
Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation



Abstract
:1. Introduction
2. Monosaccharide Composition, Combinations, and Bioactivities of Mushroom Polysaccharides
2.1. Monosaccharide Composition and Combinations
2.2. Bioactivities of Mushroom PSs with Differing Monosaccharide Composition/Combinations
3. Biosynthetic Pathways and Key Enzymes of Mushroom Polysaccharides
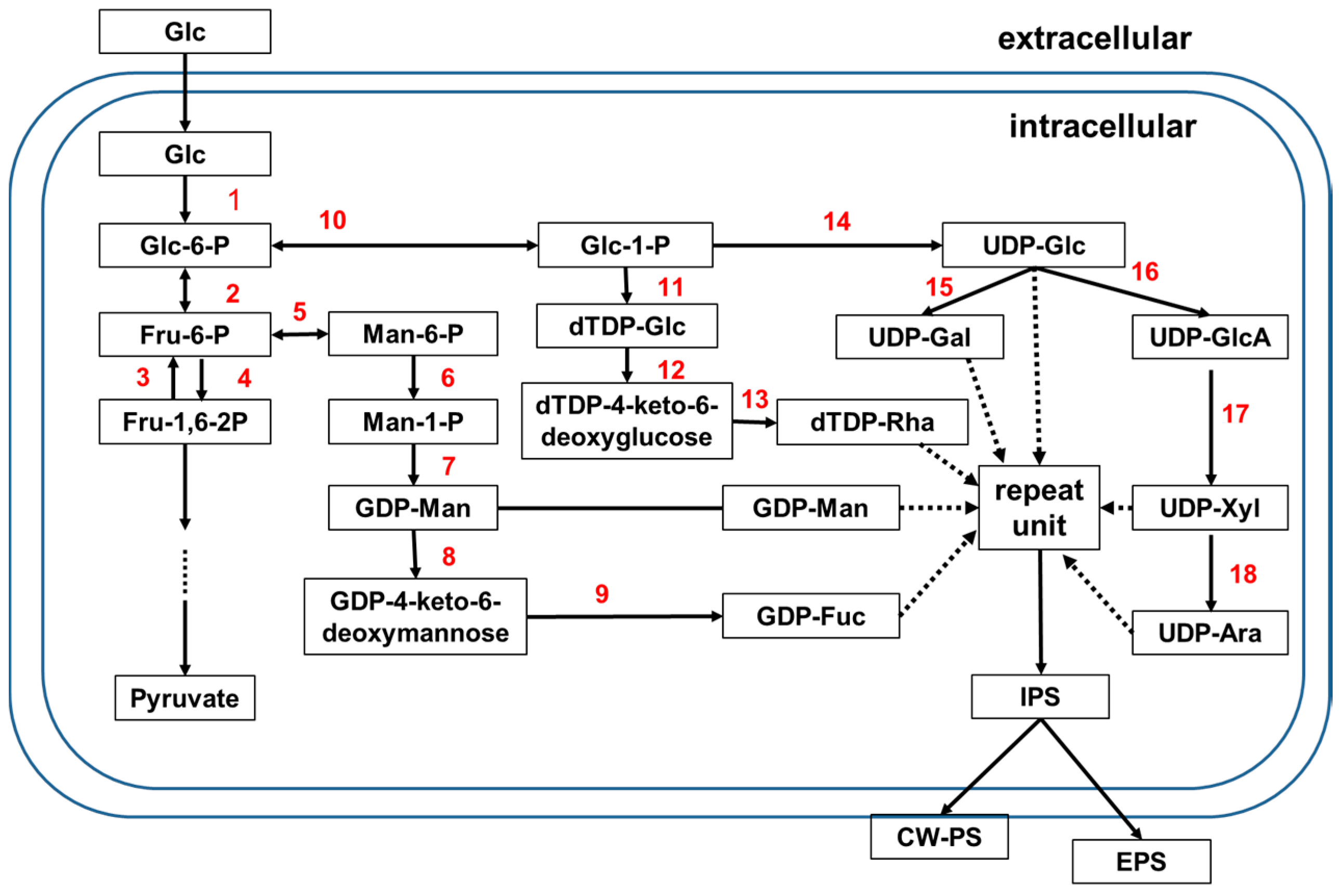
3.1. Biosynthetic Pathways
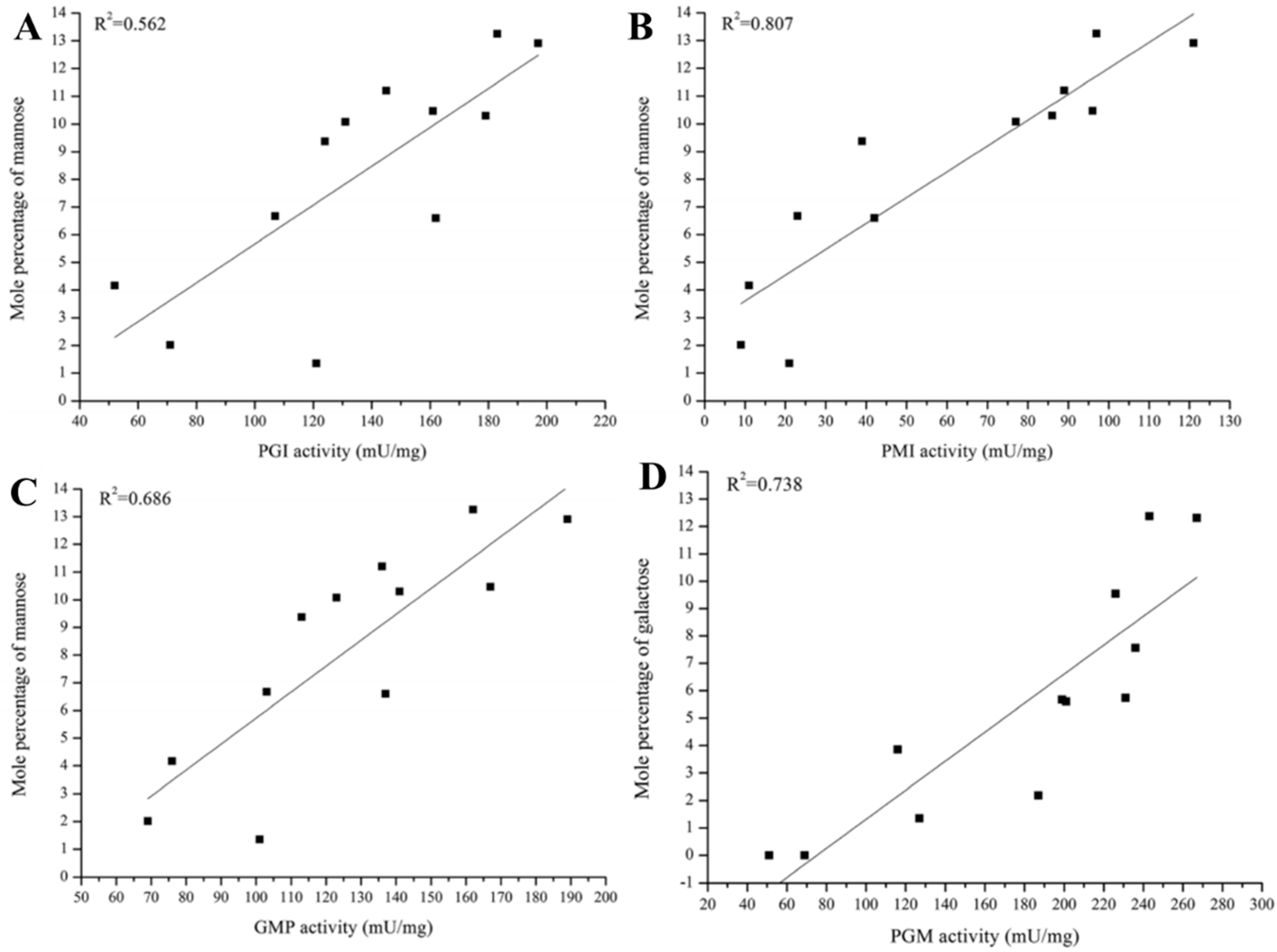
3.2. Effects of Key Enzymes on PS Production and Monosaccharide Composition
4. Factors Regulating Mushroom Polysaccharide Synthesis
5. Conclusions and Future Perspectives
Acknowledgments
Conflicts of Interest
References
- He, X.R.; Wang, X.X.; Fang, J.C.; Chang, Y.; Ning, N.; Guo, H.; Huang, L.H.; Huang, X.Q.; Zhao, Z.F. Polysaccharides in Grifola frondosa mushroom and their health promoting properties: A review. Int. J. Biol. Macromol. 2017, 101, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.F.; Yan, H.D.; Tang, J.; Chen, J.; Zhang, X.W. Polysaccharides in lentinus edodes: Isolation, structure, immunomodulating activity and future prospective. Crit. Rev. Food Sci. Nutr. 2014, 54, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.C.; Heleno, S.A.; Reis, F.S.; Stojkovic, D.; Queiroz, M.J.; Vasconcelos, M.H.; Sokovic, M. Chemical features of Ganoderma polysaccharides with antioxidant, antitumor and antimicrobial activities. Phytochemistry 2015, 114, 38–55. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.K.; Wang, W.Q.; Wu, J.Y. Recent advances in Cordyceps sinensis polysaccharides: Mycelial fermentation, isolation, structure, and bioactivities: A review. J. Funct. Foods 2014, 6, 33–47. [Google Scholar] [CrossRef]
- Ruthes, A.C.; Smiderle, F.R.; Iacomini, M. Mushroom heteropolysaccharides: A review on their sources, structure and biological effects. Carbohydr. Polym. 2016, 136, 358–375. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.T.; Cheng, A.C.; Chiu, K.H.; Tsay, P.K.; Jen, J.F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Li, Z.M.; Nie, K.Y.; Wang, Z.J.; Luo, D.H. Quantitative structure activity relationship models for the antioxidant activity of polysaccharides. PLoS ONE 2016, 11, e0163536. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Hu, Y.; Quan, L.L. Production of bioactive polysaccharides by Inonotus obliquus under submerged fermentation supplemented with lignocellulosic biomass and their antioxidant activity. Bioprocess Biosyst. Eng. 2014, 37, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Zhang, D.H.; Yue, T.H.; Jiang, L.X.; Yu, X.; Zhao, P.; Li, T.; Xu, J.W. Improved polysaccharide production in a submerged culture of Ganoderma lucidum by the heterologous expression of vitreoscilla hemoglobin gene. J. Biotechnol. 2016, 217, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.B.; Bi, P.Y.; Wu, X.T.; Huang, F.R.; Yang, H.L. Improved polysaccharide production in submerged culture of Ganoderma lucidum by the addition of coixenolide. Appl. Biochem. Biotechnol. 2014, 172, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Liu, X.C.; Dong, F.Y.; Guo, M.Z.; Wang, X.T.; Wang, Z.; Zhang, Y.M. Influence of fermentation conditions on polysaccharide production and the activities of enzymes involved in the polysaccharide synthesis of Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 3909–3921. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.H.; Liu, L.; Guo, X.F.; Li, Y.J.; Hou, B.C.; Fan, Q.L.; Wang, K.X.; Luo, Y.; Zhong, J.J. Sucrose fed-batch strategy enhanced biomass, polysaccharide, and ganoderic acids production in fermentation of Ganoderma lucidum 5.26. Bioprocess Biosyst. Eng. 2016, 39, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.L.; Liu, R.; Ren, M.F.; Li, H.J.; Xu, J.W. Enhanced production of polysaccharide through the overexpression of homologous uridine diphosphate glucose pyrophosphorylase gene in a submerged culture of lingzhi or reishi medicinal mushroom, Ganoderma lucidum (higher basidiomycetes). Int. J. Med. Mushrooms 2015, 17, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.W.; Ji, S.L.; Li, H.J.; Zhou, J.S.; Duan, Y.Q.; Dang, L.Z.; Mo, M.H. Increased polysaccharide production and biosynthetic gene expressions in a submerged culture of Ganoderma lucidum by the overexpression of the homologous α-phosphoglucomutase gene. Bioprocess Biosyst. Eng. 2015, 38, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Qiao, S.K.; Xu, Z.H.; Guang, F.; Ding, Z.Y.; Gu, Z.H.; Zhang, L.; Shi, G.Y. Effects of culture conditions on monosaccharide composition of Ganoderma lucidum exopolysaccharide and on activities of related enzymes. Carbohydr. Polym. 2015, 133, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Li, J.; Liu, Y.; Xu, Z.H.; Wu, J.Y.; Ding, Z.Y.; Gu, Z.H.; Zhang, L.; Shi, G.Y. Effects of mixed carbon sources on galactose and mannose content of exopolysaccharides and related enzyme activities in Ganoderma lucidum. RSC Adv. 2016, 6, 39284–39291. [Google Scholar] [CrossRef]
- Giavasis, I. Bioactive fungal polysaccharides as potential functional ingredients in food and nutraceuticals. Curr. Opin. Biotechnol. 2014, 26, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Q.; Nie, S.P.; Kan, L.J.; Chen, H.H.; Cui, S.W.; Phillips, A.O.; Phillips, G.O.; Xie, M.Y. Comparison of structural features and antioxidant activity of polysaccharides from natural and cultured Cordyceps sinensis. Food Sci. Biotechnol. 2017, 26, 55–62. [Google Scholar] [CrossRef]
- Cheong, K.L.; Meng, L.Z.; Chen, X.Q.; Wang, L.Y.; Wu, D.T.; Zhao, J.; Li, S.P. Structural elucidation, chain conformation and immuno-modulatory activity of glucogalactomannan from cultured Cordyceps sinensis fungus UM01. J. Funct. Foods 2016, 25, 174–185. [Google Scholar] [CrossRef]
- Yan, J.K.; Wang, W.Q.; Li, L.; Wu, J.Y. Physiochemical properties and antitumor activities of two α-glucans isolated from hot water and alkaline extracts of Cordyceps (CS-HK1) fungal mycelia. Carbohydr. Polym. 2011, 85, 753–758. [Google Scholar] [CrossRef]
- Wu, Y.L.; Sun, C.R.; Pan, Y.J. Structural analysis of a neutral (1→3), (1→4)-beta-d-glucan from the mycelia of Cordyceps sinensis. J. Nat. Prod. 2005, 68, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Smiderle, F.R.; Sassaki, G.L.; van Griensven, L.J.L.D.; Iacomini, M. Isolation and chemical characterization of a glucogalactomannan of the medicinal mushroom Cordyceps militaris. Carbohydr. Polym. 2013, 97, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.P.; Duan, Y.Q.; Yang, W.Y.; Zhang, H.H.; Li, C.Z.; Zhang, J.X. Structural elucidation and immunostimulatory activity of polysaccharide isolated by subcritical water extraction from Cordyceps militaris. Carbohydr. Polym. 2016, 157, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.C.; Zhu, Z.Y.; Tang, Y.L.; Wang, M.F.; Wang, Z.; Liu, A.J.; Zhang, Y.M. Structural properties of polysaccharides from cultivated fruit bodies and mycelium of Cordyceps militaris. Carbohydr. Polym. 2016, 142, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.K.; Gu, G.F.; Hao, L.M.; Jin, Z.N.; Wang, X. Characterization and in vitro antioxidant activity of a polysaccharide from Cordyceps sobolifera. J. Food Process. Preserv. 2016, 40, 447–452. [Google Scholar] [CrossRef]
- Sharma, S.K.; Gautam, N.; Atri, N.S. Optimized extraction, composition, antioxidant and antimicrobial activities of exo and intracellular polysaccharides from submerged culture of Cordyceps cicadae. BMC Complement. Altern. Med. 2015, 15, 446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yin, Q.; Han, T.; Zhao, Y.; Su, J.; Li, M.; Ling, J. Purification and antioxidant effect of novel fungal polysaccharides from the stroma of Cordyceps kyushuensis. Ind. Crops Prod. 2015, 69, 485–491. [Google Scholar] [CrossRef]
- Liu, C.; Sun, Y.H.; Mao, Q.; Guo, X.L.; Li, P.; Liu, Y.; Xu, N. Characteristics and antitumor activity of Morchella esculenta polysaccharide extracted by pulsed electric field. Int. J. Mol. Sci. 2016, 17, 986. [Google Scholar] [CrossRef] [PubMed]
- Li, S.H.; Gao, A.; Dong, S.; Chen, Y.; Sun, S.; Lei, Z.F.; Zhang, Z.Y. Purification, antitumor and immunomodulatory activity of polysaccharides from soybean residue fermented with Morchella esculenta. Int. J. Biol. Macromol. 2017, 96, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.L.; Chen, Y.; Wang, S.S.; Kai, G.Q.; Fang, Y.M. Isolation, partial characterisation and immunomodulatory activities of polysaccharide from Morchella esculenta. J. Sci. Food Agric. 2011, 91, 2180–2185. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, X.H.; Li, H.M.; Wang, S.H.; Chen, T.; Yuan, Z.P.; Tang, Y.J. Isolation and characterization of polysaccharides with the antitumor activity from Tuber fruiting bodies and fermentation system. Appl. Microbiol. Biotechnol. 2014, 98, 1991–2002. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.J.; Zhang, S.Q.; Ran, C.X.; Wang, L.S.; Kan, J.Q. Extraction, characterization and antioxidant activity of water-soluble polysaccharides from Tuber huidongense. Int. J. Biol. Macromol. 2016, 91, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Pattanayak, M.; Samanta, S.; Maity, P.; Manna, D.K.; Sen, I.K.; Nandi, A.K.; Panda, B.C.; Chattopadhyay, S.; Roy, S.; Sahoo, A.K. Polysaccharide of an edible Truffle tuber rufum: Structural studies and effects on human lymphocytes. Int. J. Biol. Macromol. 2016, 95, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Meng, G.Y.; Zhai, G.Y.; Yang, Y.H.; Zhao, H.J.; Jia, L. Extraction, characteristisation and antioxidant activity of polysaccharides of spent mushroom compost of Ganoderma lucidum. Int. J. Biol. Macromol. 2016, 82, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.W.; Feng, M.Y.; Zhai, X.F.; Hu, M.H.; You, L.J.; Luo, W.; Zhao, M.M. Optimization for the extraction of polysaccharides from Ganoderma lucidum and their antioxidant and antiproliferative activities. J. Taiwan Inst. Chem. Eng. 2013, 44, 886–894. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.Y.; Pang, X.B.; Yao, W.B.; Gao, X.D. Characterization and antioxidant activity of two low-molecular-weight polysaccharides purified from the fruiting bodies of Ganoderma lucidum. Int. J. Biol. Macromol. 2010, 46, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Zhang, J.S.; Tang, Q.J.; Yang, Y.; Guo, Q.B.; Wang, Q.; Wu, D.; Cui, S.W. Physicochemical characterization of a high molecular weight bioactive β-d-glucan from the fruiting bodies of Ganoderma lucidum. Carbohydr. Polym. 2014, 101, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Fang, L.; Zhang, K.C. Structure and bioactivities of a galactose rich extracellular polysaccharide from submergedly cultured Ganoderma lucidum. Carbohydr. Polym. 2007, 68, 323–328. [Google Scholar] [CrossRef]
- Ye, L.B.; Zhang, J.S.; Ye, X.J.; Tang, Q.J.; Liu, Y.F.; Gong, C.Y.; Du, X.J.; Pan, Y.J. Structural elucidation of the polysaccharide moiety of a glycopeptide (GLPCW-II) from Ganoderma lucidum fruiting bodies. Carbohydr. Res. 2008, 343, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Wang, L.Q.; Chen, C.H.; Teng, B.S.; Wang, C.D.; Xu, Z.X.; Hu, B.W.; Zhou, P. Structure characterization of a novel neutral polysaccharide isolated from Ganoderma lucidum fruiting bodies. Food Chem. 2012, 135, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Zhang, Z.Y.; Yang, Y.N. Antioxidant and immunoregulatory activity of Ganoderma lucidum polysaccharide (GLP). Carbohydr. Polym. 2013, 95, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nie, S.P.; Yin, J.Y.; Wang, Y.X.; Xie, M.Y. Structural characterization of a heterogalactan purified from fruiting bodies of Ganoderma atrum. Food Hydrocoll. 2014, 36, 339–347. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W.J.; Nie, S.P.; Chen, Y.; Wang, Y.X.; Xie, M.Y. Structural characterisation of a novel bioactive polysaccharide from Ganoderma atrum. Carbohydr. Polym. 2012, 88, 1047–1054. [Google Scholar] [CrossRef]
- Han, X.Q.; Chan, B.C.L.; Yu, H.; Yang, Y.H.; Hu, S.Q.; Ko, C.H.; Dong, C.X.; Wong, C.K.; Shaw, P.C.; Fung, K.P.; et al. Structural characterization and immuno-modulating activities of a polysaccharide from Ganoderma sinense. Int. J. Biol. Macromol. 2012, 51, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.F.; Zhang, L.N.; Zeng, F.B.; Xu, Y.X. Structure and antitumor activity of extracellular polysaccharides from mycelium. Carbohydr. Polym. 2003, 54, 297–303. [Google Scholar] [CrossRef]
- Wang, C.L.; Pi, C.C.; Kuo, C.W.; Zhuang, Y.J.; Khoo, K.H.; Liu, W.H.; Chen, C.J. Polysaccharides purified from the submerged culture of Ganoderma formosanum stimulate macrophage activation and protect mice against listeria monocytogenes infection. Biotechnol. Lett. 2011, 33, 2271. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.Y.; Hsu, T.H.; Lin, F.Y.; Lai, H.H.; Wu, J.Y. Effects on cytokine-stimulating activities of eps from Tremella mesenterica with various carbon sources. Food Chem. 2006, 99, 92–97. [Google Scholar] [CrossRef]
- Chabot, S.; Yu, H.L.; Léséleuc, L.D.; Cloutier, D.; Calsteren, M.R.V.; Lessard, M.; Roy, D.; Lacroix, M.; Oth, D. Exopolysaccharides from Lactobacillus rhamnosus RW-9595M stimulate TNF, IL-6 and IL-12 in human and mouse cultured immunocompetent cells, and IFN-γ in mouse splenocytes. Dairy Sci. Technol. 2001, 81, 683–697. [Google Scholar] [CrossRef]
- Cescutti, P.; Kallioinen, A.; Impallomeni, G.; Toffanin, R.; Pollesello, P.; Leisola, M.; Eerikäinen, T. Structure of the exopolysaccharide produced by Enterobacter amnigenus. Carbohydr. Res. 2005, 340, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, W.; Rui, X.; Li, T.; Chen, X.H.; Jiang, M.; Dong, M.S. Chemical modification, characterization and bioactivity of a released exopolysaccharide (r-EPS1) from Lactobacillus plantarum 70810. Glycoconj. J. 2015, 32, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Ostrosky-Zeichner, L.; Vitale, R.G.; Nucci, M. New serological markers in medical mycology: (1,3)-β-d-glucan and aspergillus galactomannan. Infectio 2012, 16, 59–63. [Google Scholar] [CrossRef]
- Xiang, Y.L.; Xu, X.Q.; Li, J. Chemical properties and antioxidant activity of exopolysaccharides fractions from mycelial culture of Inonotus obliquus in a ground corn stover medium. Food Chem. 2012, 134, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yan, M.C.; Zhu, J.W.; Xu, X.Q. Enhancement of exo-polysaccharide production and antioxidant activity in submerged cultures of Inonotus obliquus by lignocellulose decomposition. J. Ind. Microbiol. Biotechnol. 2011, 38, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Yong, O.K.; Park, H.W.; Kim, J.H.; Lee, J.Y.; Moon, S.H.; Shin, C.S. Anti-cancer effect and structural characterization of endo-polysaccharide from cultivated mycelia of Inonotus obliquus. Life Sci. 2006, 79, 72–80. [Google Scholar]
- Reshetnikov, S.V.; Wasser, S.P.; Nevo, E.; Duckman, I.; Tsukor, K. Medicinal value of the genus Tremella pers. (heterobasidiomycetes) (review). Int. J. Med. Mushrooms 2000, 2, 26. [Google Scholar] [CrossRef]
- Zhao, M.; Chen, Z.; Xia, G.; Cui, F.Y.; Zhang, J.J.; Jia, M.S.; Jia, S.H.; Jia, L. Enzymatic and acidic degradation effect on intracellular polysaccharide of Flammulina velutipes SF-08. Int. J. Biol. Macromol. 2015, 73, 236–244. [Google Scholar]
- Liu, M.; Jing, H.J.; Zhang, J.J.; Che, G.; Zhou, M.; Gao, Z.; Li, S.S.; Ren, Z.Z.; Hao, L.; Liu, Y. Optimization of mycelia selenium polysaccharide extraction from Agrocybe cylindracea SL-02 and assessment of their antioxidant and anti-ageing activities. PLoS ONE 2016, 11, e0160799. [Google Scholar] [CrossRef] [PubMed]
- Im, S.; Wang, W.; Lee, C.K.; Lee, Y.N. Activation of macrophages by exopolysaccharide produced by MK1 bacterial strain isolated from neungee mushroom, sarcodon aspratus. Immune Netw. 2010, 10, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Sun, S.S.; Li, R.; Shen, Z.P.; Wang, P.; Jiang, X.L. Antioxidant activity of polysaccharides produced by Hirsutella sp. And relation with their chemical characteristics. Carbohydr. Polym. 2015, 117, 452–457. [Google Scholar]
- He, P.X.; Li, F.L.; Huang, L.N.; Xue, D.D.; Liu, W.; Xu, C.P. Chemical characterization and antioxidant activity of polysaccharide extract from spent mushroom substrate of Pleurotus eryngii. J. Taiwan Inst. Chem. Eng. 2016, 69, 48–53. [Google Scholar] [CrossRef]
- Lo, T.C.T.; Yi, H.J.; Chao, A.L.J.; Cheng, A.C. Use of statistical methods to find the polysaccharide structural characteristics and the relationships between monosaccharide composition ratio and macrophage stimulatory activity of regionally different strains of Lentinula edodes. Anal. Chim. Acta 2007, 584, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.W.; Yang, Z.J.; Dan, G.; Fan, J.L.; Dai, Z.Q.; Wang, X.Q.; Bing, H.; Hong, Y.; Abid, M.; Zeng, X.X. Influences of carbon sources on the biomass, production and compositions of exopolysaccharides from Paecilomyces hepiali HN1. Biomass Bioenergy 2014, 67, 260–269. [Google Scholar] [CrossRef]
- Li, M.J.; Chen, T.X.; Gao, T.; Miao, Z.G.; Jiang, A.L.; Shi, L.; Ren, A.; Zhao, M.W. UDP-glucose pyrophosphorylase influences polysaccharide synthesis, cell wall components, and hyphal branching in Ganoderma lucidum via regulation of the balance between glucose-1-phosphate and UDP-glucose. Fungal Genet. Biol. 2015, 82, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Xu, J.; Liu, C.; Zhu, Y.J.; Nelson, D.R.; Zhou, S.G.; Li, C.F.; Wang, L.Z.; Guo, X.; Sun, Y.Z. Genome sequence of the model medicinal mushroom Ganoderma lucidum. Nat. Commun. 2011, 3, 913. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.Y.; Fan, W.L.; Wang, W.F.; Chen, T.; Tang, Y.C.; Chu, F.H.; Chang, T.T.; Wang, S.Y.; Li, M.Y.; Chen, Y.H. Genomic and transcriptomic analyses of the medicinal fungus Antrodia cinnamomea for its metabolite biosynthesis and sexual development. Proc. Natl. Acad. Sci. USA 2014, 111, 4743–4752. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.F.; Gong, Y.H.; Cai, Y.L.; Liu, W.; Zhou, Y.; Xiao, Y.; Xu, Z.Y.; Liu, Y.; Lei, X.Y.; Wang, G.Z. Genome sequence of the edible cultivated mushroom Lentinula edodes (shiitake) reveals insights into lignocellulose degradation. PLoS ONE 2016, 11, e0160336. [Google Scholar] [CrossRef] [PubMed]
- Yap, H.Y.Y.; Chooi, Y.H.; Firdausraih, M.; Fung, S.Y.; Ng, S.T.; Tan, C.S.; Tan, N.H. The genome of the tiger milk mushroom, Lignosus rhinocerotis, provides insights into the genetic basis of its medicinal properties. BMC Genom. 2014, 15, 635. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Min, W.H.; Bi, P.Y.; Zhou, H.B.; Huang, F.R. Stimulatory effects of coix lacryma-jobi oil on the mycelial growth and metabolites biosynthesis by the submerged culture of Ganoderma lucidum. Biochem. Eng. J. 2013, 76, 77–82. [Google Scholar] [CrossRef]
- Tang, Y.J.; Zhong, J.J. Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma lucidum, grown on lactose in a bioreactor. Biotechnol. Lett. 2002, 24, 1023–1026. [Google Scholar] [CrossRef]
- Chen, R.Z.; Zhao, X.; Shao, Z.; Wei, Z.; Wang, Y.Y.; Zhu, L.L.; Zhao, J.; Sun, M.X.; He, R.F.; He, G.C. Rice UDP-glucose pyrophosphorylase1 is essential for pollen callose deposition and its cosuppression results in a new type of thermosensitive genic male sterility. Plant Cell 2007, 19, 847–861. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.T.; Du, S.C.; Lo, H.H.; Hsiao, Y.M. The galu gene of Xanthomonas campestris pv. Campestris is involved in bacterial attachment, cell motility, polysaccharide synthesis, virulence, and tolerance to various stresses. Arch. Microbiol. 2014, 196, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.S.; Lin, T.Y.; Wang, W.B.; Liu, M.C.; Poren, H.; Shwujen, L. Characterization of udp-glucose dehydrogenase and UDP-glucose pyrophosphorylase mutants of proteus mirabilis: Defectiveness in polymyxin b resistance, swarming, and virulence. Antimicrob. Agents Chemother. 2010, 54, 2000–2009. [Google Scholar] [CrossRef] [PubMed]
- Diamantopoulou, P.; Papanikolaou, S.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Patterns of major metabolites biosynthesis by different mushroom fungi grown on glucose-based submerged cultures. Bioprocess Biosyst. Eng. 2014, 37, 1385–1400. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Lai, M.N.; Lin, C.C.; Ng, L.T. Comparative characterization of physicochemical properties and bioactivities of polysaccharides from selected medicinal mushrooms. Appl. Microbiol. Biotechnol. 2016, 100, 4385–4393. [Google Scholar] [CrossRef] [PubMed]
- Chien, R.C.; Yen, M.T.; Tseng, Y.H.; Mau, J.L. Chemical characteristics and anti-proliferation activities of ganoderma tsugae polysaccharides. Carbohydr. Polym. 2015, 128, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.S.; Chen, H.X.; Zhu, W.C.; Wang, Z.S. Effect of different drying methods on physicochemical properties and antioxidant activities of polysaccharides extracted from mushroom inonotus obliquus. Food Res. Int. 2013, 50, 633–640. [Google Scholar] [CrossRef]
- Xu, X.Q; Li, J.; Hu, Y. Polysaccharides from inonotus obliquus sclerotia and cultured mycelia stimulate cytokine production of human peripheral blood mononuclear cells in vitro and their chemical characterization. Int. Immunopharmacol. 2014, 21, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.J.; Zhu, L.W.; Li, H.M.; Li, D.S. Submerged culture of mushrooms in bioreactors- challenges, current state-of-the-art, and future prospects. Food Technol. Biotechnol. 2006, 45, 221–229. [Google Scholar]
- Zhao, W.; Chai, D.D.; Li, H.M.; Chen, T.; Tang, Y.J. Significance of metal ion supplementation in the fermentation medium on the structure and anti-tumor activity of tuber polysaccharides produced by submerged culture of Tuber melanosporum. Process Biochem. 2014, 49, 2030–2038. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, L.N.; Chen, L.; Chen, Y.; Cheung, P.C.; Chen, L.G. Effect of culture media on the chemical and physical characteristics of polysaccharides isolated from Poria cocos mycelia. Carbohydr. Res. 2003, 338, 1507–1515. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Olsen, L.M.; Ruthes, A.C.; Czelusniak, P.A.; Santana-Filho, A.P.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Exopolysaccharides, proteins and lipids in Pleurotus pulmonarius submerged culture using different carbon sources. Carbohydr. Polym. 2012, 87, 368–376. [Google Scholar] [CrossRef]
- Chen, X.; Wu, J.Y.; Gui, X.T. Production and characterization of exopolysaccharides in mycelial culture of Cordyceps sinensis fungus CS-HK1 with different carbon sources. Chin. J. Chem. Eng. 2016, 24, 158–162. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Source | Monosaccharide Composition | Molar Ratio | Reference |
|---|---|---|---|---|
| Cordyceps sinensis | mycelium | Man:Glc:Gal:GlcA | 9.6:4.0:4.4:1.0 | [19] |
| Cordyceps militaris | fruiting body | Man:Gal:Glc | 56.7:34.5:8.8 | [22] |
| NA | Man:Glc:Gal | 2.84:1:1.29 | [23] | |
| Man:Glc:Gal | 2.05:1:1.09 | |||
| fruiting body | Man:Glc:Gal | 1:28.63:1.41 | [24] | |
| Man:Glc:Gal | 1:12.41:0.74 | |||
| Cordyceps sobolifera | fruiting body | Man:Glc:Gal | 1.7:8.9:1 | [25] |
| Cordyceps cicadae | NA | Glc:Rha:Xyl:Man:Ara:Gal | 63.10:39.11:20.12:15.16:2.05:0.12 | [26] |
| Cordyceps kyushuensis | stroma | Fru:Man:Rha:GalN:Ara | 1.0:1.19:0.11:0.11:0.34 | [27] |
| Fru:Man:Rha:Glc:Ara | 1:1.29:0.14:0.07:0.32 | |||
| Morchella esculenta | fruiting body | Xyl:Glc:Man:Rha:Gal | 5.4:5.0:6.5:7.8:72.3 | [28] |
| fruiting body | Glc:Man:Gal:Ara:Xyl:Rha | 38.06:14.43:17.06:9.25:2.08:0.94 | [29] | |
| Glc:Man:Gal:Ara:Xyl:Rha | 27.04:28.66:11.12:9.07:6.71:3.22 | |||
| Glc:Man:Gal:Ara:Xyl:Rha | 24.69:20.46:10.22:7.91:4.05:2.83 | |||
| fermentation broth | Ara:Man:Glc:Gal | 0.7:2.8:24.8:1.0 | [30] | |
| Rha:Man:Glc:Gal | 1.8:3.1:21.4:1.0 | |||
| Tuber huidongense | fruiting body | Glc:Man:Gal | 60.56:20.12:19.32 | [32] |
| Glc | - | |||
| Tuber rufum | fruiting body | Glc:Gal:Fuc | 4:3:1 | [33] |
| Species | Source | Monosaccharide Composition | Molar Ratio | Reference |
|---|---|---|---|---|
| Ganoderma lucidum | fruiting body | Rha:Ara:Xyl:Man:Glc:Gal | 11.4:30:27.1:8:9.1:14.3 | [34] |
| Rha:Ara:Xyl:Glc:Gal | 44.7:20.9:19.9:3.6:10.8 | |||
| NA | Man:Rha:Glc:Gal | 4.7:0.65:65.22:29.43 | [35] | |
| fruiting body | Glc | - | [36] | |
| Glc:Gal:Man | 29:1.8:1.0 | |||
| fruiting body | Glc | - | [37] | |
| fermentation broth | Gal:Man:Glc:Ara:Rha | 103:17:12:10:3 | [38] | |
| fruiting body | Glc:Fuc:Gal | 1.0:1.09:4.09 | [39] | |
| fruiting body | Gal:Rha:Glc | 1.00:1.15:3.22 | [40] | |
| fermented soybean curd residue | Ara:Rha:Xyl:Man:Glc | 4.66:1.23:3.1:0.61:1.29 | [41] | |
| Ara:Xyl:Glc | 2.82:1.33:0.87 | |||
| Ara:Rha:Xyl:Gal:Man:Glc | 5.09:0.52:1.07:1.29:0.48:2.76 | |||
| Ganoderma atrum | fruiting body | Gal:Fuc:Glc | 75.87:11.83:6.02 | [42] |
| fruiting body | Glc:Man:Gal:GalA | 4.91:1:1.28:0.71 | [43] | |
| Ganoderma sinense | fruiting body | Man:Glc:Gal | 4.7:27.1:1.0 | [44] |
| Ganoderma tsugae | mycelium | Fuc:Xyl:Man:Gal:Glc:GlcNac | 2.9:16.1:66.7:9.4:0.1:4.3 | [45] |
| Fuc:Xyl:Man:Gal:Glc:GlcNac | 4.5:1.3:58.0:24.5:3.8:7.9 | |||
| Ganoderma formosanum | fruiting body | Man:Gal:Glc:Ara:Fuc:Fru:Rha | 50.13:13.1:17.47:6.94:2.71:9.21:0.45 | [46] |
| Man:Gal:Glc:Ara:Fuc:Rha | 44.91:38.64:8.26:0.08:8.02:0.09 | |||
| Man:Gal:Glc:Ara:Fuc:Fru:Rha | 33.35:30.84:20.52:8.78:4.44:1.33:0.74 |
| Species | Biological Activity | Active Component | Reference |
|---|---|---|---|
| Inonotus obliquus | antioxidant and anticancer activity | Man | [53,54,55] |
| Tremella mesenterica | cytokine-stimulating activity | Man | [47] |
| Ganoderma lucidum | antioxidant activity | Rha | [34] |
| Flammulina velutipes | antioxidant activity | Rha | [57] |
| Agrocybe cylindracea | antioxidant activity | Glc, Gal | [58] |
| Sarcodon aspratus | macrophage activation | Man, Rha | [59] |
| Inonotus obliquus | antioxidant activity | Man, Rha, Xyl | [8] |
| Hirsutella sp. | antioxidant activity | Man, Glc | [60] |
| Pleurotus eryngii | antioxidant activity | Man, Rha, GalA | [61] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Wang, F.; Xu, Z.; Ding, Z. Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules 2017, 22, 955. https://doi.org/10.3390/molecules22060955
Wang Q, Wang F, Xu Z, Ding Z. Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules. 2017; 22(6):955. https://doi.org/10.3390/molecules22060955
Chicago/Turabian StyleWang, Qiong, Feng Wang, Zhenghong Xu, and Zhongyang Ding. 2017. "Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation" Molecules 22, no. 6: 955. https://doi.org/10.3390/molecules22060955
APA StyleWang, Q., Wang, F., Xu, Z., & Ding, Z. (2017). Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules, 22(6), 955. https://doi.org/10.3390/molecules22060955