Equol, a Clinically Important Metabolite, Inhibits the Development and Pathogenicity of Magnaporthe oryzae, the Causal Agent of Rice Blast Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Growth Conditions
2.2. Assays of Mycelia Growth, Conidial Generation, Germination, and Appressorial Formation
2.3. Pathogenicity Tests
3. Results
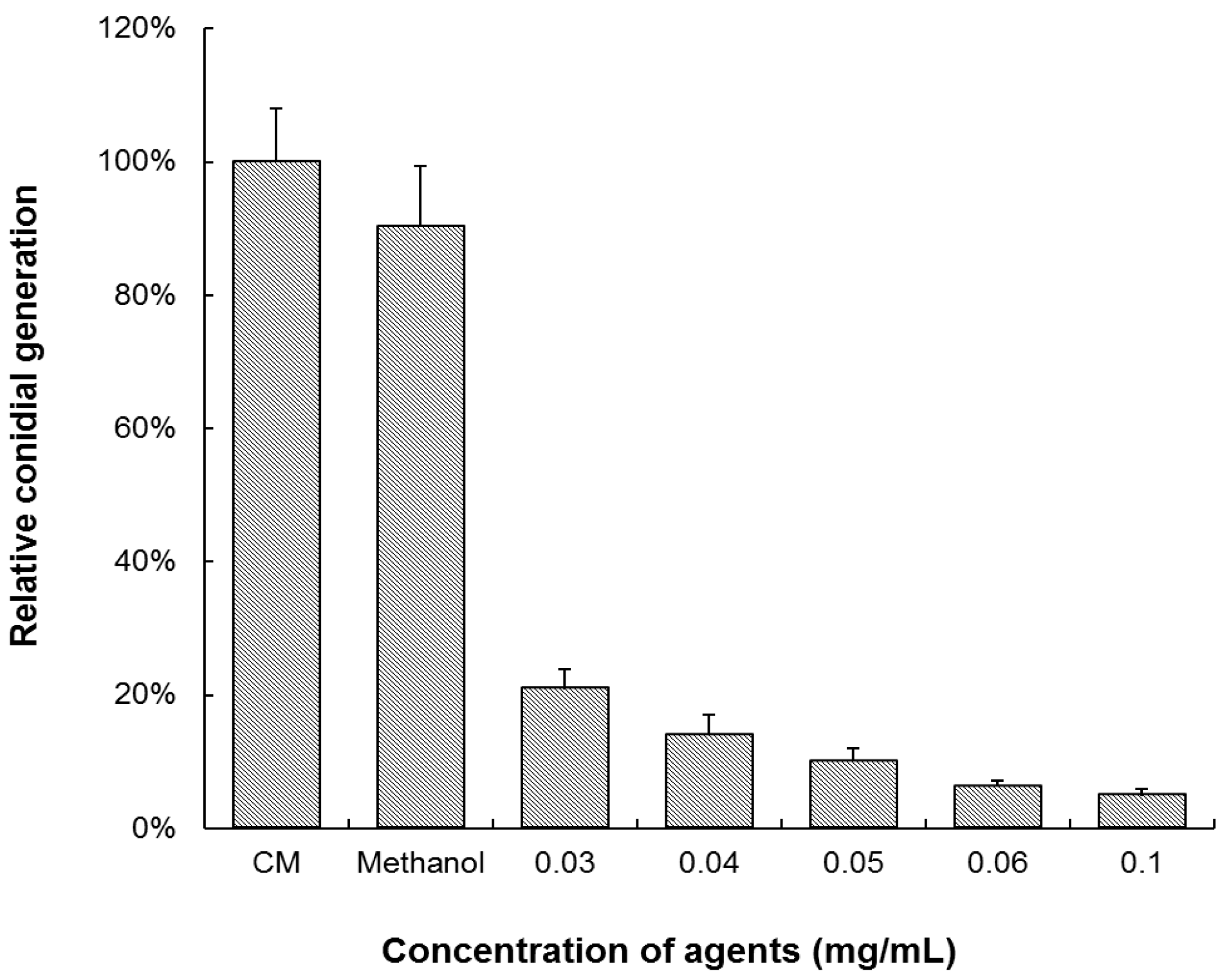
3.1. Equol Inhibits Mycelia Growth and Conidia Generation of M. oryzae
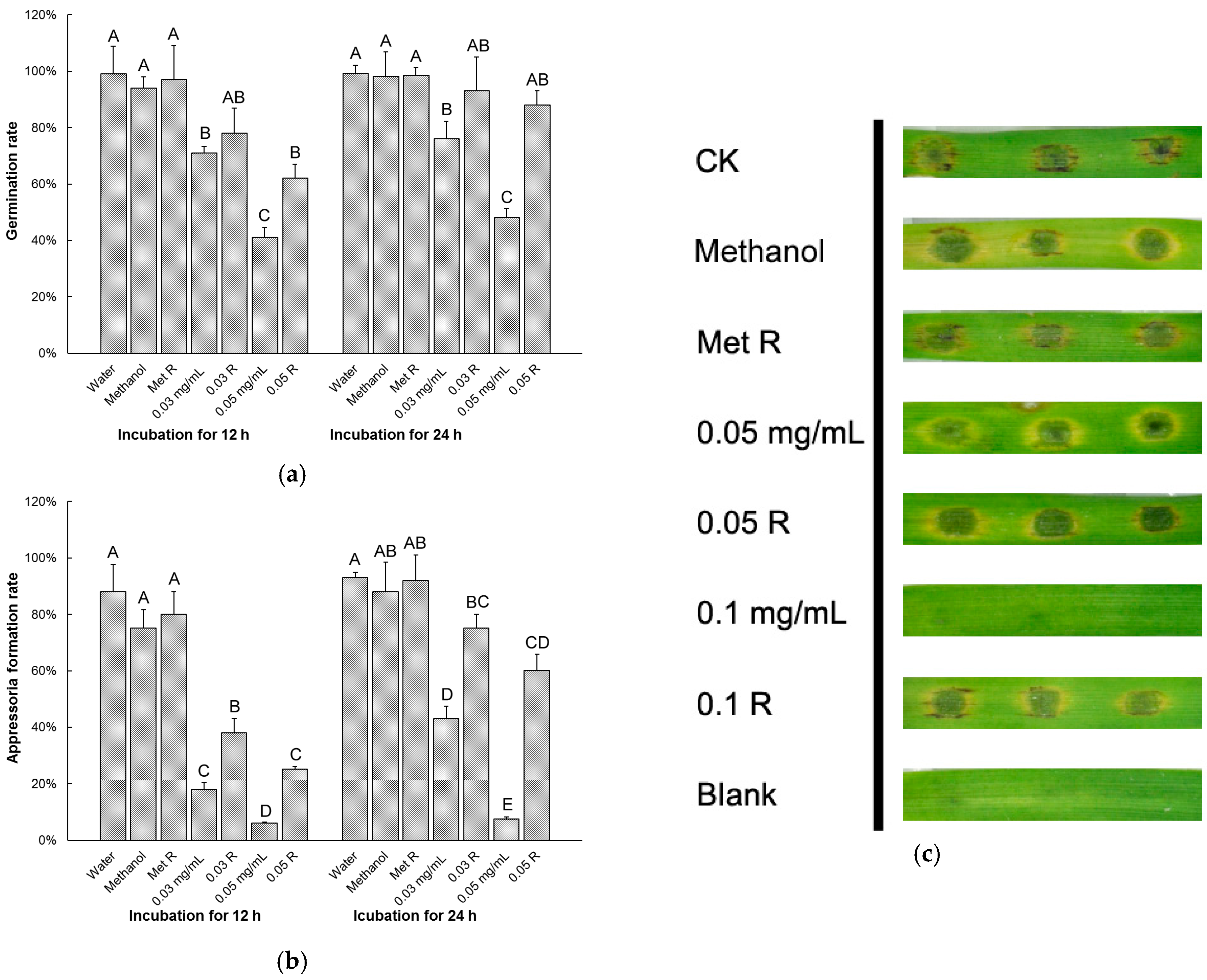
3.2. Equol Affects Conidial Germination and Appressorial Development
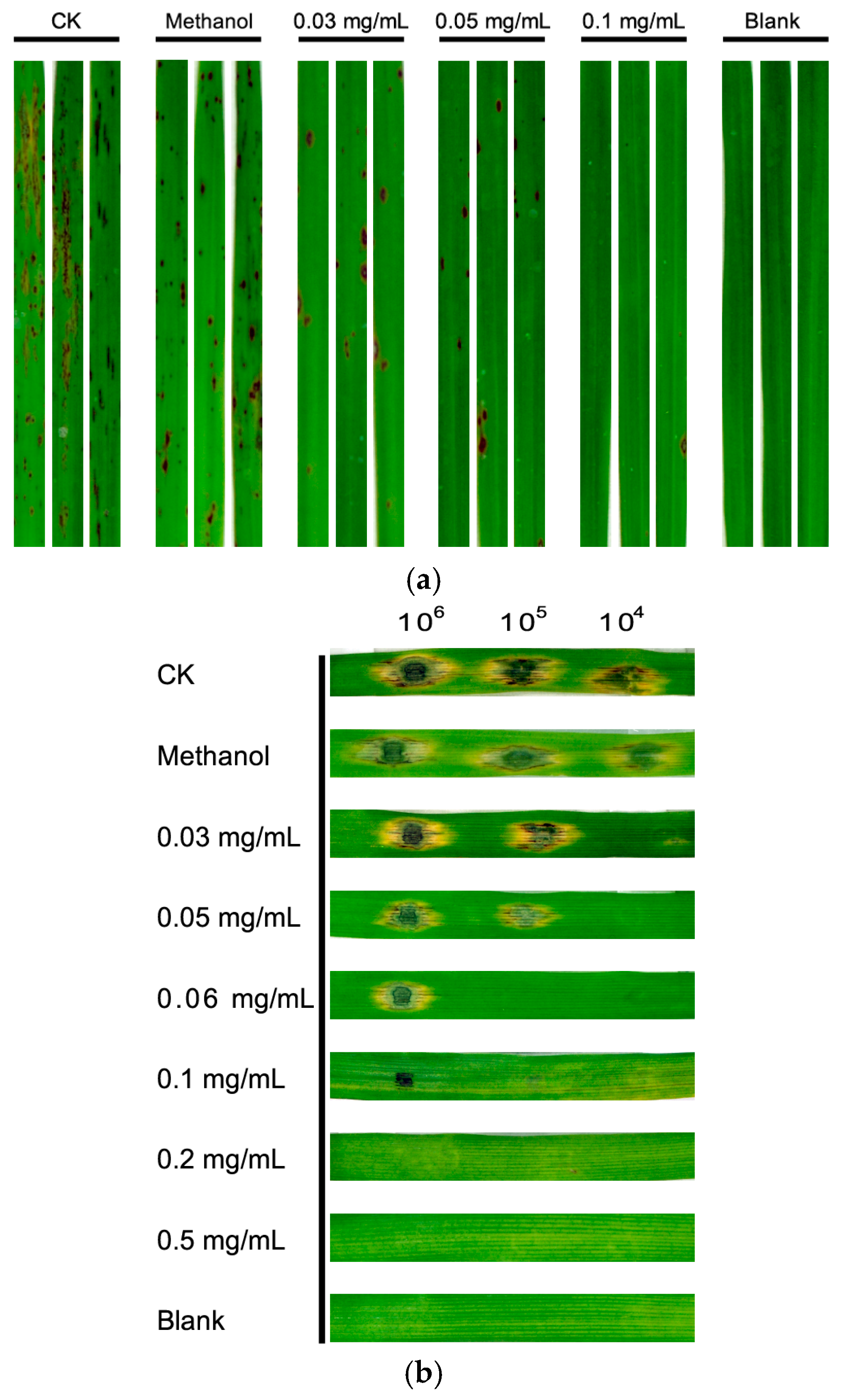
3.3. Equol Inhibits Pathogenicity of M. oryzae on Rice and Barley
3.4. Effects of Equol Are Restorable
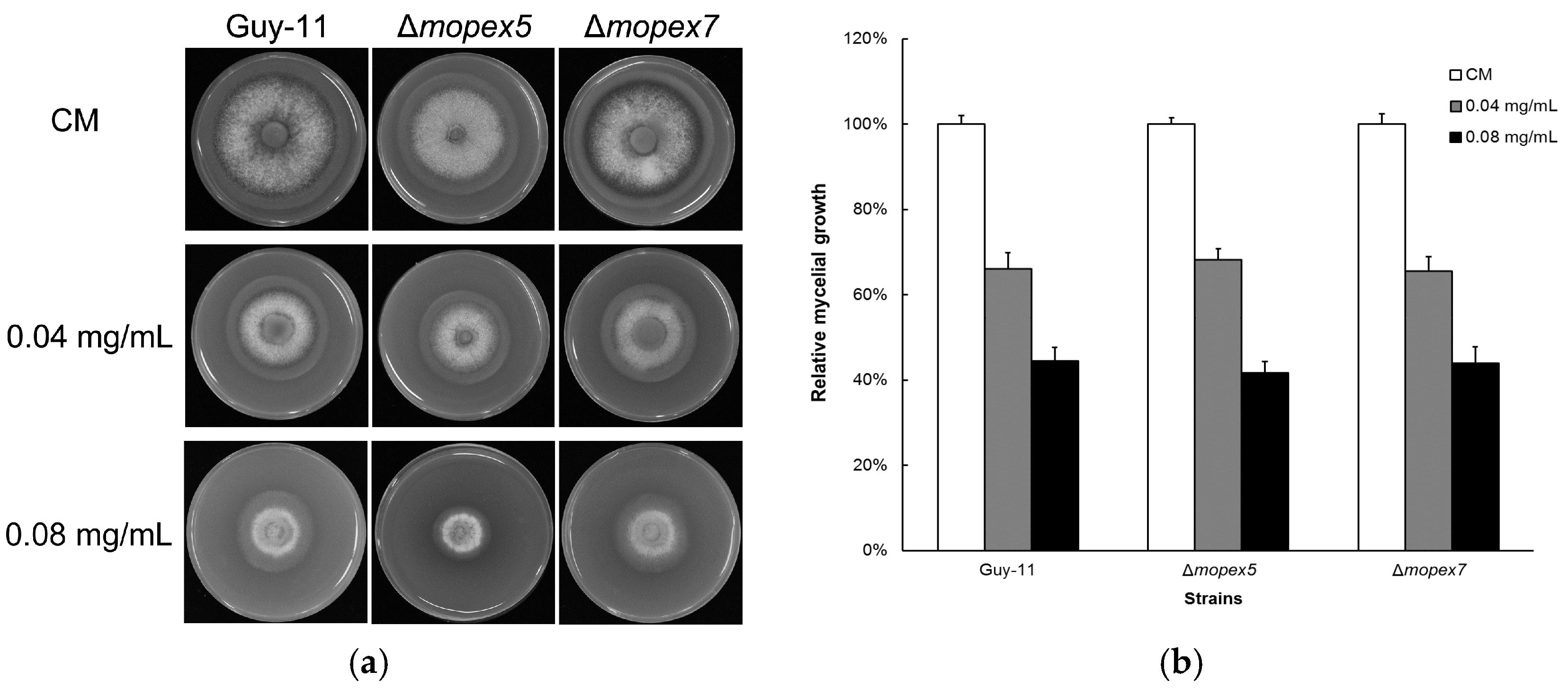
3.5. Antifungal Activity of Equol Is Not Related to Peroxisome
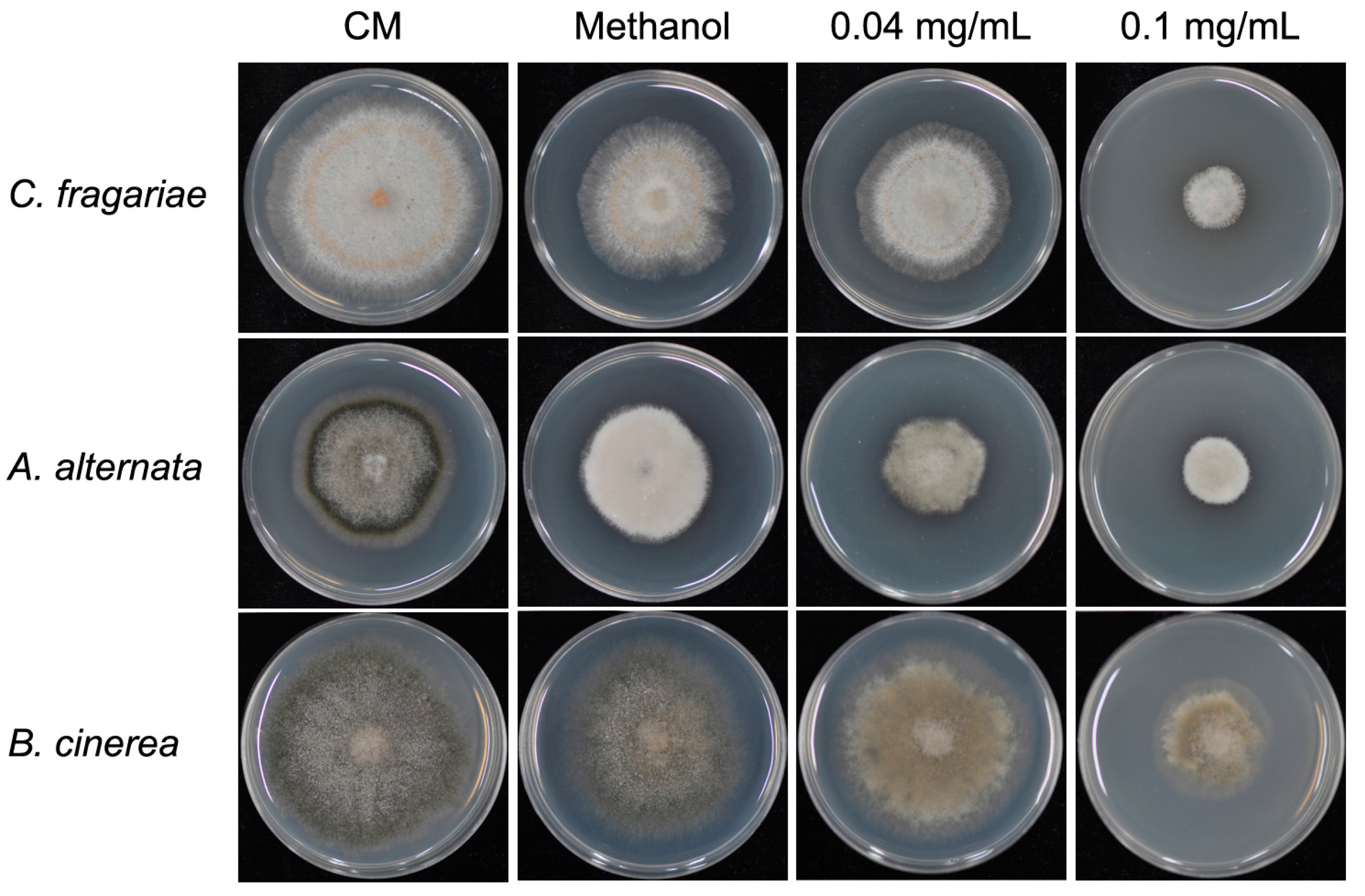
3.6. Antifungal Activity of Equol on Other Fungal Pathogens
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Marrian, G.F.; Haslewood, G.A. Equol, a new inactive phenol isolated from the ketohydroxyoestrin fraction of mares’ urine. Biochem. J. 1932, 26, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Clerici, C. Equol: History, chemistry, and formation. J. Nutr. 2010, 140, 1355S–1362S. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Hur, H.G.; Lee, J.H.; Kim, K.T.; Kim, S.I. Enantioselective synthesis of S-equol from dihydrodaidzein by a newly isolated anaerobic human intestinal bacterium. Appl. Environ. Microb. 2005, 71, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Lampe, J.W.; Karr, S.C.; Hutchins, A.M.; Slavin, J.L. Urinary equol excretion with a soy challenge: Influence of habitual diet. Exp. Biol. Med. 1998, 217, 335–339. [Google Scholar] [CrossRef]
- Setchell, K.D.; Brown, N.M.; Lydeking-Olsen, E. The clinical importance of the metabolite equol-a clue to the effectiveness of soy and its isoflavones. J. Nutr. 2002, 132, 3577–3584. [Google Scholar] [PubMed]
- Hedlund, T.E.; Johannes, W.U.; Miller, G.J. Soy isoflavonoid equol modulates the growth of benign and malignant prostatic epithelial cells in vitro. Prostate 2003, 54, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, H.; O’reilly, J.D.; Adlercreutz, H.; Mallet, A.I.; Bowey, E.A.; Rowland, I.R.; Sanders, T.A. Isoflavone phytoestrogens consumed in soy decrease F(2)-isoprostane concentrations and increase resistance of low-density lipoprotein to oxidation in humans. Am. J. Clin. Nutr. 2000, 72, 395–400. [Google Scholar] [PubMed]
- Lund, T.D.; Munson, D.J.; Haldy, M.E.; Setchell, K.D.; Lephart, E.D.; Handa, R.J. Equol is a novel anti-androgen that inhibits prostate growth and hormone feedback. Biol. Reprod. 2004, 70, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D. Soy isoflavones—Benefits and risks from nature’s selective estrogen receptor modulators (SERMs). J. Am. Coll. Nutr. 2001, 20, 354S–362S. [Google Scholar] [CrossRef] [PubMed]
- Yee, S.; Burdock, G.A.; Kurata, Y.; Enomoto, Y.; Narumi, K.; Hamada, S.; Itohc, T.; Shimomurac, Y.; Uenoc, T. Acute and subchronic toxicity and genotoxicity of SE5-OH, an equol-rich product produced by Lactococcus garvieae. Food Chem. Toxicol. 2008, 46, 2713–2720. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Valent, B. Breaking and entering: Host penetration by the fungal rice blast pathogen Magnaporthe grisea. Annu. Rev. Microbiol. 1996, 50, 491–512. [Google Scholar] [CrossRef] [PubMed]
- De Jong, J.C.; Mccormack, B.J.; Smirnoff, N.; Talbot, N.J. Glycerol generates turgor in rice blast. Nature 1997, 389, 244. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [PubMed]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 1993, 5, 1575–1590. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Z.; Wang, Y.; Li, L.; Chai, R.; Mao, X.; Jiang, H.; Qiu, H.; Du, X.; Lin, F.; et al. PTS1 peroxisomal import pathway plays shared and distinct roles to PTS2 pathway in development and pathogenicity of Magnaporthe oryzae. PLoS ONE 2013, 8, e55554. [Google Scholar] [CrossRef] [PubMed]
- Staats, M.; van Kan, J.A.L. Genome update of Botrytis cinerea strains B05. 10 and T4. Eukaryot. Cell 2012, 11, 1413–1414. [Google Scholar] [CrossRef] [PubMed]
- Dang, Z.; Lowik, C.W. The balance between concurrent activation of ERs and PPARs determines daidzein-induced osteogenesis and adipogenesis. J. Bone Miner. Res. 2004, 19, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Delp, C.J. Coping with resistance to plant disease. Plant Dis. 1980, 64, 652–657. [Google Scholar] [CrossRef]
- Filipov, N.M.; Lawrence, D.A. Developmental toxicity of a triazole fungicide: Consideration of interorgan communication. Toxicol. Sci. 2001, 62, 185–186. [Google Scholar] [CrossRef] [PubMed]
- Copping, L.G.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest Manag. Sci. 2000, 56, 651–676. [Google Scholar] [CrossRef]
- Dewick, P.M. Isoflavonoids. In The Flavonoids: Advances in Research since 1980; Harborne, J.B., Ed.; Springer: Boston, MA, USA, 1988; pp. 125–209. [Google Scholar]
- Varinska, L.; Gal, P.; Mojzisova, G.; Mirossay, L.; Mojzis, J. Soy and Breast Cancer: Focus on Angiogenesis. Int. J. Mol. Sci. 2015, 16, 11728–11749. [Google Scholar] [CrossRef] [PubMed]
- Krämer, R.P.; Hindorf, H.; Jha, H.C.; Kallage, J.; Zilliken, F. Antifungal activity of soybean and chickpea isoflavones and their reduced derivatives. Phytochemistry 1984, 23, 2203–2205. [Google Scholar]
- Smith, D.A. Some effects of the phytoalexin, kievitone, on the vegetative growth of Aphanomyces euteiches, Rhizoctonia solani and Fusarium solani f. sp. phaseoli. Physiol. Plant Pathol. 1976, 9, 45–48. [Google Scholar] [CrossRef]
- Siupp, R.A.; Bailey, J.A. The fungitoxicity of isoflavanoid phytoalexins measured using different types of bioassay. Physiol. Plant Pathol. 1977, 11, 101–112. [Google Scholar] [CrossRef]
- Perrin, D.R.; Cruickshank, I.A.M. The antifungal activity of pterocarpans towards Monilinia fructicola. Phytochemistry 1969, 8, 971–978. [Google Scholar] [CrossRef]
- Gnanamanickam, S.S.; Smith, D.A. Selective toxicity of isoflavonoid phytoalexins to gramspositive bacteria. Phytopathology 1980, 70, 894–896. [Google Scholar] [CrossRef]
- Lee, J.A.; Chee, H.Y. In Vitro Antifungal Activity of Equol against Candida albicans. Mycobiology 2010, 38, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Kuc, J. Phytoalexins. Annu. Rev. Phytopathol. 1972, 10, 207–232. [Google Scholar]
- Virtanen, A.I.; Hietala, P.K. Isolation of an anti-sclerotinia factor, 7-hydroxy-4’-methoxyisoflavone from red clover. Acta Chem. Scand. 1958, 12, 579–580. [Google Scholar] [CrossRef]
- Keen, N.T.; Sims, J.J.; Erwin, D.C.; Rice, E.; Partridge, J.E. 6a-hydroxyphasecollin: An antifungal chemical induces in soybean hypocotyls by Phytophthora megasperma var. sojae. Phytopathology 1971, 61, 1084–1089. [Google Scholar] [CrossRef]
- Naim, M.; Gestetner, B.; Zilkah, S.; Birk, Y.; Bondi, A. Soybean isoflavones. Characterization, determination, and antifungal activity. J. Agric. Food Chem. 1974, 22, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Weidenbörner, M.; Hindorf, H.; Jha, H.C.; Tsotsonos, P.; Egge, H. Antifungal activity of isoflavonoids in different reduced stages on Rhizoctonia solani and Sclerotium rolfsii. Phytochemistry 1990, 29, 801–803. [Google Scholar]
- Froyd, J.D.; Paget, C.J.; Guse, L.R.; Dreikorn, B.A.; Pafford, J.L. Tricyclazole—New systemic fungicide for control of Piricularia-oryzae on rice. Phytopathology 1976, 66, 1135–1139. [Google Scholar] [CrossRef]
- Yamaguchi, I. Overview on the chemical control of rice blast disease. In Rice Blast: Interaction with Rice and Control; Springer: Dordrecht, The Netherlands, 2004; pp. 1–13. [Google Scholar]
- Masukawa, H.; Tanaka, N.; Umezawa, H. Inhibition by kasugamygin of protein synthesis in Piricularia oryzae. J. Antibiot. 1968, 21, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Misato, T.; Ishii, I.; Asakawa, M.; Okimoto, Y.; Fukunaga, K. Antibiotics as protectant fungicides against rice blast. Jpn. J. Phytopathol. 1959, 24, 302–306. [Google Scholar] [CrossRef]
- Yoon, M.-Y.; Kim, Y.S.; Ryu, S.Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Cha, B.; Han, S.S.; Kim, J.C. In vitro and in vivo antifungal activities of decursin and decursinol angelate isolated from Angelica gigas against Magnaporthe oryzae, the causal agent of rice blast. Pestic. Biochem. Phys. 2011, 101, 118–124. [Google Scholar] [CrossRef]
- Kunova, A.; Pizzatti, C.; Cortesi, P. Impact of tricyclazole and azoxystrobin on growth, sporulation and secondary infection of the rice blast fungus, Magnaporthe oryzae. Pest Manag. Sci. 2013, 69, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.E.; Basarab, G.S.; Andersson, A.; Lindqvist, Y.; Jordan, D.B. Trihydroxynaphthalene reductase from Magnaporthe grisea: Realization of an active center inhibitor and elucidation of the kinetic mechanism. Biochemistry 1997, 36, 1852–1860. [Google Scholar] [CrossRef] [PubMed]
- Tokousbalides, M.C.; Sisler, H.D. Effect of tricyclazole on growth and secondary metabolism in Pyricularia oryzae. Pestic. Biochem. Phys. 1978, 8, 26–32. [Google Scholar] [CrossRef]
- Pak, D.; You, M.P.; Lanoiselet, V.; Barbetti, M.J. Azoxystrobin and propiconazole offer significant potential for rice blast (Pyricularia oryzae) management in Australia. Eur. J. Plant Pathol. 2017, 148, 247–259. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, X.; Yuan, S.K.; Li, Y.F.; Zhang, A.F.; Yao, J.; Gao, T.C. Effect of azoxystrobin and kresoxim-methyl on rice blast and rice grain yield in China. Ann. Appl. Biol. 2015, 166, 434–443. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Barton, M. Estrogen biology: New insights into GPER function and clinical opportunities. Mol. Cell. Endocrinol. 2014, 389, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.; Frankenfeld, C.L.; Lampe, J.W. Gut bacterial metabolism of the soy isoflavone daidzein: Exploring the relevance to human health. Exp. Biol. Med. (Maywood) 2005, 230, 155–170. [Google Scholar] [PubMed]
- Mueller, S.O.; Simon, S.; Chae, K.; Metzler, M.; Korach, K.S. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor α (ERα) and ERβ in human cells. Toxicol. Sci. 2004, 80, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Mezei, O.; Banz, W.J.; Steger, R.W.; Peluso, M.R.; Winters, T.A.; Shay, N. Soy isoflavones exert antidiabetic and hypolipidemic effects through the PPAR pathways in obese Zucker rats and murine RAW 264.7 cells. J. Nutr. 2003, 133, 1238–1243. [Google Scholar] [PubMed]
- Setchell, K.D.; Cole, S.J. Method of defining equol-producer status and its frequency among vegetarians. J. Nutr. 2006, 136, 2188–2193. [Google Scholar] [PubMed]
- Rowland, I.R.; Wiseman, H.; Sanders, T.A.; Adlercreutz, H.; Bowey, E.A. Interindividual variation in metabolism of soy isoflavones and lignans: Influence of habitual diet on equol production by the gut microflora. Nutr. Cancer 2000, 36, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Yamaguchi, M.; Sobue, T.; Takahashi, T.; Miura, T.; Arai, Y.; Mazur, W.; Wähälä, K.; Adlercreutz, H. Pharmacokinetics of soybean isoflavones in plasma, urine and feces of men after ingestion of 60 g baked soybean powder (kinako). J. Nutr. 1998, 128, 1710–1715. [Google Scholar] [PubMed]
- Arai, Y.; Uehara, M.; Sato, Y.; Kimira, M.; Eboshida, A.; Adlercreutz, H.; Watanabe, S. Comparison of isoflavones among dietary intake, plasma concentration and urinary excretion for accurate estimation of phytoestrogen intake. J. Epidemiol. 2000, 10, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Akaza, H.; Miyanaga, N.; Takashima, N.; Naito, S.; Hirao, Y.; Tsukamoto, T.; Fujioka, T.; Mori, M.; Kim, W.J.; Song, J.M.; Pantuck, A.J. Comparisons of percent equol producers between prostate cancer patients and controls: Case-controlled studies of isoflavones in Japanese, Korean and American residents. Jpn. J. Clin. Oncol. 2004, 34, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Song, K.B.; Atkinson, C.; Frankenfeld, C.L.; Jokela, T.; Wähälä, K.; Thomas, W.K.; Lampe, J.W. Prevalence of daidzein-metabolizing phenotypes differs between Caucasian and Korean American women and girls. J. Nutr. 2006, 136, 1347–1351. [Google Scholar] [PubMed]
- Vétizo, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Decroos, K.; Vanhemmens, S.; Cattoir, S.; Boon, N.; Verstraete, W. Isolation and characterisation of an equol-producing mixed microbial culture from a human faecal sample and its activity under gastrointestinal conditions. Arch. Microbiol. 2005, 183, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.T.; Yao, W.; Zhu, W.Y. Isolation and identification of equol-producing bacterial strains from cultures of pig faeces. FEMS Microbiol. Lett. 2008, 282, 73–80. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are commercially available from the companies referred. |








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, L.; Yin, Y.; Gu, Z.; Chai, R.; Wang, Y.; Sun, G. Equol, a Clinically Important Metabolite, Inhibits the Development and Pathogenicity of Magnaporthe oryzae, the Causal Agent of Rice Blast Disease. Molecules 2017, 22, 1799. https://doi.org/10.3390/molecules22101799
Wang J, Li L, Yin Y, Gu Z, Chai R, Wang Y, Sun G. Equol, a Clinically Important Metabolite, Inhibits the Development and Pathogenicity of Magnaporthe oryzae, the Causal Agent of Rice Blast Disease. Molecules. 2017; 22(10):1799. https://doi.org/10.3390/molecules22101799
Chicago/Turabian StyleWang, Jiaoyu, Ling Li, Yeshi Yin, Zhuokan Gu, Rongyao Chai, Yanli Wang, and Guochang Sun. 2017. "Equol, a Clinically Important Metabolite, Inhibits the Development and Pathogenicity of Magnaporthe oryzae, the Causal Agent of Rice Blast Disease" Molecules 22, no. 10: 1799. https://doi.org/10.3390/molecules22101799
APA StyleWang, J., Li, L., Yin, Y., Gu, Z., Chai, R., Wang, Y., & Sun, G. (2017). Equol, a Clinically Important Metabolite, Inhibits the Development and Pathogenicity of Magnaporthe oryzae, the Causal Agent of Rice Blast Disease. Molecules, 22(10), 1799. https://doi.org/10.3390/molecules22101799