
Himatanthus drasticus Leaves: Chemical Characterization and Evaluation of Their Antimicrobial, Antibiofilm, Antiproliferative Activities


 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Evaluation
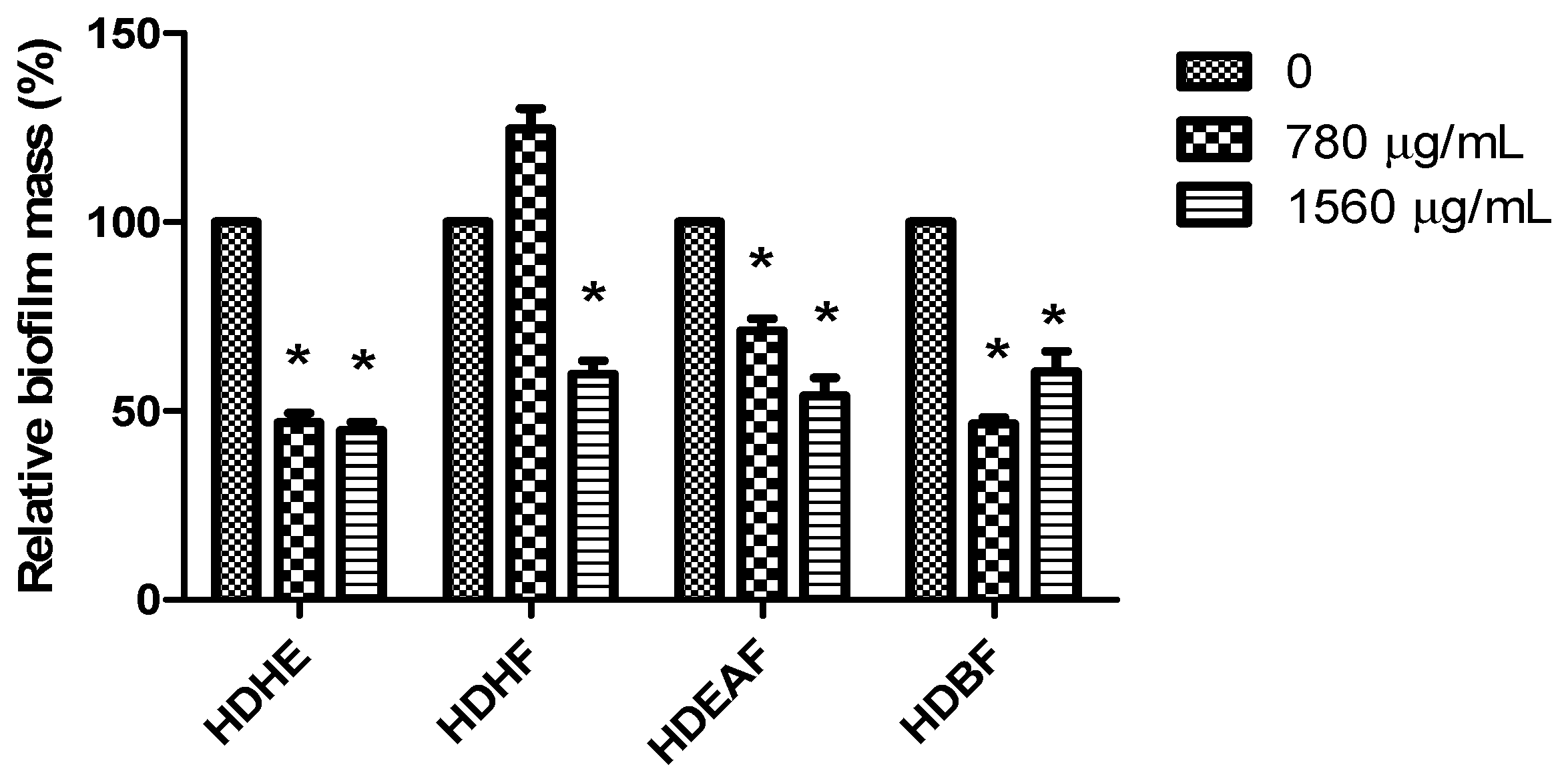
2.2. Antibiofilm Evaluation
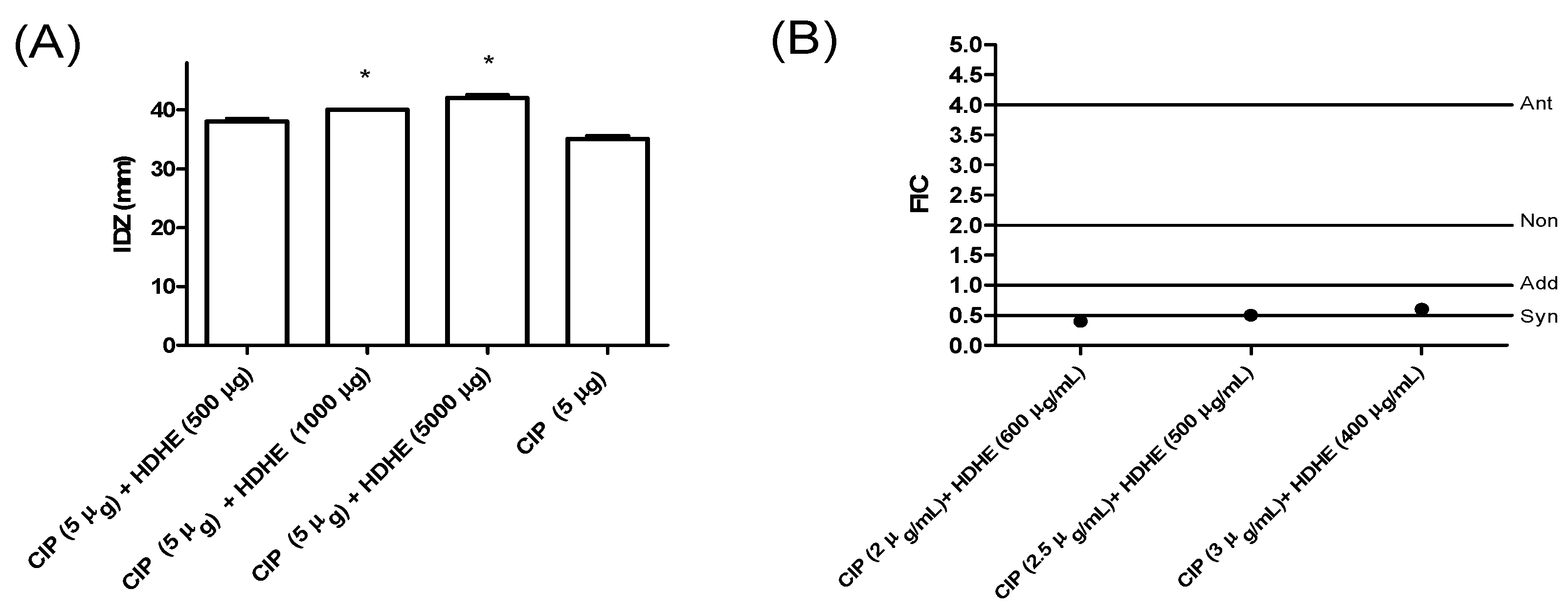
2.3. Combinatory Effects of Himatanthus drasticus Hydroalcoholic Extract and Ciprofloxacin against Klebsiella pneumoniae
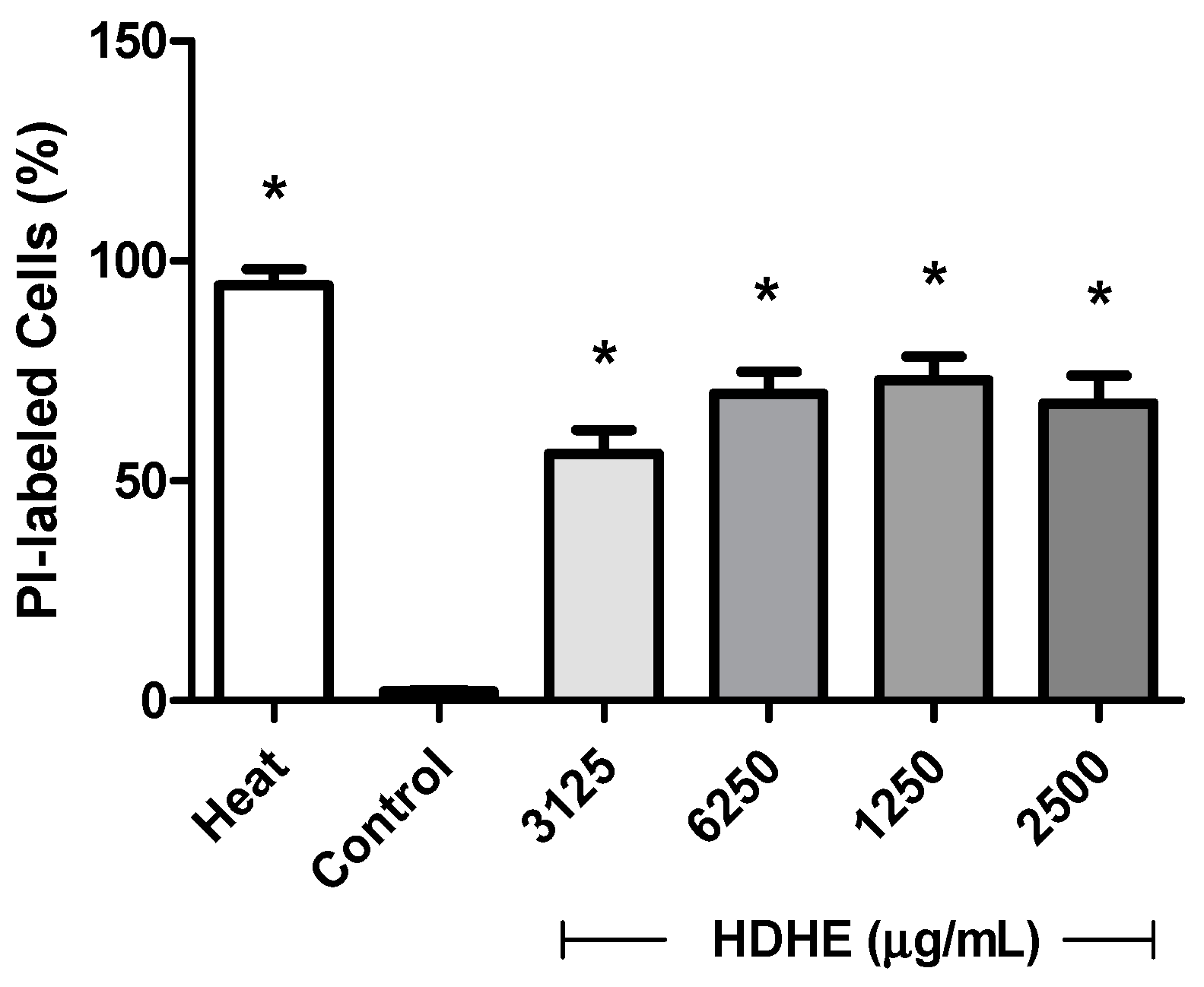
2.4. Effect of Hydroalcoholic Extract of Himatanthus drasticus on the Integrity of the Bacterial Membrane
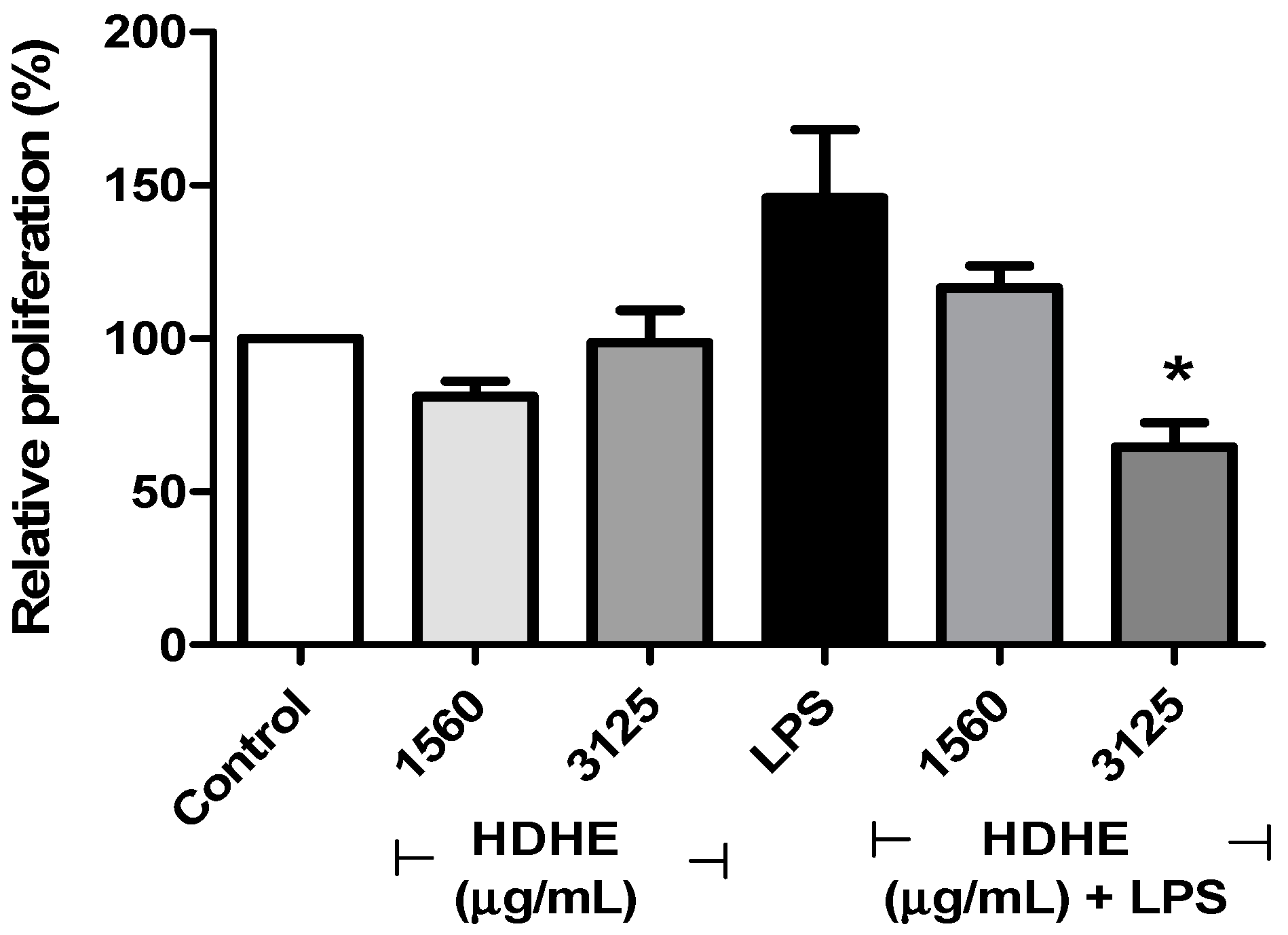
2.5. Effect of Himatanthus drasticus Hydroalcoholic Extract on Cell Viability and LPS-Induced Proliferation of PBMCs
2.6. Chemical Characterization of Himatanthus drasticus Hydroalcoholic Extract
3. Discussion
4. Material and Methods
4.1. Botanical Material
4.2. Preparation of Hydroalcoholic Extract and Its Fractions
4.3. Antimicrobial Assays
4.3.1. Test Microorganisms
4.3.2. Antimicrobial Agar Diffusion Assay
4.3.3. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
4.3.4. Biofilm Assay
4.3.5. Combined Effects of HDHE and Ciprofloxacin
| FIC ≤ 0.5: synergistic effect | |
| 0.5 < FIC ≤ 1: additive effect | |
| 1.0 < FIC ≤ 4: no interaction | |
| FIC > 4: antagonism |
4.3.6. Evaluation of Membrane Integrity
4.4. Assays Using Human Blood
4.4.1. Hemolysis Assay
4.4.2. Cell Proliferation Assay with PBMC
4.5. Chemical Characterization of HDHE
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ribeiro, P.C.; Monteiro, A.S.; Marques, S.G.; Monteiro, S.G.; Monteiro-Neto, V.; Coqueiro, M.M.; Marques, A.C.; de Jesus Gomes Turri, R.; Santos, S.G.; Bomfim, M.R. Phenotypic and molecular detection of the bla KPC gene in clinical isolates from inpatients at hospitals in Sao Luis, MA, Brazil. BMC Infect. Dis. 2016, 16, 737. [Google Scholar] [CrossRef] [PubMed]
- Rossi Goncalves, I.; Ferreira, M.L.; Araujo, B.F.; Campos, P.A.; Royer, S.; Batistao, D.W.; Souza, L.P.; Brito, C.S.; Urzedo, J.E.; Gontijo-Filho, P.P.; et al. Outbreaks of colistin-resistant and colistin-susceptible KPC-producing Klebsiella pneumoniae in a Brazilian intensive care unit. J. Hosp. Infect. 2016, 94, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Tognim, M.C.; Cardoso, C.L. Nosocomial outbreaks in Brazil: Can they be controlled? J. Hosp. Infect. 2016, 94, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Raja, B.; Goux, H.J.; Marapadaga, A.; Rajagopalan, S.; Kourentzi, K.; Willson, R.C. Development of a panel of recombinase polymerase amplification assays for detection of common bacterial urinary tract infection pathogens. J. Appl. Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.G.; Kang, D.H. Inactivation of Escherichia coli O157:H7, Salmonella Typhimurium, and Listeria monocytogenes in ready-to-bake cookie dough by gamma and electron beam irradiation. Food Microbiol. 2017, 64, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rojas, A.; Makarova, O.; Rolff, J. Antimicrobials, stress and mutagenesis. PLoS Pathog. 2014, 10, e1004445. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.S. The contribution of capsule polysaccharide genes to virulence of Klebsiella pneumoniae. Virulence 2016. [Google Scholar] [CrossRef] [PubMed]
- Mandrawa, C.L.; Cronin, K.; Buising, K.L.; Poy Lorenzo, Y.S.; Waters, M.J.; Jeremiah, C.J. Carbapenemase-producing Klebsiella pneumoniae: A major clinical challenge. Med. J. Aust. 2016, 204, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Prokesch, B.C.; TeKippe, M.; Kim, J.; Raj, P.; TeKippe, E.M.; Greenberg, D.E. Primary osteomyelitis caused by hypervirulent Klebsiella pneumoniae. Lancet Infect. Dis. 2016, 16, e190–e195. [Google Scholar] [CrossRef]
- Wu, H.; Li, D.; Zhou, H.; Sun, Y.; Guo, L.; Shen, D. Bacteremia and other body site infection caused by hypervirulent and classic Klebsiella pneumoniae. Microb. Pathog. 2017, 104, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Yu, J.L.; Ai, Q.; Liu, D.; Lu, W.; Lu, Q.; Peng, N.N. [Diversity analysis of biofilm bacteria on tracheal tubes removed from intubated neonates]. Zhonghua Er Ke Za Zhi = Chin. J. Pediatrics 2013, 51, 602–606. [Google Scholar]
- Kishibe, S.; Okubo, Y.; Morino, S.; Hirotaki, S.; Tame, T.; Aoki, K.; Ishii, Y.; Ota, N.; Shimomura, S.; Sakakibara, H.; et al. Pediatric hypervirulent Klebsiella pneumoniae septic arthritis. Pediatr. Int. 2016, 58, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Lagrafeuille, R.; Souweine, B.; Forestier, C. Anti-biofilm activity as a health issue. Front. Microbiol. 2016, 7, 592. [Google Scholar] [CrossRef] [PubMed]
- Hoiby, N.; Ciofu, O.; Johansen, H.K.; Song, Z.J.; Moser, C.; Jensen, P.O.; Molin, S.; Givskov, M.; Tolker-Nielsen, T.; Bjarnsholt, T. The clinical impact of bacterial biofilms. Int. J. Oral Sci. 2011, 3, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Biswas, N.; Datta, A.; Dey, R.; Maiti, P. Atomic force microscopy in biofilm study. Microscopy 2014, 63, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Ghafourian, S. Mutation and efflux system are responsible for ciprofloxacin resistance in Klebsiella pneumoniae. West Indian Med. J. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Yang, H.S.; Nam, Y.S.; Lee, H.J. Mutations in DNA gyrase and topoisomerase IV in ciprofloxacin-nonsusceptible extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Clin. Lab. 2017, 63, 535–541. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Y.; Wang, X.; Ding, F.; Fu, Y.; Zhao, J.; Song, W.; Opiyo, O.J.; Zhang, F.; Chen, X. Siderophores in clinical isolates of Klebsiella pneumoniae promote ciprofloxacin resistance by inhibiting the oxidative stress. Biochem. Bioph. Res. Commun. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lagerback, P.; Khine, W.W.; Giske, C.G.; Tangden, T. Evaluation of antibacterial activities of colistin, rifampicin and meropenem combinations against NDM-1-producing Klebsiella pneumoniae in 24 h in vitro time-kill experiments. J. Antimicrob. Chemother. 2016, 71, 2321–2325. [Google Scholar] [CrossRef] [PubMed]
- Mezzatesta, M.L.; Caio, C.; Gona, F.; Zingali, T.; Salerno, I.; Stefani, S. Colistin increases the cidal activity of antibiotic combinations against multidrug-resistant Klebsiella pneumoniae: An in vitro model comparing multiple combination bactericidal testing at one peak serum concentration and time-kill method. Microb. Drug Resist. 2016, 22, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Bezerra Dos Santos, A.T.; Araujo, T.F.; Nascimento da Silva, L.C.; da Silva, C.B.; de Oliveira, A.F.; Araujo, J.M.; Correia, M.T.; Lima, V.L. Organic extracts from Indigofera suffruticosa leaves have antimicrobial and synergic actions with erythromycin against Staphylococcus aureus. Front. Microbiol. 2015, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti Filho, J.R.; Silva, T.F.; Nobre, W.Q.; Oliveira de Souza, L.I.; Silva E Silva Figueiredo, C.S.; Figueiredo, R.C.; de Gusmao, N.B.; Silva, M.V.; Nascimento da Silva, L.C.; Correia, M.T. Antimicrobial activity of Buchenavia tetraphylla against Candida albicans strains isolated from vaginal secretions. Pharm. Biol. 2017, 55, 1521–1527. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.P.; da Nascimento Silva, L.C.; da Martins Fonseca, C.S.; de Araujo, J.M.; Correia, M.T.; Cavalcanti Mda, S.; Lima, V.L. Antimicrobial activity and phytochemical analysis of organic extracts from Cleome spinosa Jaqc. Front. Microbiol. 2016, 7, 963. [Google Scholar] [CrossRef] [PubMed]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.R.; Mecsas, J.; Moir, D.T. Beyond antibiotics: New therapeutic approaches for bacterial infections. Clin. Infect. Dis. 2016, 63, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Fura, J.M.; Sarkar, S.; Pidgeon, S.E.; Pires, M.M. Combatting Bacterial Pathogens with Immunomodulation and Infection Tolerance Strategies. Curr. Top. Med. Chem. 2017, 17, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.C.; Correia, M.T. Plant lectins and Toll-like receptors: Implications for therapy of microbial infections. Front. Microbiol. 2014, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.C.; da Silva, M.V.; Correia, M.T. Editorial: New frontiers in the search of antimicrobials agents from natural products. Front. Microbiol. 2017, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, B.S.; da Silva, L.C.; da Silva, T.D.; Rodrigues, J.F.; Grisotto, M.A.; Correia, M.T.; Napoleao, T.H.; da Silva, M.V.; Paiva, P.M. Application of omics technologies for evaluation of antibacterial mechanisms of action of plant-derived products. Front. Microbiol. 2016, 7, 1466. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Patridge, E.; Gareiss, P.; Kinch, M.S.; Hoyer, D. An analysis of FDA-approved drugs: Natural products and their derivatives. Drug Discov. Today 2016, 21, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Soares, F.P.; Cavalcante, L.F.; Romero, N.R.; Bandeira, M.A. Himatanthus Willd. ex Schult. (Apocynaceae): Review. Pharmacogn. Rev. 2016, 10, 6–10. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho Nilo Bitu, V.; de Carvalho Nilo Bitu, V.; Matias, E.F.; de Lima, W.P.; da Costa Portelo, A.; Coutinho, H.D.; de Menezes, I.R. Ethnopharmacological study of plants sold for therapeutic purposes in public markets in Northeast Brazil. J. Ethnopharmacol. 2015, 172, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.A.; Oliveira, L.G.; Macedo, D.G.; Menezes, I.R.; Costa, J.G.; Silva, M.A.; Lacerda, S.R.; Souza, M.M. Promising medicinal plants for bioprospection in a cerrado area of Chapada do Araripe, Northeastern Brazil. J. Ethnopharmacol. 2014, 155, 1522–1533. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; O'Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Cuyckens, F.; Claeys, M. Mass spectrometry in the structural analysis of flavonoids. J. Mass Spectrom. 2004, 39, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pankey, G.A.; Sabath, L.D. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of Gram-positive bacterial infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.C.; Sandes, J.M.; de Paiva, M.M.; de Araujo, J.M.; de Figueiredo, R.C.; da Silva, M.V.; Correia, M.T. Anti-Staphylococcus aureus action of three Caatinga fruits evaluated by electron microscopy. Nat. Prod. Res. 2013, 27, 1492–1496. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Lavigne, R. Breaking barriers: Expansion of the use of endolysins as novel antibacterials against Gram-negative bacteria. Future Microbiol. 2015, 10, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, G.V.; Tyulenev, A.V.; Muzyka, N.G.; Peters, M.A.; Oktyabrsky, O.N. Ciprofloxacin provokes SOS-dependent changes in respiration and membrane potential and causes alterations in the redox status of Escherichia coli. Res. Microbiol. 2017, 168, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Ta, C.A.; Arnason, J.T. Mini review of phytochemicals and plant taxa with activity as microbial biofilm and quorum sensing inhibitors. Molecules 2015, 21, E29. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Bruzzone, S.; De Flora, A.; Usai, C.; Graeff, R.; Lee, H.C. Cyclic ADP-ribose is a second messenger in the lipopolysaccharide-stimulated proliferation of human peripheral blood mononuclear cells. Biochem. J. 2003, 375, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Hu, D.; Hou, L.B.; Song, L.Y.; Zhang, Y.J.; Xie, Y.; Tian, L.W. Phenolic compounds from the rhizomes of Smilax china L. and their anti-inflammatory activity. Molecules 2017, 22, 515. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, N.; Sui, Z.; Bi, K.; Li, Z. An investigation on the quantitative structure-activity relationships of the anti-inflammatory activity of diterpenoid alkaloids. Molecules 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Devi, N.S.; Ramanan, M.; Paragi-Vedanthi, P.; Doble, M. Phytochemicals as multi-target inhibitors of the inflammatory pathway-A modeling and experimental study. Biochem. Biol. Res. Commun. 2017, 484, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Gao, H.; Liu, J.X.; Liu, L.; Zhou, H.; Liu, Z.Q. Triterpenoid saponins with anti-inflammatory activities from Ilex pubescens roots. Phytochemistry 2017, 134, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Nijnik, A.; Philpott, D.J. Modulating immunity as a therapy for bacterial infections. Nat. Rev. Microbiol. 2012, 10, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.; Rao, M.; Wallis, R.S.; Kaufmann, S.H.; Rustomjee, R.; Mwaba, P.; Vilaplana, C.; Yeboah-Manu, D.; Chakaya, J.; Ippolito, G.; et al. Host-Directed Therapies Network, c., Host-directed therapies for infectious diseases: Current status, recent progress, and future prospects. Lancet Infect. Dis. 2016, 16, e47–e63. [Google Scholar] [CrossRef]
- Saengsai, J.; Kongtunjanphuk, S.; Yoswatthana, N.; Kummalue, T.; Jiratchariyakul, W. Antibacterial and antiproliferative activities of plumericin, an iridoid isolated from Momordica charantia Vine. Evid. Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef]
- Fakhrudin, N.; Waltenberger, B.; Cabaravdic, M.; Atanasov, A.G.; Malainer, C.; Schachner, D.; Heiss, E.H.; Liu, R.; Noha, S.M.; Grzywacz, A.M.; et al. Identification of plumericin as a potent new inhibitor of the NF-kappaB pathway with anti-inflammatory activity in vitro and in vivo. Br. J. Pharmacol. 2014, 171, 1676–1686. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.H.; Liu, R.; Waltenberger, B.; Khan, S.; Schachner, D.; Kollmann, P.; Zimmermann, K.; Cabaravdic, M.; Uhrin, P.; Stuppner, H.; et al. Plumericin inhibits proliferation of vascular smooth muscle cells by blocking STAT3 signaling via S-glutathionylation. Sci. Rep. 2016, 6, 20771. [Google Scholar] [CrossRef] [PubMed]
- Torella, M.; Del Deo, F.; Grimaldi, A.; Iervolino, S.A.; Pezzella, M.; Tammaro, C.; Gallo, P.; Rappa, C.; De Franciscis, P.; Colacurci, N. Efficacy of an orally administered combination of hyaluronic acid, chondroitin sulfate, curcumin and quercetin for the prevention of recurrent urinary tract infections in postmenopausal women. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 207, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Usman Amin, M.; Khurram, M.; Khan, T.A.; Faidah, H.S.; Ullah Shah, Z.; Ur Rahman, S.; Haseeb, A.; Ilyas, M.; Ullah, N.; Umar Khayam, S.M.; et al. Effects of luteolin and quercetin in combination with some conventional antibiotics against methicillin-resistant Staphylococcus aureus. Int. J. Mol. Sci. 2016, 17, 1947. [Google Scholar] [CrossRef] [PubMed]
- Rajasekharan, S.K.; Ramesh, S.; Satish, A.S.; Lee, J. Antibiofilm and anti-beta-lactamase activities of burdock root extract and chlorogenic acid against Klebsiella pneumoniae. J. Microbiol. Biotechnol. 2017, 27, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, H.; Wang, Y.; Song, F.; Yuan, Y. Inhibitory effects of quercetin on the progression of liver fibrosis through the regulation of NF-small ka, CyrillicB/Ismall ka, CyrillicBalpha, p38 MAPK, and Bcl-2/Bax signaling. Int. Immunopharmacol. 2017, 47, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Haleagrahara, N.; Miranda-Hernandez, S.; Alim, M.A.; Hayes, L.; Bird, G.; Ketheesan, N. Therapeutic effect of quercetin in collagen-induced arthritis. Biomed. Pharmacother. 2017, 90, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.Y.; Jin, J.; Jin, L.W.; Chen, Y.; Zhou, Z.H.; Li, Z.Y. Chlorogenic acid attenuates lipopolysaccharide-induced acute kidney injury by inhibiting TLR4/NF-kappaB signal pathway. Inflammation 2017, 40, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Trentin Dda, S.; Giordani, R.B.; Zimmer, K.R.; da Silva, A.G.; da Silva, M.V.; Correia, M.T.; Baumvol, I.J.; Macedo, A.J. Potential of medicinal plants from the Brazilian semi-arid region (Caatinga) against Staphylococcus epidermidis planktonic and biofilm lifestyles. J. Ethnopharmacol. 2011, 137, 327–335. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of extracts or compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | HDHE | HDHF | HDEAF | HDBF | Ciprofloxacin |
|---|---|---|---|---|---|
| Inhibition Diameter Zone (mm) | |||||
| Staphylococcus aureus | - | - | - | 10 ± 0.5 | 33 ± 0.5 |
| Pseudomonas aeruginosa | - | 20 ± 0.5 | - | 25 ± 0.5 | 43 ± 0.0 |
| Salmonella enterica | - | - | - | - | 35 ± 0.5 |
| Listeria monocytogenes | 12 ± 0.5 | - | 30 ± 0 | - | 36 ± 0.5 |
| Escherichia coli | - | - | - | - | 45 ± 0.5 |
| Klebsiella. pneumoniae | 16 ± 0.5 | - | - | - | 40 ± 1.1 |
| Acinetobacter baumannii | - | - | - | - | 40 ± 0.0 |
| Enterococcus faecalis | - | - | - | - | 28 ± 1.1 |
| Bacterial Species | Header | HDHE | HDHF | HDEAF | HDBF | Ciprofloxacin |
|---|---|---|---|---|---|---|
| Staphylococcus aureus | MIC: | NE | NE | NE | >50,000 | 0.78 |
| MBC: | NE | NE | NE | >50,000 | 0.78 | |
| Pseudomonas aeruginosa | MIC: | NE | >50 | NE | >50,000 | 25 |
| MBC: | NE | >50 | NE | >50,000 | 25 | |
| Listeria monocytogenes | MIC: | 2500 | NE | >50,000 | NE | 1.56 |
| MBC: | 2500 | NE | >50,000 | NE | 1.56 | |
| Klebsiella pneumoniae | MIC: | 6250 | NE | NE | 12,500 | 12.5 |
| MBC: | 6250 | NE | NE | 12,500 | 12.5 |
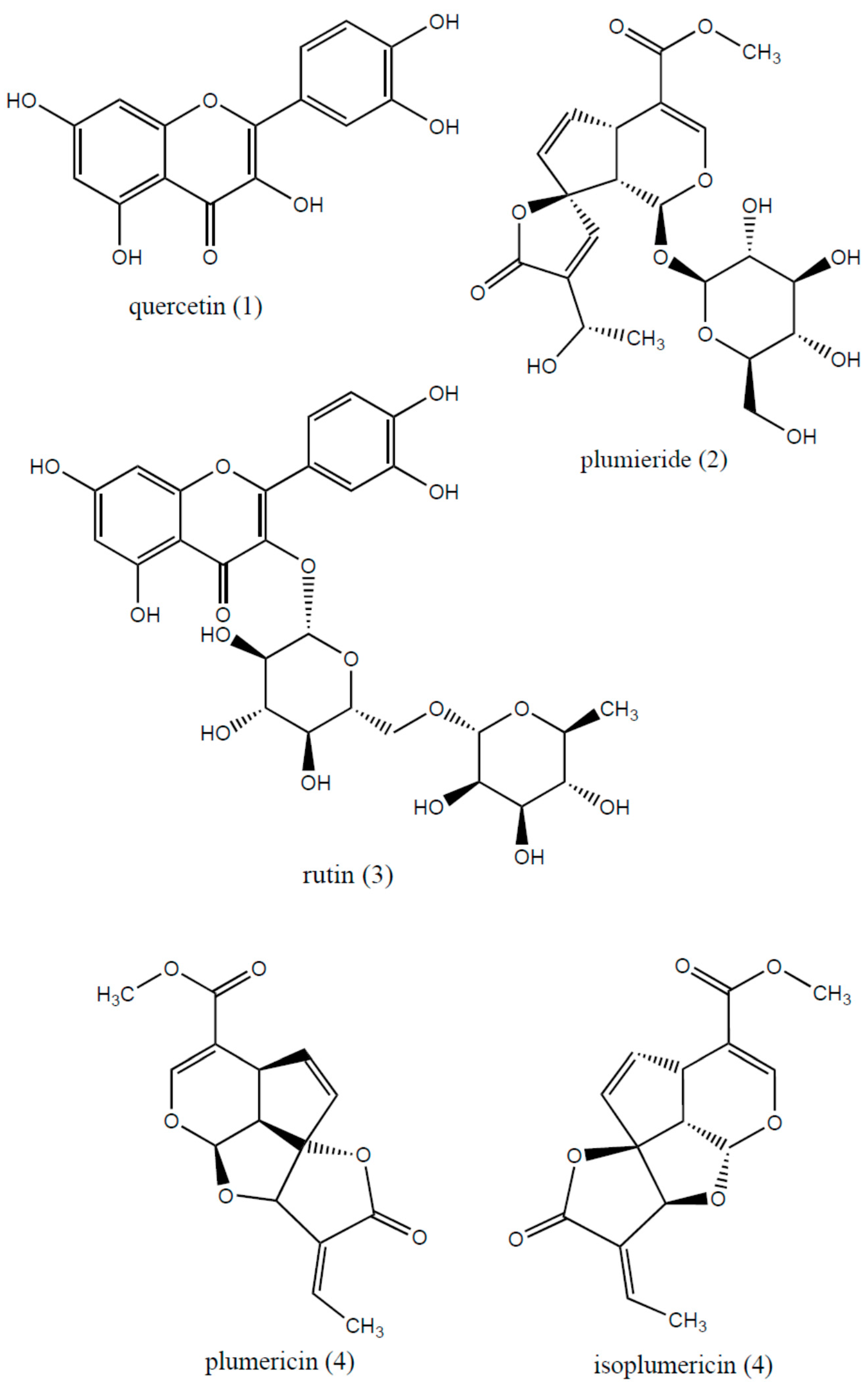
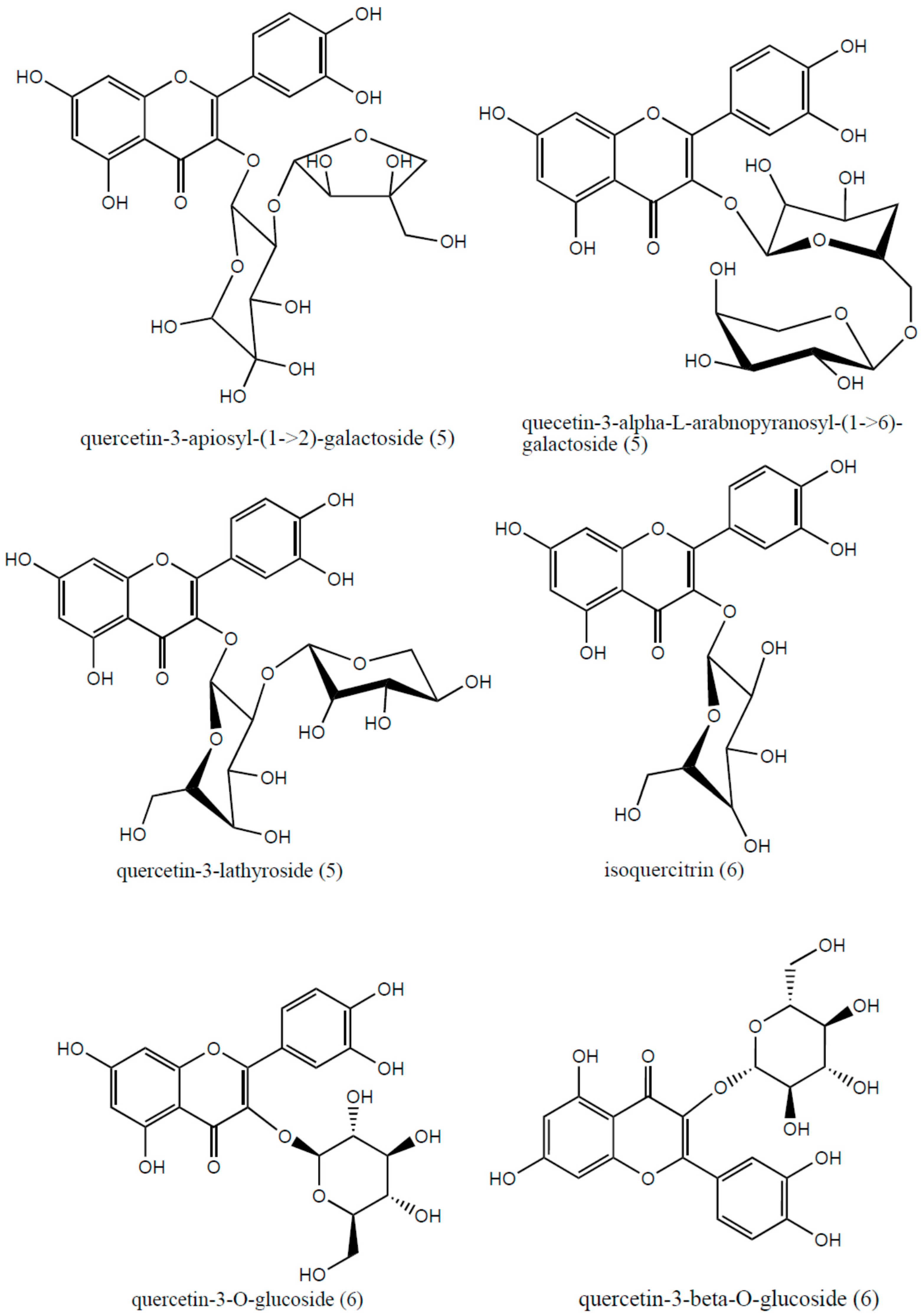
| Compound | Rt (min) | m/z [M + H]+ | λMAX (nm) | MS2 Main Fragments |
|---|---|---|---|---|
| Chlorogenic acid | 11.6 | 355.19 ([M + H]+) | 254 | 162.84 |
| Plumericin or isomers | 13.0 | 291.12 ([M + H]+) | 254, 280 | 263.06, 230.98 |
| Quercetin | 13.5 | 303.11 ([M + H]+) | 254, 280 | 285.26 |
| Quercetin 3-lathyroside or isomers | 13.5 | 597.27 ([M + H]+) | 254, 280 | 465.22, 303.06 |
| Rutin | 14.8 | 611.31 ([M + H]+) | 254, 280 | 465.20, 303.08 |
| Isoquercetrin or isomers | 15.8 | 465.20 ([M + H]+) | 254, 280 | 303.04 |
| Plumieride | 24.4 | 493.41 ([M + Na]+) | 254, 280 | 448.41 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueiredo, C.S.S.e.S.; Branco Santos, J.C.; Castro Junior, J.A.d.A.; Wakui, V.G.; Rodrigues, J.F.S.; Arruda, M.O.; Monteiro, A.D.S.; Monteiro-Neto, V.; Bomfim, M.R.Q.; Kato, L.; et al. Himatanthus drasticus Leaves: Chemical Characterization and Evaluation of Their Antimicrobial, Antibiofilm, Antiproliferative Activities. Molecules 2017, 22, 910. https://doi.org/10.3390/molecules22060910
Figueiredo CSSeS, Branco Santos JC, Castro Junior JAdA, Wakui VG, Rodrigues JFS, Arruda MO, Monteiro ADS, Monteiro-Neto V, Bomfim MRQ, Kato L, et al. Himatanthus drasticus Leaves: Chemical Characterization and Evaluation of Their Antimicrobial, Antibiofilm, Antiproliferative Activities. Molecules. 2017; 22(6):910. https://doi.org/10.3390/molecules22060910
Chicago/Turabian StyleFigueiredo, Cristiane Santos Silva e Silva, Joice Castelo Branco Santos, José Artur de Aguiar Castro Junior, Vinícius Galvão Wakui, João F. S. Rodrigues, Mariana Oliveira Arruda, Andrea De Souza Monteiro, Valério Monteiro-Neto, Maria Rosa Quaresma Bomfim, Lucília Kato, and et al. 2017. "Himatanthus drasticus Leaves: Chemical Characterization and Evaluation of Their Antimicrobial, Antibiofilm, Antiproliferative Activities" Molecules 22, no. 6: 910. https://doi.org/10.3390/molecules22060910
APA StyleFigueiredo, C. S. S. e. S., Branco Santos, J. C., Castro Junior, J. A. d. A., Wakui, V. G., Rodrigues, J. F. S., Arruda, M. O., Monteiro, A. D. S., Monteiro-Neto, V., Bomfim, M. R. Q., Kato, L., Nascimento da Silva, L. C., & Grisotto, M. A. G. (2017). Himatanthus drasticus Leaves: Chemical Characterization and Evaluation of Their Antimicrobial, Antibiofilm, Antiproliferative Activities. Molecules, 22(6), 910. https://doi.org/10.3390/molecules22060910