
The Antidiabetic Mechanisms of Polyphenols Related to Increased Glucagon-Like Peptide-1 (GLP1) and Insulin Signaling

 ,
, 
 and
and
Abstract
:1. Introduction
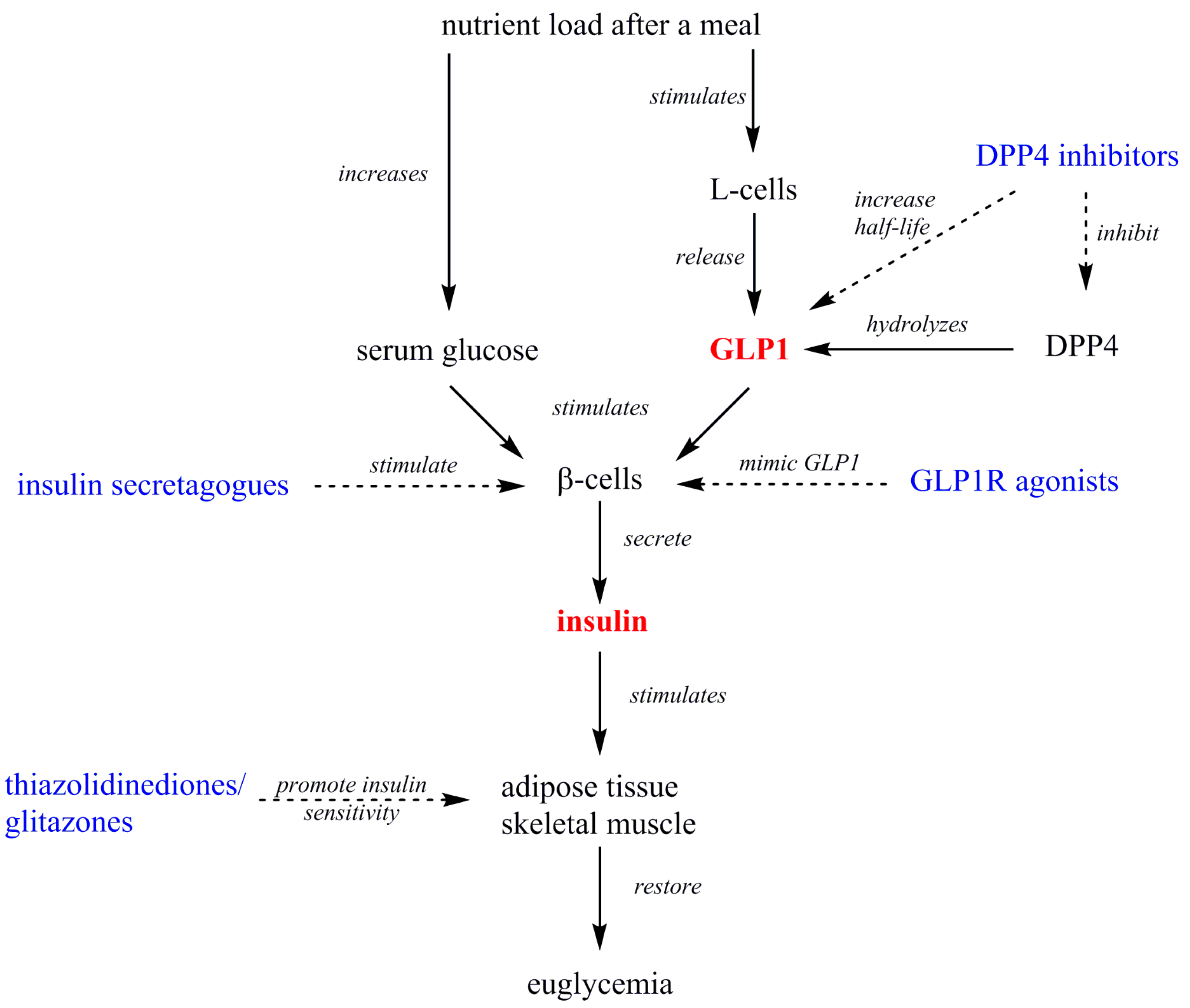
2. GLP1 and Insulin Signaling
2.1. GLP1
2.2. Insulin
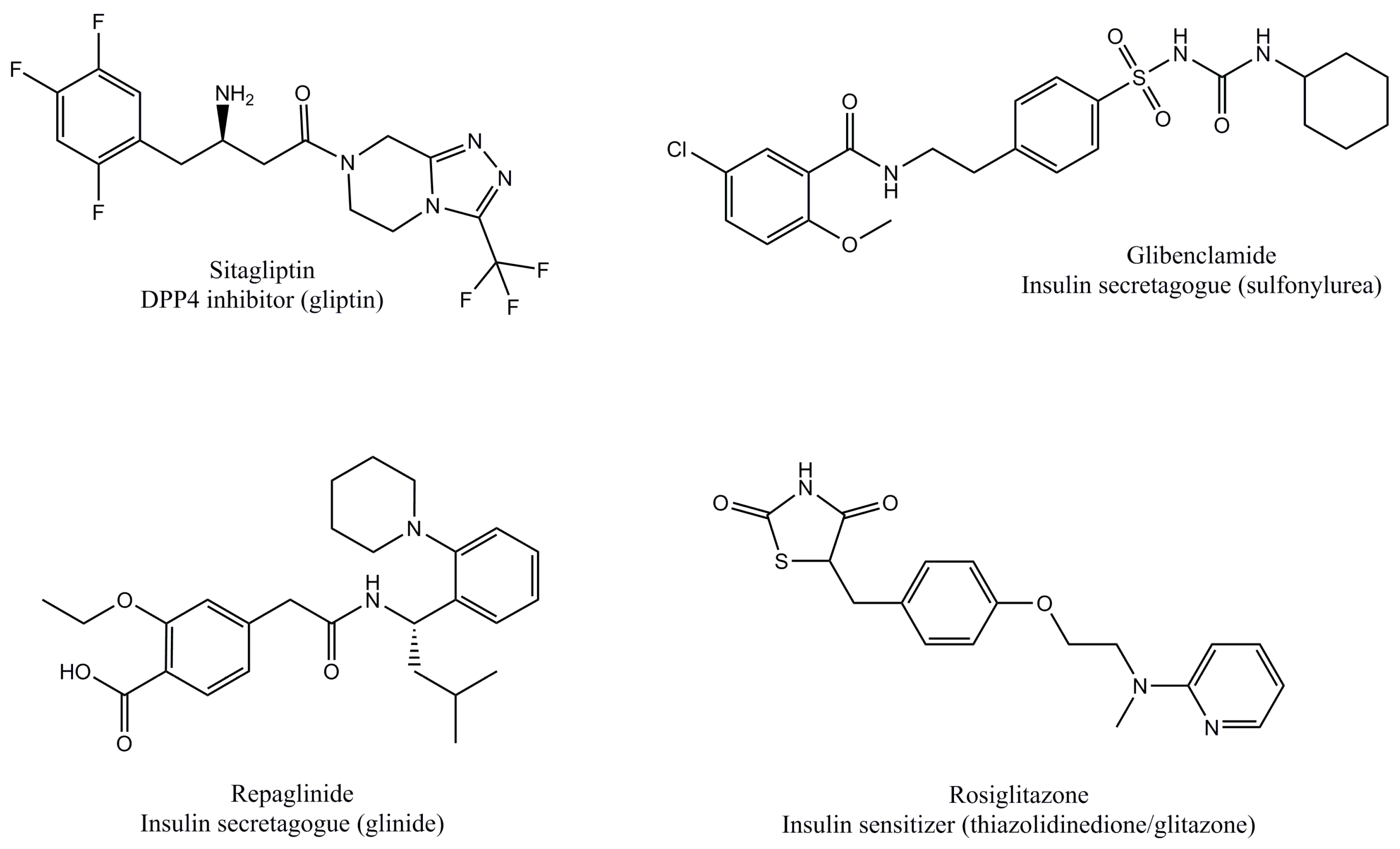
2.3. Current Antidiabetics that Target Insulin or GLP1
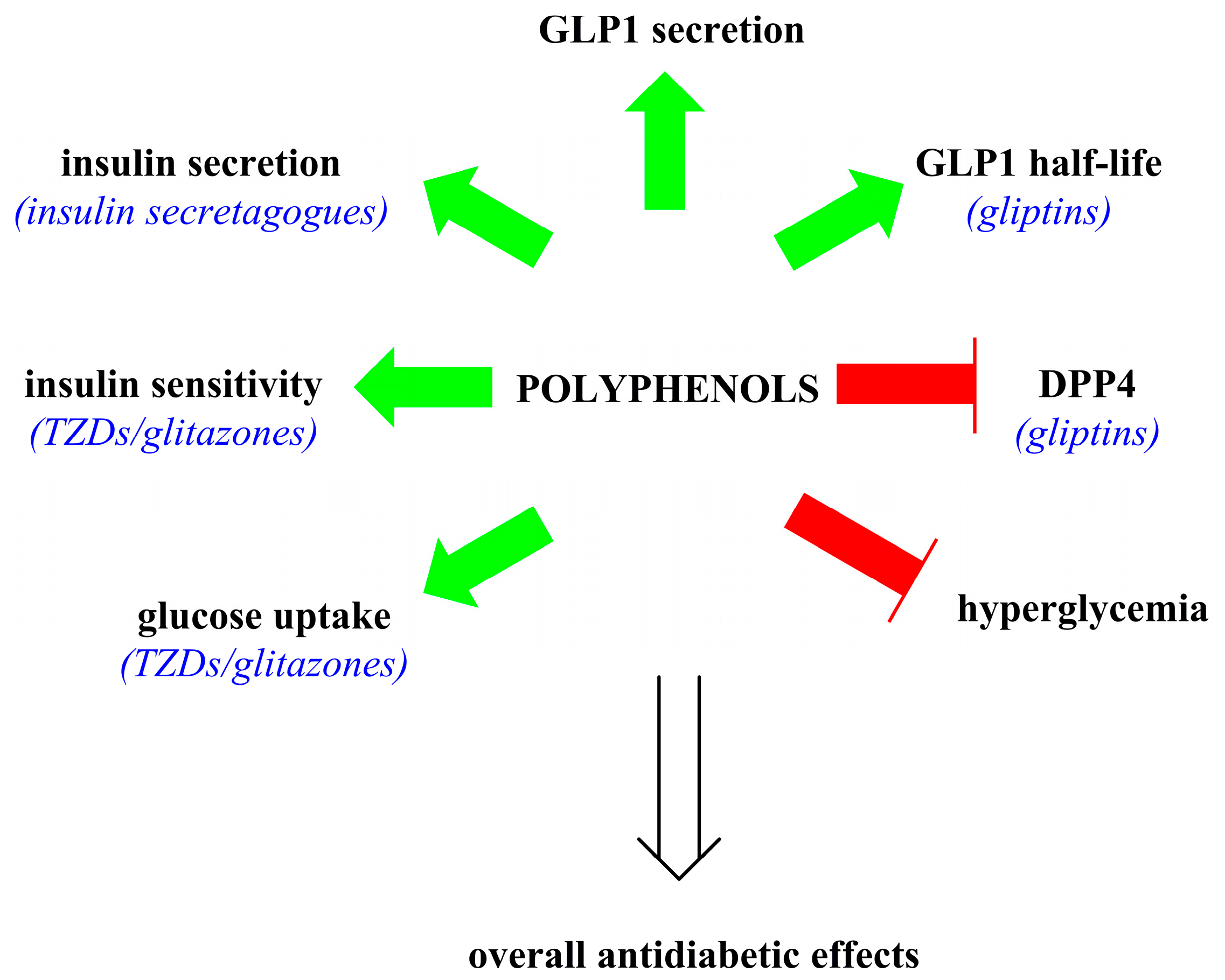
3. Effects of Polyphenols on GLP1 and Insulin Signaling
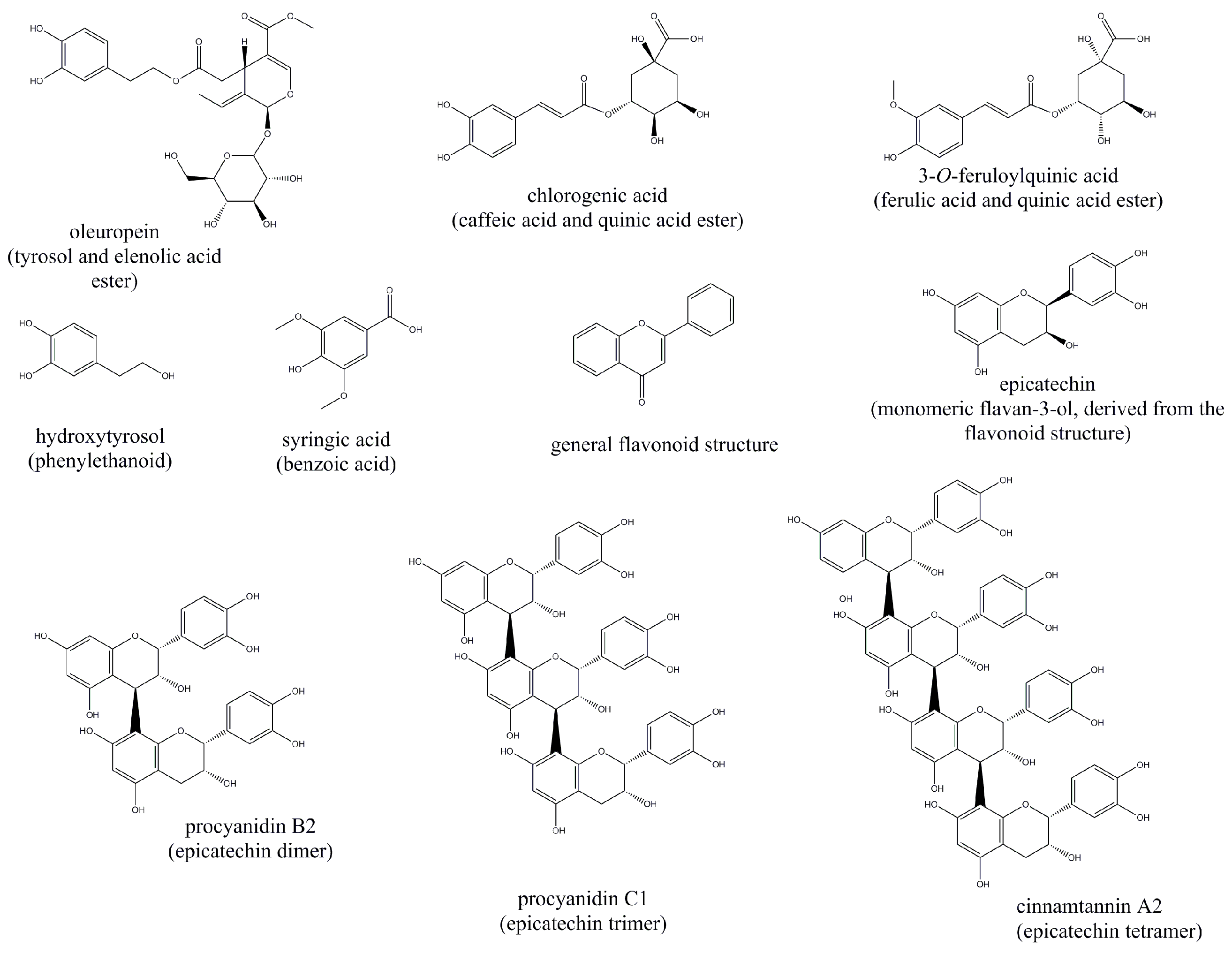
3.1. Effects of Polyphenols on GLP1
3.2. Effects of Polyphenols on Insulin
3.3. Targeted Application of Polyphenolics as Antidiabetics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Association, A.A.D. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2014, 37, S81–S90. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Whiting, D.R.; Guariguata, L.; Weil, C.; Shaw, J. IDF Diabetes Atlas: Global estimates of the prevalence of diabetes for 2011 and 2030. Diabetes Res. Clin. Pract. 2011, 94, 311–321. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. W.H.O. Global Report On Diabetes. Available online: apps.who.int/iris/bitstream/10665/204871/1/9789241565257_eng.pdf (accessed on 11 January 2017).
- Seuring, T.; Archangelidi, O.; Suhrcke, M. The Economic Costs of Type 2 Diabetes: A Global Systematic Review. Pharmacoeconomics 2015, 33, 811–831. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.I.G. Textbook of Diabetes, 4th ed.; Wiley-Blackwell: Chichester, West Sussex, UK; Hoboken, NJ, USA, 2010; p. 1119. [Google Scholar]
- Goran, M.I.; Ulijaszek, S.J.; Ventura, E.E. High fructose corn syrup and diabetes prevalence: A global perspective. Glob. Public Health 2013, 8, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Koloverou, E.; Esposito, K.; Giugliano, D.; Panagiotakos, D. The effect of Mediterranean diet on the development of type 2 diabetes mellitus: A meta-analysis of 10 prospective studies and 136,846 participants. Metabolism 2014, 63, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Kallscheuer, N.; Vogt, M.; Marienhagen, J. A Novel Synthetic Pathway Enables Microbial Production of Polyphenols Independent from the Endogenous Aromatic Amino Acid Metabolism. ACS Synth. Biol. 2017, 6, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effect-A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Nyambe-Silavwe, H.; Williamson, G. Polyphenol- and fibre-rich dried fruits with green tea attenuate starch-derived postprandial blood glucose and insulin: A randomised, controlled, single-blind, cross-over intervention. Br. J. Nutr. 2016, 116, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.L.; Foegeding, E.A.; Harris, G.K. Cocoa and Whey Protein Differentially Affect Markers of Lipid and Glucose Metabolism and Satiety. J. Med. Food 2016, 19, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Grzegorczyk-Karolak, I.; Golab, K.; Gburek, J.; Wysokinska, H.; Matkowski, A. Inhibition of Advanced Glycation End-Product Formation and Antioxidant Activity by Extracts and Polyphenols from Scutellaria alpina L. and S. altissima L. Molecules 2016, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, Physiology, and Mechanisms of Action of Dipeptidyl Peptidase-4 Inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, J.H.; Cui, J.Q.; Chen, W.; Guo, H.; Barbetti, F.; Arvan, P. INS-gene mutations: From genetics and beta cell biology to clinical disease. Mol. Asp. Med. 2015, 42, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wright, J.; Guo, H.; Xiong, Y.; Arvan, P. Proinsulin Entry and Transit through the Endoplasmic Reticulum in Pancreatic Beta Cells. Vitam. Horm. 2014, 95, 35–62. [Google Scholar] [PubMed]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin Receptor Signaling in Normal and Insulin-Resistant States. CSH Perspect. Biol. 2014, 6, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Ahren, B. DPP-4 inhibitors. Best Pract. Res. Clin. Endocrinol. 2007, 21, 517–533. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K. Dipeptidyl peptidase-4 inhibitors: Novel mechanism of actions. Indian J. Endocr. Metab. 2014, 18, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.J. GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Natr. Rev. Endocrinol. 2012, 8, 728–742. [Google Scholar] [CrossRef] [PubMed]
- Abdelmoneim, A.S.; Hasenbank, S.E.; Seubert, J.M.; Brocks, D.R.; Light, P.E.; Simpson, S.H. Variations in tissue selectivity amongst insulin secretagogues: A systematic review. Diabetes Obes. Metab. 2012, 14, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Hu, C.; Jia, W.P. Pharmacogenomics of glinides. Pharmacogenomics 2015, 16, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Seaquist, E.R.; Anderson, J.; Childs, B.; Cryer, P.; Dagogo-Jack, S.; Fish, L.; Heller, S.R.; Rodriguez, H.; Rosenzweig, J.; Vigersky, R. Hypoglycemia and Diabetes: A Report of a Workgroup of the American Diabetes Association and the Endocrine Society. Diabetes Care 2013, 36, 1384–1395. [Google Scholar] [CrossRef] [PubMed]
- Janani, C.; Ranjitha Kumari, B.D. PPAR gamma gene—A review. Diabetes Metab. Syndr. 2015, 9, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, R.; Ueno, S.; Tsubata, M.; Yamaguchi, K.; Takagaki, K.; Hira, T.; Hara, H.; Tsuda, T. Dietary sweet potato (Ipomoea batatas L.) leaf extract attenuates hyperglycaemia by enhancing the secretion of glucagon-like peptide-1 (GLP-1). Food Funct. 2014, 5, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Osaki, N.; Hase, T.; Shimotoyodome, A. Ingestion of coffee polyphenols increases postprandial release of the active glucagon-like peptide-1 (GLP-1(7–36)) amide in C57BL/6J mice. J. Nutr. Sci. 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Thom, E. The effect of chlorogenic acid enriched coffee on glucose absorption in healthy volunteers and its effect on body mass when used long-term in overweight and obese people. J. Int. Med. Res. 2007, 35, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.M.; Khoo, W.; Stewart, A.C.; O’Keefe, S.F.; Lambert, J.D.; Neilson, A.P. Flavanol concentrations do not predict dipeptidyl peptidase-IV inhibitory activities of four cocoas with different processing histories. Food Funct. 2017, 8, 746–756. [Google Scholar] [CrossRef] [PubMed]
- González-Abuín, N.; Martinez-Micaelo, N.; Margalef, M.; Blay, M.; Arola-Arnal, A.; Muguerza, B.; Ardevol, A.; Pinent, M. A grape seed extract increases active glucagon-like peptide-1 levels after an oral glucose load in rats. Food Funct. 2014, 5, 2357–2364. [Google Scholar] [CrossRef] [PubMed]
- González-Abuín, N.; Martinez-Micaelo, N.; Blay, M.; Green, B.D.; Pinent, M.; Ardevol, A. Grape-seed procyanidins modulate cellular membrane potential and nutrient-induced GLP-1 secretion in STC-1 cells. Am. J. Physiol. Cell Physiol. 2014, 306, C485–C492. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Casanova-Marti, A.; Gil-Cardoso, K.; Blay, M.T.; Terra, X.; Pinent, M.; Ardevol, A. Acutely administered grape-seed proanthocyanidin extract acts as a satiating agent. Food Funct. 2016, 7, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Sayago-Ayerdi, S.G.; Velazquez-Lopez, C.; Montalvo-Gonzalez, E.; Goni, I. By-product from decoction process of Hibiscus sabdariffa L. calyces as a source of polyphenols and dietary fiber. J. Sci. Food Agric. 2014, 94, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.H.; Yang, Y.S.; Chan, K.C.; Wang, C.J.; Chen, M.L.; Huang, C.N. Hibiscus sabdariffa Polyphenols Alleviate Insulin Resistance and Renal Epithelial to Mesenchymal Transition: A Novel Action Mechanism Mediated by Type 4 Dipeptidyl Peptidase. J. Agric. Food Chem. 2014, 62, 9736–9743. [Google Scholar] [CrossRef] [PubMed]
- Stefoska-Needham, A.; Beck, E.J.; Johnson, S.K.; Chu, J.Y.; Tapsell, L.C. Flaked sorghum biscuits increase postprandial GLP-1 and GIP levels and extend subjective satiety in healthy subjects. Mol. Nutr. Food Res. 2016, 60, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.L.; Dawson, H.D.; Schoene, N.W.; Polansky, M.M.; Anderson, R.A. Cinnamon polyphenols regulate multiple metabolic pathways involved in insulin signaling and intestinal lipoprotein metabolism of small intestinal enterocytes. Nutrition 2012, 28, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Okabe, M.; Natsume, M.; Ashida, H. Cinnamtannin A2, a Tetrameric Procyanidin, Increases GLP-1 and Insulin Secretion in Mice. Biosci. Biotechnol. Biochem. 2013, 77, 888–891. [Google Scholar] [CrossRef] [PubMed]
- Dorenkott, M.R.; Griffin, L.E.; Goodrich, K.M.; Thompson-Witrick, K.A.; Fundaro, G.; Ye, L.Y.; Stevens, J.R.; Ali, M.; O’Keefe, S.F.; Hulver, M.W.; et al. Oligomeric Cocoa Procyanidins Possess Enhanced Bioactivity Compared to Monomeric and Polymeric Cocoa Procyanidins for Preventing the Development of Obesity, Insulin Resistance, and Impaired Glucose Tolerance during High-Fat Feeding. J. Agric. Food Chem. 2014, 62, 2216–2227. [Google Scholar] [CrossRef] [PubMed]
- Muthukumaran, J.; Srinivasan, S.; Venkatesan, R.S.; Ramachandran, V.; Muruganathan, U. Syringic acid, a novel natural phenolic acid, normalizes hyperglycemia with special reference to glycoprotein components in experimental diabetic rats. J. Acute Dis. 2013, 2, 304–309. [Google Scholar] [CrossRef]
- Evans, S.F.; Meister, M.; Mahmood, M.; Eldoumi, H.; Peterson, S.; Perkins-Veazie, P.; Clarke, S.L.; Payton, M.; Smith, B.J.; Lucas, E.A. Mango supplementation improves blood glucose in obese individuals. Nutr. Metab. Insights 2014, 7, 77–84. [Google Scholar] [PubMed]
- Manzano, M.; Giron, M.D.; Vilchez, J.D.; Sevillano, N.; El-Azem, N.; Rueda, R.; Salto, R.; Lopez-Pedrosa, J.M. Apple polyphenol extract improves insulin sensitivity in vitro and in vivo in animal models of insulin resistance. Nutr. Metab. 2016, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Törrönen, R.; Sarkkinen, E.; Niskanen, T.; Tapola, N.; Kilpi, K.; Niskanen, L. Postprandial glucose, insulin and glucagon-like peptide 1 responses to sucrose ingested with berries in healthy subjects. Br. J. Nutr. 2012, 107, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- De Bock, M.; Derraik, J.G.B.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) Leaf Polyphenols Improve Insulin Sensitivity in Middle-Aged Overweight Men: A Randomized, Placebo-Controlled, Crossover Trial. PLoS ONE 2013, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing—Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Bozzetto, L.; Annuzzi, G.; Pacini, G.; Costabile, G.; Vetrani, C.; Vitale, M.; Griffo, E.; Giacco, A.; De Natale, C.; Cocozza, S.; et al. Polyphenol-rich diets improve glucose metabolism in people at high cardiometabolic risk: a controlled randomised intervention trial. Diabetologia 2015, 58, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Satija, A.; Bhupathiraju, S.N.; Rimm, E.B.; Spiegelman, D.; Chiuve, S.E.; Borgi, L.; Willett, W.C.; Manson, J.E.; Sun, Q.; Hu, F.B. Plant-Based Dietary Patterns and Incidence of Type 2 Diabetes in US Men and Women: Results from Three Prospective Cohort Studies. PLoS Med. 2016, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Tresserra-Rimbau, A.; Guasch-Ferre, M.; Salas-Salvado, J.; Toledo, E.; Corella, D.; Castaner, O.; Guo, X.H.; Gomez-Gracia, E.; Lapetra, J.; Aros, F.; et al. Intake of Total Polyphenols and Some Classes of Polyphenols Is Inversely Associated with Diabetes in Elderly People at High Cardiovascular Disease Risk. J. Nutr. 2016, 146, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.S.; Hassan, M.A.; Kondo, T. Effect of lyophilized grapefruit juice on P-glycoprotein-mediated drug transport in-vitro and in-vivo. Drug Dev. Ind. Pharm. 2015, 41, 375–381. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Mechanism of Action | Main Examples (International Non-Proprietary Name) | Reference |
|---|---|---|---|
| DPP4 inhibitors (gliptins) | Prevent GLP1 hydrolysis | Sitagliptin, vildagliptin, saxagliptin, linagliptin | [21,22] |
| GLP1R agonists (incretin mimetics) | Mimic GLP1 signaling | Exenatide, liraglutide, lixisenatide, albiglutide, dulaglutide | [23] |
| Insulin secretagogues (sulfonylureas) | Stimulate insulin secretion | Glibenclamide, glimepiride | [24,25] |
| Insulin secretagogues (glinides) | Repaglinide, nateglinide, mitiglinide | ||
| Insulin sensitizers (thiazolidinediones/glitazones) | Increase insulin sensitivity | Rosiglitazone, pioglitazone | [27] |
| Polyphenol Source | Model | Effect on GLP1 | Effect on DPP4 | Effect on Insulin | Reference |
|---|---|---|---|---|---|
| Sweet potato ethanolic extract | Murine GLUTag cells | ↑ secretion | [28] | ||
| Sweet potato ethanolic extract | Sprague–Dawley rats | ↑ secretion | [28] | ||
| Coffee extract | Human NCI-H716 cells | Dose-related ↑ secretion | [29] | ||
| Coffee extract | C57BL/6J mice | ↑ secretion | [29] | ||
| Cocoa products | Inhibit | [31] | |||
| Grape seed extract | ↑ secretion | Inhibit | ↑ secretion | ||
| Grape seed extract | STC-1 cells | ↓ secretion | [33] | ||
| Grape seed extract | Wistar rats | ↑ secretion | ↑ secretion | [34] | |
| Hibiscus sabdariffa | HK-2 cells | Inhibit | [36] | ||
| Hibiscus sabdariffa | Sprague–Dawley rats | Inhibit | [36] | ||
| Sorghum | Healthy adults | ↑ GLP1 area under the curve | [37] | ||
| Cranberry extract | C57BL/6J mice | ↓ fasting insulinemia | [38] | ||
| Cinnamon extract | Wistar rat enterocytes | ↑ sensitivity | [40] | ||
| Procyanidin oligomers from cacao liquor | ICR mice | ↑ secretion | ↑ secretion | [41] | |
| Cocoa flavanols | C57BL/6J mice | ↓ fasting insulinemia | [42] | ||
| Syringic acid | Diabetic Wistar rats | Normalized glycemia and insulinemia | [43] | ||
| Mango | Overweight adults | ↑ insulinemia in males | [44] | ||
| Apple polyphenol extract | Obese Zucker rats | No effect on postprandial insulinemia (acute effect) | [45] | ||
| Apple polyphenol extract | Obese Zucker rats | ↑ postprandial insulin response (chronic effect) | [45] | ||
| Apple polyphenol extract | Murine L6-myocytes | Polyphenol/insulin synergy on glucose uptake | [45] | ||
| Blackcurrants, bilberries, cranberries, and strawberries | Healthy adults | Prevented insulin and glucose peaks | [46] | ||
| Olive leaf extract | Overweight men | ↑ sensitivity | [47] | ||
| Dietary intervention with polyphenol-rich foods | Overweight adults | ↑ secretion and sensitivity | [49] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez Avila, J.A.; Rodrigo García, J.; González Aguilar, G.A.; De la Rosa, L.A. The Antidiabetic Mechanisms of Polyphenols Related to Increased Glucagon-Like Peptide-1 (GLP1) and Insulin Signaling. Molecules 2017, 22, 903. https://doi.org/10.3390/molecules22060903
Domínguez Avila JA, Rodrigo García J, González Aguilar GA, De la Rosa LA. The Antidiabetic Mechanisms of Polyphenols Related to Increased Glucagon-Like Peptide-1 (GLP1) and Insulin Signaling. Molecules. 2017; 22(6):903. https://doi.org/10.3390/molecules22060903
Chicago/Turabian StyleDomínguez Avila, J. Abraham, Joaquín Rodrigo García, Gustavo A. González Aguilar, and Laura A. De la Rosa. 2017. "The Antidiabetic Mechanisms of Polyphenols Related to Increased Glucagon-Like Peptide-1 (GLP1) and Insulin Signaling" Molecules 22, no. 6: 903. https://doi.org/10.3390/molecules22060903
APA StyleDomínguez Avila, J. A., Rodrigo García, J., González Aguilar, G. A., & De la Rosa, L. A. (2017). The Antidiabetic Mechanisms of Polyphenols Related to Increased Glucagon-Like Peptide-1 (GLP1) and Insulin Signaling. Molecules, 22(6), 903. https://doi.org/10.3390/molecules22060903