
In Vitro Glucuronidation and Sulfation of ε-Viniferin, a Resveratrol Dimer, in Humans and Rats

 ,
, 
Abstract
:1. Introduction
2. Results
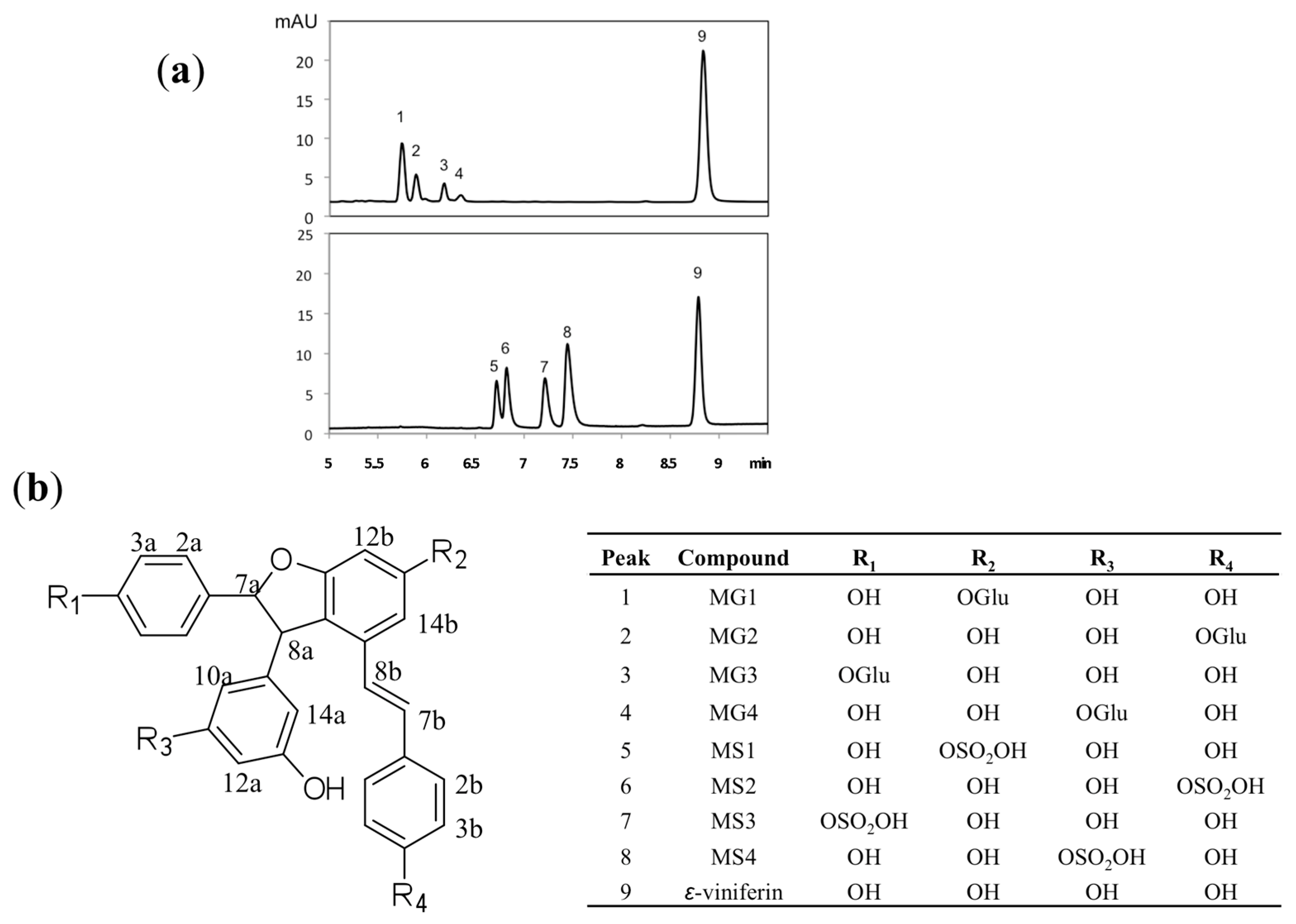
2.1. Hemi-Synthesis and Structural Identification of ε-Viniferin Metabolites
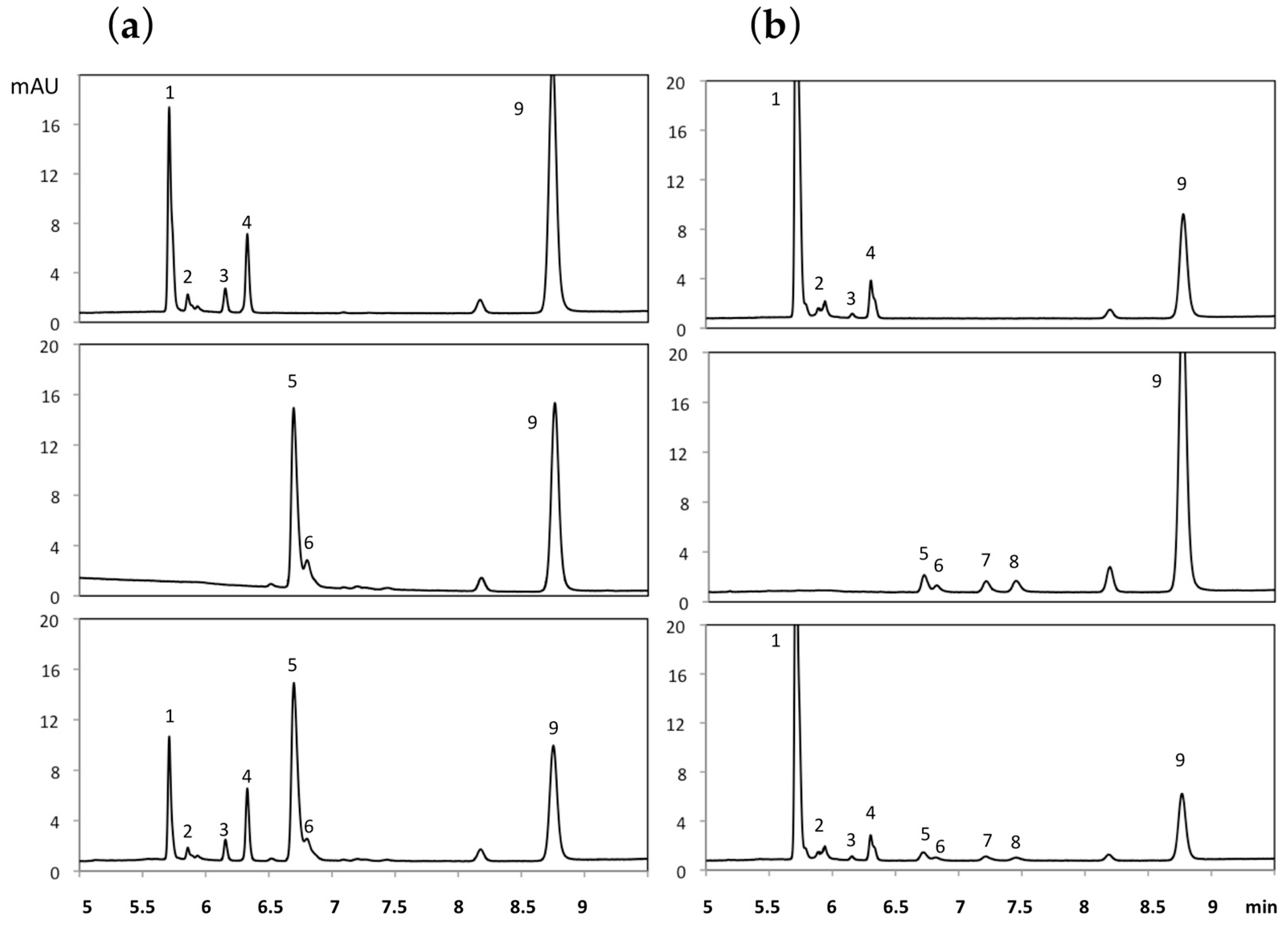
2.2. In Vitro ε-Viniferin Conjugates Profiling in Humans and Rats
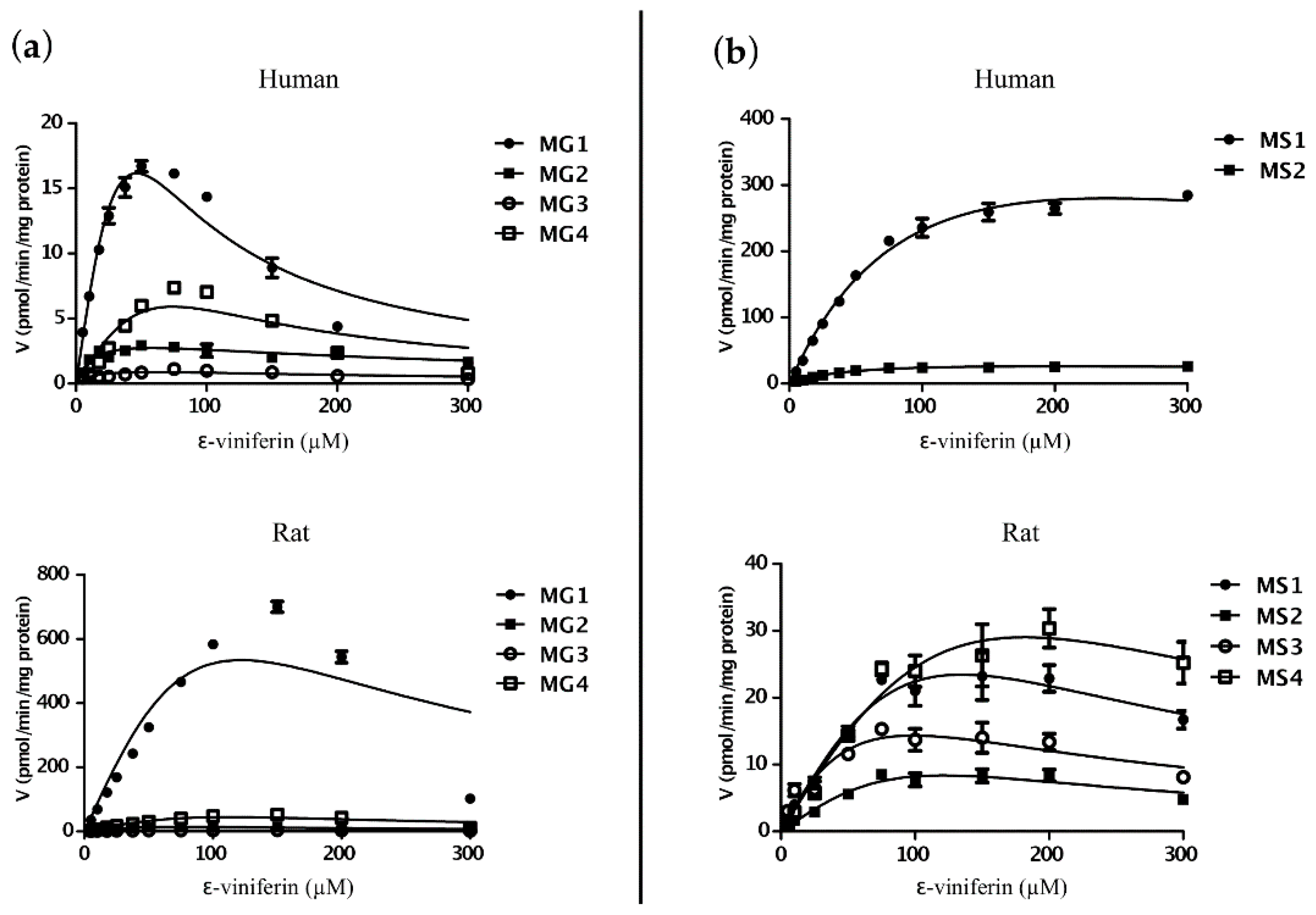
2.3. Kinetic Studies for Phase II Metabolism of ε-Viniferin
2.4. Interspecies Differences in Phase II Metabolism of ε-Viniferin
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Hemi-Synthesis of ε-Viniferin Sulfates
4.3. Hemi-Synthesis of ε-Viniferin Glucuronides
4.4. Semi-Preparative HPLC of Metabolites
4.5. UPLC-DAD-MS Analysis
4.6. NMR Analysis
4.7. Kinetic Study
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Singh, C.K.; Ndiaye, M.A.; Ahmad, N. Resveratrol and cancer: Challenges for clinical translation. Biochim. Biophys. Acta 2015, 1852, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T.; Szkudelska, K. Resveratrol and diabetes: From animal to human studies. Biochim. Biophys. Acta 2015, 1852, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Tellone, E.; Galtieri, A.; Russo, A.; Giardina, B.; Ficarra, S. Resveratrol: A Focus on Several Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2015, 2015, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Raj, P.; Zieroth, S.; Netticadan, T. An overview of the efficacy of resveratrol in the management of ischemic heart disease: Resveratrol in managing ischemic heart disease. Ann. N. Y. Acad. Sci. 2015, 1348, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Rivière, C.; Pawlus, A.D.; Mérillon, J.-M. Natural stilbenoids: Distribution in the plant kingdom and chemotaxonomic interest in Vitaceae. Nat. Prod. Rep. 2012, 29, 1317–1333. [Google Scholar] [CrossRef] [PubMed]
- Pawlus, A.D.; Waffo-Téguo, P.; Shaver, J.; Mérillon, J.-M. Stilbenoid chemistry from wine and the genus Vitis, a review. J. Int. Sci. Vigne Vin 2012, 46, 57–111. [Google Scholar] [CrossRef]
- Privat, C.; Telo, J.P.; Bernardes-Genisson, V.; Vieira, A.; Souchard, J.-P.; Nepveu, F. Antioxidant properties of trans-epsilon-viniferin as compared to stilbene derivatives in aqueous and nonaqueous media. J. Agric. Food Chem. 2002, 50, 1213–1217. [Google Scholar] [CrossRef] [PubMed]
- Nassra, M.; Krisa, S.; Papastamoulis, Y.; Kapche, G.D.; Bisson, J.; André, C.; Konsman, J.-P.; Schmitter, J.-M.; Mérillon, J.-M.; Waffo-Téguo, P. Inhibitory activity of plant stilbenoids against nitric oxide production by lipopolysaccharide-activated microglia. Planta Med. 2013, 79, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Billard, C.; Izard, J.-C.; Roman, V.; Kern, C.; Mathiot, C.; Mentz, F.; Kolb, J.-P. Comparative antiproliferative and apoptotic effects of resveratrol, epsilon-viniferin and vine-shots derived polyphenols (vineatrols) on chronic B lymphocytic leukemia cells and normal human lymphocytes. Leuk. Lymphoma 2002, 43, 1991–2002. [Google Scholar] [CrossRef] [PubMed]
- Colin, D.; Gimazane, A.; Lizard, G.; Izard, J.-C.; Solary, E.; Latruffe, N.; Delmas, D. Effects of resveratrol analogs on cell cycle progression, cell cycle associated proteins and 5fluoro-uracil sensitivity in human derived colon cancer cells. Int. J. Cancer 2009, 124, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Zghonda, N.; Yoshida, S.; Araki, M.; Kusunoki, M.; Mliki, A.; Ghorbel, A.; Miyazaki, H. Greater effectiveness of ε-viniferin in red wine than its monomer resveratrol for inhibiting vascular smooth muscle cell proliferation and migration. Biosci. Biotechnol. Biochem. 2011, 75, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Vina, J.; et al. Properties of Resveratrol: In Vitro and In Vivo Studies about Metabolism, Bioavailability, and Biological Effects in Animal Models and Humans. Oxid. Med. Cell. Longev. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, E.; Soldo, T.; Erbersdobler, H.; Somoza, V. Bioactivity and metabolism of trans-resveratrol orally administered to Wistar rats. Mol. Nutr. Food Res. 2005, 49, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Basu, S.; Meng, S.; Wang, X.; Hu, M. Regioselective sulfation and glucuronidation of phenolics: Insights into the structural basis. Curr. Drug Metab. 2011, 12, 900–916. [Google Scholar] [CrossRef] [PubMed]
- Aumont, V.; Krisa, S.; Battaglia, E.; Netter, P.; Richard, T.; Mérillon, J.M.; Magdalou, J.; Sabolovic, N. Regioselective and stereospecific glucuronidation of trans- and cis-resveratrol in human. Arch. Biochem. Biophys. 2001, 393, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Miksits, M.; Maier-Salamon, A.; Aust, S.; Thalhammer, T.; Reznicek, G.; Kunert, O.; Haslinger, E.; Szekeres, T.; Jaeger, W. Sulfation of resveratrol in human liver: Evidence of a major role for the sulfotransferases SULT1A1 and SULT1E1. Xenobiotica Fate Foreign Compd. Biol. Syst. 2005, 35, 1101–1119. [Google Scholar] [CrossRef] [PubMed]
- Lou, B.-S.; Wu, P.-S.; Hou, C.-W.; Cheng, F.-Y.; Chen, J.-K. Simultaneous quantification of trans-resveratrol and its sulfate and glucuronide metabolites in rat tissues by stable isotope-dilution UPLC-MS/MS analysis. J. Pharm. Biomed. Anal. 2014, 94, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Boocock, D.J.; Faust, G.E.S.; Patel, K.R.; Schinas, A.M.; Brown, V.A.; Ducharme, M.P.; Booth, T.D.; Crowell, J.A.; Perloff, M.; Gescher, A.J.; et al. Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Aires, V.; Limagne, E.; Cotte, A.K.; Latruffe, N.; Ghiringhelli, F.; Delmas, D. Resveratrol metabolites inhibit human metastatic colon cancer cells progression and synergize with chemotherapeutic drugs to induce cell death. Mol. Nutr. Food Res. 2013, 57, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.; Schueller, K.; Schaefer, L.-M.; Pignitter, M.; Esefelder, L.; Somoza, V. Resveratrol and its metabolites inhibit pro-inflammatory effects of lipopolysaccharides in U-937 macrophages in plasma-representative concentrations. Food Funct. 2014, 5, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Riha, J.; Brenner, S.; Böhmdorfer, M.; Giessrigl, B.; Pignitter, M.; Schueller, K.; Thalhammer, T.; Stieger, B.; Somoza, V.; Szekeres, T.; et al. Resveratrol and its major sulfated conjugates are substrates of organic anion transporting polypeptides (OATPs): Impact on growth of ZR-75-1 breast cancer cells. Mol. Nutr. Food Res. 2014, 58, 1830–1842. [Google Scholar] [CrossRef] [PubMed]
- Van de Wetering, K.; Burkon, A.; Feddema, W.; Bot, A.; de Jonge, H.; Somoza, V.; Borst, P. Intestinal breast cancer resistance protein (BCRP)/Bcrp1 and multidrug resistance protein 3 (MRP3)/Mrp3 are involved in the pharmacokinetics of resveratrol. Mol. Pharmacol. 2009, 75, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Min, J.S.; Kim, D.; Zheng, Y.F.; Mailar, K.; Choi, W.J.; Lee, C.; Bae, S.K. A simple and sensitive liquid chromatography-tandem mass spectrometry method for trans-ε-viniferin quantification in mouse plasma and its application to a pharmacokinetic study in mice. J. Pharm. Biomed. Anal. 2017, 134, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Mao, P.; Lei, Y.; Zhang, T.; Ma, C.; Jin, B.; Li, T. Pharmacokinetics, bioavailability, metabolism and excretion of δ-viniferin in rats. Acta Pharm. Sin. B 2016, 6, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Willenberg, I.; Brauer, W.; Empl, M.T.; Schebb, N.H. Development of a rapid LC-UV method for the investigation of chemical and metabolic stability of resveratrol oligomers. J. Agric. Food Chem. 2012, 60, 7844–7850. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liao, X.; Xu, J.; Zhao, J.; Luo, J.; Kong, L. Development and validation of a sensitive and selective LC-MS/MS method for the determination of trans δ-veniferin, a resveratrol dehydrodimer, in rat plasma and its application to pharmacokinetics and bioavailability studies. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 958, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Burkon, A.; Somoza, V. Quantification of free and protein-bound trans-resveratrol metabolites and identification of trans-resveratrol-C/O-conjugated diglucuronides—Two novel resveratrol metabolites in human plasma. Mol. Nutr. Food Res. 2008, 52, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Miksits, M.; Maier-Salamon, A.; Vo, T.P.N.; Sulyok, M.; Schuhmacher, R.; Szekeres, T.; Jäger, W. Glucuronidation of piceatannol by human liver microsomes: Major role of UGT1A1, UGT1A8 and UGT1A10. J. Pharm. Pharmacol. 2010, 62, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Miksits, M.; Sulyok, M.; Schuhmacher, R.; Szekeres, T.; Jäger, W. In Vitro sulfation of piceatannol by human liver cytosol and recombinant sulfotransferases. J. Pharm. Pharmacol. 2009, 61, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Maier-Salamon, A.; Böhmdorfer, M.; Thalhammer, T.; Szekeres, T.; Jaeger, W. Hepatic glucuronidation of resveratrol: Interspecies comparison of enzyme kinetic profiles in human, mouse, rat, and dog. Drug Metab. Pharmacokinet. 2011, 26, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Shin, Y.G.; Chow, A.; Li, Y.; Kosmeder, J.W.; Lee, Y.S.; Hirschelman, W.H.; Pezzuto, J.M.; Mehta, R.G.; van Breemen, R.B. Human, rat, and mouse metabolism of resveratrol. Pharm. Res. 2002, 19, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Willenberg, I.; Michael, M.; Wonik, J.; Bartel, L.C.; Empl, M.T.; Schebb, N.H. Investigation of the absorption of resveratrol oligomers in the Caco-2 cellular model of intestinal absorption. Food Chem. 2015, 167, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Waffo-Teguo, P.; Lee, D.; Cuendet, M.; Mérillon, J.; Pezzuto, J.M.; Kinghorn, A.D. Two new stilbene dimer glucosides from grape (Vitis vinifera) cell cultures. J. Nat. Prod. 2001, 64, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y.; Namao, K.; Kamijou, A.; Matsuoka, S.; Nakano, M.; Terao, K.; Ohizumi, Y. Powerful hepatoprotective and hepatotoxic plant oligostilbenes, isolated from the Oriental medicinal plant Vitis coignetiae (Vitaceae). Experientia 1995, 51, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Colin, D.; Lancon, A.; Delmas, D.; Lizard, G.; Abrossinow, J.; Kahn, E.; Jannin, B.; Latruffe, N. Antiproliferative activities of resveratrol and related compounds in human hepatocyte derived HepG2 cells are associated with biochemical cell disturbance revealed by fluorescence analyses. Biochimie 2008, 90, 1674–1684. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.-B.; Han, X.-Z.; Lou, H.-X. Oligomers of resveratrol and ferulic acid prepared by peroxidase-catalyzed oxidation and their protective effects on cardiac injury. J. Agric. Food Chem. 2007, 55, 7753–7757. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, J.; Park, E.-J.; Kondratyuk, T.P.; Marler, L.; Pezzuto, J.M.; van Breemen, R.B.; Mo, S.; Li, Y.; Cushman, M. Selective synthesis and biological evaluation of sulfate-conjugated resveratrol metabolites. J. Med. Chem. 2010, 53, 5033–5043. [Google Scholar] [CrossRef] [PubMed]
- Calamini, B.; Ratia, K.; Malkowski, M.G.; Cuendet, M.; Pezzuto, J.M.; Santarsiero, B.D.; Mesecar, A.D. Pleiotropic mechanisms facilitated by resveratrol and its metabolites. Biochem. J. 2010, 429, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Mikulski, D.; Molski, M. Quantitative structure-antioxidant activity relationship of trans-resveratrol oligomers, trans-4,4′-dihydroxystilbene dimer, trans-resveratrol-3-O-glucuronide, glucosides: Trans-piceid, cis-piceid, trans-astringin and trans-resveratrol-4′-O-beta-d-glucopyranoside. Eur. J. Med. Chem. 2010, 45, 2366–2380. [Google Scholar] [PubMed]
- Patel, K.R.; Andreadi, C.; Britton, R.G.; Horner-Glister, E.; Karmokar, A.; Sale, S.; Brown, V.A.; Brenner, D.E.; Singh, R.; Steward, W.P.; et al. Sulfate metabolites provide an intracellular pool for resveratrol generation and induce autophagy with senescence. Sci. Transl. Med. 2013, 5, 205ra133. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not Available. |




{kind=link}
{kind=link}
{kind=link}
| n° | ε-Viniferin | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|---|
| 2a/6a | 7.17 d(8) | 7.17 d(8) | 7.18 d(8) | 7.26 d(8) | 7.18 d(8) | 7.17 d(8) | 7.17 d(8) | 7.17 brs | 7.17 d(8) |
| 3a/5a | 6.73 d(8) | 6.74 d(8) | 6.74 d(8) | 6.98 d(8) | 6.77 d(8) | 6.73 d(8) | 6.73 d(8) | 7.17 brs | 6.73 d(8) |
| 7a | 5.42 d(5) | 5.45 d(5) | 5.46 d(5) | 5.42 d(5) | 5.47 d(5) | 5.44 d(5) | 5.46 d(5) | 5.46 d(5) | 5.46 d(5) |
| 8a | 4.47 d(5) | 4.52 d(5) | 4.48 d(5) | 4.49 d(5) | 4.54 d(5) | 4.51 d(5) | 4.51 d(5) | 4.48 d(5) | 4.50 d(5) |
| 10a | 6.24 brs | 6.23 brs | 6.25 d(2) | 6.24 brs | 6.59 brs | 6.25 brs | 6.26 d(2) | 6.26 brs | 6.69 t(2) (1) |
| 12a | 6.24 brs | 6.23 brs | 6.24 t(2) | 6.24 brs | 6.50 brs | 6.25 brs | 6.24 t(2) | 6.26 brs | 6.32 t(2) (1) |
| 14a | 6.24 brs | 6.23 brs | 6.25 d(2) | 6.24 Brs | 6.35 brs | 6.25 brs | 6.26 d(2) | 6.26 brs | 6.89 t(2) (1) |
| 2b/6b | 7.20 d(8) | 7.20 d(8) | 7.30 d(8) | 7.21 d(8) | 7.21 d(8) | 7.21 d(8) | 7.29 d(8) | 7.21 d(8) | 7.21 d(8) |
| 3b/5b | 6.83 d(8) | 6.83 d(8) | 7.09 d(8) | 6.83 d(8) | 6.84 d(8) | 6.83 d(8) | 7.24 d(8) | 6.83 d(8) | 6.83 d(8) |
| 7b | 6.90 d(16) | 7.00 d(16) | 6.90 d(16) | 6.94 d(16) | 6.91 d(16) | 6.93 d(16) | 6.88 d(16) | 6.91 d(16) | 6.88 d(16) |
| 8b | 6.71 d(16) | 6.73 d(16) | 6.71 d(16) | 6.78 d(16) | 6.71 d(16) | 6.71 d(16) | 6.71 d(16) | 6.77 d(16) | 6.70 d(16) |
| 12b | 6.32 d(2) | 6.58 d(2) | 6.34 d(2) | 6.34 d(2) | 6.35 d(2) | 6.87 brs | 6.34 d(2) | 6.34 d(2) | 6.33 d(2) |
| 14b | 6.72 d(2) | 6.99 d(2) | 6.72 d(2) | 6.73 d(2) | 6.72 d(2) | 7.11 brs | 6.71 d(2) | 6.73 d(2) | 6.71 d(2) |
| 1′ | 5.18 d(7) | 5.13 d(7) | 5.09 d(7) | 5.10 d(7) | |||||
| 2′ | 3.56 t(8) | 3.53 t(8) | 3.51 t(8) | 3.51 t(8) | |||||
| 3′ | 3.64 t(9) | 3.59 t(9) | 3.58 t(9) | 3.60 t(9) | |||||
| 4′ | 3.73 t(9) | 3.69 t(9) | 3.70 t(10) | 3.72 t(9) | |||||
| 5′ | 4.18 d(10) | 4.11 d(10) | 4.11 d(10) | 4.10 d(10) |
| (a) | Species | Glucuronides | Km | Vmax | Vmax/Km | ||
| Human | MG1 | 3.03 ± 3.00 | 11.27 ± 1.30 | 3.72 |  | 4.98 | |
| MG2 | 4.20 ± 1.88 | 2.54 ± 0.16 | 0.60 | ||||
| MG3 | 2.78 ± 2.00 | 0.78 ± 0.06 | 0.28 | ||||
| MG4 | 13.18 ± 9.39 | 4.91 ± 0.78 | 0.37 | ||||
| Rat | MG1 | 32.94 ± 16.94 | 548.90 ± 86.17 | 16.66 |  | 20.08 | |
| MG2 | 9.09 ± 4.26 | 12.49 ± 1.13 | 1.37 | ||||
| MG3 | 8.52 ± 2.10 | 3.06 ± 0.14 | 0.36 | ||||
| MG4 | 26.14 ± 11.55 | 43.99 ± 5.49 | 1.68 | ||||
| (b) | Species | Sulfates | Km | Vmax | Vmax/Km | ||
| Human | MS1 | 66.67 ± 6.20 | 361.60 ± 13.09 | 5.42 |  | 6.35 | |
| MS2 | 33.01 ± 3.43 | 30.46 ± 0.96 | 0.92 | ||||
| Rat | MS1 | 34.93 ± 11.33 | 25.16 ± 2.29 | 0.72 |  | 2.59 | |
| MS2 | 27.24 ± 11.15 | 8.46 ± 0.89 | 0.31 | ||||
| MS3 | 13.23 ± 5.28 | 13.47 ± 1.10 | 1.02 | ||||
| MS4 | 68.60 ± 20.35 | 36.79 ± 3.94 | 0.54 | ||||
| Human Liver | Rat Liver | |||
|---|---|---|---|---|
| ε-viniferin conversion (%) | 72.7 ± 0.7 | 78.1 ± 0.9 | ||
| Glucuronidation (%) | 29.1 ± 1.2 | MG1: 54.6 ± 2.4 | 73.8 ± 1.5 | MG1: 91.1 ± 2.4 |
| MG2: 5.0 ± 1.2 | MG2: 1.1 ± 0.2 | |||
| MG3: 8.9 ± 0.5 | MG3: 0.7 ± 0.0 | |||
| MG4: 31.5 ± 0.9 | MG4: 7.1 ± 0.1 | |||
| Sulfatation (%) | 43.6 ± 1.8 | MS1: 75.2 ± 3.8 | 4.3 ± 0.3 | MS1: 38.0 ± 1.4 |
| MS2: 24.8 ± 1.2 | MS2: 23.5 ± 3.1 | |||
| MS3: 0 | MS3: 22.0 ± 1.9 | |||
| MS4: 0 | MS4: 16.4 ± 1.3 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Courtois, A.; Jourdes, M.; Dupin, A.; Lapèze, C.; Renouf, E.; Biais, B.; Teissedre, P.-L.; Mérillon, J.-M.; Richard, T.; Krisa, S. In Vitro Glucuronidation and Sulfation of ε-Viniferin, a Resveratrol Dimer, in Humans and Rats. Molecules 2017, 22, 733. https://doi.org/10.3390/molecules22050733
Courtois A, Jourdes M, Dupin A, Lapèze C, Renouf E, Biais B, Teissedre P-L, Mérillon J-M, Richard T, Krisa S. In Vitro Glucuronidation and Sulfation of ε-Viniferin, a Resveratrol Dimer, in Humans and Rats. Molecules. 2017; 22(5):733. https://doi.org/10.3390/molecules22050733
Chicago/Turabian StyleCourtois, Arnaud, Michael Jourdes, Adeline Dupin, Caroline Lapèze, Elodie Renouf, Benoît Biais, Pierre-Louis Teissedre, Jean-Michel Mérillon, Tristan Richard, and Stéphanie Krisa. 2017. "In Vitro Glucuronidation and Sulfation of ε-Viniferin, a Resveratrol Dimer, in Humans and Rats" Molecules 22, no. 5: 733. https://doi.org/10.3390/molecules22050733
APA StyleCourtois, A., Jourdes, M., Dupin, A., Lapèze, C., Renouf, E., Biais, B., Teissedre, P.-L., Mérillon, J.-M., Richard, T., & Krisa, S. (2017). In Vitro Glucuronidation and Sulfation of ε-Viniferin, a Resveratrol Dimer, in Humans and Rats. Molecules, 22(5), 733. https://doi.org/10.3390/molecules22050733