 13683-13703. https://doi.org/10.3390/molecules190913683
Cheng G, He Y-N, Yue T-X, Wang J, Zhang Z-W. Effects of Climatic Conditions and Soil Properties on Cabernet Sauvignon Berry Growth and Anthocyanin Profiles. Molecules. 2014; 19(9):13683-13703. https://doi.org/10.3390/molecules190913683
Chicago/Turabian StyleCheng, Guo, Yan-Nan He, Tai-Xin Yue, Jun Wang, and Zhen-Wen Zhang. 2014. "Effects of Climatic Conditions and Soil Properties on Cabernet Sauvignon Berry Growth and Anthocyanin Profiles" Molecules 19, no. 9: 13683-13703. https://doi.org/10.3390/molecules190913683
APA StyleCheng, G., He, Y.-N., Yue, T.-X., Wang, J., & Zhang, Z.-W. (2014). Effects of Climatic Conditions and Soil Properties on Cabernet Sauvignon Berry Growth and Anthocyanin Profiles. Molecules, 19(9), 13683-13703. https://doi.org/10.3390/molecules190913683