MALDI-TOF MS Analysis of Proanthocyanidins in Two Lowland Tropical Forest Species of Cecropia: A First Look at Their Chemical Structures

Abstract
:1. Introduction

2. Results and Discussion




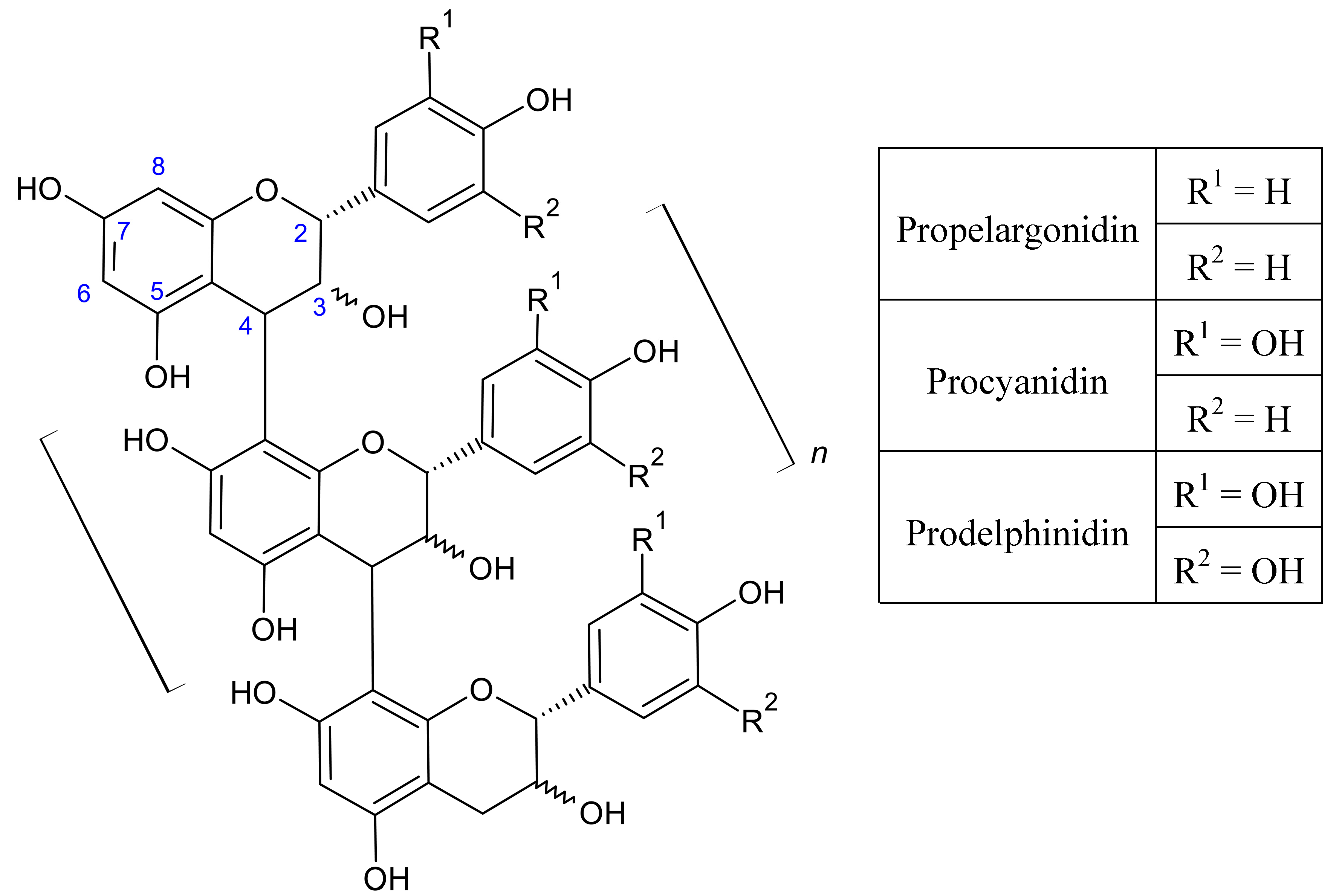{kind=link}
{kind=link}
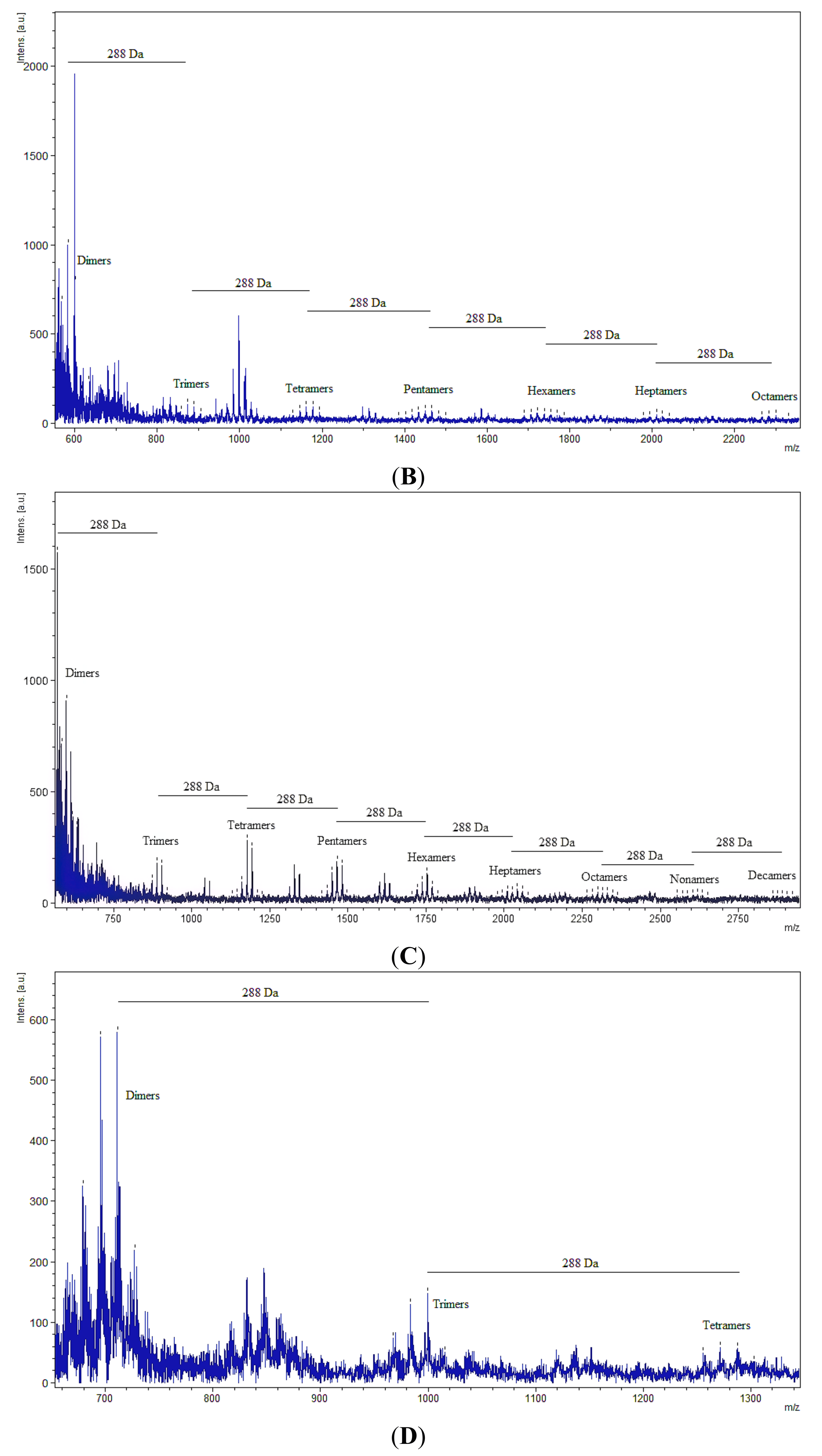{kind=link}
{kind=link}
{kind=link}
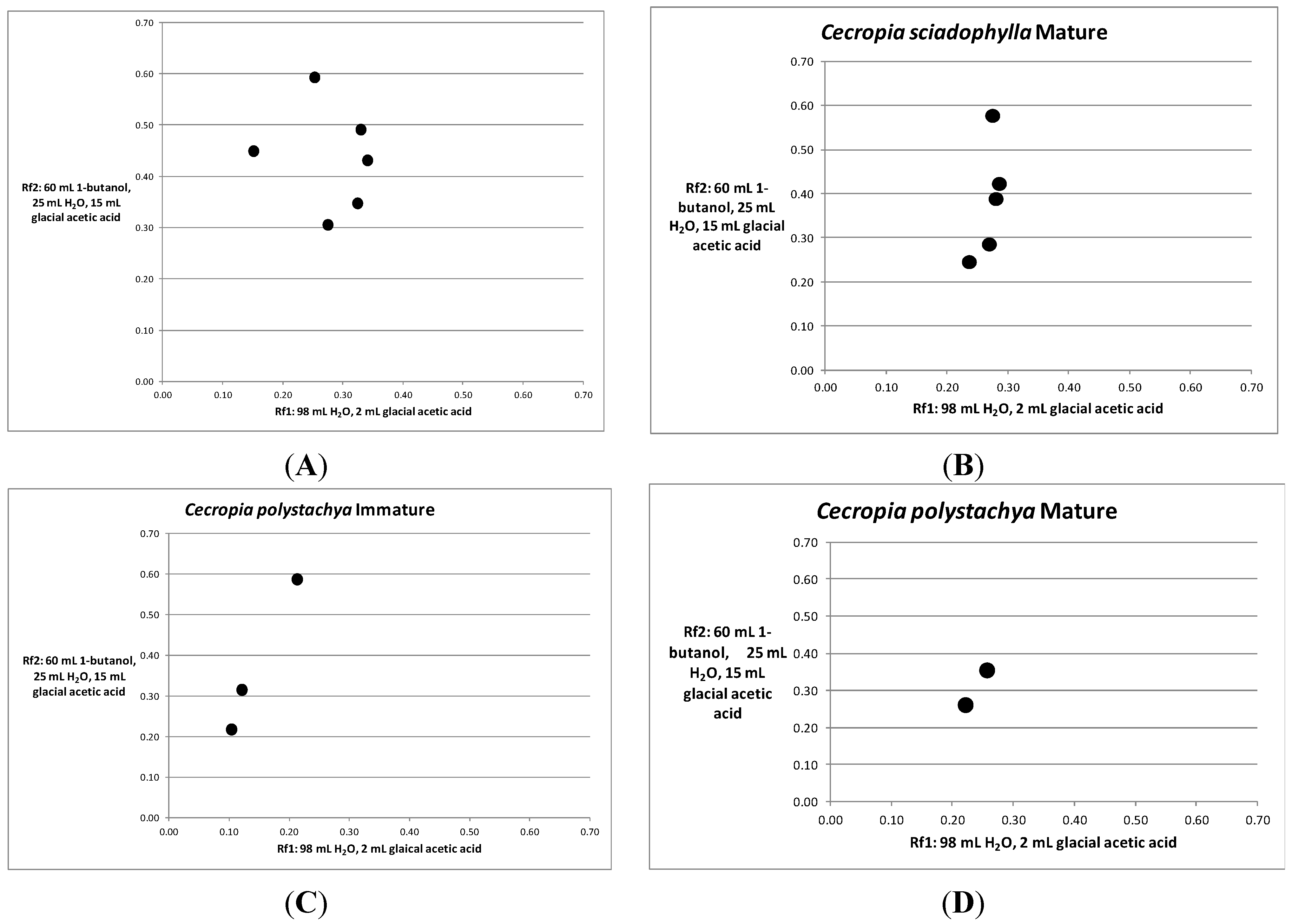{kind=link}
| Degrees of Polymerization | Number of Procyanidin Units (a) | Number of Prodelphinidin Units (b) | Number of Propelargonidin Units (c) | Calculated [M + Na]+ | Observed [M + Na]+ |
|---|---|---|---|---|---|
| Dimer | 0 | 0 | 2 | 569 | 569 |
| 1 | 0 | 1 | 585 | 585 | |
| 2 | 0 | 0 | 601 | 601 | |
| Trimer | 1 | 0 | 2 | 857 | 857 |
| 2 | 0 | 1 | 873 | 873 | |
| 3 | 0 | 0 | 889 | 889 | |
| 2 | 1 | 0 | 905 | 905 | |
| Tetramer | 1 | 0 | 3 | 1129 | 1129 |
| 2 | 0 | 2 | 1145 | 1145 | |
| 3 | 0 | 1 | 1161 | 1161 | |
| 4 | 0 | 0 | 1177 | 1177 | |
| 3 | 1 | 0 | 1193 | 1193 | |
| Pentamer | 1 | 0 | 4 | 1401 | 1402 |
| 2 | 0 | 3 | 1417 | 1418 | |
| 3 | 0 | 2 | 1433 | 1433 | |
| 4 | 0 | 1 | 1449 | 1449 | |
| 5 | 0 | 0 | 1465 | 1465 | |
| 4 | 1 | 0 | 1481 | 1482 | |
| Hexamer | 1 | 0 | 5 | 1673 | 1674 |
| 2 | 0 | 4 | 1689 | 1691 | |
| 3 | 0 | 3 | 1705 | 1706 | |
| 4 | 0 | 2 | 1721 | 1722 | |
| 5 | 0 | 1 | 1737 | 1738 | |
| 6 | 0 | 0 | 1753 | 1754 | |
| 5 | 1 | 0 | 1769 | 1770 | |
| Heptamer | 1 | 0 | 6 | 1945 | 1947 |
| 2 | 0 | 5 | 1961 | 1962 | |
| 3 | 0 | 4 | 1977 | 1978 | |
| 4 | 0 | 3 | 1993 | 1994 | |
| 5 | 0 | 2 | 2009 | 2010 | |
| 6 | 0 | 1 | 2025 | 2026 | |
| 7 | 0 | 0 | 2041 | 2042 | |
| 6 | 1 | 0 | 2057 | 2059 | |
| Octamer | 2 | 0 | 6 | 2233 | 2233 |
| 3 | 0 | 5 | 2249 | 2250 | |
| 4 | 0 | 4 | 2265 | 2267 | |
| 5 | 0 | 3 | 2281 | 2281 | |
| 6 | 0 | 2 | 2297 | 2296 | |
| 7 | 0 | 1 | 2313 | 2313 | |
| 8 | 0 | 0 | 2329 | 2329 | |
| 7 | 1 | 0 | 2345 | 2347 |


3. Experimental
3.1. Sample Collection
3.2. Proanthocyanidin Extraction
3.3. MADLI-TOF Mass Spectrometry
3.4. Thiolysis and Two Dimensional TLC
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brillouet, J.; Romieu, C.; Schoefs, B.; Solymosi, K.; Cheynier, V.; Fulerand, H.; Verdeil, J.; Gonejero, G. The tannosome is an organelle forming condensed tannins in the chlorophyllous organs of Tracheophyta. Ann. Bot. 2013, 112, 1003–1014. [Google Scholar] [CrossRef]
- Appel, H.M. The Chewing Herbivore Gut Lumen: Physicochemical Conditions and Their Impact on Plant Nutrients, Allelochemicals, and Insect Pathogens. In Insect-Plant Interactions; Bernays, E.A., Ed.; CRC Press: Boca Raton, FL, USA, 1994; pp. 209–223. [Google Scholar]
- Baxter, N.J.; Lilley, T.H.; Haslam, E.; Williamson, M.P. Multiple interactions between polyphenols and a salivary proline-rich protein repeat result in complexation and precipitation. Biochemistry 1997, 36, 5566–5577. [Google Scholar]
- Felton, G.W.; Duffey, S.S. Reassessment of the role of gut alkalinity and detergency in insect herbivory. J. Chem. Ecol. 1991, 17, 1821–1836. [Google Scholar] [CrossRef]
- Lee, K. Glutathione S-transferase activities in phytophagous insects: induction and inhibition by plant phytotoxins and phenols. Insect Biochem. 1991, 21, 353–361. [Google Scholar] [CrossRef]
- Hagerman, A.E. The Tannin Handbook. Available online: http://www.users.muohio.edu/hagermae/ (accessed on 1 September 2013).
- Porter, L.J. Tannins. In Methods in Plant Biochemistry; Herborne, J.B., Ed.; Academic Press Inc.: San Diego, CA, USA, 1989; Volume 1, pp. 389–419. [Google Scholar]
- Ayres, M.P.; Clausen, T.P.; MacLean, S.F.; Redman, A.M.; Reichardt, P.B. Diversity of structure and antiherbivore activity in condensed tannins. Ecology 1997, 78, 1696–1712. [Google Scholar] [CrossRef]
- Plumb, G.W.; de Pascual-Teresa, S.; Santos-Buelga, C.; Cheynier, V.; Williamson, G. Antioxidant properties of catechins and proanthocyanidins: Effect of polymerization, galloylation and glycosylation. Free Radic. Res. 1998, 19, 351–358. [Google Scholar]
- Feeny, P.P. Effect of oak leaf tannins on larval growth of the winter moth Operophtera brumata. J. Insect Physiol. 1968, 14, 805–817. [Google Scholar]
- Feeny, P.P. Inhibitory effect of oak leaf tannins on the hydrolysis of proteins by trypsin. Phytochemistry 1969, 8, 2119–2126. [Google Scholar] [CrossRef]
- Feeny, P.P. Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter moth caterpillars. Ecology 1970, 51, 565–581. [Google Scholar] [CrossRef]
- Forkner, R.E.; Marquis, R.J.; Lill, J.T. Feeny revisited: condensed tannins as anti-herbivore defenses in leaf-chewing herbivore communities of Quercus. Ecol. Entomol. 2004, 29, 174–187. [Google Scholar] [CrossRef]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomao, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342. [Google Scholar] [CrossRef]
- Berg, C.C.; Rosselli, P.F. Cecropia; New York Botanical Garden Press: Bronx, NY, USA, 2005. [Google Scholar]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar]
- Behrens, A.; Maie, N.; Knicker, H.; Kogel-Knabner, I. MALDI-TOF mass spectrometry and PSD fragmentation as means for the analysis of condensed tannins in plant leaves and needles. Phytochemistry 2003, 62, 1159–1170. [Google Scholar] [CrossRef]
- Foo, L.Y.; Lu, Y.; Howell, A.B.; Vorsa, N. The structure of cranberry proanthocyanidins which inhibit adherence of uropathogenic P-fimbriated Escherichia coli in vitro. Phytochemistry 2000, 54, 173–181. [Google Scholar] [CrossRef]
- Zhang, L.L.; Lin, Y.M. HPLC, NMR and MALDI-TOF MS analysis of condensed tannins from Lithocarpus glaber leaves with potent free radical scavenging activity. Molecules 2008, 13, 2986–2997. [Google Scholar] [CrossRef]
- Latterman, T.A.; Mead, J.E.; DuVall, M.A.; Bunting, C.C.; Bevington, J.M. Differences in anti-herbivore defenses in non-myrmecophyte and myrmecophyte Cecropia trees. Biotropica 2014, in press. [Google Scholar]
- Coley, P.D. Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ecol. Monogr. 1983, 53, 209–233. [Google Scholar] [CrossRef]
- Eck, G.; Fiala, B.; Linsenmair, K.E.; Hashim, R.B.; Proksch, P. Trade-off between chemical and biotic antiherbivore defense in the south east Asian plant genus Macaranga. J. Chem. Ecol. 2001, 27, 1979–1996. [Google Scholar] [CrossRef]
- Koptur, S. Alternative defenses against herbivores in Inga (Fabaceae: Mimosoidae) over an elevational gradient. Ecology 1985, 66, 1639–1650. [Google Scholar] [CrossRef]
- Rehr, S.S.; Feeny, P.P.; Janzen, D.H. Chemical defense in Central American non-ant-acacias. J. Anim. Ecol. 1973, 42, 405–416. [Google Scholar] [CrossRef]
- Jones, W.T.; Broadhurst, R.B.; Lyttleton, J.W. The condensed tannins of pasture legume species. Phytochemistry 1976, 15, 1407–1409. [Google Scholar] [CrossRef]
- Lin, Y.M.; Liu, J.W.; Xiang, P.; Lin, P.; Ye, G.F.; da Sternberg, L.S.L. Tannin dynamics of propagules and leaves of Kandelia candel and Bruguiera gymnorrhiza in the Jiulong River Estuary, Fujian, China. Biogeochemistry 2006, 78, 343–359. [Google Scholar] [CrossRef]
- Preston, C.M. Tannins of Salal (Gaultheria Shallon Pursh): A Contributing Factor to Seedling “Growth Check” on Northern Vancouver Island? In Plant Polyphenols 2: Chemistry, Biology, Pharmacology, Ecology; Gross, G.G., Hemingway, R.W., Yoshida, T., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 825–841. [Google Scholar]
- Xiang, P.; Lin, Y.; Lin, P.; Xiang, C. Effects of adduct ions on matrix-assisted laser desorption/ionization time of flight mass spectrometry of condensed tannins: A prerequisite knowledge. Chin. J. Anal. Chem. 2006, 34, 1019–1022. [Google Scholar] [CrossRef]
- Torres, J.L.; Lozano, C. Chromatographic characterization of proanthocyanidins after thiolysis with cysteamine. Chromatographica 2001, 54, 523–526. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Quantification of Tannins in Tree and Shrub Foliage; Springer: Heidelberg, The Netherlands, 2003; pp. 89–93. [Google Scholar]
- Sample Availability: Leaf samples are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Van Huynh, A.; Bevington, J.M. MALDI-TOF MS Analysis of Proanthocyanidins in Two Lowland Tropical Forest Species of Cecropia: A First Look at Their Chemical Structures. Molecules 2014, 19, 14484-14495. https://doi.org/10.3390/molecules190914484
Van Huynh A, Bevington JM. MALDI-TOF MS Analysis of Proanthocyanidins in Two Lowland Tropical Forest Species of Cecropia: A First Look at Their Chemical Structures. Molecules. 2014; 19(9):14484-14495. https://doi.org/10.3390/molecules190914484
Chicago/Turabian StyleVan Huynh, Alex, and John M. Bevington. 2014. "MALDI-TOF MS Analysis of Proanthocyanidins in Two Lowland Tropical Forest Species of Cecropia: A First Look at Their Chemical Structures" Molecules 19, no. 9: 14484-14495. https://doi.org/10.3390/molecules190914484
APA StyleVan Huynh, A., & Bevington, J. M. (2014). MALDI-TOF MS Analysis of Proanthocyanidins in Two Lowland Tropical Forest Species of Cecropia: A First Look at Their Chemical Structures. Molecules, 19(9), 14484-14495. https://doi.org/10.3390/molecules190914484