
Potential Grape-Derived Contributions to Volatile Ester Concentrations in Wine


 and
and
Abstract
:1. Introduction
2. Results and Discussion
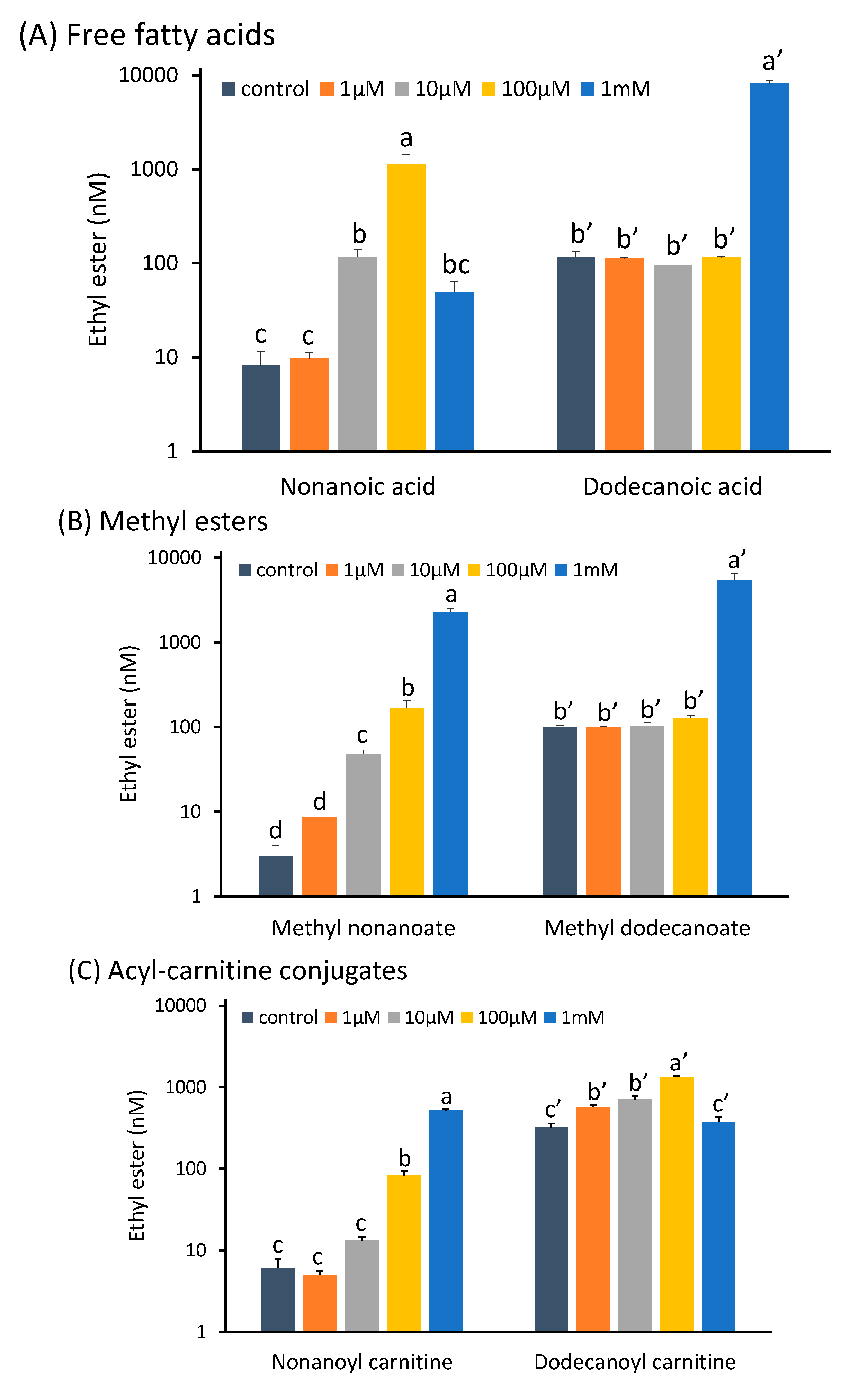
2.1. Potential Medium Chain Fatty Acid (MCFA)-Derived Precursors of Ethyl Esters
2.1.1. Free Fatty Acids as Direct Precursors

2.1.2. Methyl Esters of MCFAs as Transesterification Substrates
2.1.3. Acyl-Carnitine Conjugates, Putative Transport Forms of MCFAs
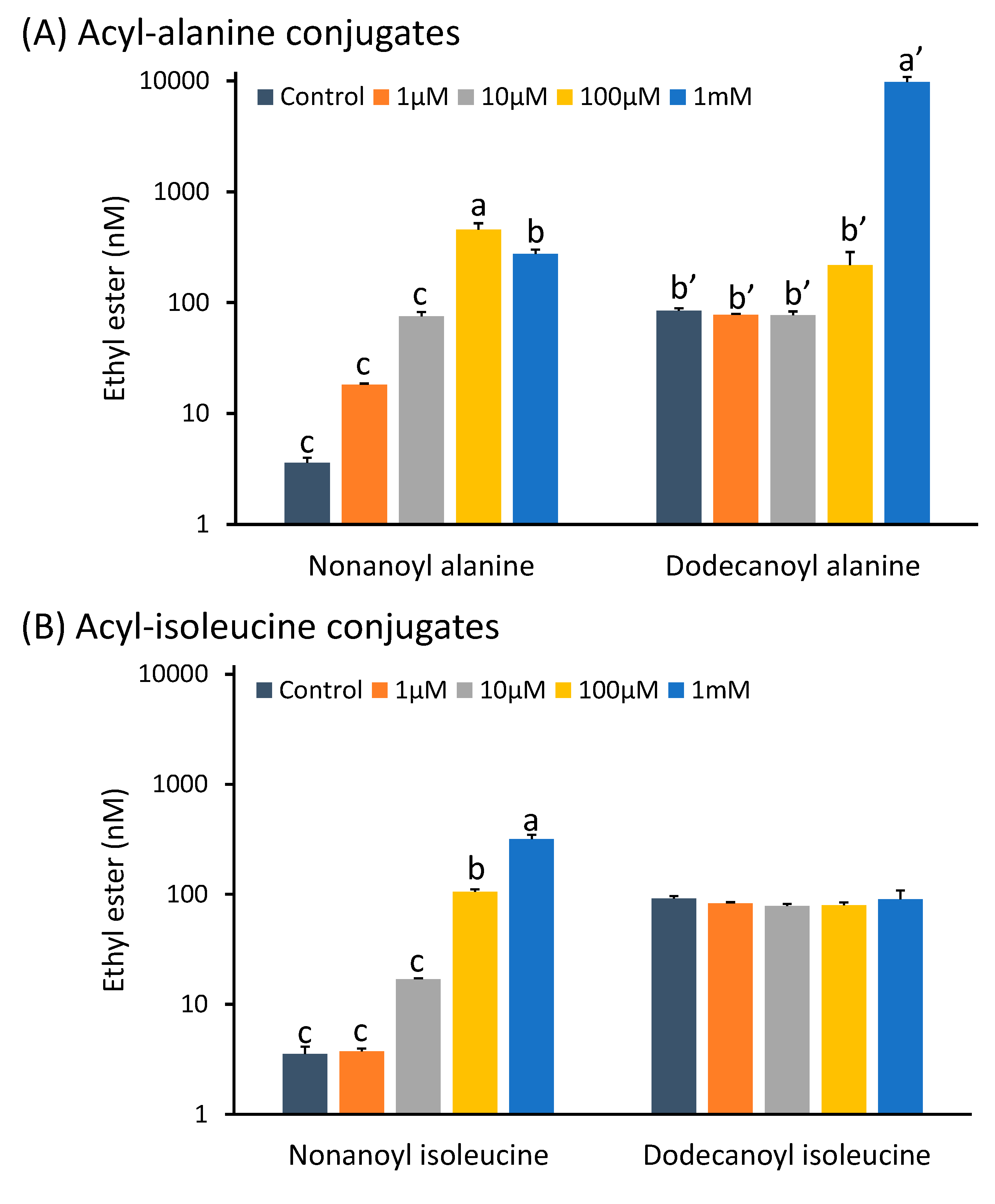
2.1.4. Acyl-Amino Acid Conjugates Potentially Exploiting Permeases
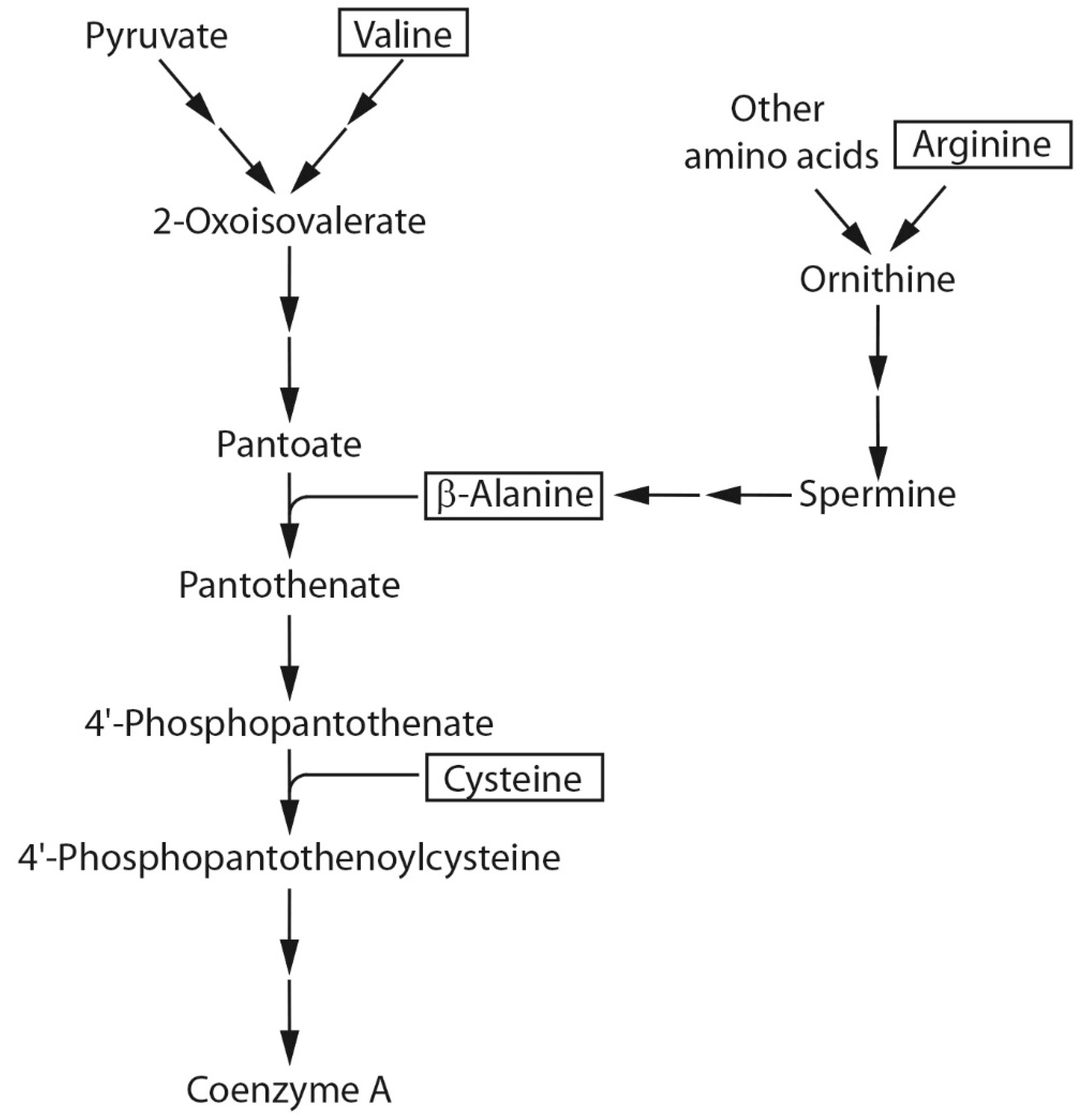
2.2. Amino Acids Involved in CoA Biosynthesis


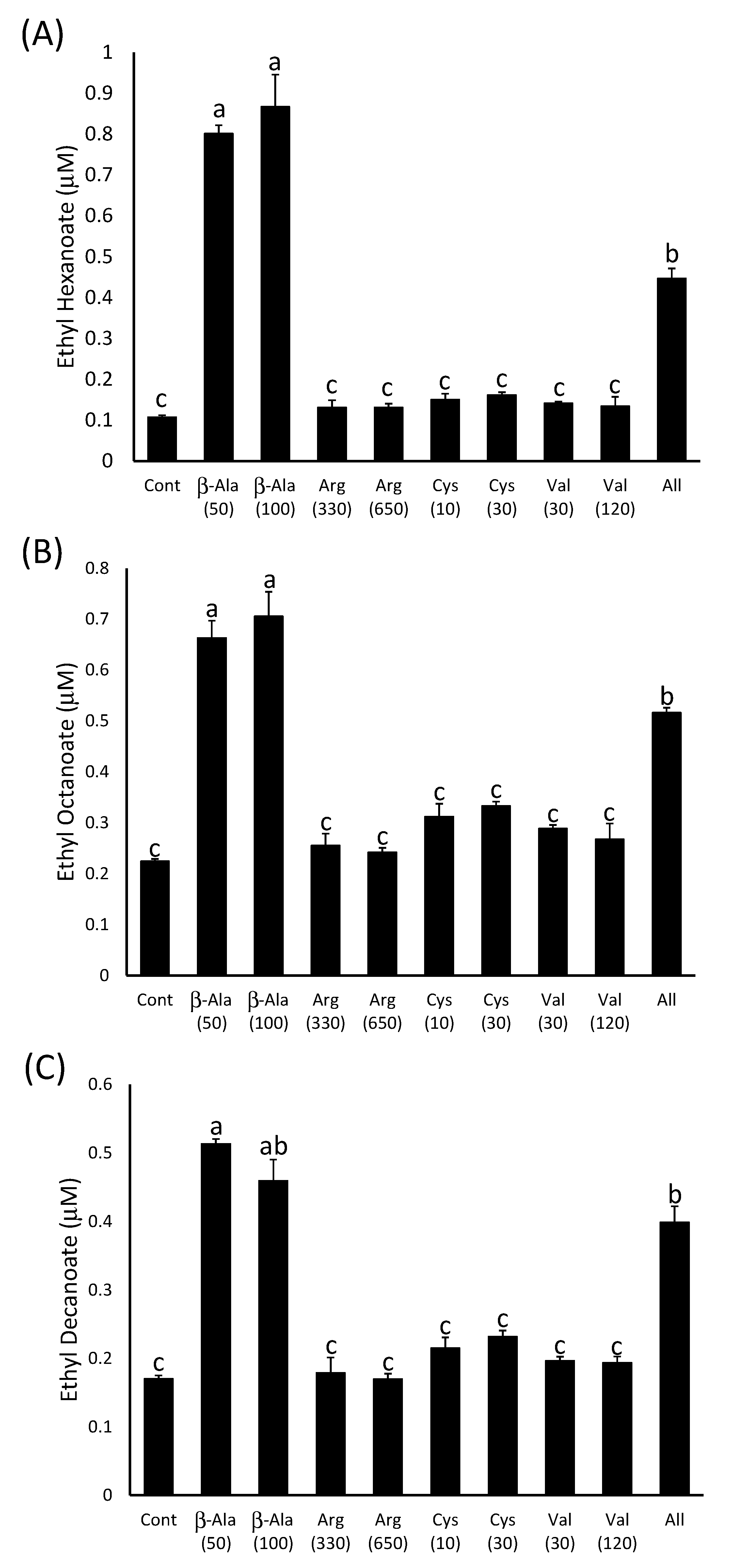
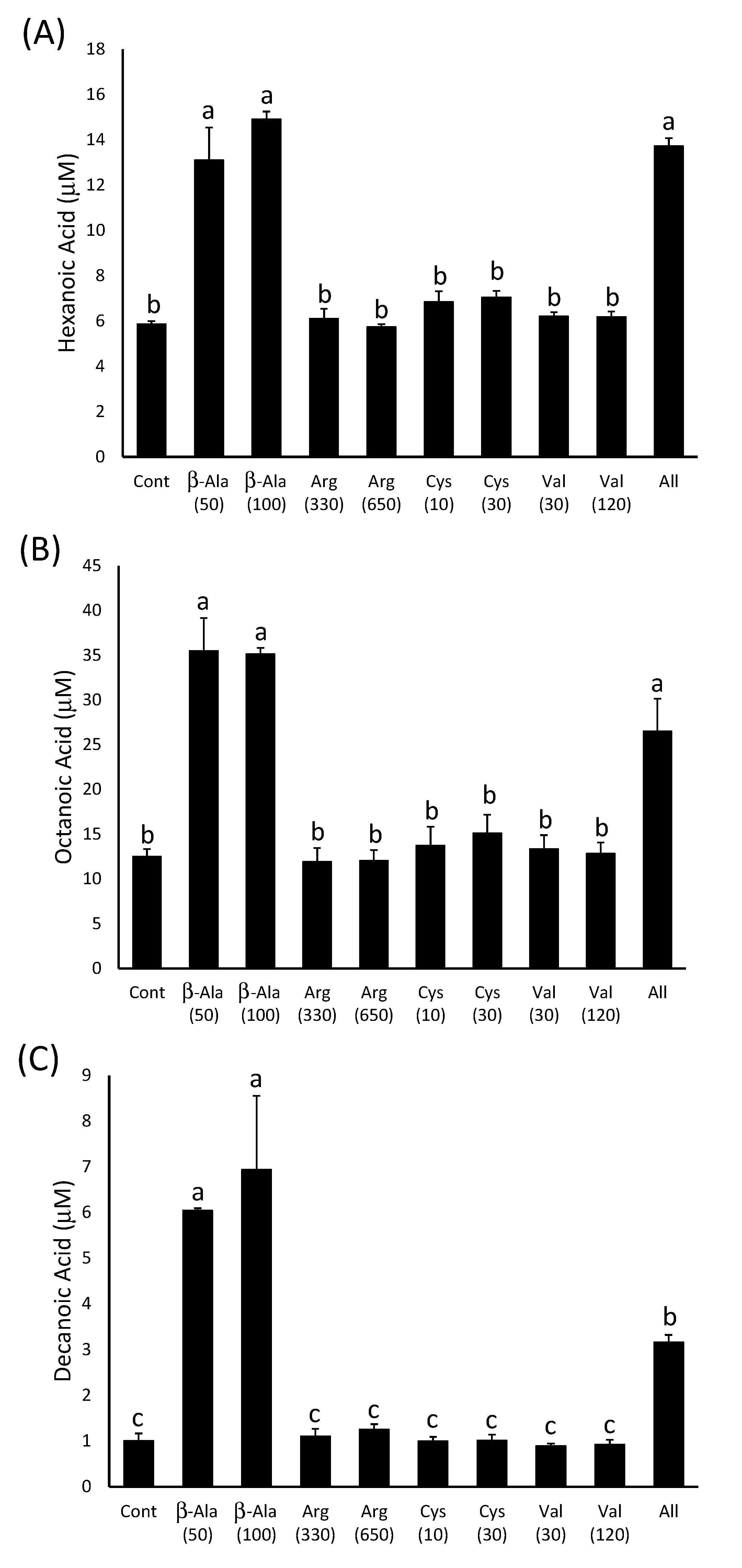
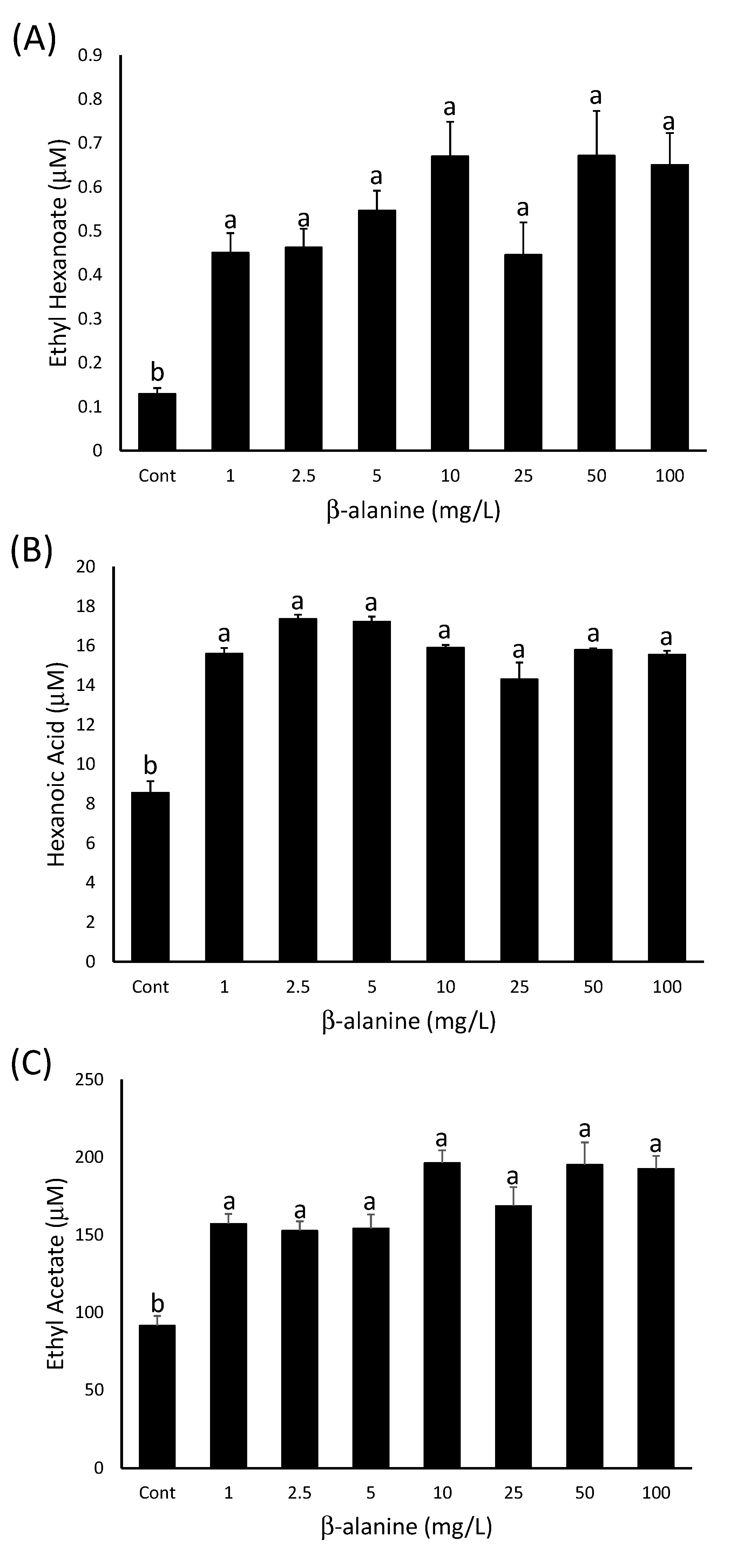
2.2.1. Stimulation of Ester Production by β-Alanine.



2.2.2. β-Alanine Concentrations in Grapes are Sufficient to Stimulate Ester Production
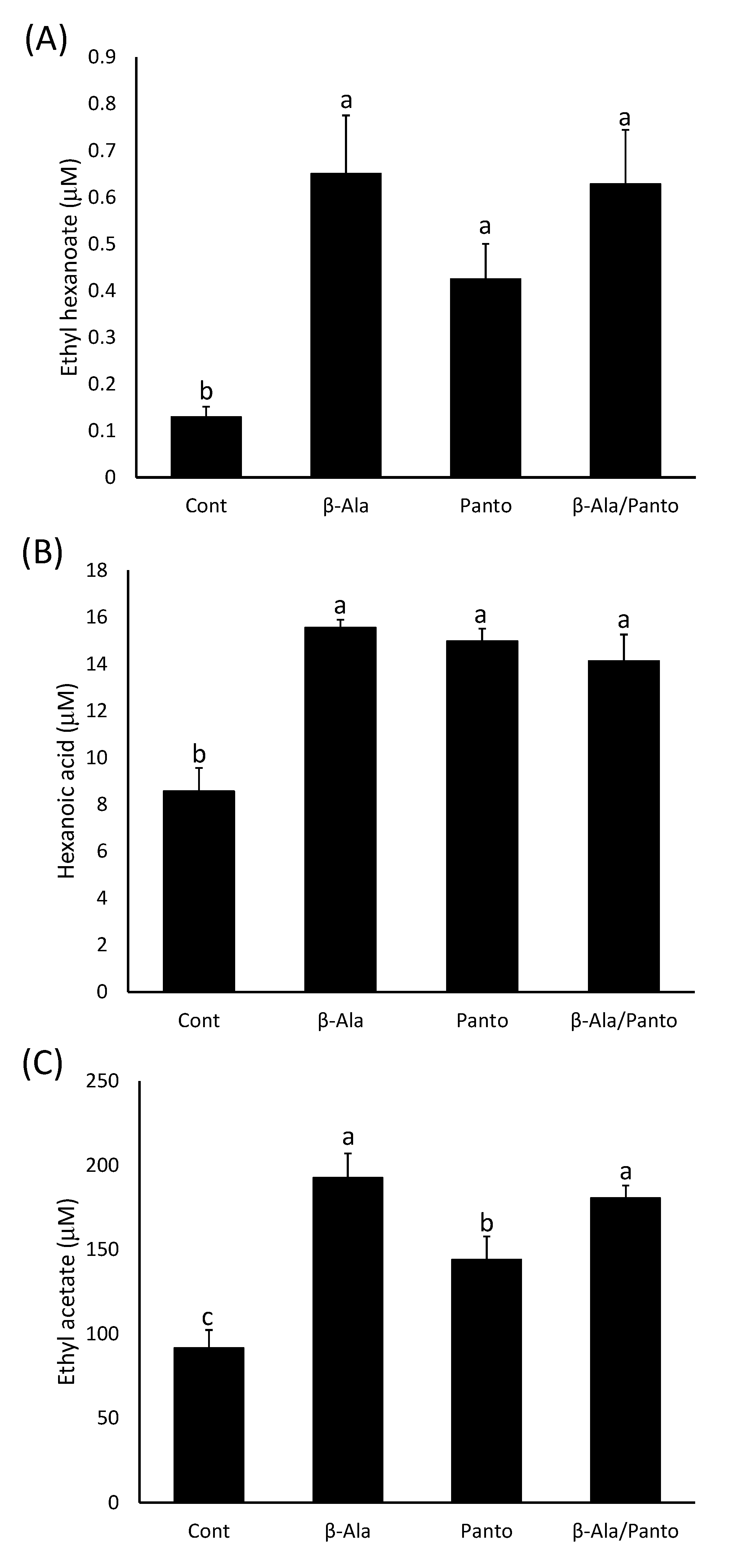
2.2.3. β-Alanine or Pantothenate Supplementation Have a Similar Effect on Ester Production


3. Experimental Section
3.1. Material
3.1.1. Chemicals
3.1.2. Nuclear Magnetic Resonance (NMR)
3.1.3. High-Resolution Mass Spectrometry (HR-MS)
3.1.4. HPLC-MS Instrumentation
3.1.5. GC-MS Instrumentation
3.1.6. Melting Points
3.2. Synthesis of Compounds
3.2.1. d5-Ethyl Nonanoate
3.2.2. d5-Ethyl Dodecanoate
3.2.3. (2R/S)-3-Carboxy-2-(nonanoyloxy)-N,N,N-trimethylpropan-1-aminium (N-Nonanoyl carnitine)
3.2.4. (2R/S)-3-Carboxy-2-(dodecanoyloxy)-N,N,N-trimethylpropan-1-aminium (N-Dodecanoyl carnitine)
3.2.5. Methyl 2-(Nonanoylamino)propanoate (N-Nonanoyl Alanine Methyl Ester)
3.2.6. 2-(Nonanoylamino)propanoic acid (N-nonanoyl alanine)
3.2.7. Methyl 2-(Dodecanoylamino)propanoate (N-dodecanoyl alanine methyl ester)
3.2.8. 2-(Dodecanoylamino)propanoic acid (N-dodecanoyl alanine)
3.2.9. Methyl 2-(Nonanoylamino)-3-methylpentanoate (N-nonanoyl isoleucine methyl ester)
3.2.10. 2-(Nonanoylamino)-3-methylpentanoic acid (N-nonanoyl isoleucine)
3.2.11. Methyl 2-(dodecanoylamino)-3-methylpentanoate (N-dodecanoyl isoleucine methyl ester)
3.2.12. 2-(Dodecanoylamino)-3-methylpentanoic acid (N-dodecanoyl isoleucine)
3.3. Yeast
3.4. Fermentation Conditions
3.4.1. Trials Involving Fatty Acids and Precursors
3.4.2. Trials Involving Amino Acids and Pantothenate
3.5. Quantitative GC-MS Analysis of Esters and Acids
3.6. Determination of β-Alanine Content of Grape Juice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte (Internal Standard) | SIM Ions (Quantifier in Bold) |
|---|---|
| Ethyl acetate | 43, 61, 70 |
| Ethyl hexanoate | 43, 60, 88 |
| Ethyl octanoate | 88, 101, 127 |
| Ethyl nonanoate | 73, 88, 101, 141, 186 |
| Ethyl decanoate | 73, 88, 101, 157 |
| Ethyl dodecanoate | 43, 88, 101, 157, 183 |
| Isoamyl acetate | 43, 55, 70, 87 |
| Phenyl ethyl acetate | 39, 43, 65, 91, 104 |
| Hexanoic acid | 41, 60, 73, 87 |
| Octanoic acid | 43, 55, 60, 73, 101 |
| Decanoic acid | 57, 60, 73, 101, 129 |
| (d3-Hexyl Acetate) | 46, 59, 84 |
| (d5-Ethyl Nonanoate) | 41, 60, 93 |
| (d11-Hexanoic Acid) | 63, 71 |
3.7. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ebeler, S.E.; Thorngate, J.H. Wine chemistry and flavor: Looking into the crystal glass. J. Agric. Food Chem. 2009, 57, 8098–8108. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.L.; Boss, P.K.; Solomon, P.S.; Trengove, R.D.; Heymann, H.; Ebeler, S.E. Origins of grape and wine aroma. Part 1. Chemical components and viticultural impacts. Am. J. Enol. Viticult. 2014, 65, 1–24. [Google Scholar] [CrossRef]
- Escudero, A.; Campo, E.; Farina, L.; Cacho, J.; Ferreira, V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agric. Food Chem. 2007, 55, 4501–4510. [Google Scholar]
- Dunlevy, J.D.; Kalua, C.M.; Keyzers, R.A.; Boss, P.K. The production of flavour and aroma compounds in grape berries. In Grapevine Molecular Physiology and Biotechnology, 2nd ed.; Roubelakis-Angelakis, K.A., Ed.; Springer Science+Business Media: Berlin, Germany, 2009; pp. 293–340. [Google Scholar]
- Ferreira, V.; Lopez, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Bakker, J.; Clarke, R.J. Wine Flavour Chemistry, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Bisson, L.F.; Karpel, J.E. Genetics of yeast impacting wine quality. In Annual Review of Food Science and Technology; Doyle, M.P., Klaenhammer, T.R., Eds.; Annual Reviews: Palo Alto, CA, USA, 2010; Volume 1, pp. 139–162. [Google Scholar]
- Verstrepen, K.J.; van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression levels of the yeast alcohol acetyltransferase genes ATF1, Lg-ATF1, and ATF2 control the formation of a broad range of volatile esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef] [PubMed]
- Peddie, H.A.B. Ester formation in brewery fermentations. J. Inst. Brew. 1990, 96, 327–331. [Google Scholar] [CrossRef]
- Olzhausen, J.; Schuebbe, S.; Schueller, H.-J. Genetic analysis of coenzyme A biosynthesis in the yeast Saccharomyces cerevisiae: Identification of a conditional mutation in the pantothenate kinase gene CAB1. Curr. Genet. 2009, 55, 163–173. [Google Scholar] [CrossRef] [PubMed]
- White, W.H.; Gunyuzlu, P.L.; Toyn, J.H. Saccharomyces cerevisiae is capable of de novo pantothenic acid biosynthesis involving a novel pathway of β-alanine production from spermine. J. Biol. Chem. 2001, 276, 10794–10800. [Google Scholar] [CrossRef] [PubMed]
- Saerens, S.M.G.; Delvaux, F.; Verstrepen, K.J.; van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Palomo, E.S.; Diaz-Maroto, M.C.; Vinas, M.A.G.; Soriano-Perez, A.; Perez-Coello, M.S. Aroma profile of wines from Albillo and Muscat grape varieties at different stages of ripening. Food Control. 2007, 18, 398–403. [Google Scholar] [CrossRef]
- Hjelmeland, A.K.; Ebeler, S.E. Glycosidically bound volatile aroma compounds in grapes and wine: A review. Am. J. Enol. Viticult. 2015, 66, 1–11. [Google Scholar] [CrossRef]
- Capone, D.L.; Sefton, M.A.; Jeffery, D.W. Analytical investigations of wine odorant 3-mercaptohexan-1-ol and its precursors. In Flavor Chemistry of Wine and Other Alcoholic Beverages; Qian, M.C., Shellhammer, T., Eds.; American Chemical Society: Washington, DC, USA, 2012; pp. 15–35. [Google Scholar]
- Keyzers, R.A.; Boss, P.K. Changes in the volatile compound production of fermentations made from musts with increasing grape content. J. Agric. Food Chem. 2010, 58, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.G.; Keyzers, R.A.; Kalua, C.M.; Maffei, S.M.; Nicholson, E.L.; Boss, P.K. Grape contribution to wine aroma: Production of hexyl acetate, octyl acetate, and benzyl acetate during yeast fermentation is dependent upon precursors in the must. J. Agric. Food Chem. 2012, 60, 2638–2646. [Google Scholar] [CrossRef] [PubMed]
- Bramley, R.G.V.; Ouzman, J.; Boss, P.K. Variation in vine vigour, grape yield and vineyard soils and topography as indicators of variation in the chemical composition of grapes, wine and wine sensory attributes. Aust. J. Grape Wine Res. 2011, 17, 217–229. [Google Scholar] [CrossRef]
- Robinson, A.L.; Boss, P.K.; Heymann, H.; Solomon, P.S.; Trengove, R.D. Influence of yeast strain, canopy management, and site on the volatile composition and sensory attributes of Cabernet Sauvignon wines from Western Australia. J. Agric. Food Chem. 2011, 59, 3273–3284. [Google Scholar] [CrossRef] [PubMed]
- Schreier, P.; Drawert, F.; Junker, A. Identification of volatile constituents from grapes. J. Agric. Food Chem. 1976, 24, 331–336. [Google Scholar] [CrossRef]
- van Roermund, C.W.T.; Hettema, E.H.; van den Berg, M.; Tabak, H.F.; Wanders, R.J.A. Molecular characterization of carnitine-dependent transport of acetyl-CoA from peroxisomes to mitochondria in Saccharomyces cerevisiae and identification of a plasma membrane carnitine transporter, Agp2p. EMBO J. 1999, 18, 5843–5852. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; O’Dell, D.K.; Yu, Y.W.; Monn, M.F.; Hughes, H.V.; Burstein, S.; Walker, J.M. Identification of endogenous acyl amino acids based on a targeted lipidomics approach. J. Lipid Res. 2010, 51, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Tortoriello, G.; Rhodes, B.P.; Takacs, S.M.; Stuart, J.M.; Basnet, A.; Raboune, S.; Widlanski, T.S.; Doherty, P.; Harkany, T.; Bradshaw, H.B. Targeted lipidomics in Drosophila melanogaster identifies novel 2-monoacylglycerols and N-acyl amides. PLoS ONE 2013, 8, e67865. [Google Scholar] [CrossRef] [PubMed]
- Halitschke, R.; Schittko, U.; Pohnert, G.; Boland, W.; Baldwin, I.T. Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. III. Fatty acid-amino acid conjugates in herbivore oral secretions are necessary and sufficient for herbivore-specific plant responses. Plant. Physiol. 2001, 125, 711–717. [Google Scholar]
- Gallander, J.F.; Peng, A.C. Lipid and fatty-acid compositions of different grape types. Am. J. Enol. Vitic. 1980, 31, 24–27. [Google Scholar]
- Lopez, R.; Ezpeleta, E.; Sanchez, I.; Cacho, J.; Ferreira, V. Analysis of the aroma intensities of volatile compounds released from mild acid hydrolysates of odourless precursors extracted from Tempranillo and Ggrenache grapes using gas chromatography-olfactometry. Food Chem. 2004, 88, 95–103. [Google Scholar] [CrossRef]
- Hunkova, Z.; Fencl, Z. Toxic effects of fatty-acids on yeast cells: dependence of inhibitory effects on fatty-acid concentration. Biotechnol. Bioeng. 1977, 19, 1623–1641. [Google Scholar] [CrossRef] [PubMed]
- Neal, A.L.; Weinstock, J.O.; Lampen, J.O. Mechanisms of fatty acid toxicity for yeast. J. Bacteriol. 1965, 90, 126–131. [Google Scholar] [PubMed]
- Stevens, S.; Hofmeyr, J.H.S. Effects of ethanol, octanoic and decanoic acids on fermentation and the passive influx of protons through the plasma membrane of Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1993, 38, 656–663. [Google Scholar] [CrossRef]
- Malcorps, P.; Dufour, J.P. Short-chain and medium-chain aliphatic-ester synthesis in Saccharomyces cerevisiae. Eur. J. Biochem. 1992, 210, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Schreier, P. Flavor composition of wines—Review. Crit. Rev. Food Sci. Nutr. 1979, 12, 59–111. [Google Scholar] [CrossRef]
- Zou, Z.Y.; DiRusso, C.C.; Ctrnacta, V.; Black, P.N. Fatty acid transport in Saccharomyces cerevisiae—Directed mutagenesis of Fat1 distinguishes the biochemical activities associated with Fat1p. J. Biol. Chem. 2002, 277, 31062–31071. [Google Scholar] [CrossRef] [PubMed]
- Seline, K.G.; Johein, H. The determination of l-carnitine in several food samples. Food Chem. 2007, 105, 793–804. [Google Scholar] [CrossRef]
- Regenberg, B.; During-Olsen, L.; Kielland-Brandt, M.C.; Holmberg, S. Substrate specificity and gene expression of the amino-acid permeases in Saccharomyces cerevisiae. Curr. Genet. 1999, 36, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Ugliano, M.; Siebert, T.; Mercurio, M.; Capone, D.; Henschke, P.A. Volatile and color composition of young and model-aged Shiraz wines as affected by diammonium phosphate supplementation before alcoholic fermentation. J. Agric. Food Chem. 2008, 56, 9175–9182. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Nicolini, G.; Ramponi, M.; Larcher, R. Free amino acid composition of juices of 12 grape varieties grown in Trentino (Italy). Ital. J. Food Sci. 2001, 13, 189–199. [Google Scholar]
- Da Cruz, S.H.; Maffud Cilli, E.; Ernandes, J.R. Structural complexity of the nitrogen source and influence on yeast growth and fermentation. J. Inst. Brew. 2002, 108, 54–61. [Google Scholar] [CrossRef]
- Kluba, R.M.; Mattick, L.R.; Hackler, L.R. Changes in free and total amino-acid composition of several Vitis labruscana grape varieties during maturation. Am. J. Enol. Viticult. 1978, 29, 102–111. [Google Scholar]
- Stolz, J.; Sauer, N. The fenpropimorph resistance gene FEN2 from Saccharomyces cerevisiae encodes a plasma membrane H+-pantothenate symporter. J. Biol. Chem. 1999, 274, 18747–18752. [Google Scholar] [CrossRef] [PubMed]
- Hagen, K.M.; Keller, M.; Edwards, C.G. Survey of biotin, pantothenic acid, and assimilable nitrogen in winegrapes from the Pacific Northwest. Am. J. Enol. Viticult. 2008, 59, 432–436. [Google Scholar]
- Bohmer, T.; Bremer, J. Propionylcarnitine physiological variations in vivo. BBA-Lipid Lipid Metab. 1968, 152, 559–567. [Google Scholar] [CrossRef]
- Montgomery, J.A.; Mamer, O.A. Measurement of urinary free and acylcarnitines—quantitative acylcarnitine profiling in normal humans and several patients with metabolic errors. Anal. Biochem. 1989, 176, 85–95. [Google Scholar] [CrossRef]
- Staab, H.A. New methods of preparative organic chemistry IV. Syntheses using heterocyclic amides (azolides). Angew. Chem. Int. Ed. 1962, 1, 351–367. [Google Scholar] [CrossRef]
- Staab, H.A.; Luking, M.; Durr, F.H. Darstellung von imidazoliden sythese von amiden, hydraziden und hydroxamsauren nach der imidazolidmethode. Chem. Ber. 1962, 95, 1275–1283. [Google Scholar] [CrossRef]
- Couffin-Hoarau, A.C.; Motulsky, A.; Delmas, P.; Leroux, J.C. In situ-forming pharmaceutical organogels based on the self-assembly of l-alanine derivatives. Pharm. Res. 2004, 21, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Henschke, P.A.; Jiranek, V. Yeasts-Metabolism of nitrogen compounds. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic: Chur, Switzerland, 1993; pp. 77–164. [Google Scholar]
- McBryde, C.; Gardner, J.M.; de Barros Lopes, M.; Jiranek, V. Generation of novel wine yeast strains by adaptive evolution. Am. J. Enol. Viticult. 2006, 57, 423–430. [Google Scholar]
- Cohen, S.A.; Michaud, D.P. Synthesis of a fluorescent derizatizing reagent, 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate, and its application for the analysis of hydrolysate amino-acids via high-performance liquid-chromotography. Anal. Biochem. 1993, 211, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-P.; Yang, X.-Y.; Hegeman, A.D.; Gray, W.M.; Cohen, J.D. Microscale analysis of amino acids using gas chromatography-mass spectrometry after methyl chloroformate derivatization. J. Chromatogr. B 2010, 878, 2199–2208. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boss, P.K.; Pearce, A.D.; Zhao, Y.; Nicholson, E.L.; Dennis, E.G.; Jeffery, D.W. Potential Grape-Derived Contributions to Volatile Ester Concentrations in Wine. Molecules 2015, 20, 7845-7873. https://doi.org/10.3390/molecules20057845
Boss PK, Pearce AD, Zhao Y, Nicholson EL, Dennis EG, Jeffery DW. Potential Grape-Derived Contributions to Volatile Ester Concentrations in Wine. Molecules. 2015; 20(5):7845-7873. https://doi.org/10.3390/molecules20057845
Chicago/Turabian StyleBoss, Paul K., Anthony D. Pearce, Yanjia Zhao, Emily L. Nicholson, Eric G. Dennis, and David W. Jeffery. 2015. "Potential Grape-Derived Contributions to Volatile Ester Concentrations in Wine" Molecules 20, no. 5: 7845-7873. https://doi.org/10.3390/molecules20057845
APA StyleBoss, P. K., Pearce, A. D., Zhao, Y., Nicholson, E. L., Dennis, E. G., & Jeffery, D. W. (2015). Potential Grape-Derived Contributions to Volatile Ester Concentrations in Wine. Molecules, 20(5), 7845-7873. https://doi.org/10.3390/molecules20057845