Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis


 ,
,
Abstract
:1. Introduction
2. Results
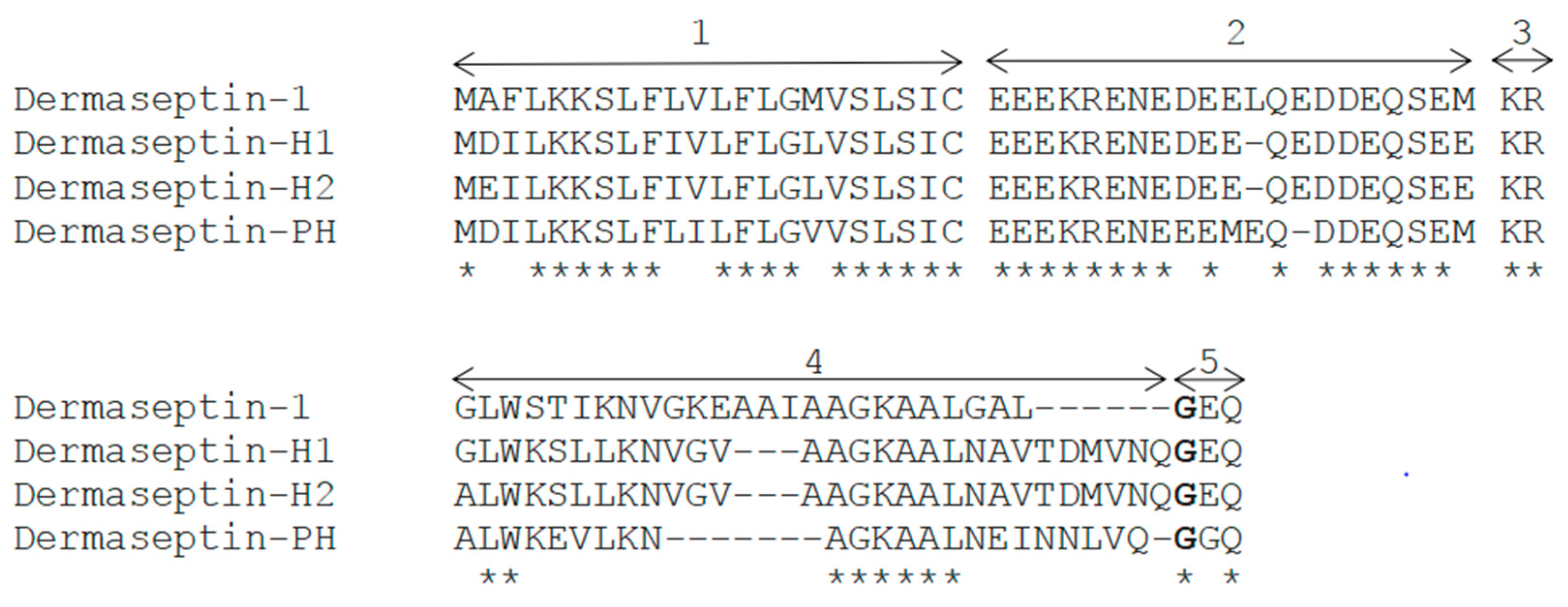
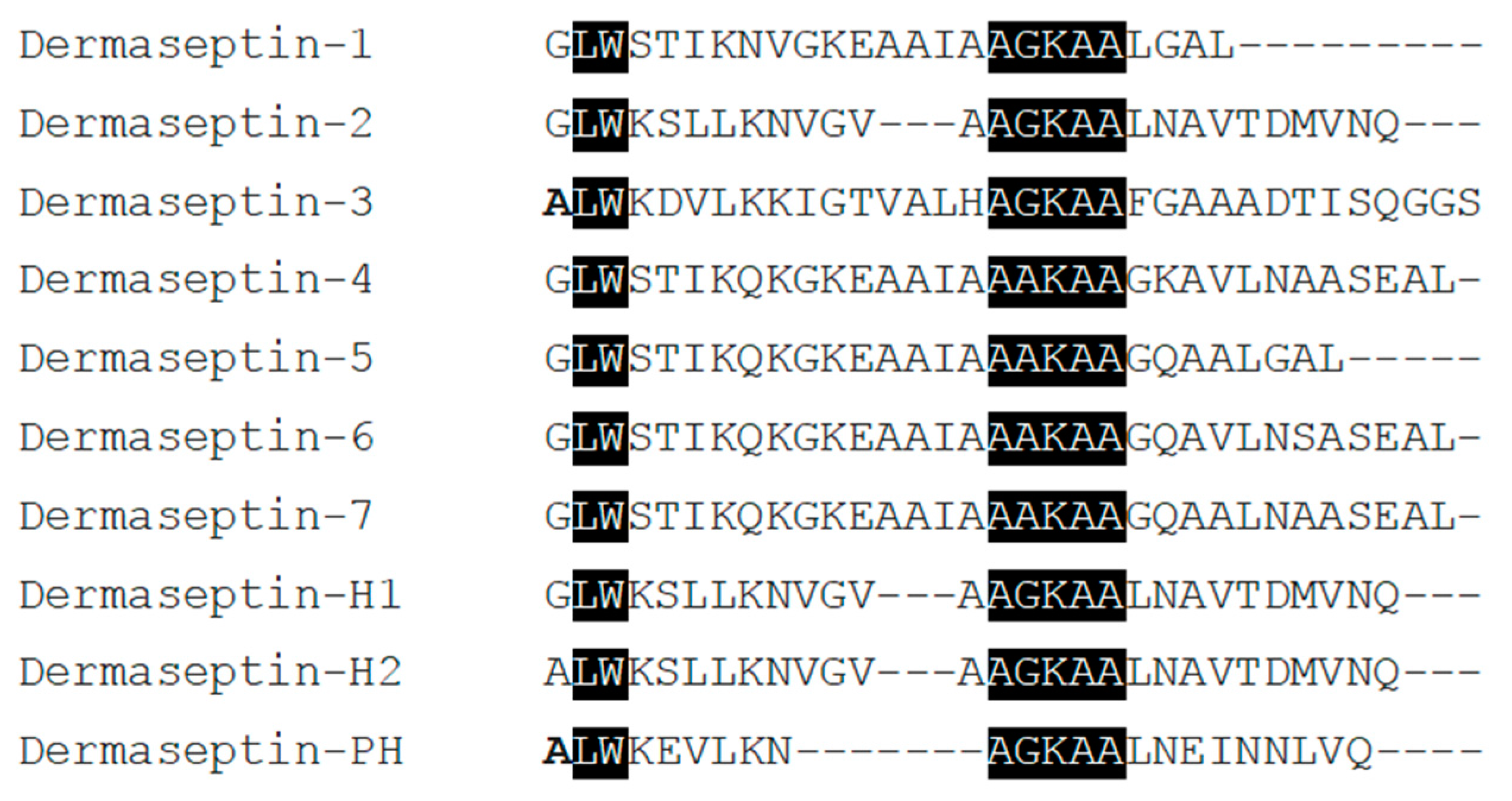
2.1. “Shotgun” Cloning of Dermaseptin-PH Precursor-Encoding cDNA from a Skin Secretion-Derived cDNA Library of Pithecopus hypochondrialis and Bioinformatic Analysis
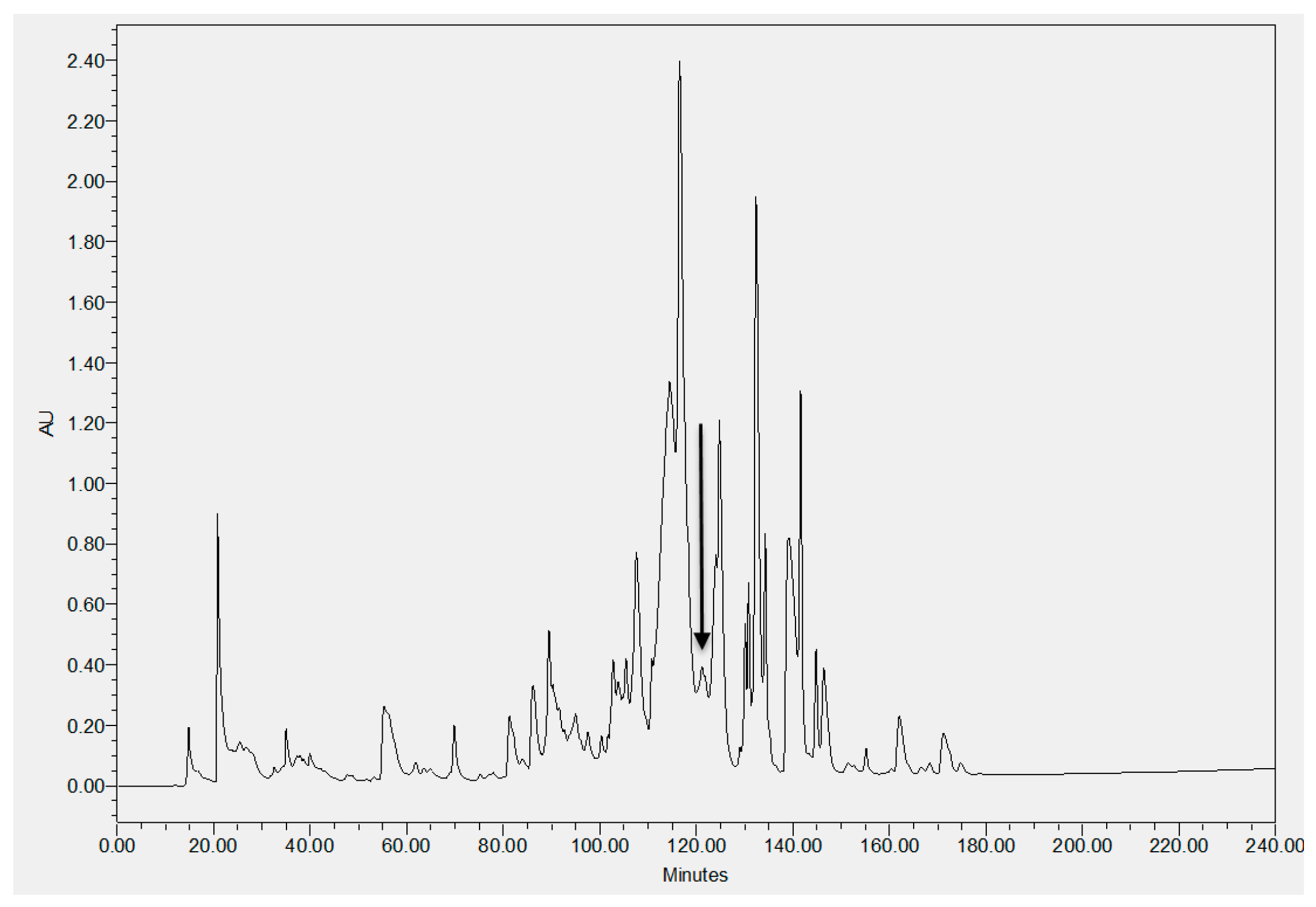
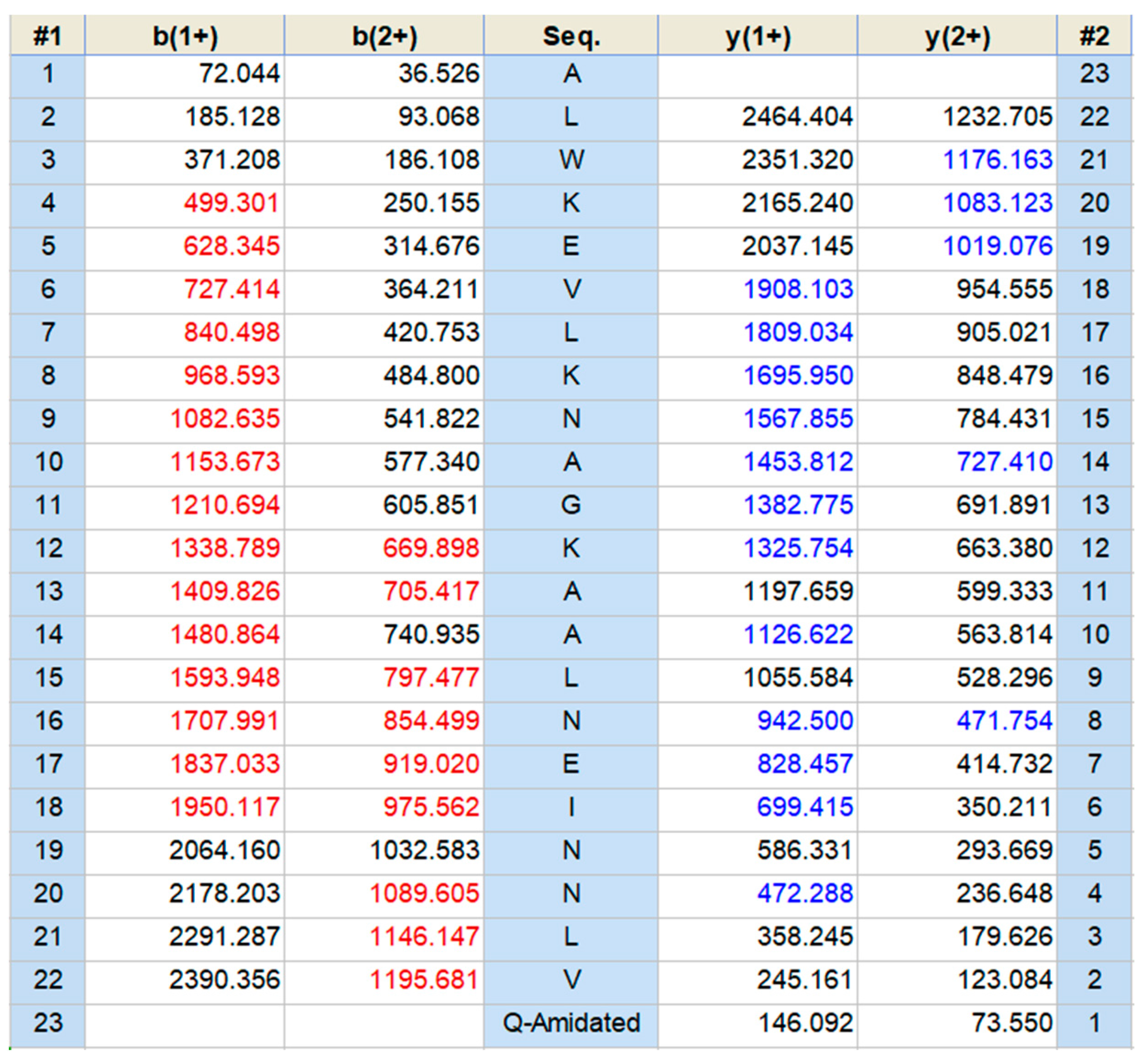
2.2. Identification and Structural Characterization of Dermaseptin-PH in Skin Secretion of Pithecopus hypochondrialis
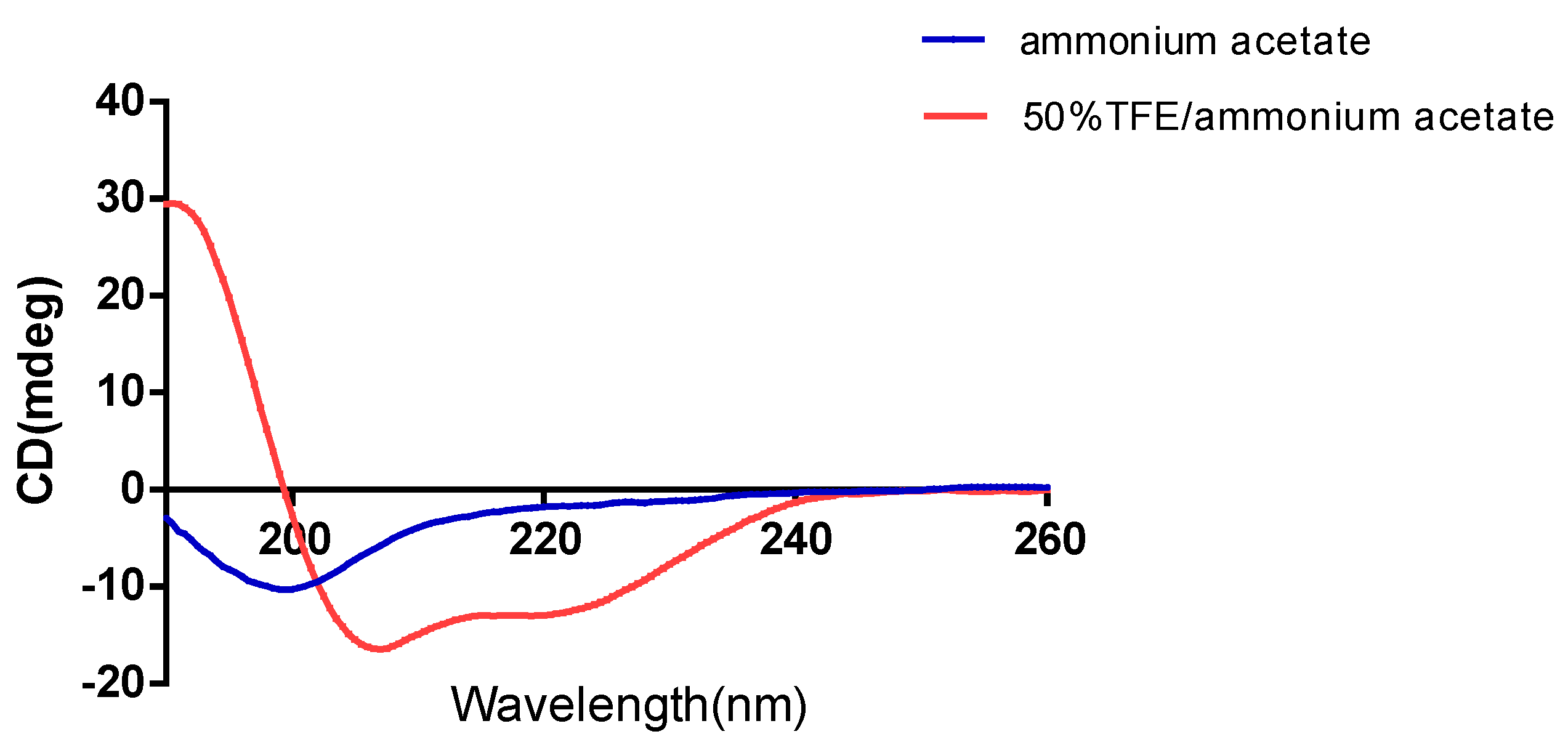
2.3. Circular Dichroism Spectroscopy
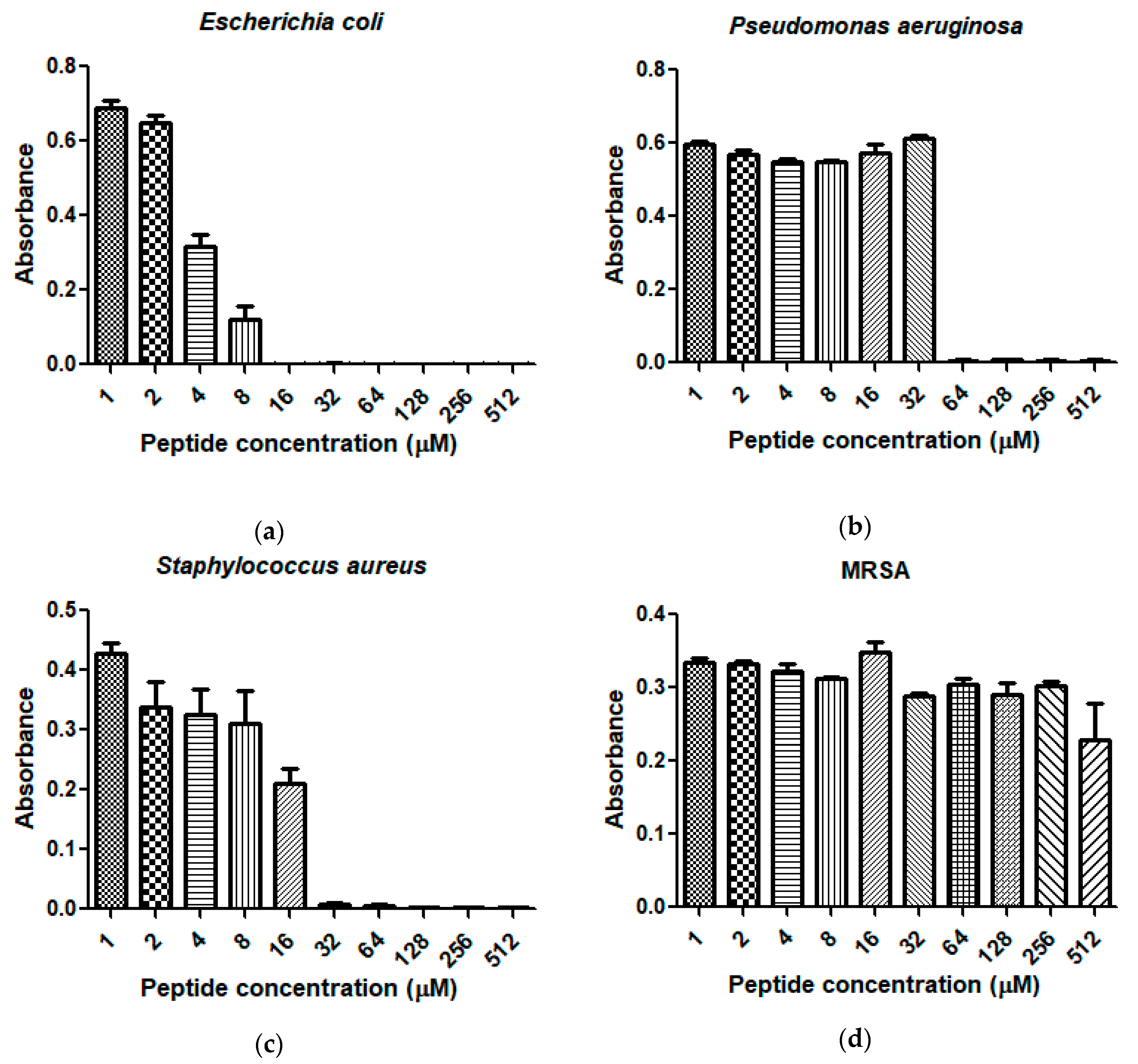
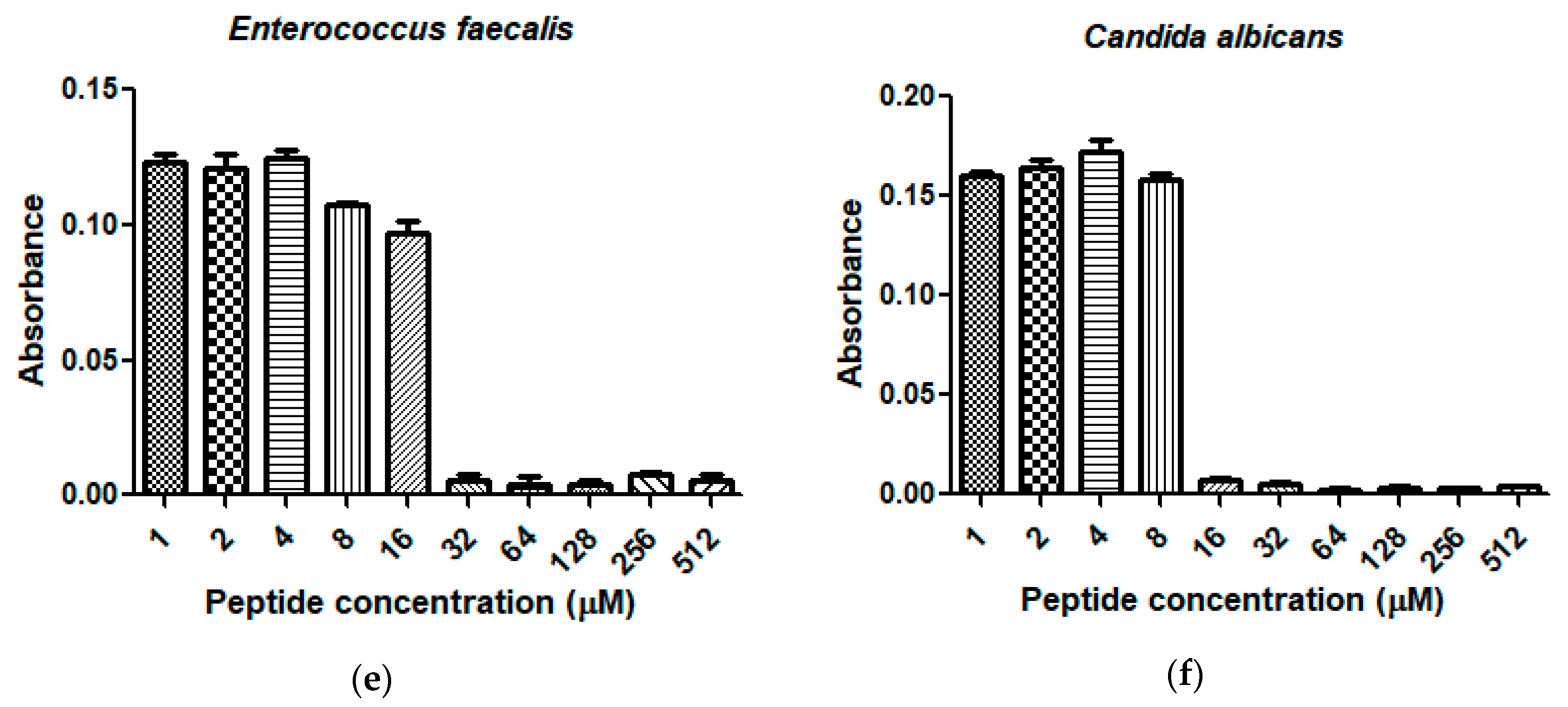
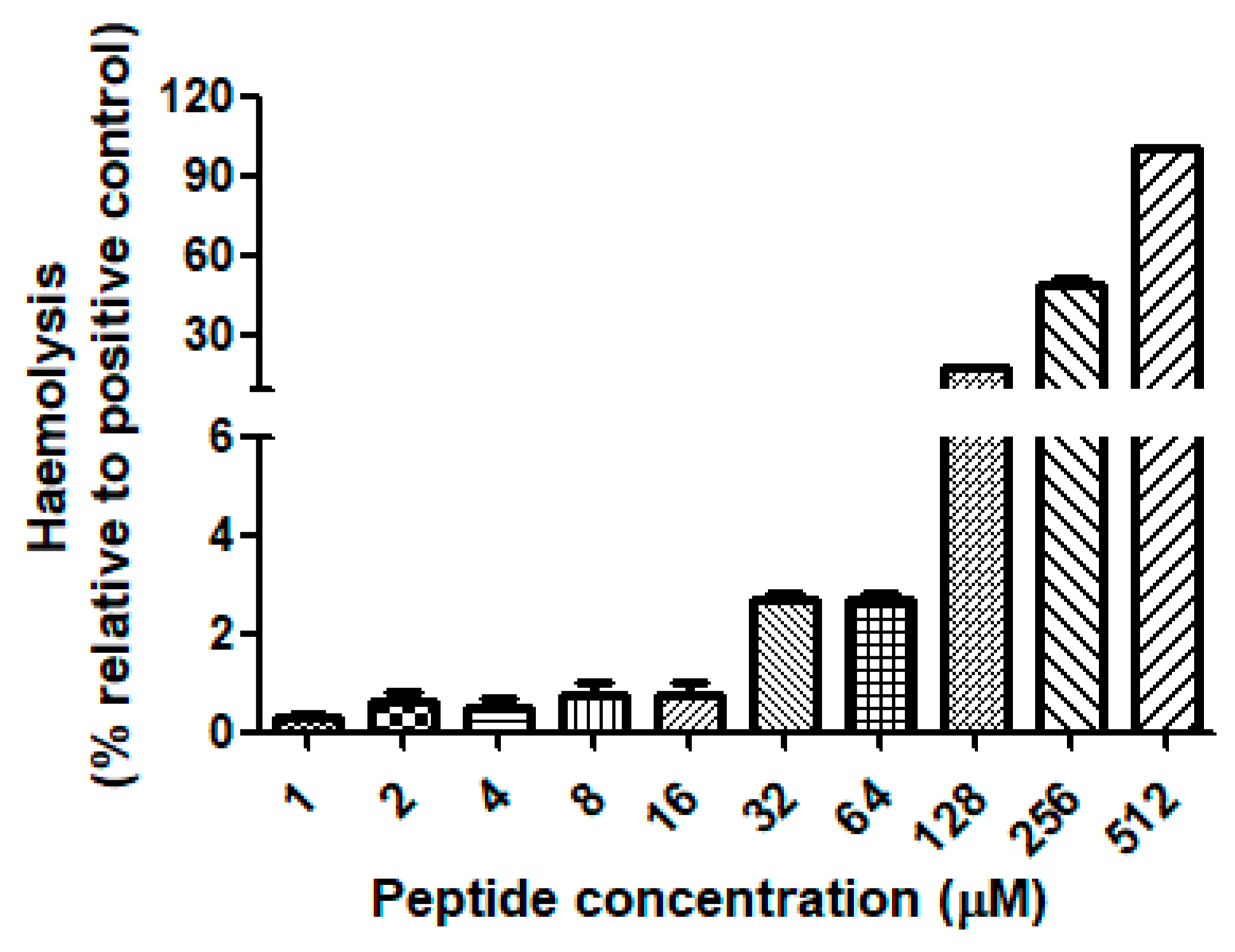
2.4. Antimicrobial Activity and Haemolytic Activity of Dermaseptin-PH
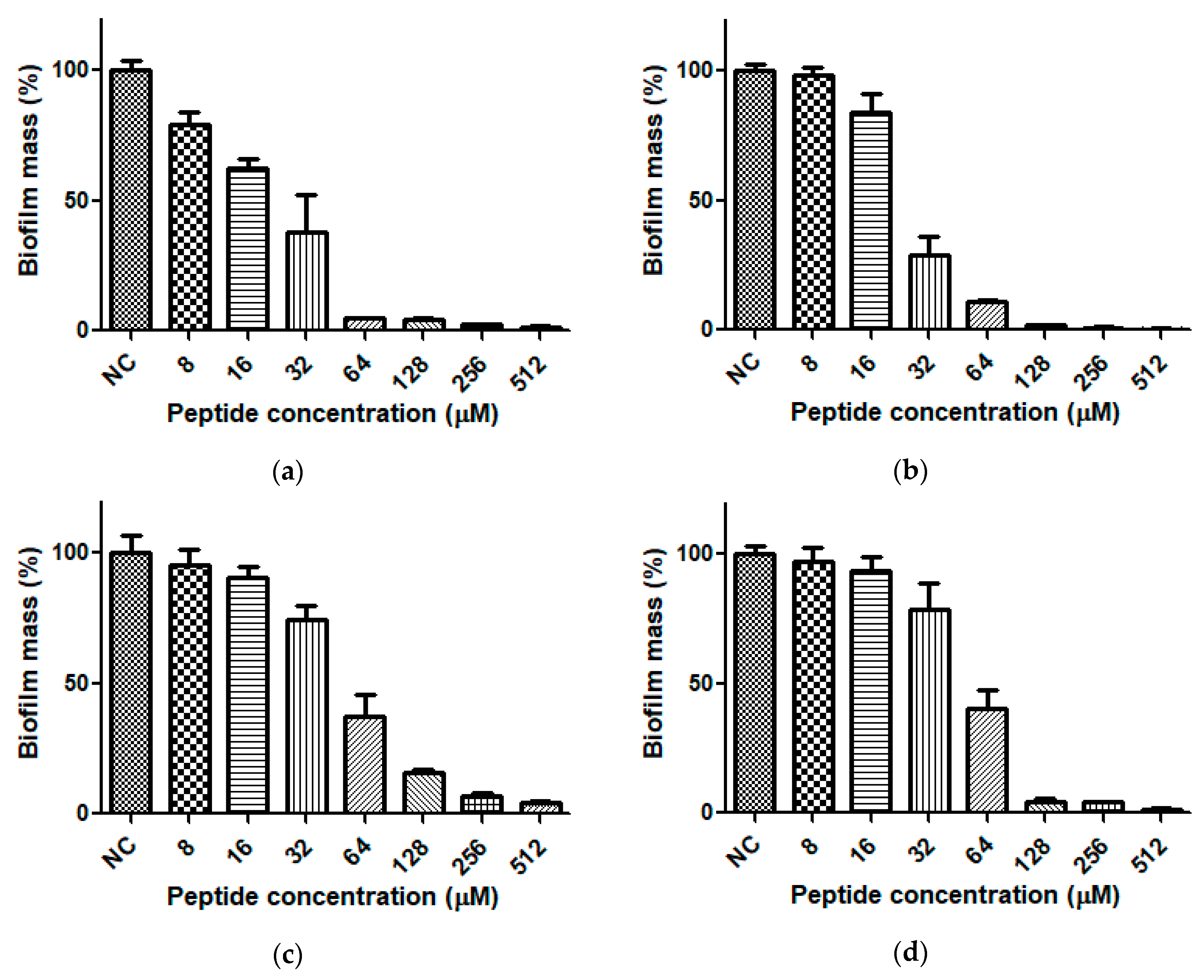
2.5. Anti-Biofilm Activity of Dermaseptin-PH
2.6. Bacteria Cell Membrane Permeabilisation Activities of Dermaseptin-PH
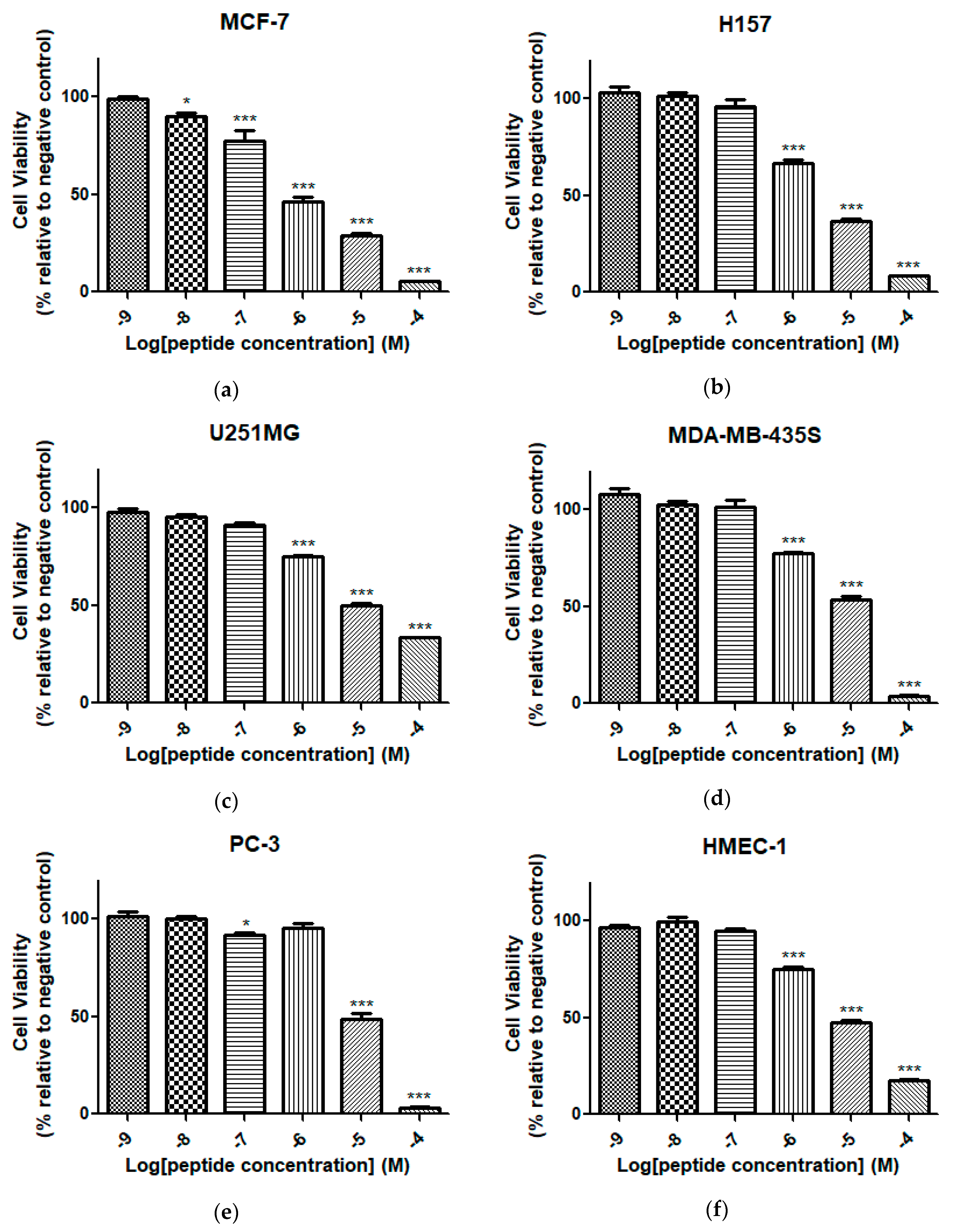
2.7. Cell Proliferation Assay of Dermaseptin-PH
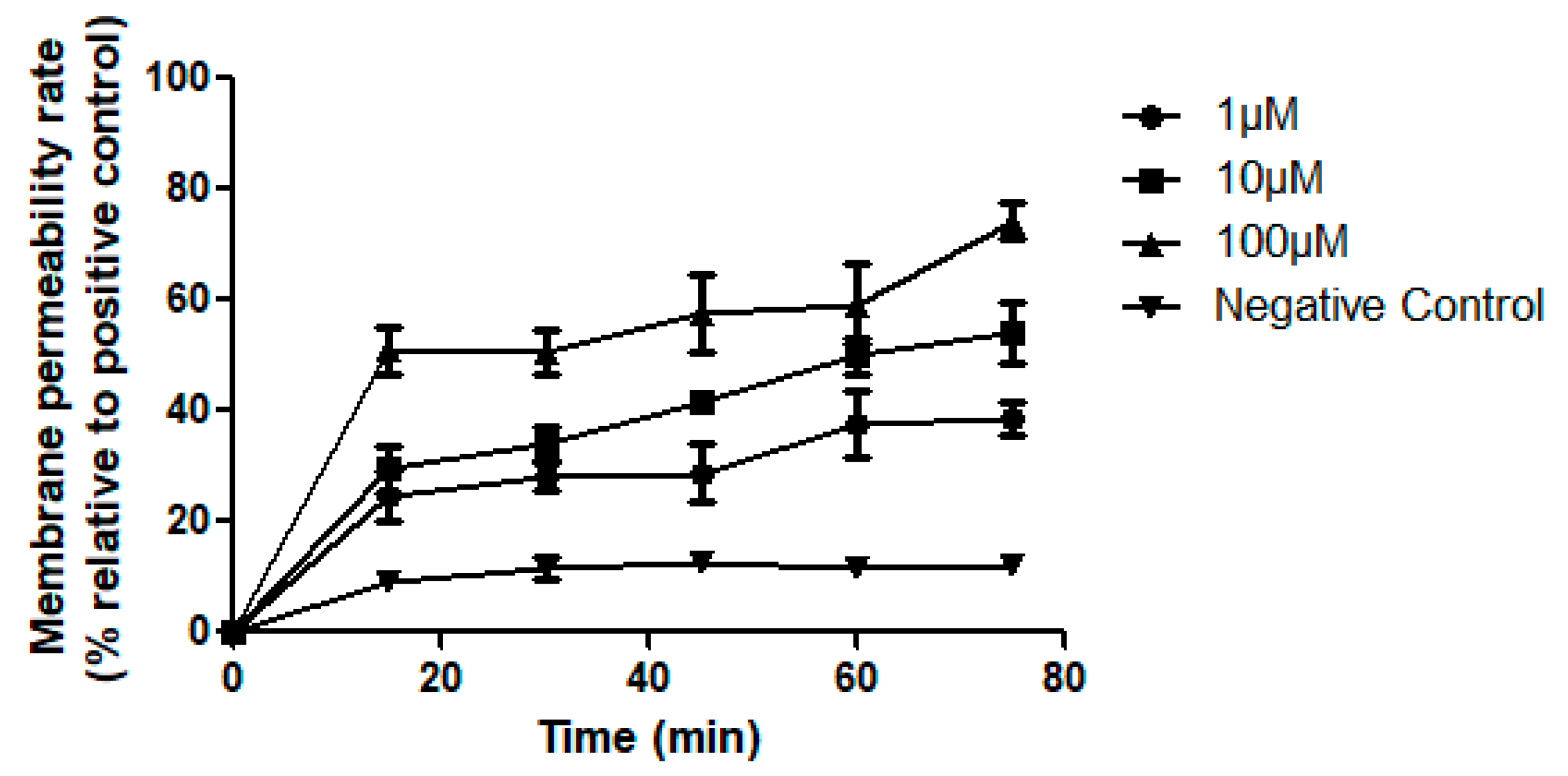
2.8. Cancer Cell Membrane Permeabilisation Activities of Dermaseptin-PH
3. Discussion
4. Materials and Methods
4.1. Acquisition of Pithecopus hypochondrialis Skin Secretion
4.2. “Shotgun” Cloning of Dermaseptin-PH Precursor-Encoding cDNA from a Skin Secretion-Derived cDNA Library of Pithecopus hypochondrialis
4.3. Identification and Structural Characterization of the Putative Dermaseptin-PH from the Skin Secretion of Pithecopus hypochondrialis
4.4. Solid-Phase Peptide Synthesis
4.5. Determination of Dermaseptin-PH Second Structure Using Circular Dichroism Spectroscopy
4.6. Antimicrobial Assays
4.7. Minimum Biofilm Inhibitory Concentration (MBIC) and Minimum Biofilm Eradication Concentration (MBEC) Assays
4.8. Bacteria Cell Membrane Permeability Assay of Dermaseptin-PH
4.9. Haemolysis Assay
4.10. MTT Cell Proliferation Assay
4.11. Cancer Cell Membrane Permeability Assay of Dermaseptin-PH
Author Contributions
Conflicts of Interest
Abbreviations
| RACE | rapid amplification of cDNA ends |
| PCR | polymerase chain reaction |
| RP-HPLC | reverse-phase high-performance liquid chromatography |
| AMPs | antimicrobial peptides |
| HDPs | host-defence peptides |
| NCBI | National Centre for Biotechnology Information |
| BLAST | basic local alignment search tool |
| MALDI-TOF-MS | matrix-assisted laser desorption/ionization time-of-flight mass spectrometry |
| TFA | trifluoracetic acid |
| CHCA | α-cyano-4-hydroxycinnamic acid |
| TFE | 2,2,2-trifluoroethanol |
| Fmoc | 9-fluorenylmethyloxycarbonyl |
References
- Da Cunha, N.B.; Cobacho, N.B.; Viana, J.F.; Lima, L.A.; Sampaio, K.B.; Dohms, S.S.; Ferreira, A.C.; de la Fuente-Nunez, C.; Costa, F.F.; Franco, O.L.; et al. The next generation of antimicrobial peptides (AMPs) as molecular therapeutic tools for the treatment of diseases with social and economic impacts. Drug Discov. Today 2017, 22, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Lehrer, R. Cationic peptides: A new source of antibiotics. Trends Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bowdish, D.; Davidson, D.; Hancock, R. A re-evaluation of the role of host defence peptides in mammalian immunity. Curr. Protein Pept. Sci. 2005, 6, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Oppenheim, J.J.; Yang, D. Alarmins: Chemotactic activators of immune responses. Curr. Opin. Immunol. 2005, 17, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Naas, T.; Fortineau, N.; Poirel, L. Superbugs in the coming new decade; multidrug resistance and prospects for treatment of Staphylococcus aureus, Enterococcus spp. and Pseudomonas aeruginosa in 2010. Curr. Opin. Microbiol. 2007, 10, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.; Kellner, J.D.; Low, D.E. The superbugs: Evolution, dissemination and fitness. Curr. Opin. Microbiol. 1998, 1, 524–529. [Google Scholar] [CrossRef]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Burns, J.L.; Montelaro, R.C. Engineered cationic antimicrobial peptides to overcome multidrug resistance by ESKAPE pathogens. Antimicrob. Agents Chemother. 2015, 59, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Karam, G.; Chastre, J.; Wilcox, M.H.; Vincent, J.L. Antibiotic strategies in the era of multidrug resistance. Crit. Care 2016, 20, 136. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, A.; Sigel, S.; Ljunggren, L. The antimicrobial peptide LL37 and its truncated derivatives potentiates proinflammatory cytokine induction by lipoteichoic acid in whole blood. Scand. J. Clin. Lab. Investig. 2010, 70, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Intratumoral Injections of LL37 for Melanoma. Available online: https://clinicaltrials.gov/show/NCT02225366 (accessed on 22 September 2017).
- Yamazaki, T.; Pitt, J.M.; Vetizou, M.; Marabelle, A.; Flores, C.; Rekdal, O.; Kroemer, G.; Zitvogel, L. The oncolytic peptide LTX-315 overcomes resistance of cancers to immunotherapy with CTLA4 checkpoint blockade. Cell Death Differ. 2016, 23, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Camilio, K.A.; Rekdal, O.; Sveinbjornsson, B. LTX-315 (Oncopore): A short synthetic anticancer peptide and novel immunotherapeutic agent. Oncoimmunology 2014, 3, e29181. [Google Scholar] [CrossRef] [PubMed]
- Spicer, J.; Awada, A.; Brunsvig, P.; Kristeleit, R.; Jøssang, D.; Saunders, A.; Olsen, W.M.; Nicolaisen, B.; Rekdal, Ø.; Laruelle, M. In Intra-tumoural treatment with LTX-315, an oncolytic peptide immunotherapy, in patients with advanced metastatic disease induces infiltration of CD8 effectors T-cells and regression in some injected tumors. In Proceedings of the 8th European Cancer Congress (ECC2015), Vienna, Austria, 25–29 September 2015. [Google Scholar]
- Duellman, W.E.; Marion, A.B.; Hedges, S.B. Phylogenetics, classification, and biogeography of the treefrogs (Amphibia: Anura: Arboranae). Zootaxa 2016, 4104, 1–109. [Google Scholar] [CrossRef] [PubMed]
- Vouille, V.; Amiche, M.; Nicolas, P. Structure of genes for Dermaseptins B, antimicrobial peptides from frog skin. FEBS Lett. 1997, 414, 27–32. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, P.; El Amri, C. The dermaseptin superfamily: A gene-based combinatorial library of antimicrobial peptides. Biochim. Biophys. Acta (BBA)—Biomembr. 2009, 1788, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Van Huong, N.; Delfour, A.; Migliore-Samour, D.; Nicolas, P. Isolation, amino acid sequence and synthesis of dermaseptin, a novel antimicrobial peptide of amphibian skin. Biochemistry 1991, 30, 8824–8830. [Google Scholar] [CrossRef] [PubMed]
- Brand, G.D.; Leite, J.R.S.; de Sá Mandel, S.M.; Mesquita, D.A.; Silva, L.P.; Prates, M.V.; Barbosa, E.A.; Vinecky, F.; Martins, G.R.; Galasso, J.H. Novel dermaseptins from Phyllomedusa hypochondrialis (amphibia). Biochem. Biophys. Res. Commun. 2006, 347, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Conceição, K.; Konno, K.; Richardson, M.; Antoniazzi, M.M.; Jared, C.; Daffre, S.; Camargo, A.C.M.; Pimenta, D.C. Isolation and biochemical characterization of peptides presenting antimicrobial activity from the skin of Phyllomedusa hypochondrialis. Peptides 2006, 27, 3092–3099. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhou, M.; Gagliardo, R.; Walker, B.; Shaw, C. Elements of the granular gland peptidome and transcriptome persist in air-dried skin of the south American orange-legged leaf frog, Phyllomedusa hypocondrialis. Peptides 2006, 27, 2129–2136. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wu, D.; Xi, X.; Wu, Y.; Ma, C.; Zhou, M.; Wang, L.; Yang, M.; Chen, T.; Shaw, C. Identification and characterisation of the antimicrobial peptide, Phylloseptin-PT, from the skin secretion of Phyllomedusa tarsius, and comparison of activity with designed, cationicity-enhanced analogues and diastereomers. Molecules 2016, 21, 1667. [Google Scholar] [CrossRef] [PubMed]
- Lequin, O.; Bruston, F.; Convert, O.; Chassaing, G.; Nicolas, P. Helical structure of Dermaseptin B2 in a membrane-mimetic environment. Biochemistry 2003, 42, 10311–10323. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.W.; Wong, G.C. Antimicrobial peptides and induced membrane curvature: Geometry, coordination chemistry, and molecular engineering. Curr. Opin. Solid State Mater. Sci. 2013, 17, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Lequin, O.; Ladram, A.; Chabbert, L.; Bruston, F.; Convert, O.; Vanhoye, D.; Chassaing, G.; Nicolas, P.; Amiche, M. Dermaseptin S9, an α-helical antimicrobial peptide with a hydrophobic core and cationic termini. Biochemistry 2006, 45, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.A.; Murray, B.E. Antibiotic-resistant bugs in the 21st century—A clinical super-challenge. N. Engl. J. Med. 2009, 360, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta BBA Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.L.; Rushton, G.; Jayson, G.C.; Avizienyte, E. Ovarian cancer cell heparan sulfate 6-O-sulfotransferases regulate an angiogenic program induced by heparin-binding epidermal growth factor (EGF)-like growth factor/EGF receptor signaling. J. Biol. Chem. 2014, 289, 10488–10501. [Google Scholar] [CrossRef] [PubMed]
- Mu, A.K.-W.; Lim, B.-K.; Hashim, O.H.; Shuib, A.S. Identification of O-glycosylated proteins that are aberrantly excreted in the urine of patients with early stage ovarian cancer. Int. J. Mol. Sci. 2013, 14, 7923–7931. [Google Scholar] [CrossRef] [PubMed]
- Osinaga, E. Expression of cancer-associated simple mucin-type O-glycosylated antigens in parasites. IUBMB Life 2007, 59, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Esteva, F.J.; Song, J.; Zhang, H. Altered expression of sialylated glycoproteins in breast cancer using hydrazide chemistry and mass spectrometry. Mol. Cell. Proteome. 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Fadnes, B.; Rekdal, Ø.; Uhlin-Hansen, L. The anticancer activity of lytic peptides is inhibited by heparan sulfate on the surface of the tumor cells. BMC Cancer 2009, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635. [Google Scholar] [CrossRef] [PubMed]
- Sok, M.; Šentjurc, M.; Schara, M.; Stare, J.; Rott, T. Cell membrane fluidity and prognosis of lung cancer. Ann. Thorac. Surg. 2002, 73, 1567–1571. [Google Scholar] [CrossRef]
- Choi, H.; Lee, W.; Lee, D. A new concept on mechanism of antimicrobial peptides: Apoptosis induction. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex Research Center: Badajoz, Spain, 2013; p. 4. [Google Scholar]
- Morton, C.O.; Dos Santos, S.C.; Coote, P. An amphibian-derived, cationic, α-helical antimicrobial peptide kills yeast by caspase-independent but AIF-dependent programmed cell death. Mol. Microbiol. 2007, 65, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Van Zoggel, H.; Carpentier, G.; Dos Santos, C.; Hamma-Kourbali, Y.; Courty, J.; Amiche, M.; Delbé, J. Antitumor and angiostatic activities of the antimicrobial peptide Dermaseptin B2. PLoS ONE 2012, 7, e44351. [Google Scholar] [CrossRef] [PubMed]
- Wäldchen, S.; Lehmann, J.; Klein, T.; Van De Linde, S.; Sauer, M. Light-induced cell damage in live-cell super-resolution microscopy. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Tyler, M.J.; Stone, D.J.; Bowie, J.H. A novel method for the release and collection of dermal, glandular secretions from the skin of frogs. J. Pharmacol. Toxicol. Methods 1992, 28, 199–200. [Google Scholar] [CrossRef]
- Kaur, G.; Balamurugan, P.; Maheswari, C.U.; Anitha, A.; Princy, S.A. Combinatorial effects of aromatic 1, 3-disubstituted ureas and fluoride on in vitro inhibition of streptococcus mutans biofilm formation. Front. Microbial. 2016, 7, 861. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (μM) | MBC (μM) | ||||
|---|---|---|---|---|---|---|
| Dermaseptin-PH | Amp | Nor | Dermaseptin-PH | Amp | Nor | |
| Escherichia coli | 16 | 36.6 | 0.6 | 16 | 36.6 | 0.6 |
| Pseudomonas aeruginosa | 64 | - | 2.5 | >512 | - | 5.2 |
| Staphylococcus aureus | 32 | 0.3 | 1.3 | 64 | 0.3 | 2.5 |
| MRSA | >512 | - | 2.5 | >512 | - | 5.2 |
| Enterococcus faecalis | 32 | 4.8 | 5.2 | 64 | 4.8 | 5.2 |
| Candida albicans | 16 | - | 1.3 | 32 | - | 2.5 |
| Strains | MBIC50 (μM) | MBIC90 (μM) | MBEC50 (μM) | MBEC90 (μM) |
|---|---|---|---|---|
| E. coli | 32 | 64 | 64 | 256 |
| S. aureus | 32 | 64 | 64 | 128 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. https://doi.org/10.3390/molecules22101805
Huang L, Chen D, Wang L, Lin C, Ma C, Xi X, Chen T, Shaw C, Zhou M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules. 2017; 22(10):1805. https://doi.org/10.3390/molecules22101805
Chicago/Turabian StyleHuang, Linyuan, Dong Chen, Lei Wang, Chen Lin, Chengbang Ma, Xinping Xi, Tianbao Chen, Chris Shaw, and Mei Zhou. 2017. "Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis" Molecules 22, no. 10: 1805. https://doi.org/10.3390/molecules22101805
APA StyleHuang, L., Chen, D., Wang, L., Lin, C., Ma, C., Xi, X., Chen, T., Shaw, C., & Zhou, M. (2017). Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules, 22(10), 1805. https://doi.org/10.3390/molecules22101805